Low Salivary Amylase Gene (AMY1) Copy Number Is Associated with Obesity and Gut Prevotella Abundance in Mexican Children and Adults

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Case-Control Studies in Children and Adults
2.2. Anthropometric and Biochemical Parameters
2.3. CNV Quantification
2.4. Gut Microbiota Analyses
2.5. Dietary Assessment
2.6. Statistical Analyses
3. Results
3.1. Clinical Characteristics of Case-Control Study
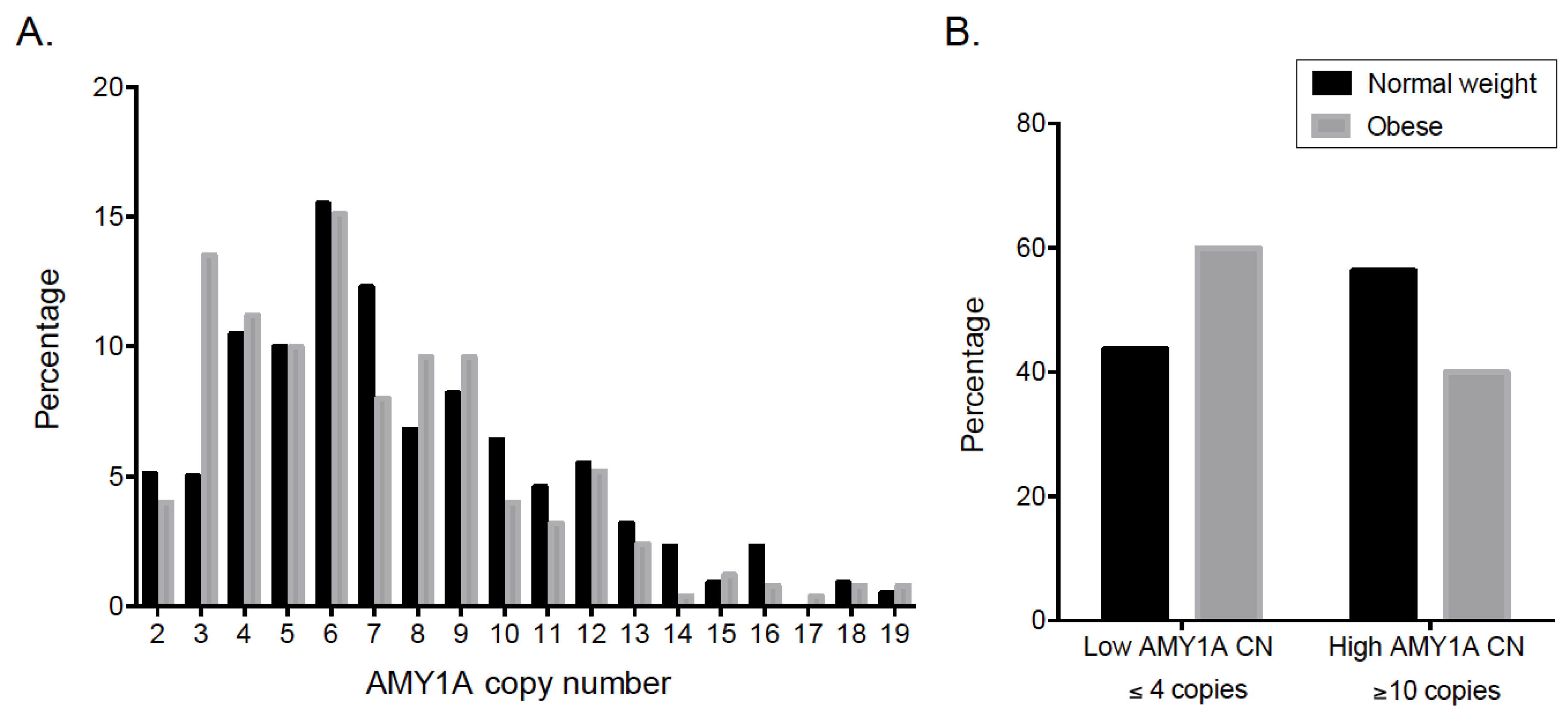
3.2. Association of 11q11and 1p21.1 (AMY1) CNVs with Obesity in Mexican Children
3.3. Association of AMY1 Copy Number with Obesity in Mexican Adults
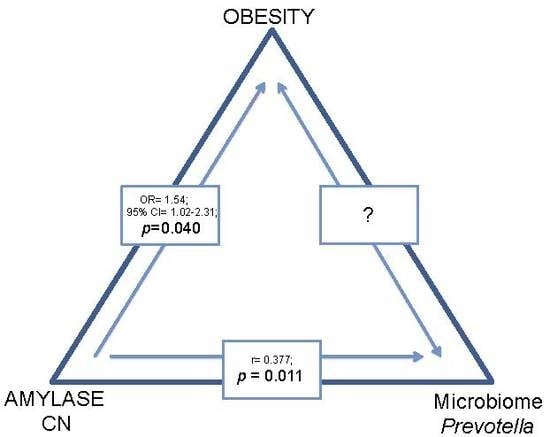
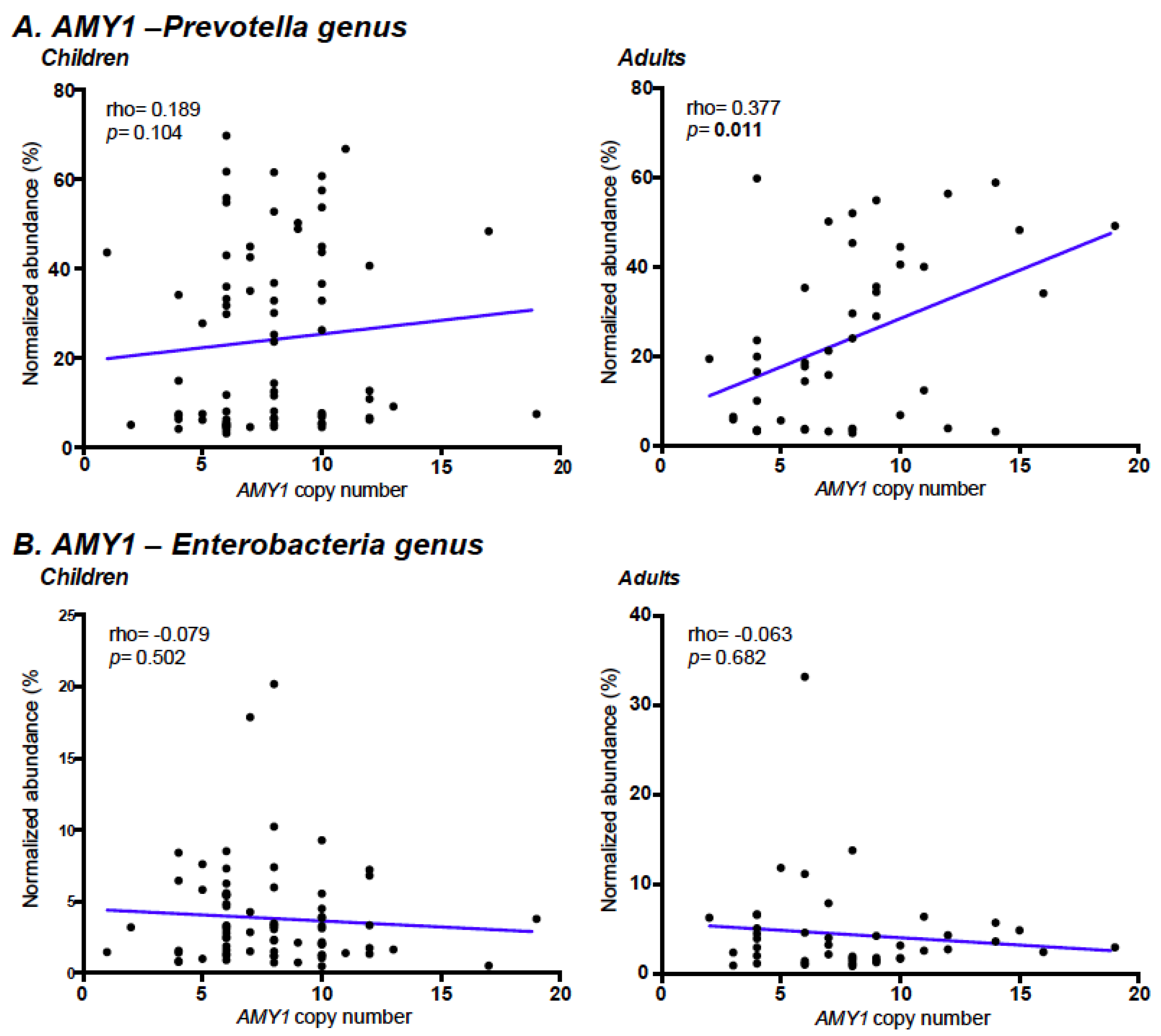
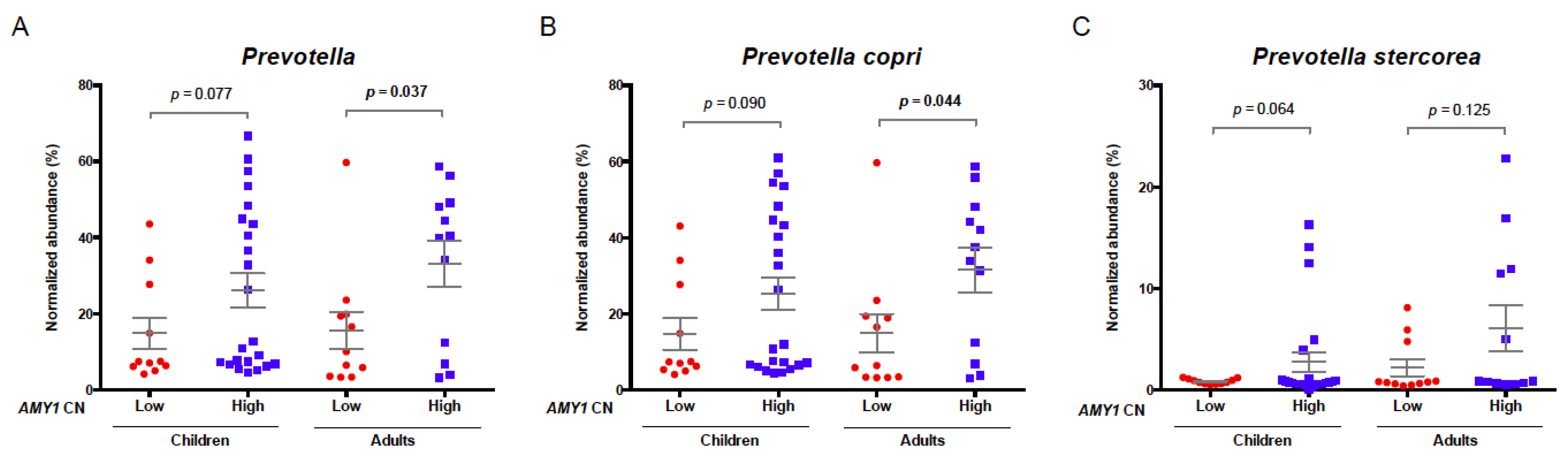
3.4. Association of AMY1 Copy Number with Prevotella Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arroyo-Johnson, C.; Mincey, K.D. Obesity Epidemiology Worldwide. Gastroenterol. Clin. N. Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Shamah-Levy, T.; Rivera-Dommarco, J.; Hernández-Ávila, M. Encuesta Nacional de Salud y Nutrición de Medio Camino; Instituto Nacional de Salud Pública: Cuernavaca, Mexico, 2016. [Google Scholar]
- Freedman, D.S.; Khan, L.K.; Serdula, M.K.; Dietz, W.H.; Srinivasan, S.R.; Berenson, G.S. The relation of childhood BMI to adult adiposity: The Bogalusa Heart Study. Pediatrics 2005, 115, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Juonala, M.; Magnussen, C.G.; Berenson, G.S.; Venn, A.; Burns, T.L.; Sabin, M.A.; Srinivasan, S.R.; Daniels, S.R.; Davis, P.H.; Chen, W.; et al. Childhood adiposity, adult adiposity, and cardiovascular risk factors. N. Engl. J. Med. 2011, 365, 1876–1885. [Google Scholar] [CrossRef] [PubMed]
- Elks, C.E.; den Hoed, M.; Zhao, J.H.; Sharp, S.J.; Wareham, N.J.; Loos, R.J.; Ong, K.K. Variability in the heritability of body mass index: A systematic review and meta-regression. Front. Endocrinol. 2012, 3, 29. [Google Scholar] [CrossRef] [PubMed]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.; Lindgren, C.M.; Li, S.; Wheeler, E.; Zhao, J.H.; Prokopenko, I.; Inouye, M.; Freathy, R.M.; Attwood, A.P.; Beckmann, J.S.; et al. Common variants near MC4R are associated with fat mass, weight and risk of obesity. Nat. Genet. 2008, 40, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Speliotes, E.K.; Loos, R.J.; Li, S.; Lindgren, C.M.; Heid, I.M.; Berndt, S.I.; Elliott, A.L.; Jackson, A.U.; Lamina, C.; et al. Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat. Genet. 2009, 41, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Speliotes, E.K.; Willer, C.J.; Berndt, S.I.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Lango Allen, H.; Lindgren, C.M.; Luan, J.; Magi, R.; et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Leon-Mimila, P.; Villamil-Ramirez, H.; Villalobos-Comparan, M.; Villarreal-Molina, T.; Romero-Hidalgo, S.; Lopez-Contreras, B.; Gutierrez-Vidal, R.; Vega-Badillo, J.; Jacobo-Albavera, L.; Posadas-Romeros, C.; et al. Contribution of common genetic variants to obesity and obesity-related traits in mexican children and adults. PLoS ONE 2013, 8, e70640. [Google Scholar] [CrossRef]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the missing heritability of complex diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Sha, B.Y.; Yang, T.L.; Zhao, L.J.; Chen, X.D.; Guo, Y.; Chen, Y.; Pan, F.; Zhang, Z.X.; Dong, S.S.; Xu, X.H.; et al. Genome-wide association study suggested copy number variation may be associated with body mass index in the Chinese population. J. Hum. Genet. 2009, 54, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Jarick, I.; Vogel, C.I.; Scherag, S.; Schafer, H.; Hebebrand, J.; Hinney, A.; Scherag, A. Novel common copy number variation for early onset extreme obesity on chromosome 11q11 identified by a genome-wide analysis. Hum. Mol. Genet. 2011, 20, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Falchi, M.; El-Sayed Moustafa, J.S.; Takousis, P.; Pesce, F.; Bonnefond, A.; Andersson-Assarsson, J.C.; Sudmant, P.H.; Dorajoo, R.; Al-Shafai, M.N.; Bottolo, L.; et al. Low copy number of the salivary amylase gene predisposes to obesity. Nat. Genet. 2014, 46, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Usher, C.L.; Handsaker, R.E.; Esko, T.; Tuke, M.A.; Weedon, M.N.; Hastie, A.R.; Cao, H.; Moon, J.E.; Kashin, S.; Fuchsberger, C.; et al. Structural forms of the human amylase locus and their relationships to SNPs, haplotypes and obesity. Nat. Genet. 2015, 47, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Yong, R.Y.; Mustaffa, S.B.; Wasan, P.S.; Sheng, L.; Marshall, C.R.; Scherer, S.W.; Teo, Y.Y.; Yap, E.P. Complex Copy Number Variation of AMY1 does not Associate with Obesity in two East Asian Cohorts. Hum. Mutat. 2016, 37, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.H.; Dominy, N.J.; Claw, K.G.; Lee, A.S.; Fiegler, H.; Redon, R.; Werner, J.; Villanea, F.A.; Mountain, J.L.; Misra, R.; et al. Diet and the evolution of human amylase gene copy number variation. Nat. Genet. 2007, 39, 1256–1260. [Google Scholar] [CrossRef] [PubMed]
- Mandel, A.L.; Peyrot des Gachons, C.; Plank, K.L.; Alarcon, S.; Breslin, P.A. Individual differences in AMY1 gene copy number, salivary alpha-amylase levels, and the perception of oral starch. PLoS ONE 2010, 5, e13352. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.M.; Lin, J.; Chen, L.H.; Zhang, M.; Chen, W.W.; Yang, X.R. The roles of AMY1 copies and protein expression in human salivary alpha-amylase activity. Physiol. Behav. 2015, 138, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Bjorck, I.; Backhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Parks, B.W.; Nam, E.; Org, E.; Kostem, E.; Norheim, F.; Hui, S.T.; Pan, C.; Civelek, M.; Rau, C.D.; Bennett, B.J.; et al. Genetic control of obesity and gut microbiota composition in response to high-fat, high-sucrose diet in mice. Cell Metab. 2013, 17, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Comparan, M.; Teresa Flores-Dorantes, M.; Teresa Villarreal-Molina, M.; Rodriguez-Cruz, M.; Garcia-Ulloa, A.C.; Robles, L.; Huertas-Vazquez, A.; Saucedo-Villarreal, N.; Lopez-Alarcon, M.; Sanchez-Munoz, F.; et al. The FTO gene is associated with adulthood obesity in the Mexican population. Obesity 2008, 16, 2296–2301. [Google Scholar] [CrossRef] [PubMed]
- Lohman, T.G.; Roche, A.F.; Martorell, R. Anthropometric Standardization Reference Manual Abridged Edition; Human Kinetics Books: Champaign, IL, USA, 1991; 90p, p. vi. [Google Scholar]
- Kuczmarski, R.J.; Ogden, C.L.; Guo, S.S.; Grummer-Strawn, L.M.; Flegal, K.M.; Mei, Z.; Wei, R.; Curtin, L.R.; Roche, A.F.; Johnson, C.L. 2000 CDC Growth Charts for the United States: Methods and development. Vital Health Stat. 11 2002, 246, 1–190. [Google Scholar]
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation; Technical Report Series; WHO: Geneva, Switzerland, 2000; pp. 1–253. [Google Scholar]
- Carpenter, D.; Dhar, S.; Mitchell, L.M.; Fu, B.; Tyson, J.; Shwan, N.A.; Yang, F.; Thomas, M.G.; Armour, J.A. Obesity, starch digestion and amylase: Association between copy number variants at human salivary (AMY1) and pancreatic (AMY2) amylase genes. Hum. Mol. Genet. 2015, 24, 3472–3480. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Contreras, B.E.; Moran-Ramos, S.; Villarruel-Vazquez, R.; Macias-Kauffer, L.; Villamil-Ramirez, H.; Leon-Mimila, P.; Vega-Badillo, J.; Sanchez-Munoz, F.; Llanos-Moreno, L.E.; Canizalez-Roman, A.; et al. Composition of gut microbiota in obese and normal-weight Mexican school-age children and its association with metabolic traits. Pediatr. Obes. 2018, 13, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.; Gonzalez, A.; Ackermann, G.; Wendel, D.; Vazquez-Baeza, Y.; Jansson, J.K.; Gordon, J.I.; Knight, R. Meta-analyses of studies of the human microbiota. Genome Res. 2013, 23, 1704–1714. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Avila, M.; Romieu, I.; Parra, S.; Hernandez-Avila, J.; Madrigal, H.; Willett, W. Validity and reproducibility of a food frequency questionnaire to assess dietary intake of women living in Mexico City. Salud Publica Mex. 1998, 40, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef]
- Stryjecki, C.; Alyass, A.; Meyre, D. Ethnic and population differences in the genetic predisposition to human obesity. Obes. Rev. 2018, 19, 62–80. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Hu, F.B. Gene-environment interactions and obesity: Recent developments and future directions. BMC Med. Genom. 2015, 8 (Suppl. 1), S2. [Google Scholar] [CrossRef] [PubMed]
- Abadi, A.; Peralta-Romero, J.; Suarez, F.; Gomez-Zamudio, J.; Burguete-Garcia, A.I.; Cruz, M.; Meyre, D. Assessing the effects of 35 European-derived BMI-associated SNPs in Mexican children. Obesity 2016, 24, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
- Antunez-Ortiz, D.L.; Flores-Alfaro, E.; Burguete-Garcia, A.I.; Bonnefond, A.; Peralta-Romero, J.; Froguel, P.; Espinoza-Rojo, M.; Cruz, M. Copy Number Variations in Candidate Genes and Intergenic Regions Affect Body Mass Index and Abdominal Obesity in Mexican Children. Biomed. Res. Int. 2017, 2017, 2432957. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.L.; Guo, Y.; Li, S.M.; Li, S.K.; Tian, Q.; Liu, Y.J.; Deng, H.W. Ethnic differentiation of copy number variation on chromosome 16p12.3 for association with obesity phenotypes in European and Chinese populations. Int. J. Obes. 2013, 37, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Aerts, E.; Beckers, S.; Zegers, D.; Van Hoorenbeeck, K.; Massa, G.; Verrijken, A.; Verhulst, S.L.; Van Gaal, L.F.; Van Hul, W. CNV analysis and mutation screening indicate an important role for the NPY4R gene in human obesity. Obesity 2016, 24, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Shebanits, K.; Andersson-Assarsson, J.C.; Larsson, I.; Carlsson, L.M.S.; Feuk, L.; Larhammar, D. Copy number of pancreatic polypeptide receptor gene NPY4R correlates with body mass index and waist circumference. PLoS ONE 2018, 13, e0194668. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Cao, M.; Shi, J.; Li, L.; Miao, L.; Hong, J.; Cui, B.; Ning, G. Copy number variations of obesity relevant loci associated with body mass index in young Chinese. Gene 2013, 516, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Choquette, A.C.; Bouchard, L.; Drapeau, V.; Lemieux, S.; Tremblay, A.; Bouchard, C.; Vohl, M.C.; Perusse, L. Association between olfactory receptor genes, eating behavior traits and adiposity: Results from the Quebec Family Study. Physiol. Behav. 2012, 105, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Riera, C.E.; Tsaousidou, E.; Halloran, J.; Follett, P.; Hahn, O.; Pereira, M.M.A.; Ruud, L.E.; Alber, J.; Tharp, K.; Anderson, C.M.; et al. The Sense of Smell Impacts Metabolic Health and Obesity. Cell Metab. 2017, 26, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, Z.; Wang, H.; Yang, M.; Liang, L.; Fu, J.; Wang, C.; Ling, J.; Zhang, Y.; Zhang, S.; et al. Interactions between obesity-related copy number variants and dietary behaviors in childhood obesity. Nutrients 2015, 7, 3054–3066. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Liu, H.; Wang, L.; Shen, H.; Hu, W. The Genome-Wide Influence on Human BMI Depends on Physical Activity, Life Course, and Historical Period. Demography 2015, 52, 1651–1670. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.R.; Song, J.Y.; Ma, J.; Liu, F.H.; Shang, X.R.; Guo, X.J.; Wang, H.J. Association study of childhood obesity with eight genetic variants recently identified by genome-wide association studies. Pediatr. Res. 2014, 76, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Felix, J.F.; Bradfield, J.P.; Monnereau, C.; van der Valk, R.J.; Stergiakouli, E.; Chesi, A.; Gaillard, R.; Feenstra, B.; Thiering, E.; Kreiner-Moller, E.; et al. Genome-wide association analysis identifies three new susceptibility loci for childhood body mass index. Hum. Mol. Genet. 2016, 25, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Viljakainen, H.; Andersson-Assarsson, J.C.; Armenio, M.; Pekkinen, M.; Pettersson, M.; Valta, H.; Lipsanen-Nyman, M.; Makitie, O.; Lindstrand, A. Low Copy Number of the AMY1 Locus Is Associated with Early-Onset Female Obesity in Finland. PLoS ONE 2015, 10, e0131883. [Google Scholar] [CrossRef] [PubMed]
- Marcovecchio, M.L.; Florio, R.; Verginelli, F.; De Lellis, L.; Capelli, C.; Verzilli, D.; Chiarelli, F.; Mohn, A.; Cama, A. Low AMY1 Gene Copy Number Is Associated with Increased Body Mass Index in Prepubertal Boys. PLoS ONE 2016, 11, e0154961. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; Yengo, L.; Dechaume, A.; Canouil, M.; Castelain, M.; Roger, E.; Allegaert, F.; Caiazzo, R.; Raverdy, V.; Pigeyre, M.; et al. Relationship between salivary/pancreatic amylase and body mass index: A systems biology approach. BMC Med. 2017, 15, 37. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.I.; Wiley, A.S. Rethinking the starch digestion hypothesis for AMY1 copy number variation in humans. Am. J. Phys. Anthropol. 2017, 163, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Inchley, C.E.; Larbey, C.D.; Shwan, N.A.; Pagani, L.; Saag, L.; Antao, T.; Jacobs, G.; Hudjashov, G.; Metspalu, E.; Mitt, M.; et al. Selective sweep on human amylase genes postdates the split with Neanderthals. Sci. Rep. 2016, 6, 37198. [Google Scholar] [CrossRef] [PubMed]
- Mejia-Benitez, M.A.; Bonnefond, A.; Yengo, L.; Huyvaert, M.; Dechaume, A.; Peralta-Romero, J.; Klunder-Klunder, M.; Garcia Mena, J.; El-Sayed Moustafa, J.S.; Falchi, M.; et al. Beneficial effect of a high number of copies of salivary amylase AMY1 gene on obesity risk in Mexican children. Diabetologia 2015, 58, 290–294. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, M.F.; Roager, H.M.; Larsen, T.M.; Poulsen, S.K.; Licht, T.R.; Bahl, M.I.; Zohar, Y.; Astrup, A. Pre-treatment microbial Prevotella-to-Bacteroides ratio, determines body fat loss success during a 6-month randomized controlled diet intervention. Int. J. Obes. 2018, 42, 284. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, M.F.; Blaedel, T.; Bendtsen, L.Q.; Lorenzen, J.K.; Holm, J.B.; Kiilerich, P.; Roager, H.M.; Kristiansen, K.; Larsen, L.H.; Astrup, A. Prevotella-to-Bacteroides ratio predicts body weight and fat loss success on 24-week diets varying in macronutrient composition and dietary fiber: Results from a post-hoc analysis. Int. J. Obes. 2018. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Gene | CN Range | Classification | Normal Weight | Obese | Association | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | OR | 95% CI | P-value | ||||
| 11q11 | OR4P4, OR4S2 | 0–8 | Deletion | 199 | 41.0 | 151 | 34.6 | 0.774 | 0.634–0.945 | 0.047 |
| OR4C6 | ≥2 copies | 286 | 59.0 | 285 | 65.4 | |||||
| 1p21.1 | AMY1 | 2–19 | Less 6 copies | 116 | 23.9 | 128 | 29.4 | 1.323 | 0.984–1.780 | 0.064 |
| ≥6 copies | 369 | 76.1 | 308 | 70.6 | ||||||
| 16p12.3 | GPRC5B | 1–4 | Deletion | 11 | 2.3 | 13 | 2.9 | 1.307 | 0.472–3.622 | 0.606 |
| ≥2 copies | 474 | 97.7 | 423 | 97.1 | ||||||
| 1p31.1 | NEGR1 | 1–4 | Deletion | 25 | 5.2 | 23 | 5.3 | 1.078 | 0.425–3.622 | 0.867 |
| ≥2 copies | 460 | 94.8 | 413 | 94.7 | ||||||
| 10q11.22 | NPY4R | 1–4 | Deletion | 21 | 4.3 | 19 | 4.4 | 1.064 | 0.411–2.758 | 0.898 |
| ≥2 copies | 464 | 95.7 | 417 | 95.6 | ||||||
| Locus | Gene | CN Range | Classification | Normal Weight | Obese | Association | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | OR | 95% CI | P-value | ||||
| 11q11 | OR4P4, OR4S2 | 0–8 | Deletion | 124 | 32.3 | 186 | 34.7 | 1.054 | 0.877–1.266 | 0.537 |
| OR4C6 | ≥2 copies | 260 | 67.7 | 350 | 65.3 | |||||
| 1p21.1 | AMY1 | 2–19 | Less 6 copies | 94 | 24.5 | 165 | 30.8 | 1.521 | 0.928–2.495 | 0.096 |
| ≥6 copies | 290 | 75.5 | 371 | 69.2 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
León-Mimila, P.; Villamil-Ramírez, H.; López-Contreras, B.E.; Morán-Ramos, S.; Macias-Kauffer, L.R.; Acuña-Alonzo, V.; Del Río-Navarro, B.E.; Salmerón, J.; Velazquez-Cruz, R.; Villarreal-Molina, T.; et al. Low Salivary Amylase Gene (AMY1) Copy Number Is Associated with Obesity and Gut Prevotella Abundance in Mexican Children and Adults. Nutrients 2018, 10, 1607. https://doi.org/10.3390/nu10111607
León-Mimila P, Villamil-Ramírez H, López-Contreras BE, Morán-Ramos S, Macias-Kauffer LR, Acuña-Alonzo V, Del Río-Navarro BE, Salmerón J, Velazquez-Cruz R, Villarreal-Molina T, et al. Low Salivary Amylase Gene (AMY1) Copy Number Is Associated with Obesity and Gut Prevotella Abundance in Mexican Children and Adults. Nutrients. 2018; 10(11):1607. https://doi.org/10.3390/nu10111607
Chicago/Turabian StyleLeón-Mimila, Paola, Hugo Villamil-Ramírez, Blanca E. López-Contreras, Sofía Morán-Ramos, Luis R. Macias-Kauffer, Víctor Acuña-Alonzo, Blanca E. Del Río-Navarro, Jorge Salmerón, Rafael Velazquez-Cruz, Teresa Villarreal-Molina, and et al. 2018. "Low Salivary Amylase Gene (AMY1) Copy Number Is Associated with Obesity and Gut Prevotella Abundance in Mexican Children and Adults" Nutrients 10, no. 11: 1607. https://doi.org/10.3390/nu10111607
APA StyleLeón-Mimila, P., Villamil-Ramírez, H., López-Contreras, B. E., Morán-Ramos, S., Macias-Kauffer, L. R., Acuña-Alonzo, V., Del Río-Navarro, B. E., Salmerón, J., Velazquez-Cruz, R., Villarreal-Molina, T., Aguilar-Salinas, C. A., & Canizales-Quinteros, S. (2018). Low Salivary Amylase Gene (AMY1) Copy Number Is Associated with Obesity and Gut Prevotella Abundance in Mexican Children and Adults. Nutrients, 10(11), 1607. https://doi.org/10.3390/nu10111607