
Alpine Forest Drought Monitoring in South Tyrol: PCA Based Synergy between scPDSI Data and MODIS Derived NDVI and NDII7 Time Series

Abstract
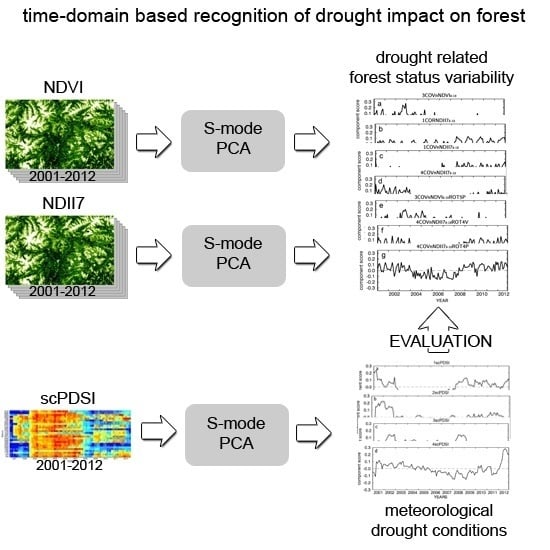
:
1. Introduction
2. Materials and Methods
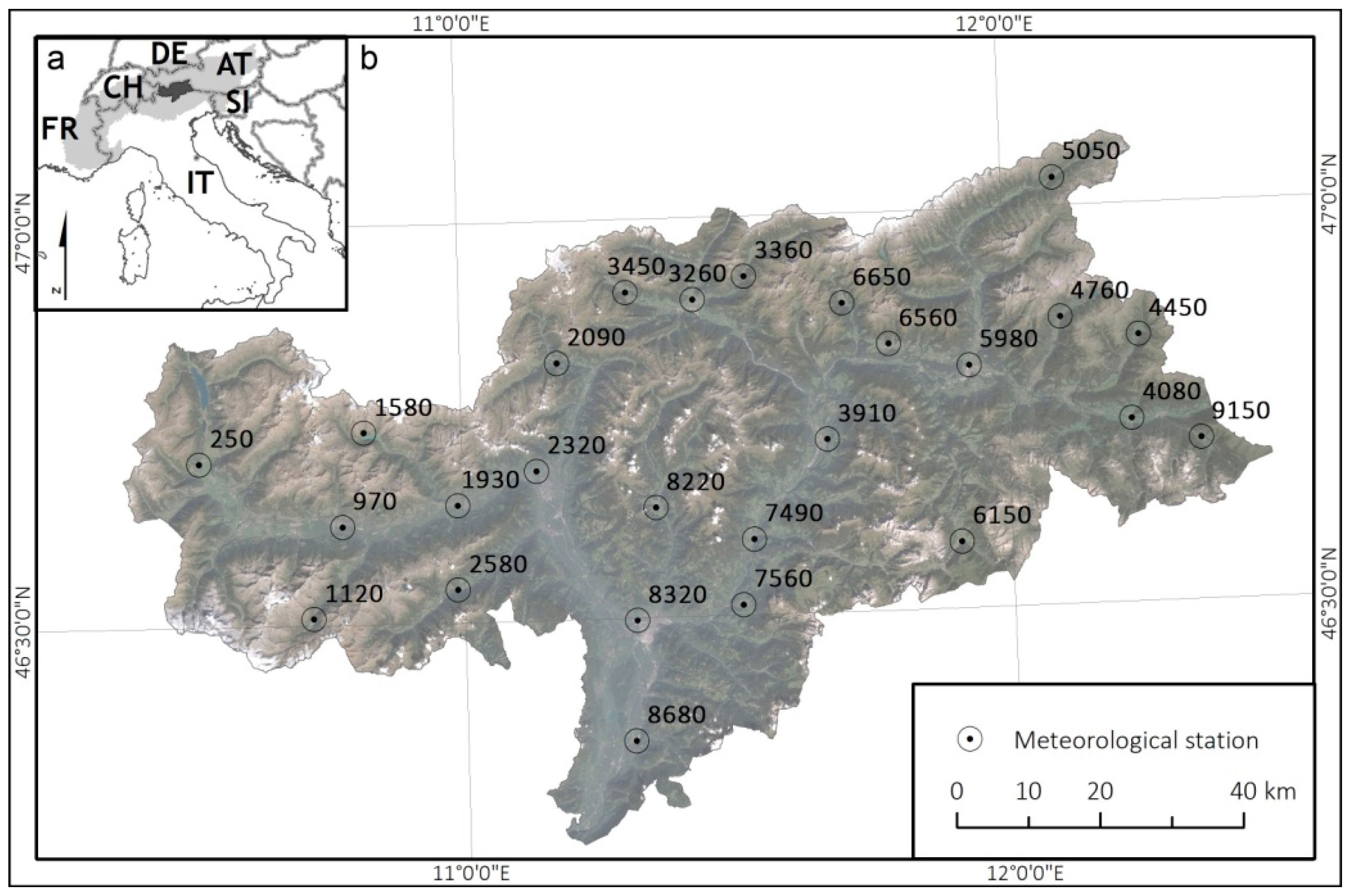
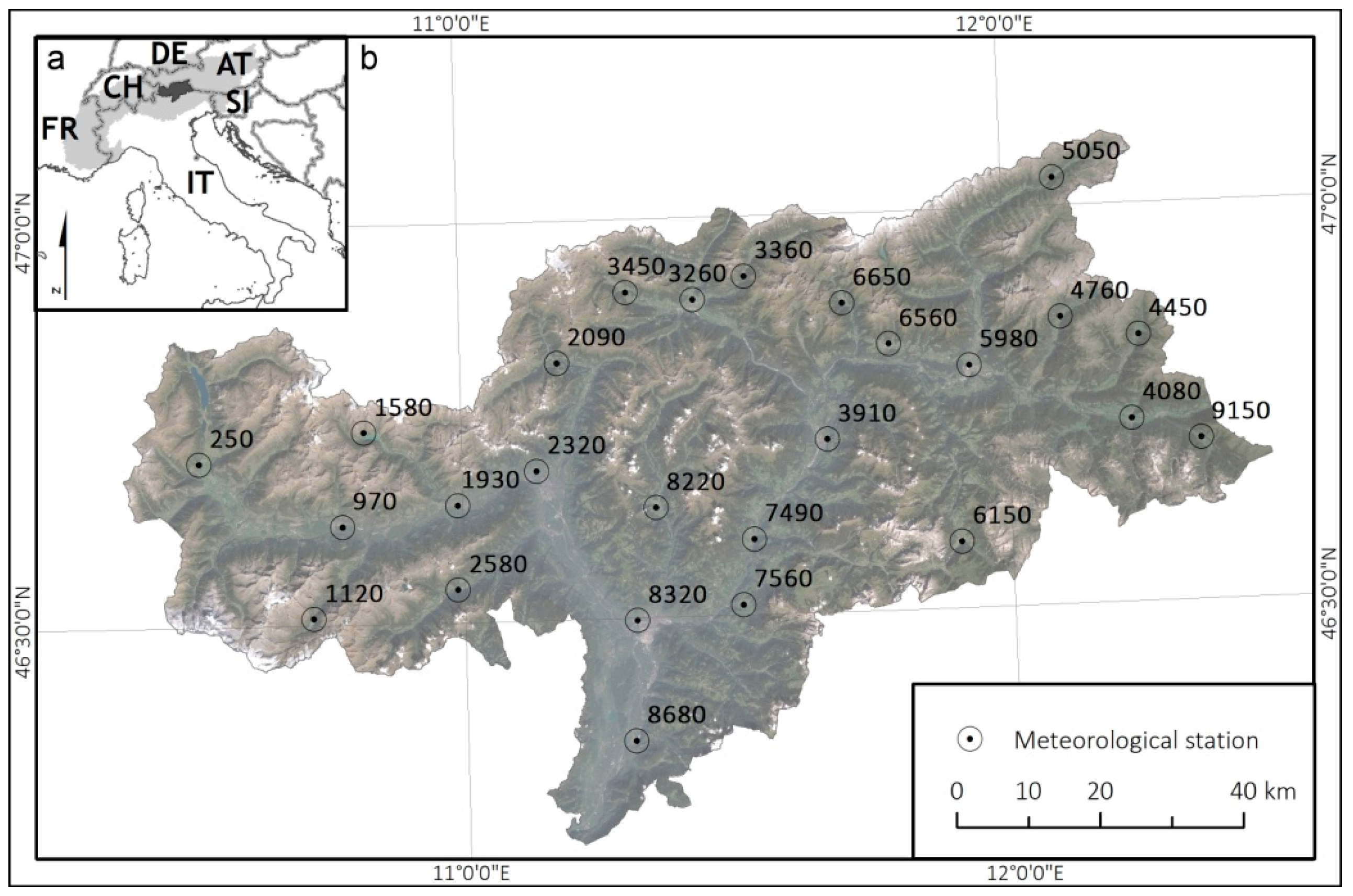
2.1. Study Area
2.2. Data
2.2.1. Meteorological Data and scPDSI
2.2.2. MODIS Derived Time Series of Vegetation Indices
2.2.3. Ancillary Data—Forest Mask
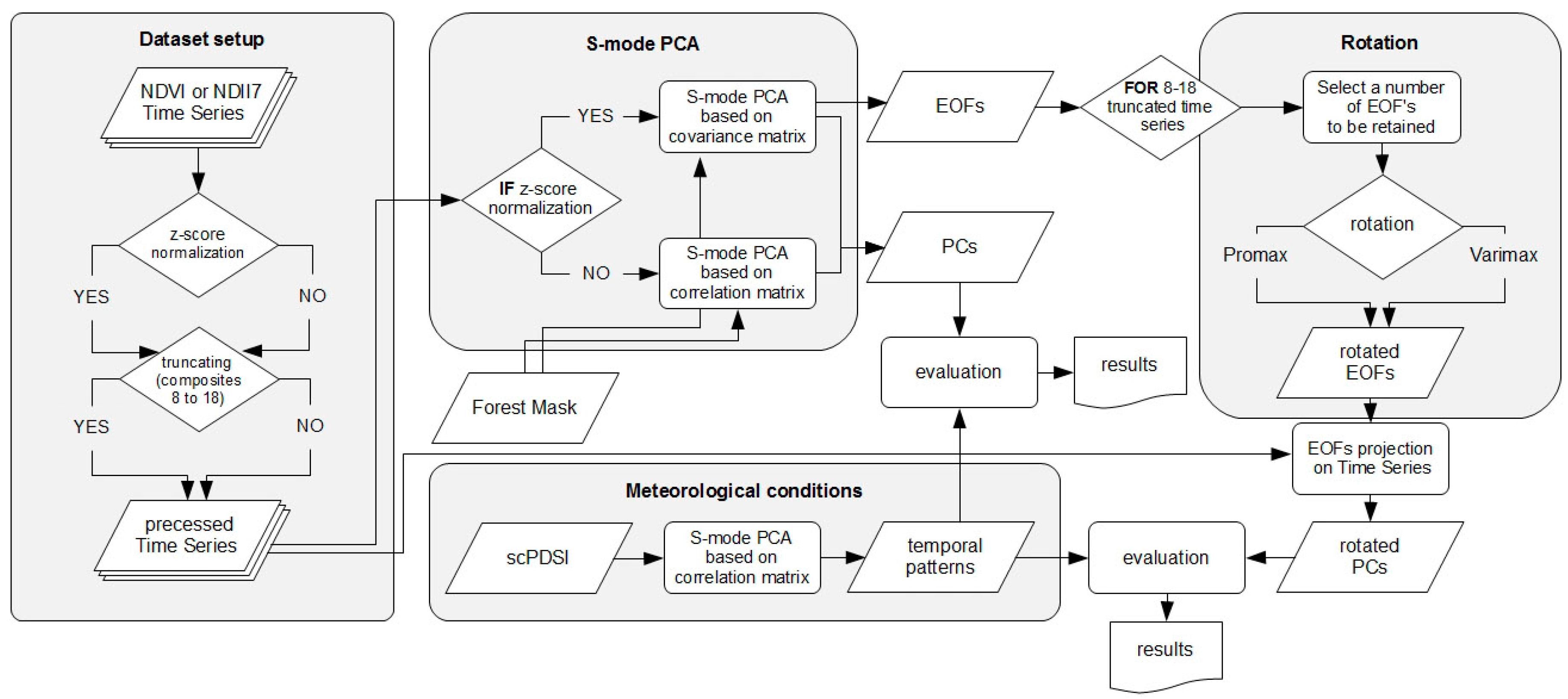
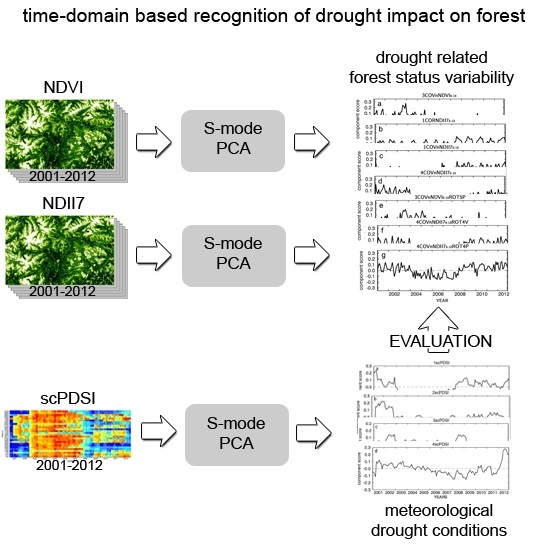
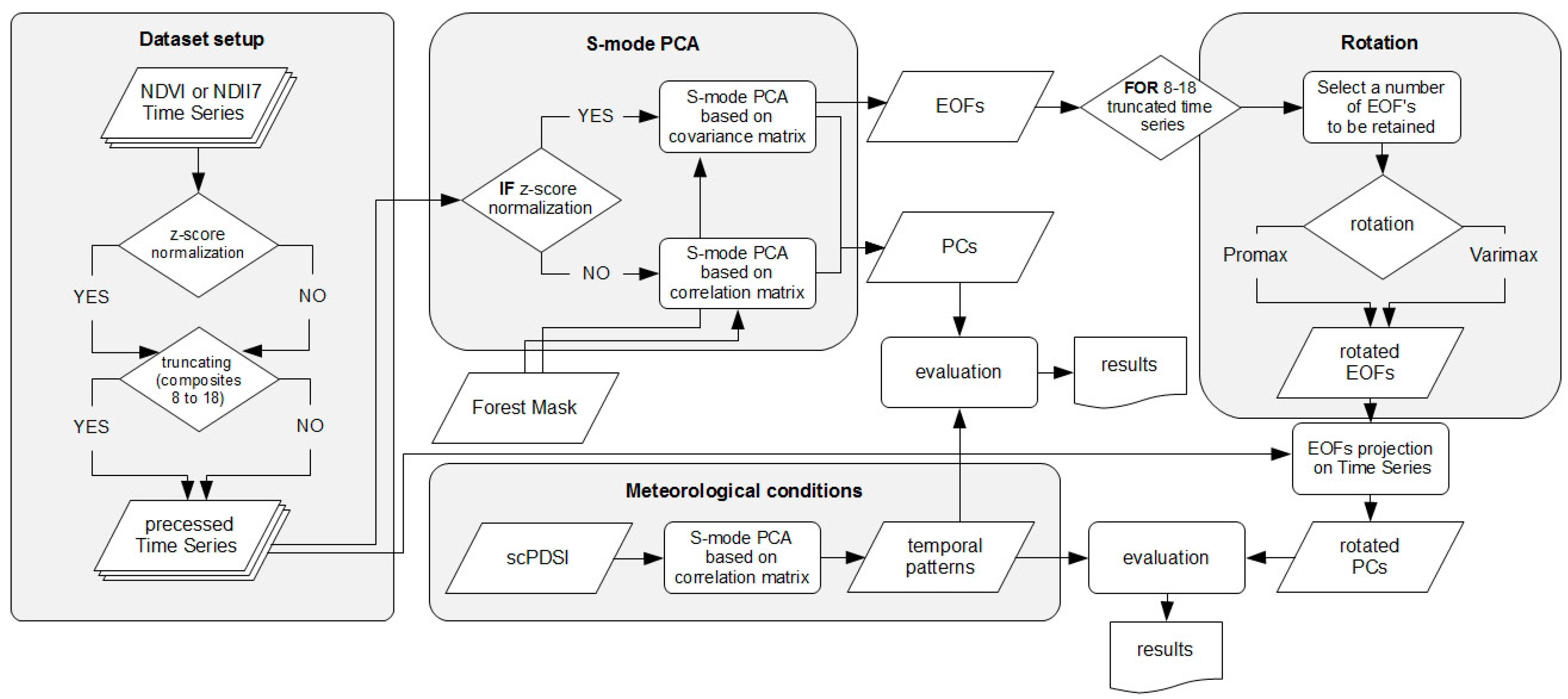
2.3. PCA Analyses Design
2.3.1. scPDSI Based Meteorological Variability
2.3.2. MODIS Derived NDVI and NDII7 Time Series
- NDVI and NDII7 time series, where the former relates to the photosynthetic activity, while the latter approximates foliage water content;
- removal of seasonality through a per-pixel z-score normalizationwhere Xij denotes a j-th composite () of an i-th year () of the complete 2001–2012 time series, µj-com stands for an average value for all j-th composites across all the years, and σj-com is a standard deviation for all the j-th composite in the time series;
- two different lengths of NDVI and NDII7 time series (applied to the original and z-score normalized datasets alike) where beside the complete MODIS time series comprising all 23 annual composites (1–23), a vegetation season time series focusing on a period between end of April and mid-October (corresponding with 8th to 18th MODIS annual composites (8–18)) were also exploited as time series restricted to vegetation season allow excluding dormancy state signal and potential impact of snow cover; and
3. Results
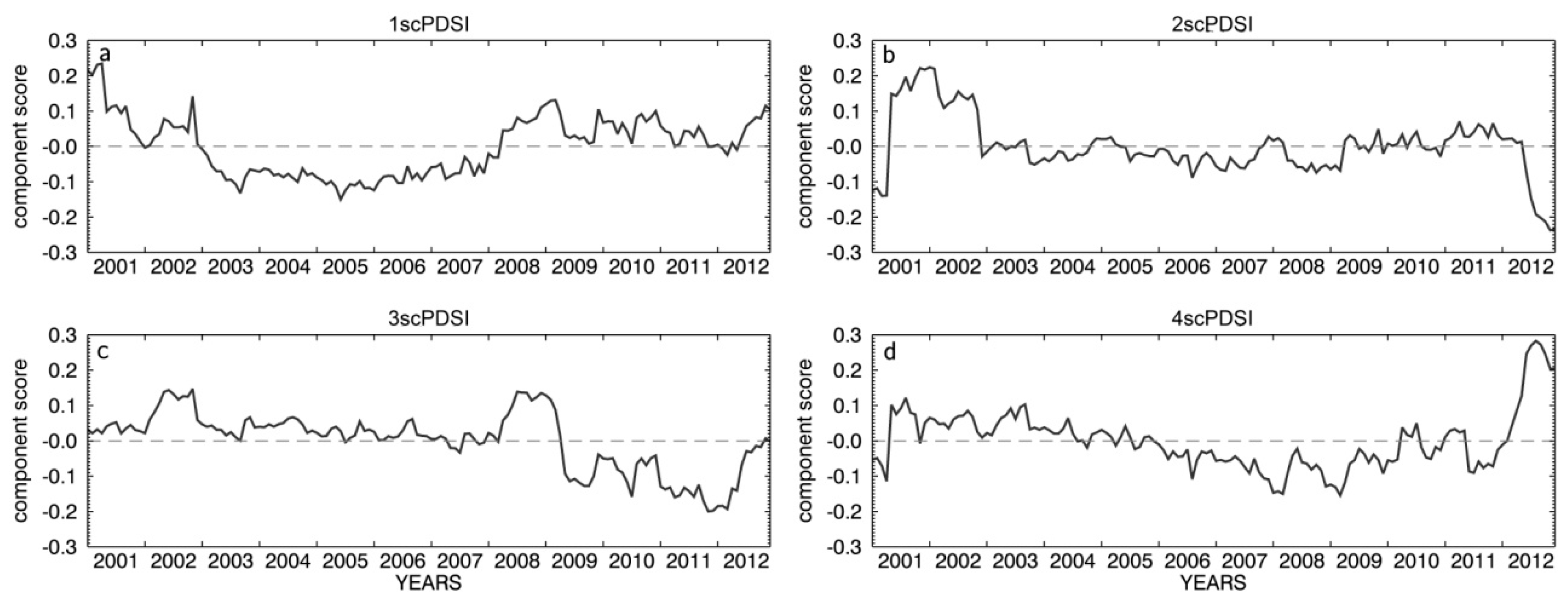
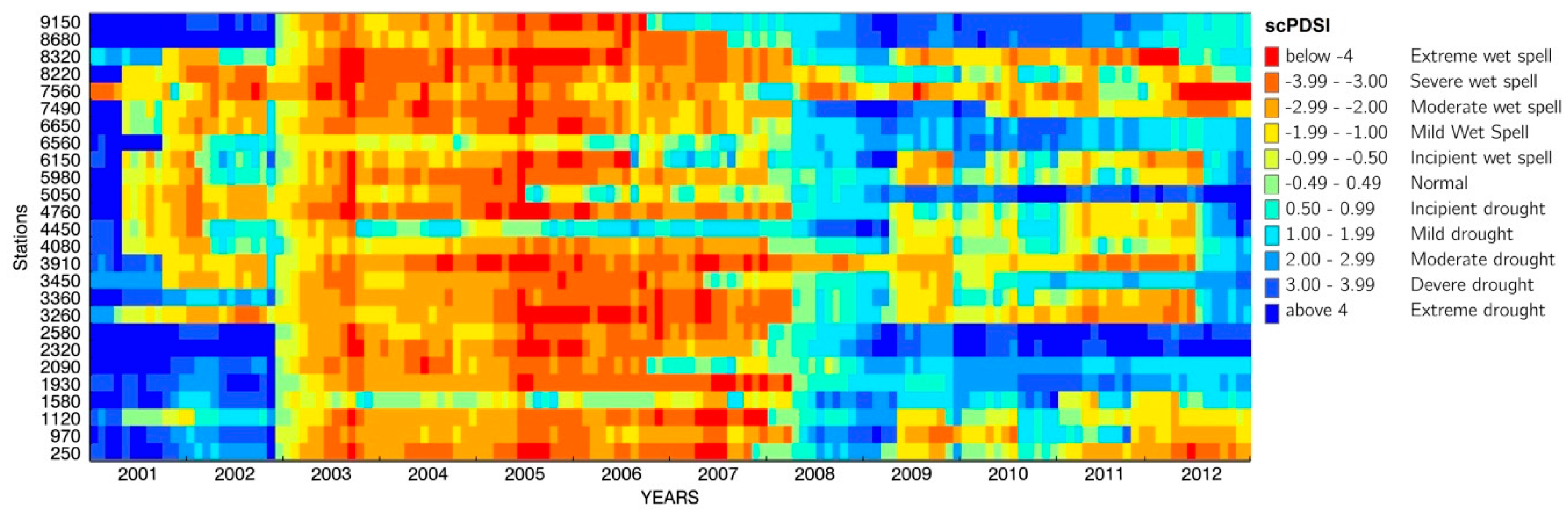
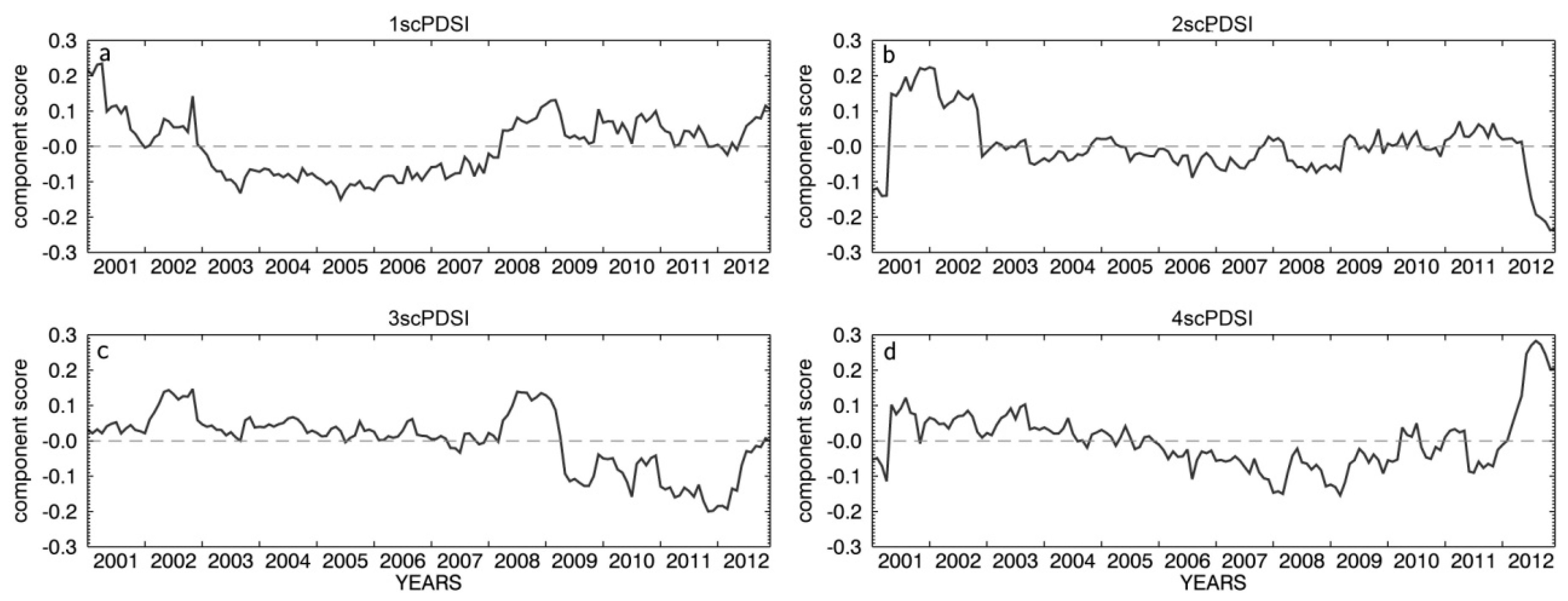
3.1. Meteorological Conditions in South Tyrol
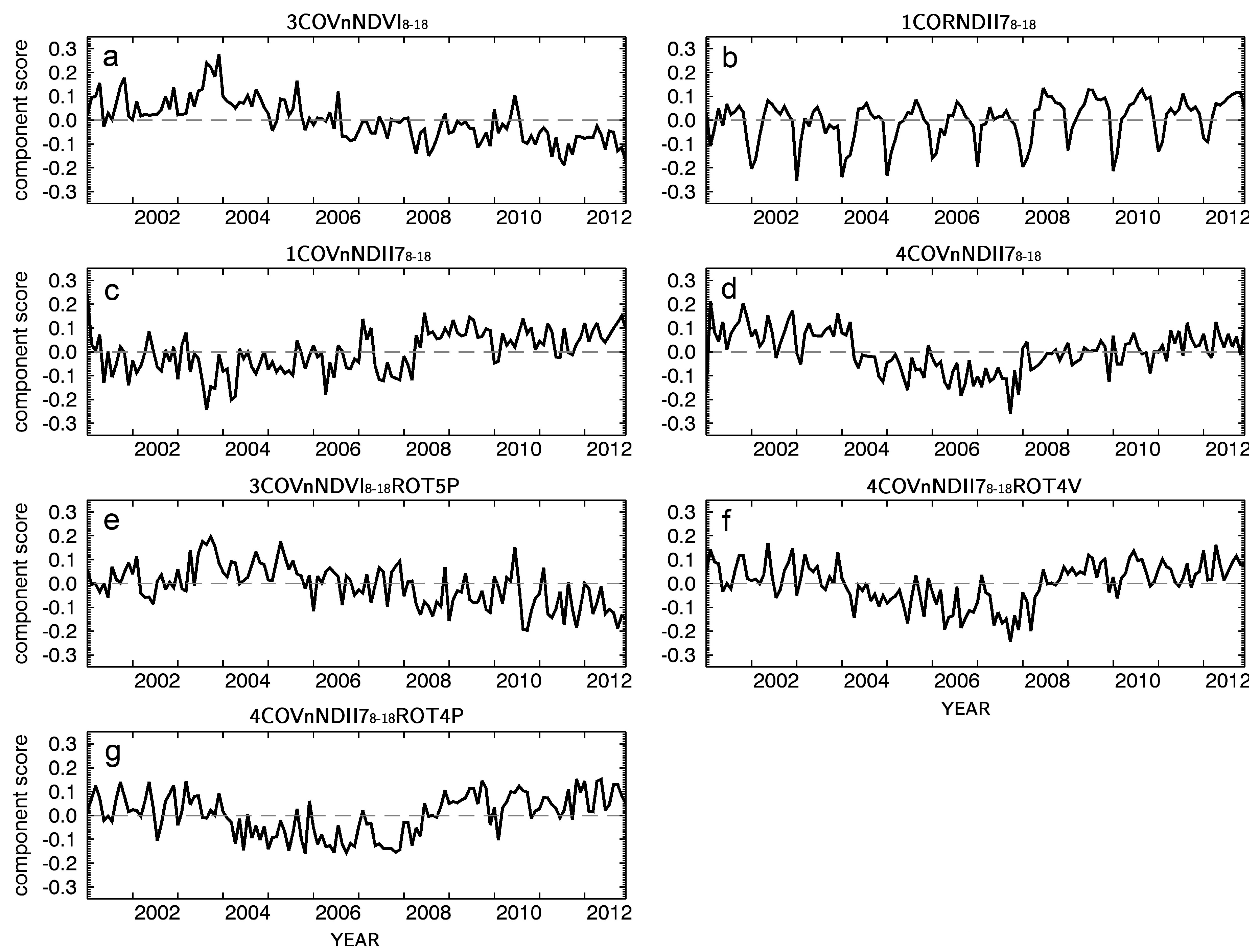
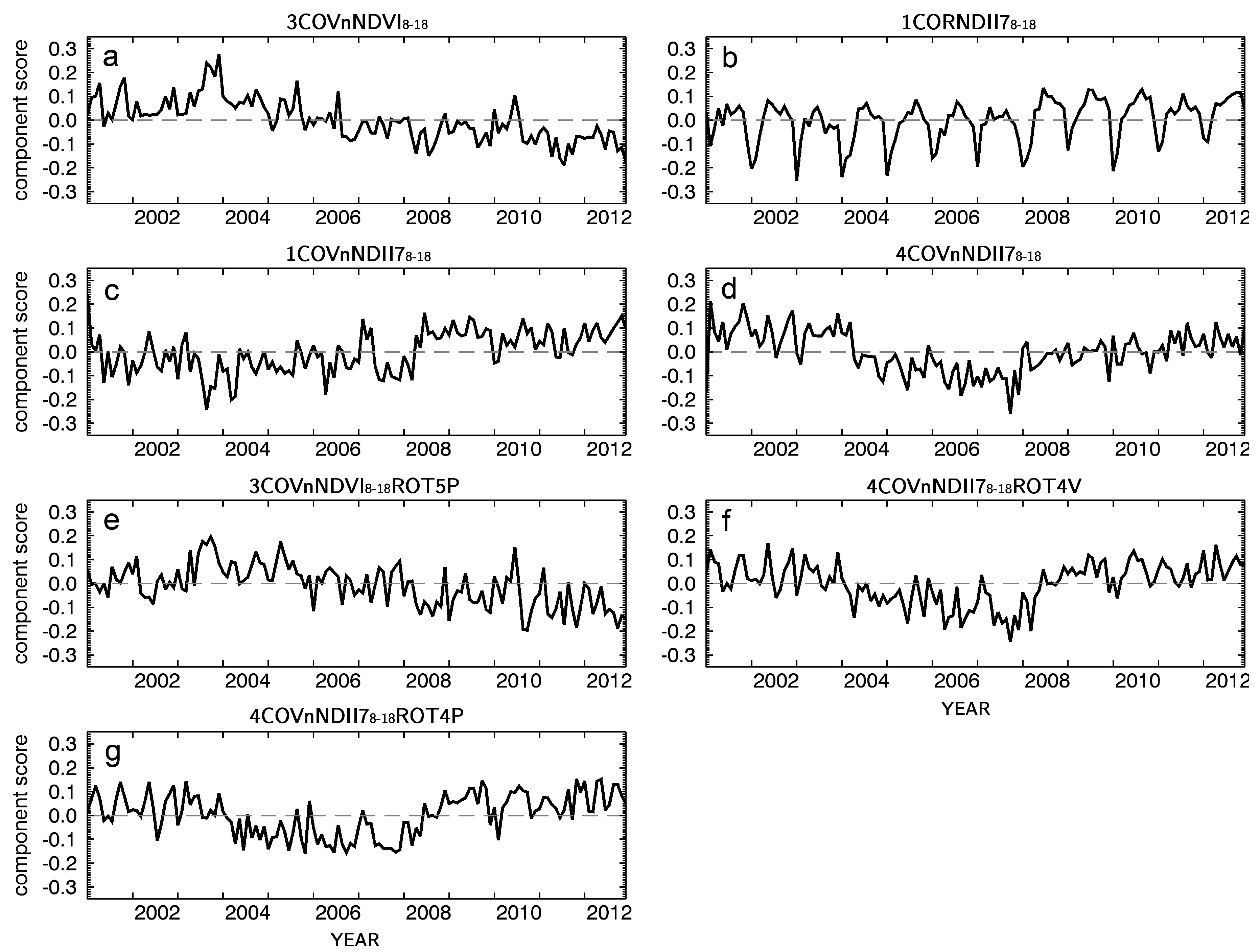
3.2. Forest Photosynthetic Activity Captured by PCA of MODIS NDVI Time Series
3.3. Forest Foliage Water Content Indicated by PCA of Vegetation Season MODIS NDII7 Time Series
3.4. Loadings Rotation
3.5. Comparison of Identified Potential Forest Responses to Meteorological Drought Conditions
4. Discussion
4.1. Drought Conditions in South Tyrol
4.2. Evaluation of Multiple S-mode PCA Decomposition Approaches and Data Setups
4.3. Forest Drought Response Identified through PCA Decomposition of MODIS NDVI and NDII7 Time Series
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| NDII7 | Normalized Difference Infrared Index based on MODIS band 7 |
| NDVI | Normalized Difference Vegetation Index |
| PC | Principal component |
| PCA | Principal Component Analysis |
| scPDSI | self-calibrated Palmer Drought Severity Index |
Appendix A
- Kaiser’s stopping rule [101], which proposes to rotate all loadings with eigenvalue ≥1.
- Cattell’s scree test [68], in which the selection is based on a visual interpretation of the eigenvalues plot and identification of a transition point between incline and leveled line. Because the transition point belongs to the leveled part, only loadings of a lower order than the transition point are rotated. Cattell’s scree test can be recognized as simplified graphical solution of the N rule [44].
- A priori criterion, where a number of rotated factors is set beforehand.
- Non-trivial factors approach, in which only these loadings are rotated that have at least three variables loadings above a certain threshold (customary 0.3).
- Percent of cumulative variance criterion, in which rotated are these foremost loadings that eigenvalues sum up to a predefined value.
References
- Mishra, A.K.; Singh, V.P. A review of drought concepts. J. Hydrol. 2010, 391, 202–216. [Google Scholar] [CrossRef]
- European Environment Agency (EEA). Europe’s Ecological Backbone: Recognising the True Value of Our Mountains; European Environment Agency (EEA): Copenhagen, Denmark, 2010. [Google Scholar]
- European Environment Agency (EEA). Climate Change, Impacts and Vulnerability in Europe 2012; European Environment Agency (EEA): Copenhagen, Denmark, 2012. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Dai, A. Drought under global warming: A review. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef]
- European Environment Agency (EEA). Regional Climate Change and Adaptation; European Environment Agency (EEA): Copenhagen, Denmark, 2009. [Google Scholar]
- Philipona, R.; Behrens, K.; Ruckstuhl, C. How declining aerosols and rising greenhouse gases forced rapid warming in Europe since 1980s. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Dai, A. Characteristics and trends in various forms of the palmer drought severity index during 1900–2008. Clim. Dyn. 2011, 116. [Google Scholar] [CrossRef]
- Lloyd-Hughes, B.; Saunders, M.A. A drought climatology for Europe. Int. J. Climatol. 2002, 22, 1571–1592. [Google Scholar] [CrossRef]
- Schär, C.; Vidale, P.L.; Lüthi, D.; Frei, C.; Häberli, C.; LiNiger, M.A.; Appenzeller, C. The role of increasing temperature variability in European summer heat waves. Nature 2004, 427, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Migliavacca, M.; Cremonese, E.; Colombo, R.; Busetto, L.; Galvagno, M.; Ganis, L.; Meroni, M.; Pari, E.; Rossini, M.; Siniscalco, C.; et al. European larch phenology in the Alps: Can we grasp the role of ecological factors by combining field observations and inverse modelling? Int. J. Biometeorol. 2008, 52, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- Wilhite, D.A.; Glantz, M.H. Understanding: The drought phenomenon: The role of definitions. Water Int. 1985, 10, 111–120. [Google Scholar] [CrossRef]
- He, B.; Cui, X.; Wang, H.; Chen, A. Drought: The most important physical stress of terrestrial ecosystems. Acta Ecol. Sin. 2014, 34, 179–183. [Google Scholar] [CrossRef]
- Ma, Z.; Peng, C.; Zhu, Q.; Chen, H.; Yu, G.; Li, W.; Zhou, X.; Wang, W.; Zhang, W. Regional drought-induced reduction in the biomass carbon sink of Canada’s boreal forests. Proc. Natl. Acad. Sci. USA 2012, 109, 2423–2427. [Google Scholar] [CrossRef] [PubMed]
- Schoene, D.H.F.; Bernier, P.Y. Adapting forestry and forests to climate change: A challenge to change the paradigm. For. Policy Econ. 2012, 24, 12–19. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Buffo, E.; Larsson, S. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob. Chang. Biol. 2006, 12, 662–671. [Google Scholar] [CrossRef]
- Rigling, A.; Bigler, C.; Eilmann, B.; Feldmeyer-Christe, E.; Gimmi, U.; Ginzler, C.; Graf, U.; Mayer, P.; Vacchiano, G.; Weber, P.; et al. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob. Chang. Biol. 2013, 19, 229–240. [Google Scholar] [CrossRef] [PubMed]
- The European Commission. Report of the Standing Forestry Committee Ad Hoc Working Group III on “Climate Change and Forestry”; The European Commission: Brussels, Belgium; Luxembourg, 2010. [Google Scholar]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Scherrer, D.; Bader, M.K.-F.; Körner, C. Drought-sensitivity ranking of deciduous tree species based on thermal imaging of forest canopies. Agric. For. Meteorol. 2011, 151, 1632–1640. [Google Scholar] [CrossRef]
- Etzold, S.; Waldner, P.; Thimonier, A.; Schmitt, M.; Dobbertin, M. Tree growth in Swiss forests between 1995 and 2010 in relation to climate and stand conditions: Recent disturbances matter. For. Ecol. Manag. 2014, 311, 41–55. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Jandl, R.; Hanewinkel, M.; Kunstler, G.; Kölling, C.; Gasparini, P.; Breznikar, A.; Meier, E.S.; Normand, S.; Ulmer, U.; et al. Potential future ranges of tree species in the Alps. In Management Strategies to Adapt Alpine Space Forests to Climate Change Risks; Cerbu, G.A., Hanewinkel, M., Gerosa, G., Jandl, R., Eds.; InTech: New York, NY, USA, 2013; pp. 37–48. [Google Scholar]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Rigling, A.; Waldner, P.O.; Forster, T.; Bräker, O.U.; Pouttu, A. Ecological interpretation of tree-ring width and intraannual density fluctuations in Pinus sylvestris on dry sites in the central Alps and Siberia. Can. J. For. Res. 2001, 31, 18–31. [Google Scholar] [CrossRef]
- Weber, P.; Bugmann, H.; Rigling, A. Radial growth responses to drought of Pinus sylvestris and Quercus pubescens in an inner-Alpine dry valley. J. Veg. Sci. 2007, 18, 777–792. [Google Scholar] [CrossRef]
- Pichler, P.; Oberhuber, W. Radial growth response of coniferous forest trees in an inner Alpine environment to heat-wave in 2003. For. Ecol. Manag. 2007, 242, 688–699. [Google Scholar] [CrossRef]
- Castagneri, D.; Nola, P.; Motta, R.; Carrer, M. Summer climate variability over the last 250 years differently affected tree species radial growth in a mesic Fagus–Abies–Picea old-growth forest. For. Ecol. Manag. 2014, 320, 21–29. [Google Scholar] [CrossRef]
- Eilmann, B.; Weber, P.; Rigling, A.; Eckstein, D. Growth reactions of Pinus sylvestris L. and Quercus pubescens Willd. to drought years at a xeric site in Valais, Switzerland. Dendrochronologia 2006, 23, 121–132. [Google Scholar] [CrossRef]
- Lévesque, M.; Rigling, A.; Bugmann, H.; Weber, P.; Brang, P. Growth response of five co-occurring conifers to drought across a wide climatic gradient in Central Europe. Agric. For. Meteorol. 2014, 197, 1–12. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photograpic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Hardisky, M.A.; Klemas, V.; Smart, R. The influence of soil salinity, growth form, and leaf moisture on the spectral radiance of Spartan alterniflora canopies. Am. Soc. Photogramm. 1983, 49, 77–83. [Google Scholar]
- Van Wagtendonk, J.W.; Root, R.R.; Key, C.H. Comparison of AVIRIS and Landsat ETM+ detection capabilities for burn severity. Remote Sens. Environ. 2004, 92, 397–408. [Google Scholar] [CrossRef]
- Rahimzadeh, B.P.; Shimizu, Y.; Hosoi, F.; Omasa, K. MODIS vegetation and water indices for drought assessment in semi-arid ecosystems of Iran. J. Agric. Meteorol. 2009, 65, 349–355. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.; Gao, X.; Ferreira, L. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Gao, B. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Horion, S.; Cornet, Y.; Erpicum, M.; Tychon, B. Studying interactions between climate variability and vegetation dynamic using a phenology based approach. Int. J. Appl. Earth Obs. Geoinf. 2013, 20, 20–32. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P. Temporal responses of NDVI to precipitation and temperature in the central Great Plains, USA. Int. J. Remote Sens. 2003, 24, 2345–2364. [Google Scholar] [CrossRef]
- Ji, L.; Peters, A.J. Assessing vegetation response to drought in the northern Great Plains using vegetation and drought indices. Remote Sens. Environ. 2003, 87, 85–98. [Google Scholar] [CrossRef]
- Peters, A.J.; Walter-Shea, E.A.; Ji, L.; Vina, A.; Hayes, M.; Svoboda, M.D. Drought monitoring with NDVI-based standardized vegetation index. Photogramm. Eng. Remote Sens. 2002, 68, 71–75. [Google Scholar]
- Gouveia, C.; Trigo, R.M.; DaCamara, C.C. Drought and vegetation stress monitoring in Portugal using satellite data. Nat. Hazards Earth Syst. Sci. 2009, 9, 185–195. [Google Scholar] [CrossRef]
- Loveland, T.R.; Dwyer, J.L. Landsat: Building a strong future. Remote Sens. Environ. 2012, 122, 22–29. [Google Scholar] [CrossRef]
- Venegas, S.A. Statistical Methods for Signal Detection in Climate; Danish Center for Earth System Science (DCESS): Copenhagen, Denmark, 2001. [Google Scholar]
- Preisendorfer, R.W. Principal Component Analysis in Meteorology and Oceanography; Elsevier: Amsterdam, The Netherlands, 1988. [Google Scholar]
- Chen, C.H.; Peter Ho, P.-G. Statistical pattern recognition in remote sensing. Pattern Recognit. 2008, 41, 2731–2741. [Google Scholar] [CrossRef]
- Neeti, N.; Ronald, E.J. Novel approaches in extended principal component analysis to compare spatio-temporal patterns among multiple image time series. Remote Sens. Environ. 2014, 148, 84–96. [Google Scholar] [CrossRef]
- Small, C. Spatiotemporal dimensionality and Time-Space characterization of multitemporal imagery. Remote Sens. Environ. 2012, 124, 793–809. [Google Scholar] [CrossRef]
- Ivits, E.; Cherlet, M.; Mehl, W.; Sommer, S. Ecosystem functional units characterized by satellite observed phenology and productivity gradients: A case study for Europe. Ecol. Indic. 2013, 27, 17–28. [Google Scholar] [CrossRef]
- Lasaponara, R. On the use of principal component analysis (PCA) for evaluating interannual vegetation anomalies from SPOT/VEGETATION NDVI temporal series. Ecol. Model. 2006, 194, 429–434. [Google Scholar] [CrossRef]
- Eastman, R.J.; Fulk, M. Long sequence time series evaluation using standardized principal components. Photogramm. Eng. Remote Sens. 1993, 59, 991–996. [Google Scholar]
- MaChado-MaChado, E.A.; Neeti, N.; Eastman, J.R.; Chen, H. Implications of space-time orientation for principal components analysis of earth observation image time series. Earth Sci. Inform. 2011, 4, 117–124. [Google Scholar] [CrossRef]
- Richman, M.B. Rotation of principal components. J. Climatol. 1986, 6, 293–335. [Google Scholar] [CrossRef]
- Kaiser, H. The varimax criterion for analytic rotation in factor analysis. Psychometrika 1958, 23, 187–200. [Google Scholar] [CrossRef]
- Hendrickson, A.E.; White, P.O. PROMAX: A quick method for rotation to oblique simple structure. Br. J. Stat. Psychol. 1964, 17, 65–70. [Google Scholar] [CrossRef]
- Minerbi, S.; Cescatti, A.; Cherubini, P.; Hellrigl, K.; Markart, G.; Saurer, M.; Mutinelli, C. Scots pine dieback in the Isarco Valley due to severe drought in the summer of 2003. For. Obs. 2006, 2, 89–144. [Google Scholar]
- Auer, I.; Reinhard, B.; Jurkovic, A.; Lipa, W.; Orlik, A.; Potzmann, R.; Sch, W.; Ungersb, M.; Matulla, C.; Briffa, K.; et al. HISTALP—Historical instrumental climatological surface time series of the Greater Alpine Region. Int. J. Climatol. 2007, 27, 17–46. [Google Scholar] [CrossRef]
- Wells, N.; Steve, G.; Hayes, M.J. A self-calibrating palmer drought severity index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- Büntgen, U.; Trouet, V.; Frank, D.; Leuschner, H.H.; Friedrichs, D.; Luterbacher, J.; Esper, J. Tree-ring indicators of German summer drought over the last millennium. Quat. Sci. Rev. 2010, 29, 1005–1016. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought matters—Declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in North-Eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Gillner, S.; Vogt, J.; Roloff, A. Climatic response and impacts of drought on oaks at urban and forest sites. Urban For. Urban Green. 2013, 12, 597–605. [Google Scholar] [CrossRef]
- Provincial Statistics Institute of Autonomous Province of South Tyrol. South Tyrol in Figures; Provincial Statistics Institute of Autonomous Province of South Tyrol: Bolzano, Italy, 2012. [Google Scholar]
- Wells, N. PDSI Users Manual; Version 2.0; National Agricultural Decision Support System, University of Nebraska-Lincoln: Lincoln, NE, USA, 2003. [Google Scholar]
- Colditz, R.R.; Conrad, C.; Wehrmann, T.; Schmidt, M.; Dech, S. TiSeG: A flexible software tool for time-series generation of MODIS data utilizing the quality assessment science data set. IEEE Trans. Geosci. Remote Sens. 2008, 46, 3296–3308. [Google Scholar] [CrossRef]
- Udelhoven, T. TimeStats: A software tool for the retrieval of temporal patterns from global satellite archives. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2011, 4, 310–317. [Google Scholar] [CrossRef]
- Wang, D.; Morton, D.; Masek, J.; Wu, A.; Nagol, J.; Xiong, X.; Levy, R.; Vermote, E.; Wolfe, R. Impact of sensor degradation on the MODIS NDVI time series. Remote Sens. Environ. 2012, 119, 55–61. [Google Scholar] [CrossRef]
- Geoland2. Technical Note on HR Forest Layer Product Specification, Issue 1.4, Publication of the FP7 geoland2 project. 2012.
- European Environment Agency (EEA). Corine Land Cover 2006; European Environment Agency: Kobenhavn, Denmark, 2006. [Google Scholar]
- Cattell, R.B. The scree test for the number of factors. Multivar. Behav. Res. 1966, 1, 245–276. [Google Scholar] [CrossRef] [PubMed]
- Rebetez, M.; Mayer, H.; Dupont, O. Heat and drought 2003 in Europe: A climate synthesis. Ann. For. 2006, 63, 569–577. [Google Scholar] [CrossRef]
- Spinoni, J.; Naumann, G.; Vogt, J.V.; Barbosa, P. The biggest drought events in Europe from 1950 to 2012. J. Hydrol. Reg. Stud. 2015, 3, 1–16. [Google Scholar] [CrossRef]
- García-Herrera, R.; Hernández, E.; Barriopedro, D.; Paredes, D.; Trigo, R.M.; Trigo, I.F.; Mendes, M.A. The outstanding 2004/05 drought in the Iberian Peninsula: Associated atmospheric circulation. J. Hydrometeorol. 2007, 8, 483–498. [Google Scholar] [CrossRef]
- Rebetez, M.; Dupont, O.; Giroud, M. An analysis of the July 2006 heatwave extent in Europe compared to the record year of 2003. Theor. Appl. Climatol. 2008, 95, 1–7. [Google Scholar] [CrossRef]
- Hydrographic Office of the Autonomous Province of Bolzano-Südtirol. Climareport Südtirol-Alto Adige; Hydrographic Office of the Autonomous Province of Bolzano-Südtirol: Bolzano, Italy, 2006. [Google Scholar]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef] [PubMed]
- August, D.; Geiger, M. Drought in the Mediterranean. Recent Developments; World Wide Fund for Nature: Berlin, Germany, 2008. [Google Scholar]
- Ivits, E.; Horion, S.; Fensholt, R.; Cherlet, M. Drought footprint on European ecosystems between 1999 and 2010 assessed by remotely sensed vegetation phenology and productivity. Glob. Chang. Biol. 2014, 20, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Auer, I.; Bühm, R.; Jurković, A.; Orlik, A.; Potzmann, R.; Schöner, W.; Ungersböck, M.; Brunetti, M.; Nanni, T.; Maugeri, M.; et al. A new instrumental precipitation dataset for the greater alpine region for the period 1800–2002. Int. J. Climatol. 2005, 25, 139–166. [Google Scholar] [CrossRef]
- Toothill, J. Central European Flooding, August 2002. An EQECAT Technical Report; ABS Consulting: Vancouver, BC, Canada, 2002. [Google Scholar]
- Van der Schrier, G.; Barichivich, J.; Briffa, K.R.; Jones, P.D. A scPDSI-based global data set of dry and wet spells for 1901–2009. J. Geophys. Res. Atmos. 2013, 118, 4025–4048. [Google Scholar] [CrossRef]
- Van der Schrier, G.; Briffa, K.R.; Jones, P.D.; Osborn, T.J. Summer moisture variability across Europe. J. Clim. 2006, 19, 2818–2834. [Google Scholar] [CrossRef]
- Van der Schrier, G.; Efthymiadis, D.; Briffa, K.R.; Jones, P.D. European Alpine moisture variability for 1800–2003. Int. J. Climatol. 2007, 27, 415–427. [Google Scholar] [CrossRef]
- DeVries, B.; Decuyper, M.; Verbesselt, J.; Zeileis, A.; Herold, M.; Joseph, S. Tracking disturbance-regrowth dynamics in tropical forests using structural change detection and Landsat time series. Remote Sens. Environ. 2015, 169, 320–334. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Gebetsroither, E.; Züger, J.; Schmatz, D.; Psomas, A. Future climate of the European Alps. In Management Strategies to Adapt Alpine Space Forests to Climate Change Risks; Zimmermann, N.E., Ed.; InTech: New York, NY, USA, 2013; pp. 27–36. [Google Scholar]
- Körner, C. Alpine Plant Life; Springer: Berlin, Germany; Heidelberg, Germany, 2003. [Google Scholar]
- Courbaud, B.; Kunstler, G.; Morin, X. What is the future of the ecosystem services of the Alpine forest against a backdrop of climate change? J. Alp. Res. 2010, 98. [Google Scholar] [CrossRef]
- Bussotti, F.; Pollastrini, M.; Holland, V.; Brüggemann, W. Functional traits and adaptive capacity of European forests to climate change. Environ. Exp. Bot. 2014, 111, 91–113. [Google Scholar] [CrossRef]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Vittoz, P.; Rulence, B.; Largey, T.; Freléchoux, F. Effects of climate and land-use change on the establishment and growth of Cembran Pine (Pinus cembra L.) over the altitudinal treeline ecotone in the Central Swiss Alps. Arct. Antarct. Alp. Res. 2008, 40, 225–232. [Google Scholar] [CrossRef]
- Studer, S.; Appenzeller, C.; Defila, C. Inter-annual variability and decadal trends in alpine spring phenology: A multivariate analysis approach. Clim. Chang. 2005, 73, 395–414. [Google Scholar] [CrossRef]
- Theurillat, J.-P.; Guisan, A. Potential impact of climate change on vegetation in the European Alps: A review. Clim. Chang. 2001, 50, 77–109. [Google Scholar] [CrossRef]
- Studer, S.; Stöckli, R.; Appenzeller, C.; Vidale, P.L. A comparative study of satellite and ground-based phenology. Int. J. Biometeorol. 2007, 51, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Provincia Autonomica di Bolzano. Tipologie Forestali Dell’ Alto Adige Volume 1 Tipi Forestali, Regioni Forestali, Chiave dei Tipi Forestali; Provincia Autonomica di Bolzano: Bolzano, Italy, 2010. [Google Scholar]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Jolly, W.M.; Dobbertin, M.; Zimmermann, N.E.; Reichstein, M. Divergent vegetation growth responses to the 2003 heat wave in the Swiss Alps. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Vacchiano, G.; Garbarion, M.; Borgogno Mondino, E.; Renzo, M. Evidences of drought stress as a predisposing factor to Scots pine decline in Valle d’Aosta. Eur. J. For. Res. 2012, 131, 989–1000. [Google Scholar] [CrossRef]
- Chauchard, S.; Beilhe, F.; Denis, N.; Carcaillet, C. An increase in the upper tree-limit of silver fir (Abies alba Mill.) in the Alps since the mid-20th century: A land-use change phenomenon. For. Ecol. Manag. 2010, 259, 1406–1415. [Google Scholar] [CrossRef]
- Giuggiola, A.; Bugmann, H.; Zingg, A.; Dobbertin, M.; Rigling, A. Reduction of stand density increases drought resistance in xeric Scots pine forests. For. Ecol. Manag. 2013, 310, 827–835. [Google Scholar] [CrossRef]
- Bigler, C.; Bräker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an inciting mortality factor in Scots pine stands of the Valais, Switzerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef]
- Björnsson, H.; Venegas, S. A Manual for EOF and SVD Analyses of Climatic Data; McGill University: Montréal, QC, Canada, 1997. [Google Scholar]
- Brown, J. Choosing the right type of rotation in PCA and EFA. JALT Test. Eval. SIG Newsl. 2009, 13, 20–25. [Google Scholar]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Costello, A.B.; Osborne, J.W. Best practices in exploratory factor analysis: Four recommendations for getting the most from your analysis. Pract. Assess. Res. Eval. 2005, 10, 1–9. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | z-Score Data Normalization | Annual Time Window (MODIS Composites) | Dataset Short Name | EOF Matrix | Rotation (Retained Scores) |
|---|---|---|---|---|---|
| NDVI | no | full year (1–23) | NDVI1–23 | cor. | no |
| no | veg. season (8–18) | NDVI8–18 | cor. | no | |
| yes | full year (1–23) | nNDVI1–23 | cov. | no | |
| yes | veg. season (8–18) | nNDVI8–18 | cov. | yes (1–5) | |
| NDII7 | no | veg. season (8–18) | NDII78–18 | cor. | no |
| yes | veg. season (8–18) | nNDII78–18 | cov. | yes (1–4) |
| PCs | |||||||
|---|---|---|---|---|---|---|---|
| a | b | c | d | e | f | g | |
| 1scPDSI | −0.310 | 0.736 * | 0.717 * | 0.608 * | −0.590 * | 0.751 * | 0.772 * |
| 2scPDSI | 0.360 | −0.172 | −0.199 | 0.502 | 0.186 | 0.301 | 0.241 |
| 3scPDSI | 0.632 * | −0.576 * | −0.559 | −0.023 | 0.607 * | −0.331 | −0.397 |
| 4scPDSI | 0.257 | 0.030 | −0.010 | 0.583* | 0.029 | 0.466 | 0.400 |
| Index | PC Name | Score Order | EOF Matrix Rotation | Short Name of Original Dataset | Scores Retained for Rotation | Rotation Approach |
|---|---|---|---|---|---|---|
| a | 3COVnNDVI8–18 | 3 | cov. | nNDVI8–18 | - | - |
| b | 1CORNDII78–18 | 1 | cor. | NDII78–18 | - | - |
| c | 1COVnNDII78–18 | 1 | cov. | nNDII78–18 | - | - |
| d | 4COVnNDII78–18 | 4 | cov. | nNDII78–18 | - | - |
| e | 3COVnNDVI8–18ROT5P | 3 | cov. | nNDVI8–18 | (1–5) | Promax |
| f | 4COVnNDII78–18ROT4V | 4 | cov. | nNDII78–18 | (1–4) | Varimax |
| g | 4COVnNDII78–18ROT4V | 4 | cov. | nNDII78–18 | (1–4) | Promax |
| Index | PC Name | PCs | ||||||
|---|---|---|---|---|---|---|---|---|
| a | b | c | d | e | f | g | ||
| a | 3COVnNDVI8–18 | 1.000 * | ||||||
| b | 1CORNDII78–18 | −0.207 * | 1.000 * | |||||
| c | 1COVnNDII78–18 | −0.422 * | 0.487 * | 1.000 * | ||||
| d | 4COVnNDII78–18 | 0.267 * | 0.032 | 0.000 | 1.000 * | |||
| e | 3COVnNDVI8–18ROT5P | 0.714 * | −0.352 * | −0.674 * | −0.031 * | 1.000 * | ||
| f | 4COVnNDII78–18ROT4V | 0.320 * | 0.187 * | 0.339 * | 0.617 * | 0.025 | 1.000 * | |
| g | 4COVnNDII78–18ROT4V | 0.285 * | 0.226 * | 0.497 * | 0.659 * | −0.051 | 0.924 * | 1.000 * |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewińska, K.E.; Ivits, E.; Schardt, M.; Zebisch, M. Alpine Forest Drought Monitoring in South Tyrol: PCA Based Synergy between scPDSI Data and MODIS Derived NDVI and NDII7 Time Series. Remote Sens. 2016, 8, 639. https://doi.org/10.3390/rs8080639
Lewińska KE, Ivits E, Schardt M, Zebisch M. Alpine Forest Drought Monitoring in South Tyrol: PCA Based Synergy between scPDSI Data and MODIS Derived NDVI and NDII7 Time Series. Remote Sensing. 2016; 8(8):639. https://doi.org/10.3390/rs8080639
Chicago/Turabian StyleLewińska, Katarzyna Ewa, Eva Ivits, Mathias Schardt, and Marc Zebisch. 2016. "Alpine Forest Drought Monitoring in South Tyrol: PCA Based Synergy between scPDSI Data and MODIS Derived NDVI and NDII7 Time Series" Remote Sensing 8, no. 8: 639. https://doi.org/10.3390/rs8080639