1. Introduction
The plant species
Zanthoxylum armatum DC. (ZA; also known as Nepalese or Timur pepper) is from the
Rutaceae or citrus, family and its bark, seeds and fruits have a long history as sources of medicinal treatments. This is particularly prevalent in Chinese, Nepalese and Ayurvedic medical systems with indications including as a treatment for depression, gastro-intestinal and digestive disorders and as a topical treatments for toothache and skin irritation/wounds [
1,
2,
3]. It is also one of several species of the
Zanthoxylum genus generically known as Szechuan pepper; a traditional Chinese and Nepalese culinary spice comprising the fruit pericarp which is typically added to wok and dumpling dishes to provide a characteristic grapefruit-like taste and flavor and a tingling numbness in the mouth.
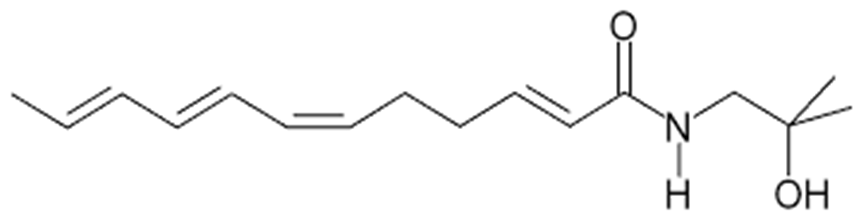
Whilst the various tissues from ZA have complex phytochemistry, the tingling mouth sensation, and any gastro-intestinal or dental applications, are related to the presence of a number of alkamides and unsaturated fatty acid amides; most notably hydroxy α-sanshool (see
Figure 1) and related structures [
4]. The alkamide group of compounds, which are also found in 300+ species from the
Zanthoxylum,
Acmella (Spilanthes),
Anacyclus,
Echinacea and
Heliopsis genera, typically also have insecticidal, larvicidal and antimicrobial properties and act as plant growth factors [
4]. In mammals, they have anti-inflammatory, immunomodulatory and analgesic properties and may underlie the putative medicinal effects of plants such as Echinacea and ‘maca’ (
Lepidium meyeni) [
1,
4]. Evidence suggests that alkamides can readily cross the skin, oral and gut mucosa and that they can traverse the blood brain barrier [
5]. In rodent models, the alkamide riparin II has been shown to reduce anxiety [
6] and attenuate depression via monoaminergic mechanisms [
7].
In a medicinal context, ZA is most likely to be encountered as a component of the traditional Japanese herbal medicine Daikenchuto, which comprises ginger rhizome, ginseng root, rice gluten and
Zanthoxylum fruit. Daikenchuto is generally indicated for gastrointestinal disorders [
8], with some evidence of efficacy following abdominal surgery [
9,
10]. In a study assessing the pharmacokinetics of Daikenchuto, it was demonstrated that hydroxy α-sanshool achieved by far the highest plasma levels of the six compounds measured. Its bioavailability peaked after 30 min with a mean half-life of approximately 2 h [
11]. A single study [
12] demonstrated that Daikenchuto, administered for 3 days, improved mouse learning and memory and attenuated the cognitive decrements associated with the muscarinic receptor antagonist scopolamine. The learning and memory effects were also evident following both the ZA and hydroxy α-sanshool components by themselves, confirming that hydroxy α-sanshool was the active component in terms of learning and memory benefits.
In mechanistic terms, the gastrointestinal effects of Daikenchuto have been attributed to increased acetylcholine release [
13]. In line with this, an attenuation of the learning and memory decrements associated with scopolamine in rodents suggests cholinergic activity as a potential mechanism of any brain function effects of Daikenchuto and its active ingredients [
12]. However, it is also notable that alkamides such as γ-sanshool have been shown to interact with cannabinoid CB
1 and CB
2 receptors; due to the structural similarity between the sanshool group and endocannabinoids [
14]. In general, the oral sensations associated with a variety of spices and food additives are attributable to interactions with differing transient receptor potential (TRP) channels. These plasma membrane ion channels modulate the entry of positively charged ions into numerous cell types and mediate the perception of sensations such as pressure, temperature and pain [
15]. The tingling numbness associated with hydroxy α-sanshool is specifically due to interactions with TRPV1 (also known as the vanilloid receptor) and TRPA1 receptors in sensory neurons, which result in an influx of Ca
2+ [
16]. Hydroxy α-sanshool also inhibits members of the two-pore domain potassium ion channel family (designated as the KCNK family by the Human Genome Organization) that are sensitive to mechanical stimuli, pH, signaling lipids and other G-protein receptor ligands including neurotransmitters and anesthetics. In this regard hydroxy α-sanshool specifically inhibits family members TASK-1 (TWIK-related acid-sensitive potassium-1), TASK-3 and TRESK [
4]. Interestingly, evidence shows that both the TRP and KCNK ion channels that are modulated by hydroxy α-sanshool are widely distributed in cognition relevant brain regions [
17,
18].
Alongside significant levels of monoterpenes, which have been shown to exert independent beneficial effects on cognitive function [
19,
20], it is possible that an extract rich in hydroxy α-sanshool will have multifarious effects on human brain function. The current double-blind, randomized, parallel groups study therefore investigated the effects of a single 80 mg dose and 56-days supplementation with a lipid extract of ZA on cognitive function, mood and cerebral blood-flow parameters in the frontal cortex.
2. Methods
2.1. Design
The study adopted a double-blind, placebo-controlled, parallel-groups design, in which the acute and chronic effects of Zanthoxylum armatum DC. (Nepalese pepper) extract (ZA) and placebo were assessed pre-dose and at 1, 3 and 5 h post-dose on the first day (i.e., acute effects) and following 56 days (+/−3 days) consumption of the intervention. An optional, concomitant investigation of the cerebral blood-flow effects of the extract was carried out in ~50% of the total sample.
2.2. Participants
The final sample comprised a total of 82 participants, aged 30 to 55 years, who self-reported themselves to be in good health. Participants were excluded from the study if they: Had any pre-existing medical condition/illness or were currently taking prescription medications which might have an impact on their ability to take part in the study; were taking any dietary supplements including omega fatty acids/fish oils; had high blood pressure (systolic over 159 mm Hg or diastolic over 99 mm Hg); had a Body Mass Index (BMI) outside of the range 18–35 kg/m2; were pregnant, seeking to become pregnant or lactating; had learning difficulties or dyslexia; had an uncorrected visual impairment; were smokers or regular consumers of nicotine containing products; had a history of alcohol or drug abuse; consumed caffeine in excess of 500 mg per day; had any food intolerances/sensitivities; were unable to complete all of the study assessments or were non-compliant with regards treatment consumption (<80%; although this did not apply to any participants in this study).
The group demographics are shown in
Table 1. below. Two participants dropped out after their Day 1 assessment (for reasons unrelated to treatment) and were only included in the analysis of Day 1 data.
A small number of data points were either missing or omitted due to incorrect performance of tasks. Individual sample sizes for each task (n) are shown in the data tables in the Online
Supplementary Materials.
Near-infrared Spectroscopy (NIRS) Cerebral Blood-Flow Participants
A sub-set of 41 participants (21 placebo, 20 ZA) took part in the cerebral blood-flow (CBF) assessment. Two participants in the placebo group were excluded from the assessment of the global effects of the treatments on resting CBF on Day 1/Day 56 and the acute CBF effects on Day 1 due to aberrant pre-dose baseline data due to a technical issue. The data from both participants was included in the assessment of hemodynamic responses on Day 1 and Day 56 (which used pre-task resting data as the baseline).
The study received ethical approval from the Northumbria University Psychology department (within the faculty of Health and Life Sciences) staff ethics committee (Reference 7905) and was conducted according to the Declaration of Helsinki (1964). All participants gave their written informed consent prior to their inclusion in the study.
2.3. Treatments
Treatments comprised 4 dark-brown soft gel capsules/day containing either:
The medium-chain triglyceride (MCT) oil extract of Zanthozylum armatum DC. (SaraPEPP™ Nu) was provided by Mibelle Group Biochemistry, Mibelle AG, Switzerland. Each 2.8 g dose contained alkamides including 7.8 mg hydroxy α-sanshool and monoterpenes including 5.3 mg limonene, 3.6 mg linalool and 2.3 mg methyl cinnamate.
Participants consumed the first and last dose of their 56-day treatment regimen under supervision within the laboratory. Otherwise, they consumed their treatment twice daily (2 capsules in the morning, 2 capsules in the evening, together with a meal) at home. The average compliance (as assessed by treatment diary and returned pill count) was 98%.
A treatment guess at the end of the study showed that there was no significant difference between the accuracy of guessing in the two treatment groups.
2.4. Cognitive and Mood Measures
All cognitive tasks and mood measures were delivered via the Computerized Mental Performance Assessment System (COMPASS; see:
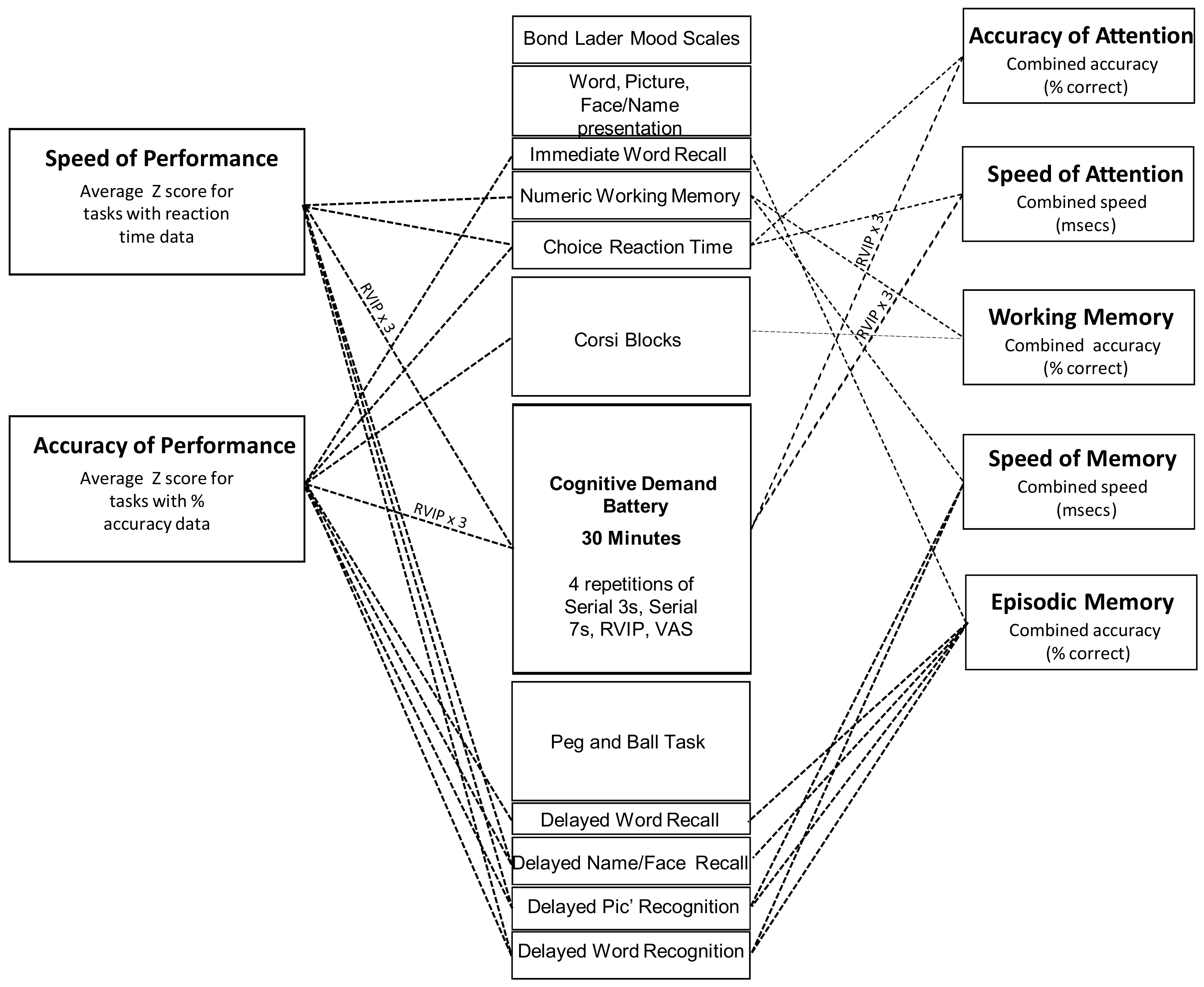
www.cognitivetesting.co.uk), a software platform for the presentation of classic and bespoke computerized cognitive tasks, with fully randomized parallel versions of each task delivered at each assessment for each individual. Tasks were presented on a laptop PC with responses made either via a four-button response box, with mouse and cursor or by the keyboard’s linear number pad. The tasks and other components of each assessment are described below in order of completion. The timelines of each assessment are shown in
Figure 2. Given the exploratory nature of the study, a wide spectrum of classic, widely employed cognitive tasks, assessing attention, secondary and working memory and executive function, were employed. A similar selection of tasks has previously been shown to be sensitive to nutritional interventions [
20,
21,
22,
23,
24].
2.4.1. Bond-Lader Mood Scales
The Bond-Lader Mood Scales [
25] have been utilised in numerous pharmacological, psychopharmacological and medical trials. These scales comprise a total of sixteen 100 mm lines anchored at either end by antonyms (e.g., ‘alert-drowsy,’ ‘calm-excited’). Participants indicate their current subjective position between the antonyms on the line. Outcomes comprise three factor analysis derived scores—‘Alertness,’ ‘Calmness’ and ‘Contentment.’
2.4.2. Stimuli (Picture, Name/Face, Word) Presentation
Fifteen black-and-white photographic images of objects were presented sequentially on screen for the participant to remember at the rate of 1 every 3 s, with a stimulus duration of one second. Following this a set of twelve passport-style photographic images of people were presented sequentially to participants. A first and last name was assigned to each photograph and presented on the screen underneath the person’s face. Stimulus duration was one second, with a 3-s inter-stimulus duration. Finally, a unique set of fifteen words was presented. Words were selected at random from a large bank of words derived from the Medical Research Council Psycholinguistic Database [
26] and matched for word length, frequency, familiarity and concreteness. Stimulus duration was one second, as was the inter-stimulus duration.
2.4.3. Immediate Word Recall
The participant was allowed 60 s to write down as many of the words as possible. The task was scored for number correct and errors.
2.4.4. Numeric Working Memory
Five random digits from 1–9 were presented sequentially for the participant to hold in memory. This was followed by a series of 30 probe digits (15 targets and 15 distractors) for each of which the participant indicated whether or not it had been in the original series by a simple key press. The task consisted of 3 separate trials. Accuracy (% correct) and mean reaction time (ms) were recorded.
2.4.5. Corsi Blocks Task
In this task, nine identical blue squares appeared on screen in non-overlapping random positions. A set number of blocks changed color from blue to red in a randomly generated sequence. The cursor was locked in position until the entire sequence had been presented, at which point the participants were instructed to repeat the sequence by clicking on the blocks using the mouse and cursor. The task was repeated five times at each level of difficulty. The sequence span increased from 4 upwards, until the participant could no longer correctly recall the sequence, resulting in a span measure of nonverbal working memory, calculated by averaging the level of the last five correctly completed trials.
2.4.6. Choice Reaction Time (CRT)
The Choice Reaction Task (CRT) task required participants to indicate, by pressing the ‘left’ or ‘right’ response box button, the direction of the arrow presented on the computer screen. Fifty stimuli (arrows) were presented, with a randomly varying delay of 1 to 3 s between stimuli. The task took approximately 2 min to complete, depending on participant reaction time. The task wass scored for % correct responses and reaction time (ms).
2.4.7. Cognitive Demand Battery
Multiple completions of this 10 min battery of tasks reliably increases self-ratings of mental fatigue and it has been shown to be sensitive to many natural interventions [
19,
27,
28,
29,
30]. Two minutes each of Serial 3s and 7s subtractions is first completed and followed immediately by 5 min of Rapid Visual Information Processing. Mental fatigue is self-rated after each completion of the three tasks. In the current study the tasks were repeated 3 times, giving a total of 30 min of continuous demanding cognitive task performance.
Serial 3s and 7s subtractions—At the start of the 2 min task a standard instruction screen informs the participant to count backwards in 3s or 7s as quickly and accurately as possible, using the keyboards linear number keys to enter each response. Participants were instructed verbally at the outset that if they were to make a mistake they should carry on subtracting from the new incorrect number with subsequent responses scored as correct in relation to the new number. To begin, a random starting number between 800 and 999 was presented on the computer screen, which was cleared by the entry of the first response. Each three-digit response was represented on screen by asterisks and pressing the enter key signalled the end of each response and cleared the three asterisks from the screen. Outcomes were the total number of correct subtractions and the number of incorrect responses.
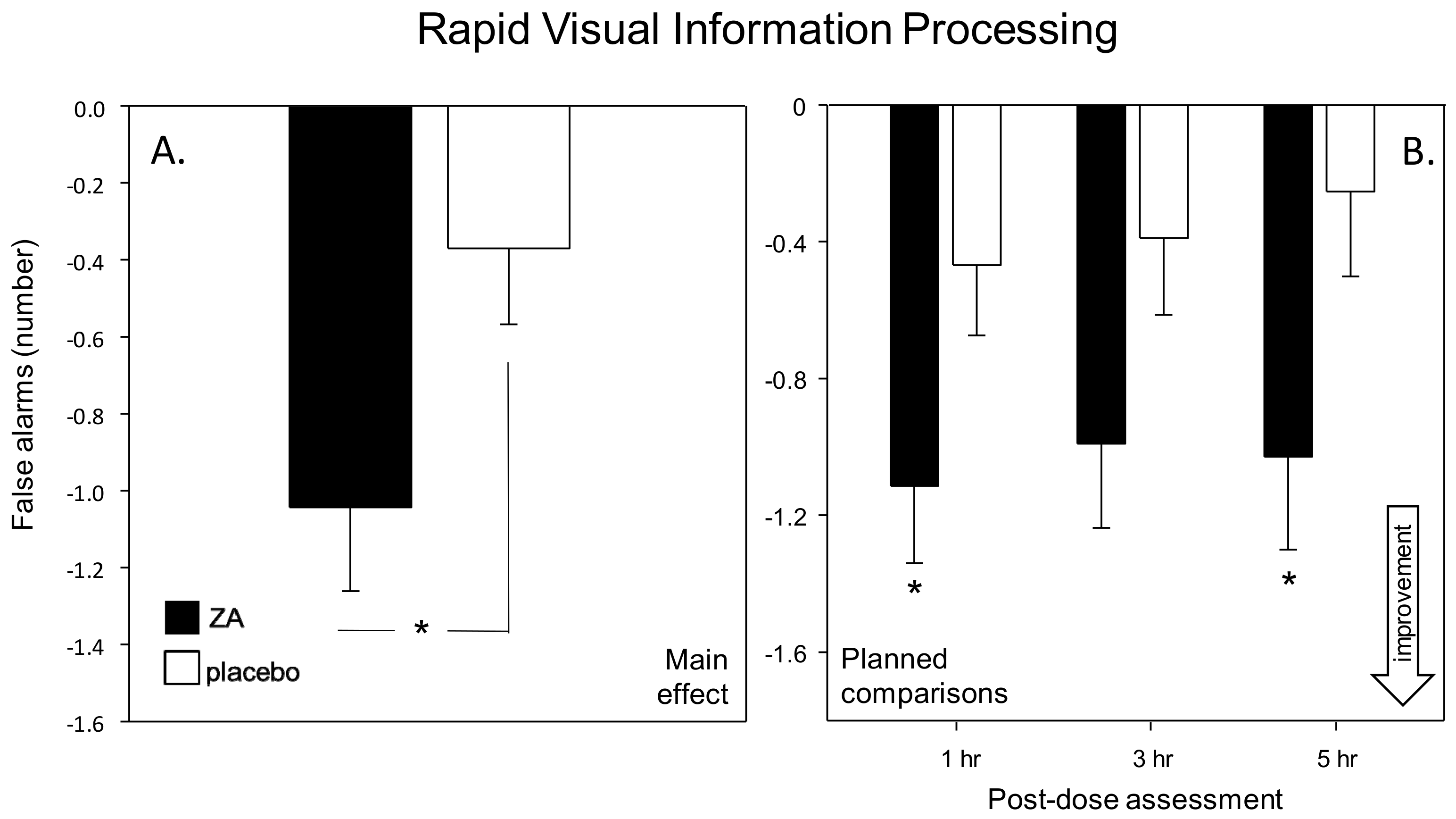
Rapid Visual Information Processing (RVIP) task—The RVIP task required the participant to monitor a continuous series of single digits for targets of three consecutive odd or even numbers. The white digits are presented on the black computer screen at the rate of 100 per minutes; with eight correct target strings in each minute presented in pseudo-random order. The participant responded to the detection of a target string by pressing the appropriate response button as quickly as possible. The RVIP was scored for the % target strings correctly detected, false alarms and the average reaction time (ms) for correct detections.
2.4.8. Peg and Ball
Two configurations of wooden peg (×3) and ball (×3; blue, green and red) diagrams were displayed, on screen, with the top diagram denoting the ‘goal’ configuration of balls on pegs. Participants were required to rearrange the balls on the ‘starting’ configuration below this to match the ‘goal’ configuration. They were instructed to do this in the least number of moves possible. The task was scored for average thinking time (ms), average completion time (ms) and errors (total number of moves in excess of minimum required to complete all trials).
2.4.9. Delayed Word Recall
Participants were informed, via a computerized instruction page, that they had 60 s to note down as many of the words from the list presented at the beginning of the task battery as they could remember. The task outcomes were the number of correctly recalled words and the number of incorrect words.
2.4.10. Delayed Name to Face Recall
The target faces presented at the beginning of the battery were displayed on the screen one at a time. Below each face was a list of 4 forenames and a list of 4 surnames. Participants used the mouse to select the forename and surname that they believed were associated with each face as presented at the beginning of the assessment. The task outcomes comprised % accuracy for overall correct forenames and correct surnames and reaction time (ms).
2.4.11. Delayed Picture and Word Recognition
Word and picture recognition were completed separately but both required participants to differentiate, by pressing ‘yes’ or ‘no’ on the response box, between the 15 target words and pictures presented at the beginning of the test battery and 15 randomly interspersed decoy words and pictures. The tasks took ~2 min to complete and were scored for % of correctly recognized words/pictures and reaction time (ms).
All of the computerized cognitive assessments were identical, with the exception of the presentation of randomly generated parallel versions of each task. The individual tasks making up the cognitive/mood assessment, alongside the timelines of each assessment are shown in
Figure 2.
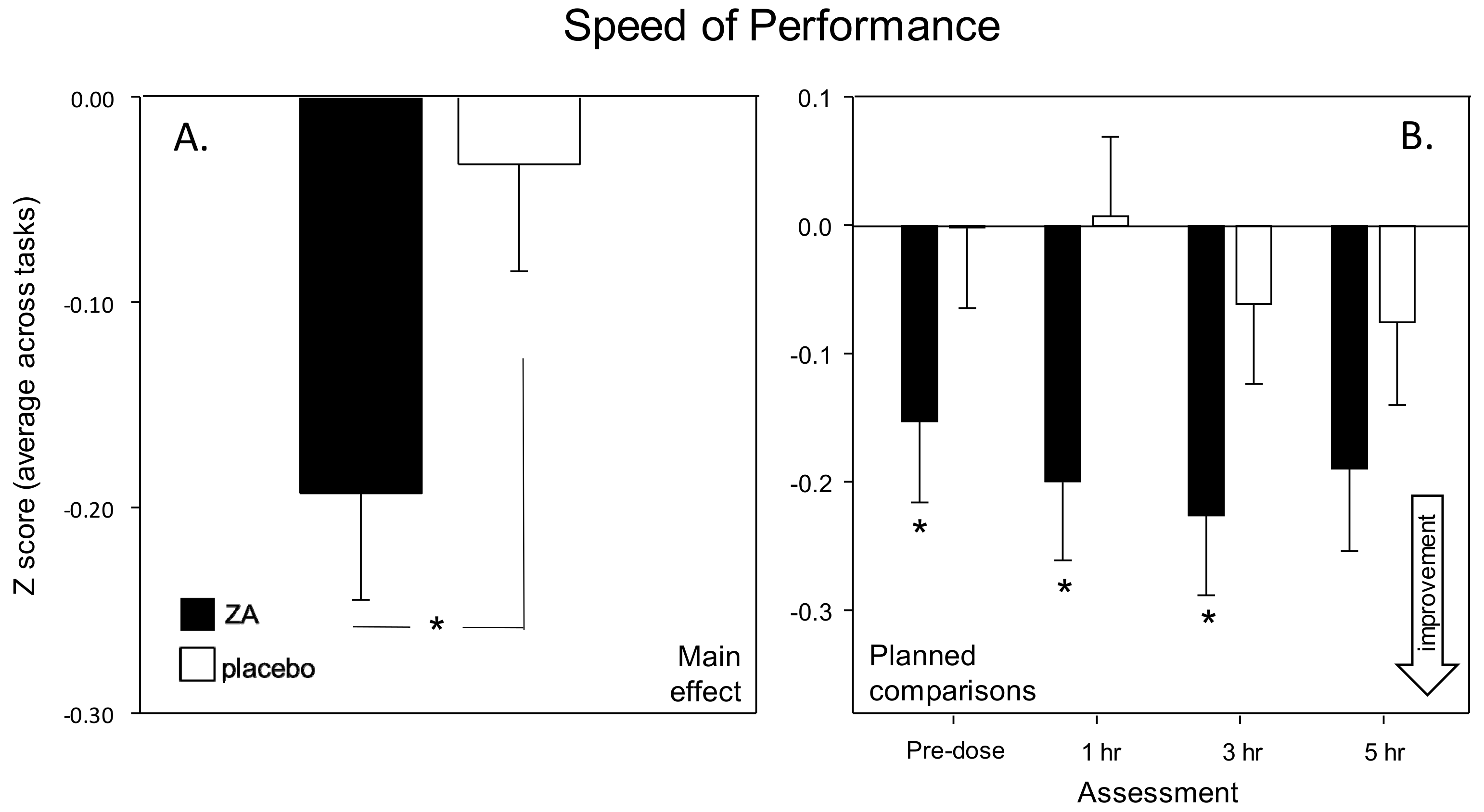
Figure 2 also shows the contribution of individual tasks to the measures that were derived by collapsing the data from individual tasks into cognitive domain factor scores representing the average speed (ms) of performance (Speed of Attention, Speed of Memory) or average (% correct) accuracy (Accuracy of Attention, Working Memory, Secondary Memory) of the attention tasks and working/episodic memory tasks within the battery. Two global performance measures were also calculated; ‘Speed of Performance’ and ‘Accuracy of Performance.’ These comprised averaged Z scores for speed and accuracy data respectively (for all tasks that collected such data). The composite measures have previously been shown to be sensitive to nutritional interventions ([
23,
31]). The task running order and contribution to cognitive factors and global measures are shown in
Figure 2 below.
2.5. Cerebral Blood-Flow (via NIRS) Assessment
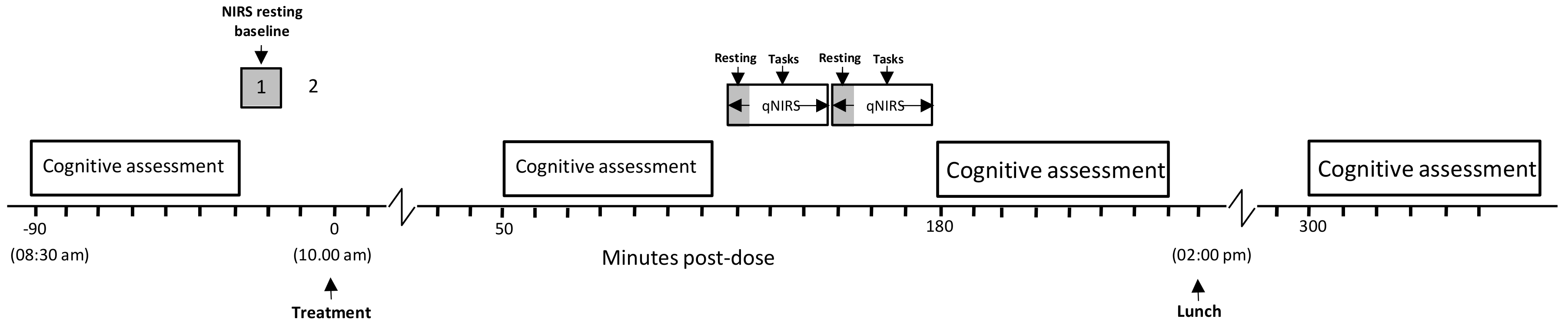
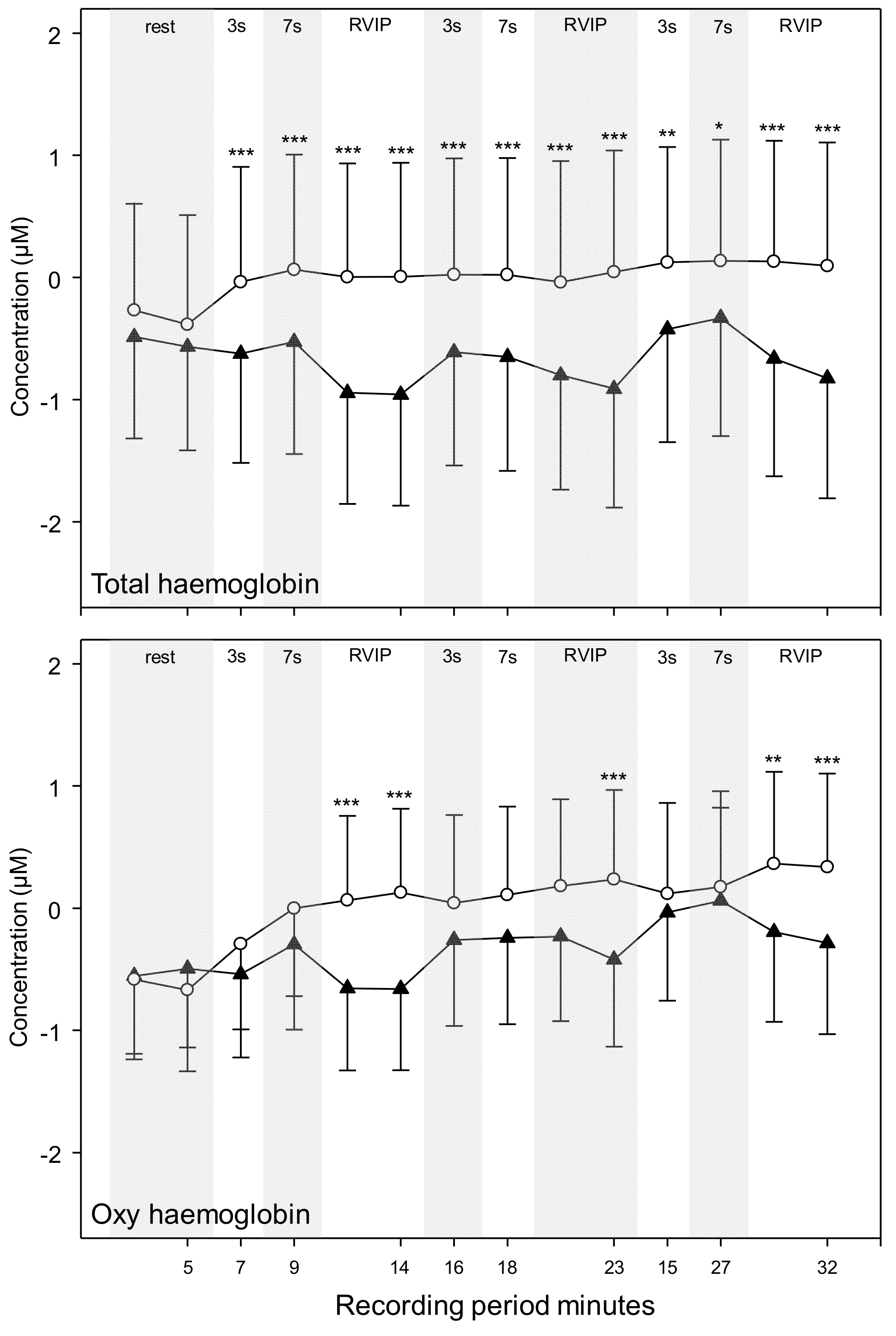
Forty one of the participants took part in an additional NIRS assessment, with CBF parameters (oxygenated hemoglobin [oxy-Hb], deoxygenated hemoglobin [deoxy-Hb], total-hemoglobin [total-Hb—the sum of oxy-Hb and deoxy-Hb, a marker of overall CBF] and % oxygen saturation [% oxy]) measured in the frontal cortex using NIRS at rest and during cognitive task performance. Each assessment comprised a 5 min pre-dose resting measurement and a post-dose assessment commencing either at 120 or 150 min post-dose. The post-dose assessment comprised a 5 min resting measurement followed by 27 min (3 repetitions) of performing the Cognitive Demand Battery tasks (Serial 3s – 2 min, Serial 7s – 2 min, RVIP—5 min) all of which activate the frontal cortex. The NIRS assessment is shown in
Figure 3.
Near-Infrared Spectroscopy (NIRS)
Cerebral blood-flow (CBF) parameters and hemodynamic responses during task performance were monitored using a quantitative NIRS system (OxiplexTS Frequency-Domain Near-Infrared Tissue Oximeter; ISS, Inc., Champaign, IL, USA). NIRS provides a proxy measure of CBF parameters and has been shown to sensitively measure resting CBF and hemodynamic responses in humans during differing levels of brain activation [
32]. The system employed here gives absolute measurements of absorption of near-infrared light emitted at two distinct wavelengths. This allows the quantification of oxy-Hb and deoxy-Hb. These values are then used to determine total-Hb (oxy-Hb + deoxy-Hb) and % oxy (oxy-Hb/total-Hb × 100). This system is ideal for quantifying acute changes in hemodynamic responses over an extended period (i.e., with intermittent testing throughout one visit) and in a chronic context (here comparing CBF parameters between Day 1 and Day 56).
Light was emitted at 691 and 830 nm by optical fibers glued in pairs to four prisms (eight fibers in total) that were separated from the collector bundle, also glued to a prism, by 2.0-, 2.5-, 3.0- or 3.5 cm. Each of the emitter and collector bundle prisms were embedded into a flexible polyurethane resin to form a sensor with the overall dimensions of 7.6 cm × 2.5 cm × 0.3 cm. Identical sensors were attached to either side of the forehead of participants with medical tape and secured in place with a self-adhering bandage. The sensors were positioned so that the bottom edge was level with the top of the participants’ eyebrows and the middle edge touching at the midline of the forehead. Data were collected at a rate of 5 Hz.
2.6. Procedure
Testing took place in a suite of testing facilities with participants visually isolated from each other. Participants attended the laboratory on 3 separate occasions; an introductory visit between 1 and 14 days before the first day of treatment and two testing days (Day 1 and Day 56 of treatment).
The Introductory visit to the laboratory comprised—briefing on requirements of the study, obtaining of informed consent, health screening, completion of the Caffeine Consumption Questionnaire (CCQ) and State-Trait Anxiety Inventory (STAI) trait subscale, training on the cognitive and mood measures and collection of demographic data.
For the two subsequent laboratory-based testing sessions (Day 1, Day 56) participants attended the laboratory at 8:30 am having consumed a standardized breakfast of cereal and toast at home no later than an hour before arrival. They had refrained from alcohol for 24 h and caffeine for 18 h before each visit. On arrival on each day, participants completed the STAI state subscale and the computerized cognitive assessment (as per
Figure 2). The participants who were taking part in the CBF study (maximum 2 participants per day) were then equipped with the NIRS for a 5-min resting measurement of CBF parameters. Thirty minutes after the initial cognitive assessment participants consumed their treatment for that day. Three further cognitive assessments, identical to the pre-dose assessment commenced at 1 h, 3 h and 5 h post-dose, with the CBF subset of participants undergoing their post-dose NIRS assessment between the 1 h and 3 h cognitive assessments (i.e., commencing at either 120 or 150 min post-dose). Participants were given a standardized lunch at approximately 2:00 pm. Testing was identical on Day 56 with the exception that—following the completion of Day 1 testing, participants were provided with the treatment they would be consuming between study days and a treatment diary; and following the completion of testing on Day 56, participants were asked to guess which treatment they had consumed and they returned their remaining treatment and diary for analysis of compliance and adverse events. The testing day timeline is shown in
Figure 4.
2.7. Analysis
2.7.1. Cognitive and Mood Outcomes
The data from the individual tasks was collapsed into five separate cognitive factor scores (accuracy measures = average % correct: speed measures = average reaction time in ms) and two global performance scores (average of accuracy or speed Z scores) as shown in
Figure 2. The outcomes from the individual tasks shown in
Figure 2 were also analyzed as secondary outcomes.
The 3 repetitions (per assessment) of the Cognitive Demand Battery also returned the outcomes described above for the mental fatigue visual analogue scale, RVIP task, Serial 3sand Serial 7s.
Acute (Day 1) effects—In order to explore the acute effects of ZA, data from the Day 1 post-dose assessments at 1 h, 3 h and 5 h were baseline adjusted with regards the pre-dose baseline data collected on Day 1 and entered into a two way (treatment [placebo, ZA] × assessment [1 h, 3 h, 5 h]) ANOVA. Analyses of the Cognitive Demand Battery outcomes were by three-way (treatment [placebo, ZA] × assessment [1, 3 h, 5 h] × repetition [1, 2, 3]) ANOVA.
Chronic (Day 56) effects—In order to explore the chronic effects of ZA after 56 days administration, data from the pre-dose and 1 h, 3 h and 5 h post-dose assessments on Day 56 were baseline adjusted with respect to the Day 1 pre-dose baseline data and analyzed via two-way (treatment [placebo, ZA] × assessment [pre-dose, 1 h, 3 h, 5 h]) ANOVA to compare performance between treatments. Analyses of the Cognitive Demand Battery outcomes were by three-way (treatment [placebo, ZA] × assessment [1 h, 3 h, 5 h] × repetition [1, 2, 3]) ANOVA.
For both the acute and chronic analyses, planned comparisons between data from each condition during each assessment were also conducted using t-tests, utilizing Mean Squares Error (MSE) from the ANOVA. These comparisons were Bonferroni adjusted. Only those planned comparisons relating to an outcome that generated a significant ANOVA result are reported.
Day 1 baseline differences between placebo and ZA were explored using t-tests for those measures that had a single repetition and with reference to the main treatment effect of two-way ANOVAs (repetition [
3] × treatment [placebo/ZA]) for those measures (Cognitive Demand Battery outcomes) that had three repetitions.
2.7.2. Frontal Cortex Cerebral Blood Flow analysis
There were three separate analyses of data for these CBF parameters:
Global CBF effects on Day 1 and Day 56—The effects of ZA on global CBF parameters in the absence of task related brain activation (i.e., resting CBF) were analysed with unadjusted averaged data from the pre and post-dose resting assessments on Day 1 and Day 56. The analysis was by three-way ANOVA (pre/post dose × Day 1/56 × treatment) of averaged data from each 5 min rest period. Bonferroni adjusted post-hoc comparisons were then carried out between ZA and placebo as appropriate. This analysis also provided a test of baseline differences between groups on Day 1 and at rest (post-dose) on Day 1/Day 56.
Acute (Day 1) effects during brain activation—Post-dose data were averaged across 14 epochs (rest: 2 × 2.5 min, Serial 3s subtractions: 3 × 2 min, Serial 7s subtractions: 3 × 2 min, RVIP: 6 × 2.5 min: See Figures 9 and 10). The acute effects of a single dose of ZA on CBF parameters on Day 1 were measured as the change in CBF parameters during each 2 or 2.5 min epoch of the 32-min post-dose assessment, baseline adjusted to the pre-dose resting baseline. The analysis of change from baseline data was by two-way ANOVA (treatment [ZA/placebo] × epoch [14 × 2/2.5 min epochs). Bonferroni adjusted planned comparisons were then carried out between ZA and placebo during each epoch of the Day 1 post-dose assessment. Only those planned comparisons associated with a significant treatment related effect (multivariate or between-subjects) are reported below.
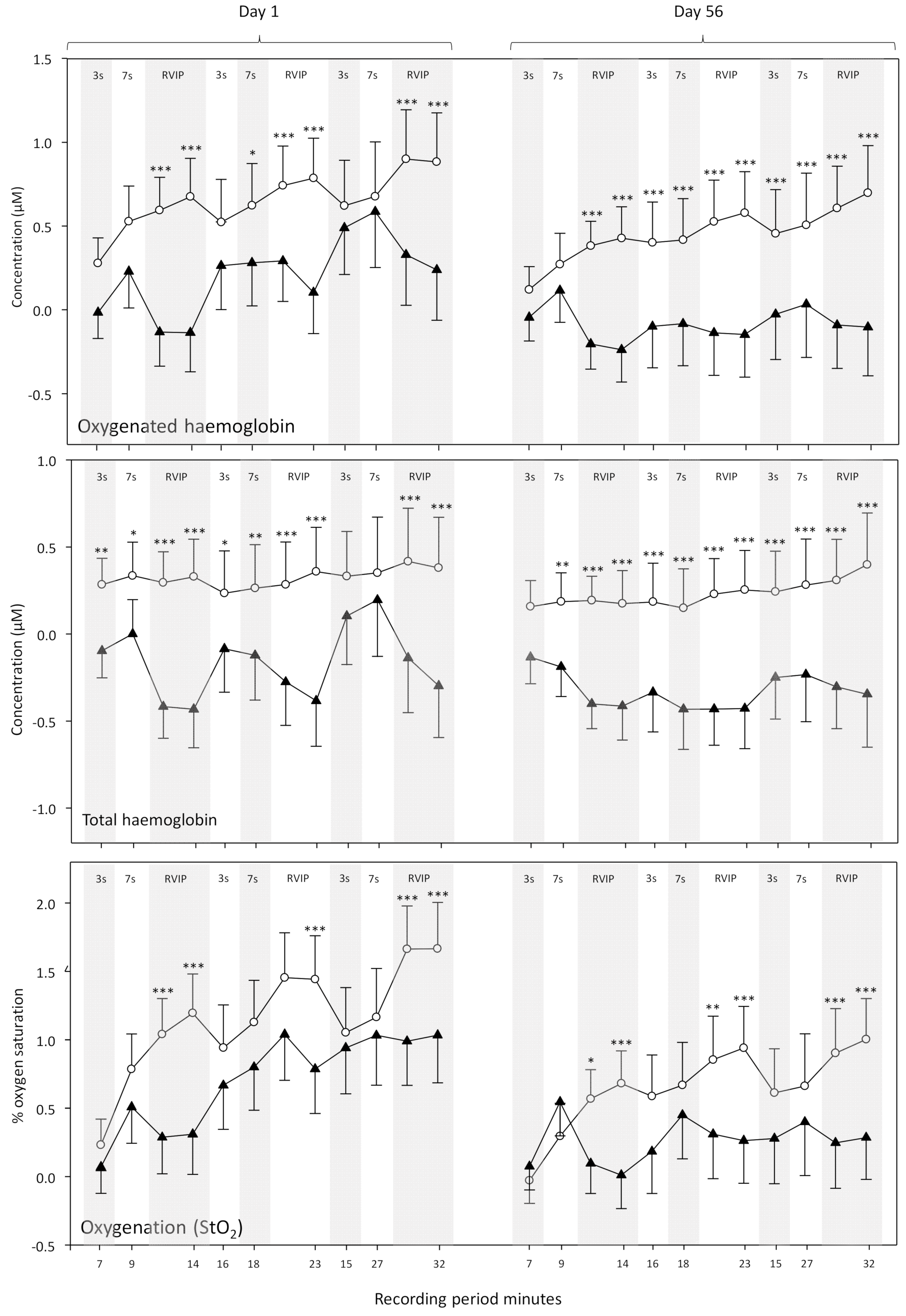
Hemodynamic responses on Day 1 and Day 56—In the absence of any effect on global blood-flow on Day 1/56 (see results below) the effects of ZA on hemodynamic responses (the CBF response to local brain activation; i.e., how much blood flow and oxygenation change in response to increased activity) were measured as changes in CBF parameters during each 2/2.5 min epoch of active task performance and baseline adjusted to the respective 5 min resting period immediately before task performance. The analysis of change from resting baseline data was performed by three-way ANOVA (treatment [ZA/placebo] × day [1/56] × epoch [12 × 2/2.5 min epochs]). Bonferroni adjusted planned comparisons were then carried out as described above.
4. Discussion
The results demonstrate that a single dose and 56-days administration of Zanthoxylum armatum DC. MCT oil extract (ZA; product name SaraPEPP™ Nu) lead to significant improvements in cognitive function and modulation of cerebral blood flow (CBF) parameters.
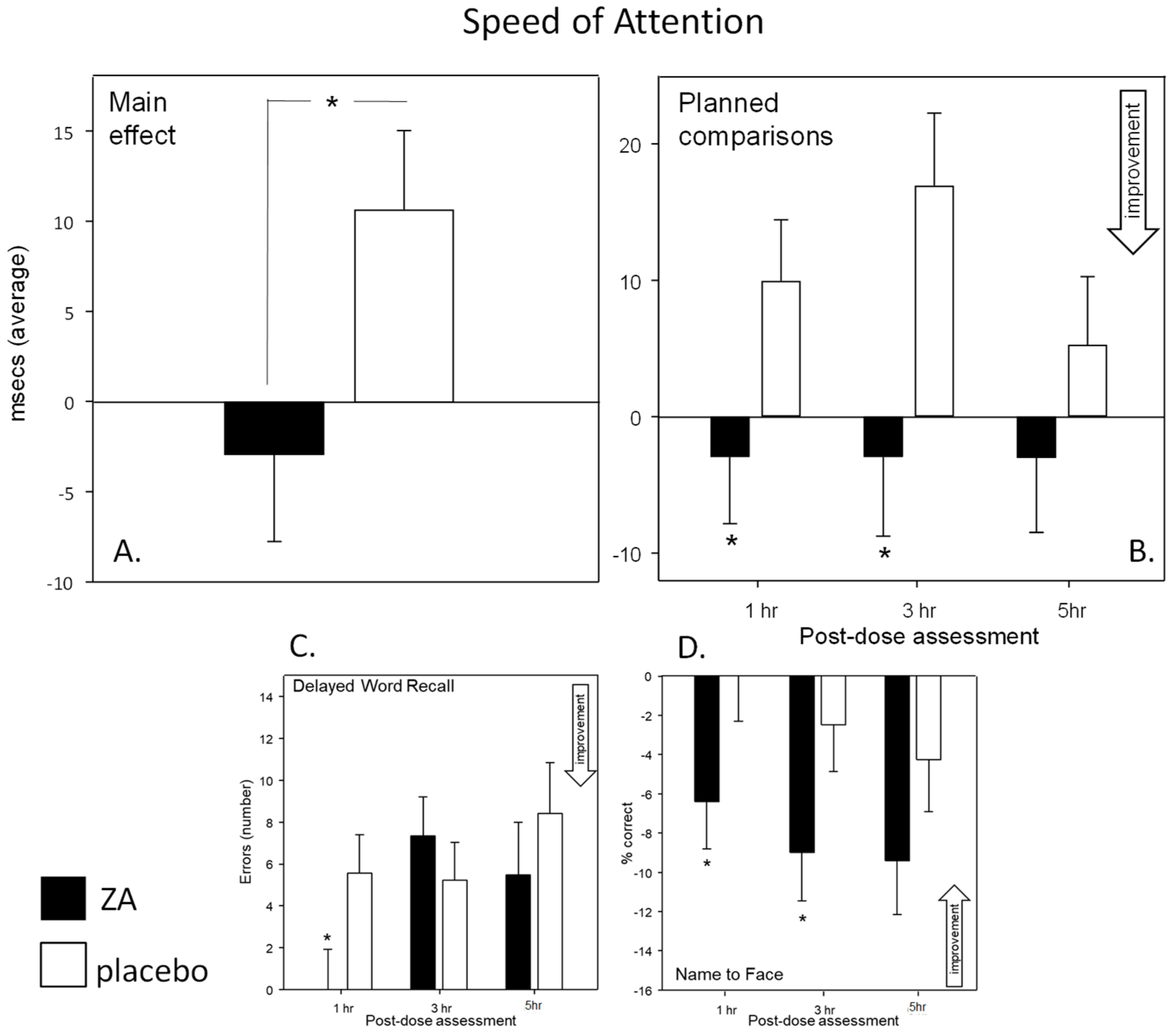
With regards cognitive function, the most striking finding was an increased speed of task performance. This was seen in terms of significantly faster attention task performance (Speed of Attention) on Day 1 and significantly faster performance across all tasks (Speed of Performance) on Day 56. These findings were also supported by corresponding trends towards faster task performance on both days (Speed of Performance on Day 1 and Speed of Attention on Day 56). Improvements were also seen within the Cognitive Demand Battery outcomes. Firstly, improved accuracy of Rapid Visual Information Processing (RVIP) performance, in terms of false alarms after a single dose (Day 1), was observed. Reduced ratings of Mental Fatigue across all assessments and increased correct Serial 3 s were also evident during the 3 h post-dose assessment, on Day 56. Of note, the beneficial effects of ZA on Speed of Performance and Mental Fatigue on Day 56 were evident both before and, to a greater extent, after the consumption of the day’s treatments, suggesting that the beneficial effects of ZA, at least in part, were related to chronic consumption.
The predominant cognitive effects were a general increase in speed of processing, rather than domain specific changes and these might be expected to correlate with a similar general change in CBF during task performance. With reference to the NIRS results, whilst ZA had no effect on gross, resting CBF following either a single dose or 56-days administration, it did result in modulation of several hemodynamic parameters during the performance of tasks that activate the pre-frontal cortex; both following a single dose and following 56-days administration. Broadly, the pattern of results was for reduced hemodynamic responses during task performance across both days. For the analysis combining data from Day 1 and Day 56 this was seen in terms of reduced oxygenated hemoglobin, reduced total hemoglobin (a proxy for overall CBF) and reduced % oxygenation, with these effects seen predominantly during the RVIP attention task. A further analysis of CBF data (task × treatment × day) confirmed that the significant effects seen here were task dependent and primarily seen during the RVIP task. It is notable that performance of the RVIP task has previously been shown to activate the prefrontal cortex [
33,
34]. Interestingly, the ANOVAs here did not show any interactions between visit and treatment; suggesting that the hemodynamic effects during task performance were similar after acute and chronic supplementation. However, it might be noteworthy that the pattern of modulation after 56-days administration could be described descriptively as being stronger, on the basis of the number of epochs that saw a significant difference between treatments.
One important point here is that the reduced hemodynamic response, particularly during the RVIP focused attention task, was not associated with poorer cognitive function. Indeed, as well as the significant improvements in the speed of performing the attention tasks on Day 1 and all of the timed tasks on Day 56, performance on the RVIP task for the full cohort was more accurate on Day 1 with trends towards increased accuracy and faster speed on this task on Day 56. Taken together, these findings strongly suggest that the pattern of hemodynamic effects seen here reflect improved physiological brain function. It would, of course, be interesting to explore the nature and spatial properties of the brain function effects seen here with alternative imaging techniques. NIRS only provides proxy measures of CBF and these can be conflated with changes in cerebral blood volume and cerebral rates of oxygen metabolism. In common with NIRS, most other brain imaging techniques also interpret proxy markers. However, investigations of ZA’s effects on regional electrophysiological activity (e.g., via Electroencephalography (EEG)), gross cerebral blood flow parameters (e.g., via Trans-Cranial Doppler) or the spatial resolution of any brain function effects (via Functional Magnetic Resonance Imaging (FMRI) would be of interest in future trials. Interestingly, recently developed neuroimaging techniques, such as Event-related optical signal (EROS) offer a direct measure of neuronal cellular activity and might be useful in disentangling any changes in local neuronal activity and blood flow.
Interestingly, the CBF results in the current study bear a striking resemblance to those seen following the consumption of dietary nitrate [
35]. In this previous study, which involved similar methodology as employed here, the reduction in CBF following a single dose of nitrate-rich beetroot juice was only seen during the RVIP task. There were no nitrate related effects on CBF during the serial subtraction tasks but improved performance on the Serial 3s subtraction task was seen nonetheless. In this earlier study it was noted that the RVIP task is a less cognitively demanding task and that the pattern of reductions in CBF represented an exaggeration of the typical response to reduced demand. The authors also attributed the findings to increased neural efficiency related to a nitrate related increase in the synthesis of the nitric oxide, with an attendant enhancement of cellular metabolic processes [
35].
In terms of mechanisms underpinning the effects seen in the current study, previous research suggests that both the gastrointestinal effects of Daikenchuto, which contains hydroxy-alpha-sanshool from
Z. piperitum [
13] and the learning and memory effects of ZA in rodents [
12] may be underpinned by increased cholinergic activity. Indeed, monoterpenes, which are represented in ZA extracts at significant levels, have been shown to have consistent cholinesterase inhibitory properties [
36,
37] and to exert beneficial cognitive effects [
20]; including at doses as low as 25 µL [
38]. A cholinergic mechanism certainly could be responsible for the cognitive effects seen here, although the pattern here on Day 56 was for a speeding of performance across cognitive domains; suggesting a general improvement in neural efficiency and processing speeds rather than the focused cognitive benefits to memory/attention that cholinergic agents are typically associated with. It is, however, unlikely that the interactions with cannabinoid CB1 and CB2 receptors attributed to some sanshools (specifically γ-sanshool) [
14] played any role here, as preliminary screening demonstrated that the extract administered here did not exhibit any cannabinoid receptor binding activity (data not presented but available on request).
One alternative possibility then, is that the cellular ion channel mechanisms that drive the sensory tingling-numbness following topical/oral application of hydroxy α-sanshool; namely interactions with TRP and KCNK ion channels, may also be responsible for the cognitive and CBF effects seen here. In this respect it is notable that structurally similar alkamides have been shown to have good bioavailability and can readily cross the blood brain barrier [
5]. Recent evidence also shows that the TRPV1 vanilloid receptors with which hydroxy α-sanshool interacts are widely distributed in the brain. Indeed, TRPV1 has a key role in the regulation of neuronal excitability and synaptic plasticity, including long-term potentiation, potentially mediated by glutamate release [
17]. Similarly, several members of the KCNK two-pore domain potassium ion channel family that are inhibited by hydroxy α-sanshool, specifically TASK-1, TASK-3 and TRESK [
4] are also expressed, often as TASK-1/TASK-3 heterodimers, throughout multiple brain regions, with particular concentration in the cerebellum, somatic motor-neurons and hypothalamus. These channels contribute to the excitability of distinct populations of neurons [
18]. As an example, TASK-1 and TASK-3 channels contribute to the excitability of several types of hippocampal neurons, including CA1 pyramidal cell and inhibitory interneurons [
39] and modulate the excitability of the key acetylcholine producing basal forebrain cholinergic neurons which drive elements of cortical arousal [
40]. Clearly, interactions with neuronal mechanisms such as these might be expected to have a general, rather than cognitive-domain specific effect in terms of neural activation/efficiency and hemodynamic responses to brain activation.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}