Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target



 , and
, and
Abstract
1. Introduction
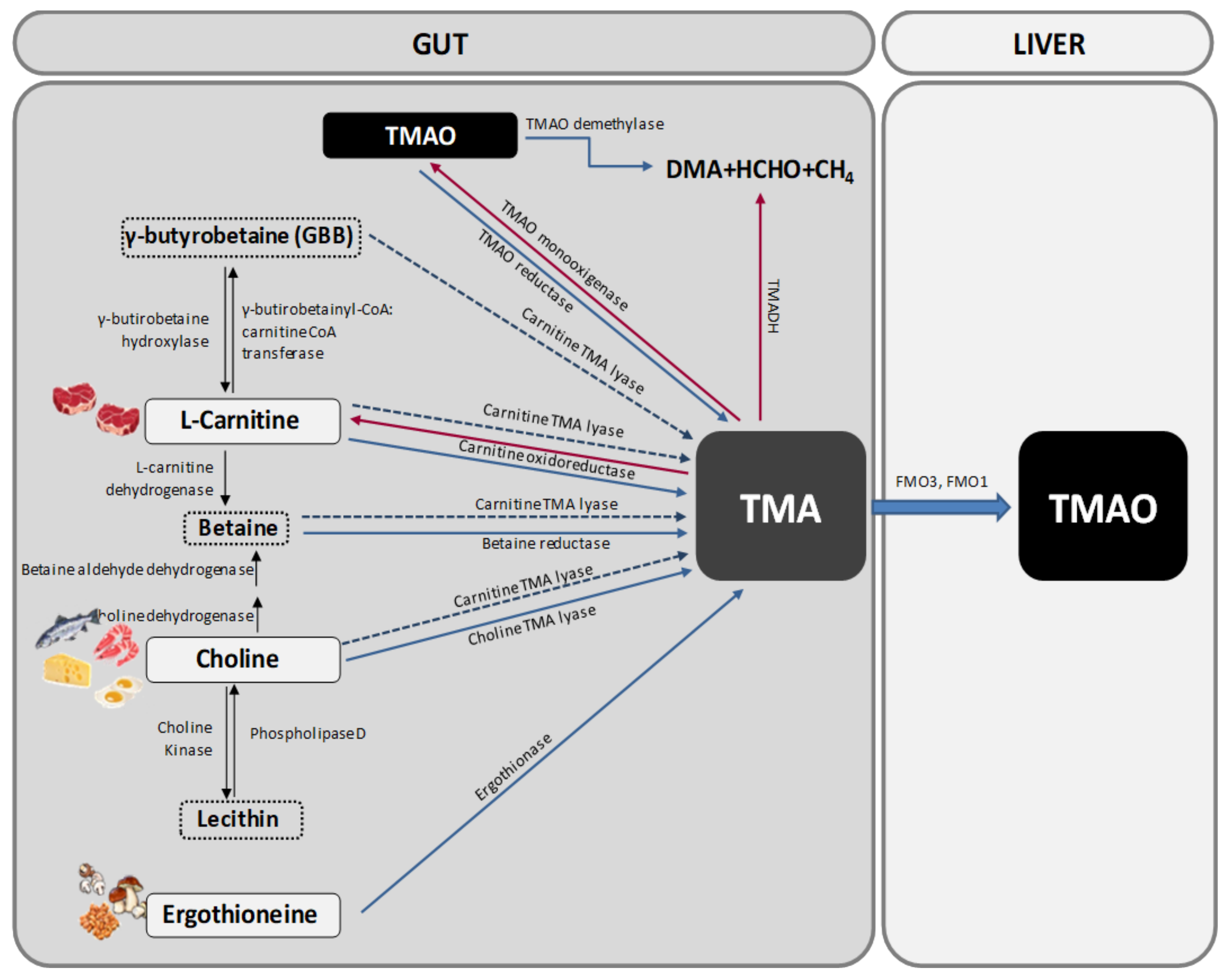
1.1. TMAO Metabolism
1.2. TMA and TMAO Distribution and Excretion
1.3. TMAO Detection and Measurement
1.4. Variations in TMAO Levels
2. Importance of Gut Microbiota in TMAO Metabolism
3. Relationship between Atherosclerosis, Cardiovascular Disease and TMAO
4. Relationship between TMAO and Neurological Disorders
5. Inflammation as the Underlying Mechanism of the Deleterious Effects of TMAO
6. Therapeutic Strategies
7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dumas, M.E.; Barton, R.H.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.C.; et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. USA 2006, 103, 12511–12516. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.; Wong, F.S.; Szot, G.L.; Bluestone, J.A.; et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.L.; Gordon, J.I. Our unindicted coconspirators: Human metabolism from a microbial perspective. Cell Metab. 2010, 12, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zheng, X.; Feng, M.; Li, D.; Zhang, H. Gut Microbiota-Dependent Metabolite Trimethylamine N-Oxide Contributes to Cardiac Dysfunction in Western Diet-Induced Obese Mice. Front. Physiol. 2017, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Zhu, X.H.; Ran, L.; Lang, H.D.; Yi, L.; Mi, M.T. Trimethylamine-N-Oxide Induces Vascular Inflammation by Activating the NLRP3 Inflammasome Through the SIRT3-SOD2-mtROS Signaling Pathway. J. Am. Heart Assoc. 2017, 6, e006347. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, S.; Linseisen, J.; Allenspach, M.; von Eckardstein, A.; Müller, D. Plasma Concentrations of Trimethylamine-N-oxide Are Directly Associated with Dairy Food Consumption and Low-Grade Inflammation in a German Adult Population. J. Nutr. 2016, 146, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jiao, X.; Ma, Y.; Liu, Y.; Zhang, L.; He, Y.; Chen, Y. Trimethylamine N-oxide induces inflammation and endothelial dysfunction in human umbilical vein endothelial cells via activating ROS-TXNIP-NLRP3 inflammasome. Biochem. Biophys. Res. Commun. 2016, 481, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Yang, X.; Li, J.; Chen, X.; Zhao, X.; Chen, Y.; Wen, Y. Trimethylamine N-oxide prime NLRP3 inflammasome via inhibiting ATG16L1-induced autophagy in colonic epithelial cells. Biochem. Biophys. Res. Commun. 2017, 490, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Yang, C.; Wang, B.; Zhang, X.; Hu, T.; Gu, Y.; Li, J. Trimethylamine N-oxide promotes atherosclerosis via CD36-dependent MAPK/JNK pathway. Biomed. Pharmacother. 2018, 97, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Levison, B.S.; Culley, M.K.; Buffa, J.A.; Wang, Z.; Gregory, J.C.; Org, E.; Wu, Y.; Li, L.; Smith, J.D.; Tang, W.H.; et al. γ-Butyrobetaine is a proatherogenic intermediate in gut microbial metabolism of L-carnitine to TMAO. Cell Metab. 2014, 20, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Najar, A.G.; Yaghoobi, M.M.; Jahani, Y.; Vahabzadeh, Z. Trimethylamine-N-Oxide Treatment Induces Changes in the ATP-Binding Cassette Transporter A1 and Scavenger Receptor A1 in Murine Macrophage J774A.1 cells. Inflammation 2016, 39, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Roberts, A.B.; Buffa, J.A.; Levison, B.S.; Zhu, W.; Org, E.; Gu, X.; Huang, Y.; Zamanian-Daryoush, M.; Culley, M.K.; et al. Non-lethal Inhibition of Gut Microbial Trimethylamine Production for the Treatment of Atherosclerosis. Cell 2015, 163, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H.; Warrier, M. Trimethylamine N-Oxide, the Microbiome, and Heart and Kidney Disease. Annu. Rev. Nutr. 2017, 37, 157–181. [Google Scholar] [CrossRef] [PubMed]
- Fennema, D.; Phillips, I.R.; Shephard, E.A. Trimethylamine and Trimethylamine N-Oxide, a Flavin-Containing Monooxygenase 3 (FMO3)-Mediated Host-Microbiome Metabolic Axis Implicated in Health and Disease. Drug Metab. Dispos. 2016, 44, 1839–1850. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Jameson, E.; Crosatti, M.; Schäfer, H.; Rajakumar, K.; Bugg, T.D.; Chen, Y. Carnitine metabolism to trimethylamine by an unusual Rieske-type oxygenase from human microbiota. Proc. Natl. Acad. Sci. USA 2014, 111, 4268–4273. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Patel, N.A.; Crombie, A.; Scrivens, J.H.; Murrell, J.C. Bacterial flavin-containing monooxygenase is trimethylamine monooxygenase. Proc. Natl. Acad. Sci. USA 2011, 108, 17791–17796. [Google Scholar] [CrossRef] [PubMed]
- Chhibber-Goel, J.; Gaur, A.; Singhal, V.; Parakh, N.; Bhargava, B.; Sharma, A. The complex metabolism of trimethylamine in humans: Endogenous and exogenous sources. Expert Rev. Mol. Med. 2016, 18, e8. [Google Scholar] [CrossRef] [PubMed]
- Taesuwan, S.; Cho, C.E.; Malysheva, O.V.; Bender, E.; King, J.H.; Yan, J.; Thalacker-Mercer, A.E.; Caudill, M.A. The metabolic fate of isotopically labeled trimethylamine-N-oxide (TMAO) in humans. J. Nutr. Biochem. 2017, 45, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Veeravalli, S.; Karu, K.; Phillips, I.R.; Shephard, E.A. A highly sensitive liquid chromatography electrospray ionization mass spectrometry method for quantification of TMA, TMAO and creatinine in mouse urine. MethodsX 2017, 4, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Wahlang, B.; Barney, J.; Thompson, B.; Wang, C.; Hamad, O.M.; Hoffman, J.B.; Petriello, M.C.; Morris, A.J.; Hennig, B. Editor’s Highlight: PCB126 Exposure Increases Risk for Peripheral Vascular Diseases in a Liver Injury Mouse Model. Toxicol. Sci. 2017, 160, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. EBioMedicine 2015, 2, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Wekell, J.C.; Barnett, H. New method for analysis of trimethylamine oxide using ferrous sulfate and EDTA. J. Food Sci. 1991, 56, 132–138. [Google Scholar] [CrossRef]
- Laxson, C.J.; Condon, N.E.; Drazen, J.C.; Yancey, P.H. Decreasing urea: Trimethylamine N-oxide ratios with depth in chondrichthyes: A physiological depth limit? Physiol. Biochem. Zool. 2011, 84, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Zerbst-Boroffka, I.; Kamaltynow, R.M.; Harjes, S.; Kinne-Saffran, E.; Gross, J. TMAO and other organic osmolytes in the muscles of amphipods (Crustacea) from shallow and deep water of Lake Baikal. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Zimetti, F.; Caffarra, P.; Tassotti, M.; Bernini, F.; Brighenti, F.; Zini, A.; Zanotti, I. The Gut Microbial Metabolite Trimethylamine-N-Oxide Is Present in Human Cerebrospinal Fluid. Nutrients 2017, 9, 1053. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, M.T.; Ramezani, A.; Manal, A.; Raj, D.S. Trimethylamine N-Oxide: The Good, the Bad and the Unknown. Toxins 2016, 8, 326. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, Y.; Gua, C.; Li, X. Elevated Circulating Trimethylamine N-Oxide Levels Contribute to Endothelial Dysfunction in Aged Rats through Vascular Inflammation and Oxidative Stress. Front. Physiol. 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Levison, B.S.; Hazen, J.E.; Donahue, L.; Li, X.M.; Hazen, S.L. Measurement of trimethylamine-N-oxide by stable isotope dilution liquid chromatography tandem mass spectrometry. Anal. Biochem. 2014, 455, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Boutagy, N.E.; Neilson, A.P.; Osterberg, K.L.; Smithson, A.T.; Englund, T.R.; Davy, B.M.; Hulver, M.W.; Davy, K.P. Probiotic supplementation and trimethylamine-N-oxide production following a high-fat diet. Obesity 2015, 23, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Boutagy, N.E.; Neilson, A.P.; Osterberg, K.L.; Smithson, A.T.; Englund, T.R.; Davy, B.M.; Hulver, M.W.; Davy, K.P. Short-term high-fat diet increases postprandial trimethylamine-N-oxide in humans. Nutr. Res. 2015, 35, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xu, J.; Jiang, C.; Zhang, Y.; Xue, Y.; Li, Z.; Wang, J.; Xue, C.; Wang, Y. Fish oil ameliorates trimethylamine N-oxide-exacerbated glucose intolerance in high-fat diet-fed mice. Food Funct. 2015, 6, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.Y. Intestinal phospholipid and lysophospholipid metabolism in cardiometabolic disease. Curr. Opin. Lipidol. 2016, 27, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Yin, Z.; Liu, N.; Bian, X.; Yu, R.; Su, X.; Zhang, B.; Wang, Y. Gut microbial metabolite TMAO contributes to renal dysfunction in a mouse model of diet-induced obesity. Biochem. Biophys. Res. Commun. 2017, 493, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, F.; Wang, Y.; Ma, X.; Zhao, M.; Zhao, C. Metabonomics study of the therapeutic mechanism of Gynostemma pentaphyllum and atorvastatin for hyperlipidemia in rats. PLoS ONE 2013, 8, e78731. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.G.; Winning, H.; Savorani, F.; Toft, H.; Larsen, T.M.; Dragsted, L.O.; Astrup, A.; Engelsen, S.B. Assessment of the effect of high or low protein diet on the human urine metabolome as measured by NMR. Nutrients 2012, 4, 112–131. [Google Scholar] [CrossRef] [PubMed]
- Ufnal, M.; Zadlo, A.; Ostaszewski, R. TMAO: A small molecule of great expectations. Nutrition 2015, 31, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Mafra, D.; Borges, N.A.; Cardozo, L.F.M.F.; Anjos, J.S.; Black, A.P.; Moraes, C.; Bergman, P.; Lindholm, B.; Stenvinkel, P. Red meat intake in chronic kidney disease patients: Two sides of the coin. Nutrition 2018, 46, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, N.; Williams, P.T.; Lamendella, R.; Faghihnia, N.; Grube, A.; Li, X.; Wang, Z.; Knight, R.; Jansson, J.K.; Hazen, S.L.; et al. Diets high in resistant starch increase plasma levels of trimethylamine-N-oxide, a gut microbiome metabolite associated with CVD risk. Br. J. Nutr. 2016, 116, 2020–2029. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alonso, P.; Cañueto, D.; Giardina, S.; Salas-Salvadó, J.; Cañellas, N.; Correig, X.; Bulló, M. Effect of pistachio consumption on the modulation of urinary gut microbiota-related metabolites in prediabetic subjects. J. Nutr. Biochem. 2017, 45, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Sun, S.; Liu, L.; Zhang, Q.; Guo, F.; Li, C.; Feng, R.; Sun, C. Effects of Histidine Supplementation on Global Serum and Urine (1)H NMR-based Metabolomics and Serum Amino Acid Profiles in Obese Women from a Randomized Controlled Study. J. Proteome Res. 2017, 16, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Awwad, H.M.; Rabagny, Y.; Graeber, S.; Herrmann, W.; Geisel, J. Plasma trimethylamine N-oxide concentration is associated with choline, phospholipids, and methyl metabolism. Am. J. Clin. Nutr. 2016, 103, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Mueller, D.M.; Allenspach, M.; Othman, A.; Saely, C.H.; Muendlein, A.; Vonbank, A.; Drexel, H.; von Eckardstein, A. Plasma levels of trimethylamine-N-oxide are confounded by impaired kidney function and poor metabolic control. Atherosclerosis 2015, 243, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, J.R.; House, J.A.; Ocque, A.J.; Zhang, S.; Johnson, C.; Kimber, C.; Schmidt, K.; Gupta, A.; Wetmore, J.B.; Nolin, T.D.; et al. Serum Trimethylamine-N-Oxide is Elevated in CKD and Correlates with Coronary Atherosclerosis Burden. J. Am. Soc. Nephrol. 2016, 27, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Missailidis, C.; Hällqvist, J.; Qureshi, A.R.; Barany, P.; Heimbürger, O.; Lindholm, B.; Stenvinkel, P.; Bergman, P. Serum Trimethylamine-N-Oxide Is Strongly Related to Renal Function and Predicts Outcome in Chronic Kidney Disease. PLoS ONE 2016, 11, e0141738. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, B.; Fitzpatrick, M.; Al-Nuaimi, H.; Coughlan, A.M.; Hickey, F.B.; Pusey, C.D.; Savage, C.; Benton, C.M.; O’Brien, E.C.; O’Toole, D.; et al. Changes in urinary metabolomic profile during relapsing renal vasculitis. Sci. Rep. 2016, 6, 38074. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Lever, M.; George, P.M.; Slow, S.; Bellamy, D.; Young, J.M.; Ho, M.; McEntyre, C.J.; Elmslie, J.L.; Atkinson, W.; Molyneux, S.L.; et al. Betaine and Trimethylamine-N-Oxide as Predictors of Cardiovascular Outcomes Show Different Patterns in Diabetes Mellitus: An Observational Study. PLoS ONE 2014, 9, e114969. [Google Scholar] [CrossRef] [PubMed]
- Mente, A.; Chalcraft, K.; Ak, H.; Davis, A.D.; Lonn, E.; Miller, R.; Potter, M.A.; Yusuf, S.; Anand, S.S.; McQueen, M.J. The Relationship Between Trimethylamine-N-Oxide and Prevalent Cardiovascular Disease in a Multiethnic Population Living in Canada. Can. J. Cardiol. 2015, 31, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Trøseid, M.; Ueland, T.; Hov, J.R.; Svardal, A.; Gregersen, I.; Dahl, C.P.; Aakhus, S.; Gude, E.; Bjørndal, B.; Halvorsen, B.; et al. Microbiota-dependent metabolite trimethylamine-N-oxide is associated with disease severity and survival of patients with chronic heart failure. J. Intern. Med. 2015, 277, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Wang, Z.; Shrestha, K.; Borowski, A.G.; Wu, Y.; Troughton, R.W.; Klein, A.L.; Hazen, S.L. Intestinal microbiota-dependent phosphatidylcholine metabolites, diastolic dysfunction, and adverse clinical outcomes in chronic systolic heart failure. J. Card. Fail. 2015, 21, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, W.H.; Buffa, J.A.; Fu, X.; Britt, E.B.; Koeth, R.A.; Levison, B.S.; Fan, Y.; Wu, Y.; Hazen, S.L. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur. Heart J. 2014, 35, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Pan, B.; Chen, Y.; Guo, C.; Zhao, M.; Zheng, L.; Chen, B. Trimethylamine N-oxide in atherogenesis: Impairing endothelial self-repair capacity and enhancing monocyte adhesion. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Seldin, M.M.; Meng, Y.; Qi, H.; Zhu, W.; Wang, Z.; Hazen, S.L.; Lusis, A.J.; Shih, D.M. Trimethylamine N-Oxide Promotes Vascular Inflammation Through Signaling of Mitogen-Activated Protein Kinase and Nuclear Factor-κB. J. Am. Heart Assoc. 2016, 5, e002767. [Google Scholar] [CrossRef] [PubMed]
- Al-Obaide, M.A.I.; Singh, R.; Datta, P.; Rewers-Felkins, K.A.; Salguero, M.V.; Al-Obaidi, I.; Kottapalli, K.R.; Vasylyeva, T.L. Gut Microbiota-Dependent Trimethylamine-N-oxide and Serum Biomarkers in Patients with T2DM and Advanced CKD. J. Clin. Med. 2017, 6, e86. [Google Scholar] [CrossRef] [PubMed]
- Hove-Skovsgaard, M.; Gaardbo, J.C.; Kolte, L.; Winding, K.; Seljeflot, I.; Svardal, A.; Berge, R.K.; Gerstoft, J.; Ullum, H.; Trøseid, M.; Nielsen, S.D. HIV-infected persons with type 2 diabetes show evidence of endothelial dysfunction and increased inflammation. BMC Infect. Dis. 2017, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Charach, G.; Rabinovich, P.D.; Konikoff, F.M.; Grosskopf, I.; Weintraub, M.S.; Gilat, T. Decreased fecal bile acid output in patients with coronary atherosclerosis. J. Med. 1998, 29, 125–136. [Google Scholar] [PubMed]
- Charach, G.; Rabinovich, A.; Argov, O.; Weintraub, M.; Rabinovich, P. The role of bile Acid excretion in atherosclerotic coronary artery disease. Int. J. Vasc. Med. 2012, 2012, 949672. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Feskens, E.J.; Boer, J.M.; Müller, M. The potential influence of genetic variants in genes along bile acid and bile metabolic pathway on blood cholesterol levels in the population. Atherosclerosis 2010, 210, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Narath, S.H.; Mautner, S.I.; Svehlikova, E.; Schultes, B.; Pieber, T.R.; Sinner, F.M.; Gander, E.; Libiseller, G.; Schimek, M.G.; Sourij, H.; et al. An Untargeted Metabolomics Approach to Characterize Short-Term and Long-Term Metabolic Changes after Bariatric Surgery. PLoS ONE 2016, 11, e0161425. [Google Scholar] [CrossRef] [PubMed]
- Trøseid, M.; Hov, J.R.; Nestvold, T.K.; Thoresen, H.; Berge, R.K.; Svardal, A.; Lappegård, K.T. Major Increase in Microbiota-Dependent Proatherogenic Metabolite TMAO One Year After Bariatric Surgery. Metab. Syndr. Relat. Disord. 2016, 14, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Nowiński, A.; Ufnal, M. Trimethylamine N-oxide: A harmful, protective or diagnostic marker in lifestyle diseases? Nutrition 2018, 46, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Johri, A.M.; Heyland, D.K.; Hétu, M.F.; Crawford, B.; Spence, J.D. Carnitine therapy for the treatment of metabolic syndrome and cardiovascular disease: Evidence and controversies. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Yamagishi, S.; Sakai, K.; Kaida, Y.; Yokoro, M.; Ueda, S.; Wada, Y.; Takeuchi, M.; Shimizu, M.; Yamazaki, H.; et al. Oral L-carnitine supplementation increases trimethylamine-N-oxide but reduces markers of vascular injury in hemodialysis patients. J. Cardiovasc. Pharmacol. 2015, 65, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Collins, H.L.; Drazul-Schrader, D.; Sulpizio, A.C.; Koster, P.D.; Williamson, Y.; Adelman, S.J.; Owen, K.; Sanli, T.; Bellamine, A. L-Carnitine intake and high trimethylamine N-oxide plasma levels correlate with low aortic lesions in ApoE(-/-) transgenic mice expressing CETP. Atherosclerosis 2016, 244, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Bain, M.A.; Fornasini, G.; Evans, A.M. Trimethylamine: Metabolic, pharmacokinetic and safety aspects. Curr. Drug Metab. 2005, 6, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.E.; Taesuwan, S.; Malysheva, O.V.; Bender, E.; Tulchinsky, N.F.; Yan, J.; Sutter, J.L.; Caudill, M.A. Trimethylamine-N-oxide (TMAO) response to animal source foods varies among healthy young men and is influenced by their gut microbiota composition: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, R.; Knekt, P.; Rissanen, H.; Reunanen, A. Intake of fish and long-chain n-3 fatty acids and the risk of coronary heart mortality in men and women. Br. J. Nutr. 2006, 95, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, K.; Iso, H.; Date, C.; Fukui, M.; Wakai, K.; Kikuchi, S.; Inaba, Y.; Tanabe, N.; Tamakoshi, A.; Japan Collaborative Cohort Study for Evaluation of Cancer Risk Study Group. Fish, omega-3 polyunsaturated fatty acids, and mortality from cardiovascular diseases in a nationwide community-based cohort of Japanese men and women the JACC (Japan Collaborative Cohort Study for Evaluation of Cancer Risk) Study. J. Am. Coll. Cardiol. 2008, 52, 988–996. [Google Scholar] [PubMed]
- Warrier, M.; Shih, D.M.; Burrows, A.C.; Ferguson, D.; Gromovsky, A.D.; Brown, A.L.; Marshall, S.; McDaniel, A.; Schugar, R.C.; Wang, Z.; et al. The TMAO-Generating Enzyme Flavin Monooxygenase 3 Is a Central Regulator of Cholesterol Balance. Cell Rep. 2015, 10, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Ling, A.V.; Manthena, P.V.; Gearing, M.E.; Graham, M.J.; Crooke, R.M.; Croce, K.J.; Esquejo, R.M.; Clish, C.B.; Morbid Obesity Study Group; Vicent, D.; et al. Flavin-containing monooxygenase 3 as a potential player in diabetes-associated atherosclerosis. Nat. Commun. 2015, 6, 6498. [Google Scholar] [CrossRef] [PubMed]
- Vernetti, L.; Gough, A.; Baetz, N.; Blutt, S.; Broughman, J.R.; Brown, J.A.; Foulke-Abel, J.; Hasan, N.; In, J.; Kelly, E.; et al. Functional Coupling of Human Microphysiology Systems: Intestine, Liver, Kidney Proximal Tubule, Blood-Brain Barrier and Skeletal Muscle. Sci. Rep. 2017, 7, 42296. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Fletcher, C. Trimethylamine N-oxide: Breathe new life. Br. J. Pharmacol. 2018, 175, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Wang, Q. Towards understanding brain-gut-microbiome connections in Alzheimer’s disease. BMC Syst. Biol. 2016, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.S.; Yip, C.M.; Huang, T.H.; Chakrabartty, A.; Fraser, P.E. Manipulating the amyloid-beta aggregation pathway with chemical chaperones. J. Biol. Chem. 1999, 274, 32970–32974. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Graves, D.J. Natural methylamine osmolytes, trimethylamine N-oxide and betaine, increase tau-induced polymerization of microtubules. Biochem. Biophys. Res. Commun. 1998, 250, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Crowther, R.A.; Goedert, M. The natural osmolyte trimethylamine N-oxide (TMAO) restores the ability of mutant tau to promote microtubule assembly. FEBS Lett. 2000, 484, 265–270. [Google Scholar] [CrossRef]
- Tseng, H.C.; Lu, Q.; Henderson, E.; Graves, D.J. Phosphorylated tau can promote tubulin assembly. Proc. Natl. Acad. Sci. USA 1999, 96, 9503–9508. [Google Scholar] [CrossRef] [PubMed]
- Tatzelt, J.; Prusiner, S.B.; Welch, W.J. Chemical chaperones interfere with the formation of scrapie prion protein. EMBO J. 1996, 15, 6363–6373. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Yoshizawa, T.; Shibasaki, F.; Shoji, S.; Kanazawa, I. Chemical chaperones reduce aggregate formation and cell death caused by the truncated Machado-Joseph disease gene product with an expanded polyglutamine stretch. Neurobiol. Dis. 2002, 10, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Getter, T.; Zaks, I.; Barhum, Y.; Ben-Zur, T.; Böselt, S.; Gregoire, S.; Viskind, O.; Shani, T.; Gottlieb, H.; Green, O.; et al. A chemical chaperone-based drug candidate is effective in a mouse model of amyotrophic lateral sclerosis (ALS). ChemMedChem 2015, 10, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Jamal, S.; Kumari, A.; Singh, A.; Goyal, S.; Grover, A. Conformational Ensembles of α-Synuclein Derived Peptide with Different Osmolytes from Temperature Replica Exchange Sampling. Front. Neurosci. 2017, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Li, J.; Fink, A.L. Trimethylamine-N-oxide-induced folding of alpha-synuclein. FEBS Lett. 2001, 509, 31–35. [Google Scholar] [CrossRef]
- Boini, K.M.; Hussain, T.; Li, P.L.; Koka, S. Trimethylamine-N-Oxide Instigates NLRP3 Inflammasome Activation and Endothelial Dysfunction. Cell. Physiol. Biochem. 2017, 44, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Saco, T.; Parthasarathy, P.T.; Cho, Y.; Lockey, R.F.; Kolliputi, N. Inflammasome: A new trigger of Alzheimer’s disease. Front. Aging Neurosci. 2014, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.P.; Wang, Y.; Sprenger, N.; Yap, I.K.; Lundstedt, T.; Lek, P.; Rezzi, S.; Ramadan, Z.; van Bladeren, P.; Fay, L.B.; et al. Probiotic modulation of symbiotic gut microbial-host metabolic interactions in a humanized microbiome mouse model. Mol. Syst. Biol. 2008, 4, 157. [Google Scholar] [CrossRef] [PubMed]
- Brugère, J.F.; Borrel, G.; Gaci, N.; Tottey, W.; O’Toole, P.W.; Malpuech-Brugère, C. Archaebiotics: Proposed therapeutic use of archaea to prevent trimethylaminuria and cardiovascular disease. Gut Microbes 2014, 5, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Dridi, B.; Fardeau, M.L.; Ollivier, B.; Raoult, D.; Drancourt, M. Methanomassiliicoccus luminyensis gen. nov., sp. nov., a methanogenic archaeon isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2012, 62, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, A.R.; Larrick, J.W. Dietary modification of the microbiome affects risk for cardiovascular disease. Rejuvenation Res. 2013, 16, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, Y.; Tang, W.H. Intersections Between Microbiome and Heart Failure: Revisiting the Gut Hypothesis. J. Card. Fail. 2015, 21, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Spector, R. New Insight into the Dietary Cause of Atherosclerosis: Implications for Pharmacology. J. Pharmacol. Exp. Ther. 2016, 358, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Dambrova, M.; Skapare-Makarova, E.; Konrade, I.; Pugovics, O.; Grinberga, S.; Tirzite, D.; Petrovska, R.; Kalvins, I.; Liepins, E. Meldonium decreases the diet-increased plasma levels of trimethylamine N-oxide, a metabolite associated with atherosclerosis. J. Clin. Pharmacol. 2013, 53, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Kuka, J.; Liepinsh, E.; Makrecka-Kuka, M.; Liepins, J.; Cirule, H.; Gustina, D.; Loza, E.; Zharkova-Malkova, O.; Grinberga, S.; Pugovics, O.; et al. Suppression of intestinal microbiota-dependent production of pro-atherogenic trimethylamine N-oxide by shifting L-carnitine microbial degradation. Life Sci. 2014, 117, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.M.; Wang, Z.; Lee, R.; Meng, Y.; Che, N.; Charugundla, S.; Qi, H.; Wu, J.; Pan, C.; Brown, J.M.; et al. Flavin containing monooxygenase 3 exerts broad effects on glucose and lipid metabolism and atherosclerosis. J. Lipid Res. 2015, 56, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wang, X.; Cao, R.; Zhang, Q.; Liu, Q.; Xu, M.; Zhang, M.; Du, X.; Dong, F.; Yan, X. NMR-based metabonomics study on the effect of Gancao in the attenuation of toxicity in rats induced by Fuzi. J. Ethnopharmacol. 2016, 193, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.D.; Zhang, Q.Y.; Mi, M.T. Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. MBio 2016, 7, e02210–15. [Google Scholar] [CrossRef] [PubMed]
- Konop, M.; Radkowski, M.; Grochowska, M.; Perlejewski, K.; Samborowska, E.; Ufnal, M. Enalapril decreases rat plasma concentration of TMAO, a gut bacteria-derived cardiovascular marker. Biomarkers 2018, 23, 380–385. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of diet | Influence on microbiota and TMAO | Consequences and Remarks | References |
|---|---|---|---|
| High fat diet (mice) | ↑ Plasma TMAO | Obesity and metabolic problems (not prevented with the use of DMB) Renal fibrosis, oxidative stress and inflammation of the kidney (prevented with DMB). | Sun G et al., 2017 |
| High fat diet enriched in phosphatydilcholine (rats) | ↑ TMAO in plasma and liver | Hyperlipidemia. TMAO levels decrease and lecithin levels increase with treatment of Gynostemma pentaphyllum, but not with treatment of atorvastatin. | Wang M et al., 2013 |
| High fat diet (human) | ↑ Postpandrial plasma TMAO levels, but not fasting ones. | In the short term, a reduction of plasma TMAO clearance is observed. | Boutagy NE et al., 2015a |
| High fat diet (human) | ↑ Plasma TMAO | The increase of TMAO levels is not prevented with the use of probiotics (VSL#3®), though there is less weight gain and fat. The magnitude of the change in the levels of TMAO is correlated with systolic pressure and carotid pulse. | Boutagy NE et al., 2015b |
| High fat diet | ↑ Firmicutes and Proteobacteria ↓ Bacteroidetes | More production of TMAO. | Hui D, 2016 |
| Low fat diet | ↑ Bacteroidetes and ↓ Firmicutes | Lower production of TMAO. | Hui D, 2016 |
| High fat diet enriched in fish oils (FO) (mice) | ↑ TMAO plasma | FO improve the adverse effects produced by TMAO (tolerance to glucose and adipose tissue inflammation). | Gao X et al., 2015 |
| High protein diet | ↑ TMAO urine | High correlation with daily nitrogen excreted through the urinary tract. | Rasmussen LG et al., 2012 |
| Low protein diet | ↓ TMAO plasma | A diet low in proteins in patients with CKD resulted in lower plasma TMAO levels. | Mafra D et al., 2017 |
| Similar to Western (mice) | ↑ TMAO plasma | Obesity and Dyslipidemia (not prevented with the use of DMB) Cardiac dysfunction and fibrosis of heart with increased expression of Pro-Inflammatory Cytokines, tumor necrosis factor and interleukin IL-1β and reduced expression of anti-inflammatory cytokines (IL-10) (prevented with DMB). | Chen K et al., 2017 |
| Rich in indigestible CH (human) | ↓ Production of TMAO Gut microbiota alterations | Significant weight loss in children with simple obesity or Prader Willi Syndrome (PWS). Better state of inflammation. | Zhang C et al., 2015 |
| Rich in CH and/or low in CH and rich in indigestible starch (human) | ↑ TMAO plasma | The diet does not improve short-term biomarkers of CVR. It mitigates the postprandial glucose and insulin response to hearty meals. | Bergeron N et al., 2016 |
| Supplementation with pistachios (human) | ↓ Production of TMAO | Improvement of metabolic disorders associated with IR and DMII. | Hernández-Alonso P et al., 2017 |
| Supplementation with histidine (human) | ↑ TMAO in plasma and urine. | Lower production of lipids and glucose. | Du et al., 2017 |
| Vegetarian (human) | Changes in gut microbiota | Vegetables of the family Cruciferae can reduce FMO3 activity Reduced ability to produce TMA from L-carnitine. | Koeth RA et al., 2013 |
| Species/Cells | Alterations of TMAO levels, consequences and remarks // Proposed Mechanisms | References |
|---|---|---|
| THP-1 y HUVECs (Human Umbilical Vein Endothelial Cells) | ↑ TMAO levels relates to: ↑ Endothelial dysfunction ↓ Endothelial self-reparation ↑ Adhesion of monocytes through activation of: PKC/NF-κB/VCAM-1 pathways | Ma G et al., 2017 |
| Mouse, HAECs (Human Aoric Endothelial Cells) and VSMC (Vascular Smooth Muscle Cells) | ↑ TMAO levels relates to: ↑ Proinflammatory cytokines via MAPK and NF-κB ↑ Leukocyte adhesion to endothelial wall | Seldin MM et al., 2016 |
| Mouse | ↑ TMAO levels relates to: ↑ Proinflammatory cytokines ↑ Tumor necrosis factor ↑ Interleukin IL-1β ↓ Anti-inflammatory cytokines (IL-10) | Chen K et al., 2017 |
| Human | ↑ TMAO levels relates to prevalent CVD | Wang Z et al., 2011 Mente A et al., 2015 |
| Human | Higher TMAO levels in patients after bariatric surgery | Troseid M et al., 2016 |
| Human | Higher TMAO levels in patients after bariatric surgery in the short and long term | Narath S et al., 2016 |
| Human and mice | TMAO alters cholesterol and sterol metabolism in various compartment TMAO lowers the expression of the main bile acid synthetic enzyme (Cyp7a1) Supplementation of TMAO precursors or even TMAO itself, could also promote suppression of reverse cholesterol transport TMAO also reduces the expression of intestinal cholesterol transporters Niemann-Pick C1-like1 (Npc1L1) | Koeth RA et al., 2013; 2014 Wang Z et al., 2011 |
| Mice | TMAO increases the expression in macrophages of scavenger receptors CD36 and SR-A1, which promote lipid accumulation and foam cell formation | Wang Z et al 2011, 2015 |
| Mice | TMAO enhanced CD36 expression and foam cell formation, which is induced by oxidatively modified low density lipoprotein (ox-LDL). Foam cell formation was also attenuated by the use of siRNA-mediated knockdown of CD36 | Geng J et al., 2018 |
| Rat | Circulating TMAO levels increase with age ↑ Endothelial dysfunction and vascular inflammation via oxidative stress ↑ Expression of pro-inflammatory cytokines ↓ eNOS expression (corrected with DMB) | Li T et al., 2017 |
| Human | Patients with T2D and chronic kidney disease have more amount of TMAO producing microbiota. There is a positive correlation with endothelial dysfunction and inflammatory biomarkers | Al-Obaide MAI et al., 2017 |
| Human | TMAO is correlated with ADMA (marker of endothelial dysfunction) in patients with DMII and HIV, not in the other groups. Uncorrelated to hsCRP | Hove-Skovsgaard et al., 2017 |
| Human | High levels of betaine were associated with CVR only in diabetic patients | Lever M et al., 2014 |
| Human | Elevated levels of TMAO precursors are only associated with higher risk of MACE when high TMAO levels are present concomitantly | Wang Z et al., 2014b |
| Human | Supplementation with L-carnitine seems to improve some features of CVD although it raises plasma TMAO and TMA levels | Fukami K et al., 2015 |
| Mice | TMAO shows positive effects against atherosclerosis in ApoE-/-transgenic mice expressing cholesteryl ester transfer protein CETP | Collins HL et al., 2016 |
| Human | ↑ TMAO levels relates to: ↑ Cardiac failure ↓ survival Diastolic dysfunction Uncorrelated with markers of inflammation | Tang WH et al., 2015b |
| Murine macrophage J774A.1 cells | ↑ TMAO levels relates to: ↑ Expression of SR-A1 (proatherogenic), ↑ Stress in endoplasmic reticulum ↓ ATP-binding cassette transporter A1 | Mohammadi et al., 2016 |
| Experimental conditions | Remarks | References |
|---|---|---|
| Human | Detection of TMAO in cerebrospinal fluid. It seems that TMAO levels are not related to neurological disorders (although it was not the objective of the study). | Del Rio D et al., 2017 |
| Synthesized and purified Aβ peptides | TMAO is able to stabilize and modify the aggregation of the peptide Aβ, favouring and accelerating the transformation of the random string of the Aβ peptide to its β-conformation and stabilizing the resulting protofibrils, that can originate fibers that tend to aggregate and form tangled plates. | Yang DS Et al., 1999 |
| Wild and mutant tau proteins | TMAO is able to promote and enhance the assembly of microtubules in mutant and hyperphosphorylated tau protein, reaching in the majority of cases a greater protein efficiency ratio than in wild-type tau. | Smith MJ Et al., 2000 |
| Purified human recombinant tau | TMAO does not act by dephosphorylating tau protein; it facilitates the binding between tau protein and tubulin by reducing the critical concentration of tubulin necessary for assembly. | Tseng HC et al., 1999 |
| Human | TMAO has been suggested to cause blood brain barrier disruption by reducing the expression of tight junction proteins like claudin-5 and tight junction protein-1 (ZO-1). | Subramaniam S et al., 2018 |
| Purified tau proteins | TMAO can act as a natural osmolyte and stimulates tau-induced tubulin assembly | Tseng HC and Graves DJ, 1998 |
| Scrapie-infected mouse neuroblastoma cells | TMAO inhibits the conversion of the scrapie prion protein (PrPC) into its pathogenic isoform (PrPSc), which is associated with transmissible spongiform encephalopathies. | Tatzelt J et al., 1996 |
| BHK-21 and Neuro2a cells transfected with N-terminal truncated ataxin-3 with an expanded polyglutamine stretch | TMAO has been shown to reduce aggregate formation, cell death and cytotoxicity induced by truncated expanded ataxin-3, which is involved in Machado-Joseph disease/spinocerebellar ataxia-3. | Yoshida H et al., 2002 |
| Mice | A lipophilic derivative of TMAO showed an improvement in neurological functions in mice, preventing endothelial reticulum-stress induced apoptosis of NSC-34 motor neuron-like cells and primary mouse astrocytes. | Getter T et al., 2015 |
| α-synuclein peptides | TMAO suppresses the formation of extended conformations and can act as a protecting osmolyte leading to compact and folded forms of α-synuclein. This effect could probably prevent the alpha-synuclein aggregation and formation of insoluble fibrils that cause Parkinson disease. | Jamal S et al., 2017 |
| Purified recombinantα –synuclein | When the concentration of TMAO is high enough, α-synuclein forms oligomers in which the subunits are folded and are not able to fibrillate. | Uversky V et al., 2001 |
| Species // Cell lines | Alterations of TMAO levels, consequences and remarks // Proposed mechanisms | References |
|---|---|---|
| Human | ↑ PlasmaTMAO levels relates to: ↑ TNF-α, ↑ IL-6 ↑ C-reactive protein ↑ Inflammation | Rohrmann S et al., 2016 |
| Fetal human colon cells (FHC) | TMAO increases state inflammation via NLRP3 inflammasome activation (gets reversed with ATG16L1 overexpression or siRNA-NLRP3 KO) ↑ TMAO levels inhibit cell growth and ↑ apoptosis. It also induces oxidative stress and inhibits the expression of ATG16L1 and p62 and the autophagy of LC3-II | Yue C et al., 2017 |
| Human umbilical vein endothelial cells (HUVEC) | TMAO increases oxidative stress and inflammation via ROS-TXNIP-inflammasome NLRP3. It also increases IL-1β and IL-18 and inhibits eNOS and NO. The effects are reversed with the use of NAC and siRNA-mediated knockdown TXNIP - NLRP3 | Sun X et al., 2016 |
| Mouse and HUVEC | TMAO promotes vascular inflammation by activating the NLRP3 inflammasome ↑ IL-1β, ICAM-1 and MMP-9 ↑ Monocyte adhesion to endothelial cells NLRP3 activation is mediated by inhibition of SIRT3-SOD2-mtROS signalling pathway | Chen ML et al., 2017 |
| Carotid artery endothelial cells (CAEC) | TMAO significantly increases the activation and formation of NLRP3 and caspase-1 activity, ↑ IL-1β production and cell permeability Activation of NLRP3 was abolished with NLRP3 siRNA or caspase-1 inhibitor, WEHD | Boini KM et al., 2017 |
| Therapy | Effects | Remarks and Issues |
|---|---|---|
| Prebiotics | Elicit a favourable impact on gut microbiota composition to decrease TMA formation in the intestine. | Unclear effects in humans. Several factors influence gut microbiota composition |
| Probiotics (I): Bacteria unable to transform precursors into TMA | Decrease TMA formation in the gut | Beneficial effects in mice. However, the effects are not clear in humans |
| Probiotics (II): Methanogenic bacteria | Deplete TMA and TMAO | Safety and engraftment remain unclear in humans |
| Antibiotics | Eliminate TMA-forming microbiota. Nearly total suppression of plasma TMAO levels | Nonspecific, beneficial bacteria are also eradicated. Chronic use is not viable. Repopulation and resistant bacterial strains are likely |
| Oral non-absorbent binders | Remove TMAO or its precursors in the gut | Hypothetical approach. A compound that removes specifically TMAO has not yet been discovered |
| FMO3 enzyme inhibition | Prevents TMA oxidization to TMAO | An accumulation of TMA produces trimethylaminuria, characterized by fishy odor, and could cause inflammation. FMO3 also metabolizes other molecules |
| Gynostemma pentaphyllum | Reduces plasma TMAO levels and increase lecithin levels | The effect of this plant in the other precursors has not been studied. Studies performed in rats |
| Gancao | Avoids the increase in TMAO levels when Fuzi is co-administered | It does not reduce TMAO levels when administered alone. Studies performed in rats |
| Resveratrol | Modulates gut microbiota composition. ↓ TMA and TMAO production | ↑ Lactobacillus and Bifidobacterium No effects when antibiotic are used. Studies performed in mice |
| 3,3-Dimethyldimethyl-1-butanol (DMB) | Inhibits transformation of choline, carnitine and crotonobetaine into TMA through inhibition of microbial TMA lyases | Not able to inhibit the conversion of γ-butyrobetaine to TMA. Studies performed in mice and rats |
| Meldonium | Reduces TMAO biosynthesis from L-carnitine (inhibits the conversion of GBB into L-carnitine) | Not able to reduce TMAO formation from choline. It may increase TMAO urinary excretion in humans |
| Enalapril | Increases urine TMAO excretion | Unknown mechanism. Studies performed in rats. It does not affect TMA production or gut bacteria composition |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janeiro, M.H.; Ramírez, M.J.; Milagro, F.I.; Martínez, J.A.; Solas, M. Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target. Nutrients 2018, 10, 1398. https://doi.org/10.3390/nu10101398
Janeiro MH, Ramírez MJ, Milagro FI, Martínez JA, Solas M. Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target. Nutrients. 2018; 10(10):1398. https://doi.org/10.3390/nu10101398
Chicago/Turabian StyleJaneiro, Manuel H., María J. Ramírez, Fermin I. Milagro, J. Alfredo Martínez, and Maite Solas. 2018. "Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target" Nutrients 10, no. 10: 1398. https://doi.org/10.3390/nu10101398
APA StyleJaneiro, M. H., Ramírez, M. J., Milagro, F. I., Martínez, J. A., & Solas, M. (2018). Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target. Nutrients, 10(10), 1398. https://doi.org/10.3390/nu10101398