
Estimation of Daily Maize Gross Primary Productivity by Considering Specific Leaf Nitrogen and Phenology via Machine Learning Methods


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Ground-Measured Data
2.2.1. Solar Shortwave Radiation (SSR), Air Temperature (Tair), and GPP Data
2.2.2. Specific Leaf Nitrogen (SLN)
2.3. Remotely Sensed NDVI
2.4. Relationship between Phenology and the Maximum Allowable SLN
2.5. Methodology
2.5.1. ML Methods
2.5.2. Input Variable Combinations
2.5.3. The Importance of SLN and NMP
- (1)
- NE1 for training and NE2 for testing: these two sites exhibit similar moisture levels, yet possess varying data quantities, and the larger data set is utilized to validate the smaller data set.
- (2)
- NE1 for training and NE3 for testing: the water conditions at the two sites differ, and the smaller data set is validated using the larger data set.
- (3)
- NE3 for training and NE2 for testing: due to the varying water conditions at the two sites, the smaller data set is employed to validate the larger data set.
2.5.4. Comparison of Different ML Methods
2.6. Evaluation Metrics
3. Results
3.1. Relationship between Phenology and the Maximum Allowable SLN
3.2. Comparison of Input Variable Combinations Based on RF
3.2.1. RF Model Calibration and Input Variable Importance
3.2.2. RF Performance in NE1, NE2, and NE3 Sites with Different Input Variable Combinations
3.2.3. RF Performance in RO1 Site While Trained in NE1, NE2, and NE3 Sites
3.3. Comparison of Different ML Model Performances
3.3.1. Comparison of Model Performance in NE1, NE2, and NE3 Sites
3.3.2. Comparison of Model Performances in RO1 Site
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Full Name | Units | Source |
|---|---|---|---|
| Tmean | Daily mean air temperature | °C | FLUXNET 2015 |
| Tmin | Daily minimum air temperature | °C | FLUXNET 2015 |
| Tmax | Daily maximum air temperature | °C | FLUXNET 2015 |
| SSR | Solar shortwave radiation | MJ·m−2·day−1 | FLUXNET 2015 |
| NDVI | Normalized difference vegetation index | - | MOD09GQ, MOD09Q1 |
| SLN | Specific leaf nitrogen | gN·m−2(leaf) | CSP of the University of Nebraska |
| NMP | Normalized maize phenology | - | Wang-Engel model [64] |
| Acronym | Full Name |
|---|---|
| RF | Random forest |
| SVM | Support vector machine |
| CNN | Convolutional neural network |
| ELM | Extreme learning machine |
| NSE | Nash efficiency coefficient (-) |
| RMSE | Root mean square efficiency (gC·m−2·day−1) |
| CV | Coefficient of variation (-) |
| URMSE | Unbiased root mean square efficiency (gC·m−2·day−1) |
References
- Wu, C.; Munger, J.W.; Niu, Z.; Kuang, D. Comparison of Multiple Models for Estimating Gross Primary Production Using MODIS and Eddy Covariance Data in Harvard Forest. Remote Sens. Environ. 2010, 114, 2925–2939. [Google Scholar] [CrossRef]
- Wang, M.; Wang, S.; Zhao, J.; Ju, W.; Hao, Z. Global Positive Gross Primary Productivity Extremes and Climate Contributions during 1982–2016. Sci. Total Environ. 2021, 774, 145703. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Gilabert, M.; Sánchez-Ruiz, S.; Moreno, Á. Annual Gross Primary Production from Vegetation Indices: A Theoretically Sound Approach. Remote Sens. 2017, 9, 193. [Google Scholar] [CrossRef]
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the Planet: 1. Geographic Distribution of Global Agricultural Lands in the Year 2000: GLOBAL AGRICULTURAL LANDS IN 2000. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef]
- Chen, T.; Van Der Werf, G.R.; Gobron, N.; Moors, E.J.; Dolman, A.J. Global Cropland Monthly Gross Primary Production in the Year 2000. Biogeosciences 2014, 11, 3871–3880. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B. Terrestrial Gross Carbon Dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Sims, D.; Rahman, A.; Cordova, V.; Elmasri, B.; Baldocchi, D.; Bolstad, P.; Flanagan, L.; Goldstein, A.; Hollinger, D.; Misson, L. A New Model of Gross Primary Productivity for North American Ecosystems Based Solely on the Enhanced Vegetation Index and Land Surface Temperature from MODIS. Remote Sens. Environ. 2008, 112, 1633–1646. [Google Scholar] [CrossRef]
- Keenan, T.F.; Davidson, E.; Moffat, A.M.; Munger, W.; Richardson, A.D. Using Model-Data Fusion to Interpret Past Trends, and Quantify Uncertainties in Future Projections, of Terrestrial Ecosystem Carbon Cycling. Glob. Chang. Biol. 2012, 18, 2555–2569. [Google Scholar] [CrossRef]
- Zhu, A.X.; Scott Mackay, D. Effects of Spatial Detail of Soil Information on Watershed Modeling. J. Hydrol. 2001, 248, 54–77. [Google Scholar] [CrossRef]
- Running, S.W.; Nemani, R.R.; Heinsch, F.A.; Zhao, M.; Reeves, M.; Hashimoto, H. A Continuous Satellite-Derived Measure of Global Terrestrial Primary Production. BioScience 2004, 54, 547. [Google Scholar] [CrossRef]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The Photochemical Reflectance Index: An Optical Indicator of Photosynthetic Radiation Use Efficiency across Species, Functional Types, and Nutrient Levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Suyker, A.E.; Verma, S.B. Gross Primary Production and Ecosystem Respiration of Irrigated and Rainfed Maize–Soybean Cropping Systems over 8 Years. Agric. For. Meteorol. 2012, 165, 12–24. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, Q.; Hollinger, D.; Aber, J.; Moore, B. Modeling Gross Primary Production of an Evergreen Needleleaf Forest Using Modis and Climate Data. Ecol. Appl. 2005, 15, 954–969. [Google Scholar] [CrossRef]
- Yuan, W.; Cai, W.; Nguy-Robertson, A.L.; Fang, H.; Suyker, A.E.; Chen, Y.; Dong, W.; Liu, S.; Zhang, H. Uncertainty in Simulating Gross Primary Production of Cropland Ecosystem from Satellite-Based Models. Agric. For. Meteorol. 2015, 207, 48–57. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the Satellite-Derived NDVI to Assess Ecological Responses to Environmental Change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Crane-Droesch, A. Machine Learning Methods for Crop Yield Prediction and Climate Change Impact Assessment in Agriculture. Environ. Res. Lett. 2018, 13, 114003. [Google Scholar] [CrossRef]
- Veenadhari, S.; Misra, B.; Singh, C. Machine Learning Approach for Forecasting Crop Yield Based on Climatic Parameters. In Proceedings of the 2014 International Conference on Computer Communication and Informatics, Coimbatore, India, 3–5 January 2014; IEEE: Piscataway, NJ, USA, 2014; pp. 1–5. [Google Scholar]
- Palanivel, K.; Surianarayanan, C. An Approach for Prediction of Crop Yield Using Machine Learning and Big Data Techniques. Int. J. Comput. Eng. Technol. 2019, 10, 110–118. [Google Scholar] [CrossRef]
- Schlenker, W.; Roberts, M.J. Nonlinear Effects of Weather on Corn Yields. Rev. Agric. Econ. 2006, 28, 391–398. [Google Scholar] [CrossRef]
- Khaki, S.; Wang, L. Crop Yield Prediction Using Deep Neural Networks. Front. Plant Sci. 2019, 10, 621. [Google Scholar] [CrossRef]
- Benos, L.; Tagarakis, A.C.; Dolias, G.; Berruto, R.; Kateris, D.; Bochtis, D. Machine Learning in Agriculture: A Comprehensive Updated Review. Sensors 2021, 21, 3758. [Google Scholar] [CrossRef] [PubMed]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random Forests for Classification in Ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Liang, S.; Yuan, W. Estimating Global Gross Primary Production from Sun-Induced Chlorophyll Fluorescence Data and Auxiliary Information Using Machine Learning Methods. Remote Sens. 2021, 13, 963. [Google Scholar] [CrossRef]
- Dou, X.; Yang, Y. Comprehensive Evaluation of Machine Learning Techniques for Estimating the Responses of Carbon Fluxes to Climatic Forces in Different Terrestrial Ecosystems. Atmosphere 2018, 9, 83. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, D.; Santra, G.H. Applications of Machine Learning Techniques in Agricultural Crop Production: A Review Paper. Indian J. Sci. Technol. 2016, 9, 1–14. [Google Scholar] [CrossRef]
- Prakash Sarkar, D.; Uma Shankar, B.; Ranjan Parida, B. Machine Learning Approach to Predict Terrestrial Gross Primary Productivity Using Topographical and Remote Sensing Data. Ecol. Inform. 2022, 70, 101697. [Google Scholar] [CrossRef]
- Zhu, X.-J.; Yu, G.-R.; Chen, Z.; Zhang, W.-K.; Han, L.; Wang, Q.-F.; Chen, S.-P.; Liu, S.-M.; Wang, H.-M.; Yan, J.-H.; et al. Mapping Chinese Annual Gross Primary Productivity with Eddy Covariance Measurements and Machine Learning. Sci. Total Environ. 2023, 857, 159390. [Google Scholar] [CrossRef]
- Gu, L.; Post, W.M.; Baldocchi, D.; Andy Black, T.; Verma, S.B.; Vesala, T.; Wofsy, S.C. Phenology of Vegetation Photosynthesis. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Tasks for Vegetation Science; Springer: Dordrecht, The Netherlands, 2003; Volume 39, pp. 467–485. ISBN 978-1-4020-1580-9. [Google Scholar]
- Peñuelas, J.; Rutishauser, T.; Filella, I. Phenology Feedbacks on Climate Change. Science 2009, 324, 887–888. [Google Scholar] [CrossRef]
- Richardson, A.D.; Andy Black, T.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of Spring and Autumn Phenological Transitions on Forest Ecosystem Productivity. Phil. Trans. R. Soc. B 2010, 365, 3227–3246. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf Chlorophyll Content as a Proxy for Leaf Photosynthetic Capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef]
- Schlemmer, M.; Gitelson, A.; Schepers, J.; Ferguson, R.; Peng, Y.; Shanahan, J.; Rundquist, D. Remote Estimation of Nitrogen and Chlorophyll Contents in Maize at Leaf and Canopy Levels. Int. J. Appl. Earth Obs. Geoinf. 2013, 25, 47–54. [Google Scholar] [CrossRef]
- Gitelson, A.A. Novel Technique for Remote Estimation of CO2 Flux in Maize. Geophys. Res. Lett. 2003, 30, 1486. [Google Scholar] [CrossRef]
- Schepers, J.S.; Francis, D.D.; Vigil, M.; Below, F.E. Comparison of Corn Leaf Nitrogen Concentration and Chlorophyll Meter Readings. Commun. Soil Sci. Plant Anal. 1992, 23, 2173–2187. [Google Scholar] [CrossRef]
- Daughtry, C. Estimating Corn Leaf Chlorophyll Concentration from Leaf and Canopy Reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Muchow, R.C.; Sinclair, T.R. Nitrogen Response of Leaf Photosynthesis and Canopy Radiation Use Efficiency in Field-Grown Maize and Sorghum. Crop Sci. 1994, 34, 721–727. [Google Scholar] [CrossRef]
- Allison, J.C.S.; Williams, H.T.; Pammenter, N.W. Effect of Specific Leaf Nitrogen Content on Photosynthesis of Sugarcane. Ann. Appl. Biol. 1997, 131, 339–350. [Google Scholar] [CrossRef]
- Houborg, R.; Cescatti, A.; Migliavacca, M.; Kustas, W.P. Satellite Retrievals of Leaf Chlorophyll and Photosynthetic Capacity for Improved Modeling of GPP. Agric. For. Meteorol. 2013, 177, 10–23. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Horie, T. Leaf Nitrogen, Photosynthesis, and Crop Radiation Use Efficiency: A Review. Crop Sci. 1989, 29, 90–98. [Google Scholar] [CrossRef]
- Muchow, R.C. Effect of Nitrogen Supply on the Comparative Productivity of Maize and Sorghum in a Semi-Arid Tropical Environment I. Leaf Growth and Leaf Nitrogen. Field Crops Res. 1988, 18, 1–16. [Google Scholar] [CrossRef]
- Hammer, G.L.; Van Oosterom, E.; McLean, G.; Chapman, S.C.; Broad, I.; Harland, P.; Muchow, R.C. Adapting APSIM to Model the Physiology and Genetics of Complex Adaptive Traits in Field Crops. J. Exp. Bot. 2010, 61, 2185–2202. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.R. AFRCWHEAT2: A Model of the Growth and Development of Wheat Incorporating Responses to Water and Nitrogen. Eur. J. Agron. 1993, 2, 69–82. [Google Scholar] [CrossRef]
- Wu, A.; Song, Y.; Van Oosterom, E.J.; Hammer, G.L. Connecting Biochemical Photosynthesis Models with Crop Models to Support Crop Improvement. Front. Plant Sci. 2016, 7, 1518. [Google Scholar] [CrossRef]
- Huang, X.; Xiao, J.; Ma, M. Evaluating the Performance of Satellite-Derived Vegetation Indices for Estimating Gross Primary Productivity Using FLUXNET Observations across the Globe. Remote Sens. 2019, 11, 1823. [Google Scholar] [CrossRef]
- Wang, J.; Dong, J.; Yi, Y.; Lu, G.; Oyler, J.; Smith, W.K.; Zhao, M.; Liu, J.; Running, S. Decreasing Net Primary Production Due to Drought and Slight Decreases in Solar Radiation in China from 2000 to 2012: Decreasing NPP Due To Solar Radiation. J. Geophys. Res. Biogeosci. 2017, 122, 261–278. [Google Scholar] [CrossRef]
- Liu, Y.; Ju, W.; He, H.; Wang, S.; Sun, R.; Zhang, Y. Changes of Net Primary Productivity in China during Recent 11 Years Detected Using an Ecological Model Driven by MODIS Data. Front. Earth Sci. 2013, 7, 112–127. [Google Scholar] [CrossRef]
- Verma, S.B.; Dobermann, A.; Cassman, K.G.; Walters, D.T.; Knops, J.M.; Arkebauer, T.J.; Suyker, A.E.; Burba, G.G.; Amos, B.; Yang, H.; et al. Annual Carbon Dioxide Exchange in Irrigated and Rainfed Maize-Based Agroecosystems. Agric. For. Meteorol. 2005, 131, 77–96. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring Vegetation Phenology Using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Wang, R.; Shan, J.; Tadesse, T.; Hayes, M.J.; Li, D. A Hybrid Approach for Detecting Corn and Soybean Phenology with Time-Series MODIS Data. Remote Sens. Environ. 2016, 181, 237–250. [Google Scholar] [CrossRef]
- Amani, M.; Ghorbanian, A.; Ahmadi, S.A.; Kakooei, M.; Moghimi, A.; Mirmazloumi, S.M.; Moghaddam, S.H.A.; Mahdavi, S.; Ghahremanloo, M.; Parsian, S.; et al. Google Earth Engine Cloud Computing Platform for Remote Sensing Big Data Applications: A Comprehensive Review. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 5326–5350. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Hu, S.; Zhang, X.; Zhou, G.; Peng, G.; Xiang, D.; Wang, R.; Meng, R.; Wu, W. A Novel Strategy to Reconstruct NDVI Time-Series with High Temporal Resolution from MODIS Multi-Temporal Composite Products. Remote Sens. 2021, 13, 1397. [Google Scholar] [CrossRef]
- Dash, J.; Curran, P.J.; Tallis, M.J.; Llewellyn, G.M.; Taylor, G.; Snoeij, P. Validating the MERIS Terrestrial Chlorophyll Index (MTCI) with Ground Chlorophyll Content Data at MERIS Spatial Resolution. Int. J. Remote Sens. 2010, 31, 5513–5532. [Google Scholar] [CrossRef]
- Wu, B.; Zhang, M.; Zeng, H.; Tian, F.; Potgieter, A.B.; Qin, X.; Yan, N.; Chang, S.; Zhao, Y.; Dong, Q.; et al. Challenges and Opportunities in Remote Sensing-Based Crop Monitoring: A Review. Natl. Sci. Rev. 2023, 10, nwac290. [Google Scholar] [CrossRef]
- Godwin, D.C.; Singh, U. Nitrogen Balance and Crop Response to Nitrogen in Upland and Lowland Cropping Systems. In Understanding Options for Agricultural Production; Tsuji, G.Y., Hoogenboom, G., Thornton, P.K., Eds.; Systems Approaches for Sustainable Agricultural Development; Springer: Dordrecht, The Netherlands, 1998; Volume 7, pp. 55–77. ISBN 978-90-481-4940-7. [Google Scholar]
- Boote, K. (Ed.) Advances in Crop Modelling for a Sustainable Agriculture; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; ISBN 978-0-429-26659-1. [Google Scholar]
- Manschadi, A.M.; Eitzinger, J.; Breisch, M.; Fuchs, W.; Neubauer, T.; Soltani, A. Full Parameterisation Matters for the Best Performance of Crop Models: Inter-Comparison of a Simple and a Detailed Maize Model. Int. J. Plant Prod. 2021, 15, 61–78. [Google Scholar] [CrossRef]
- Soufizadeh, S.; Munaro, E.; McLean, G.; Massignam, A.; Van Oosterom, E.J.; Chapman, S.C.; Messina, C.; Cooper, M.; Hammer, G.L. Modelling the Nitrogen Dynamics of Maize Crops—Enhancing the APSIM Maize Model. Eur. J. Agron. 2018, 100, 118–131. [Google Scholar] [CrossRef]
- Groenendijk, P.; Boogaard, H.; Heinen, M.; Kroes, J.G.; Supit, I.; de Wit, A. Simulation Nitrogen-Limited Crop Growth with SWAP/WOFOST: Process Descriptions and User Manual; Wageningen Environmental Research: Wageningen, The Netherlands, 2016. [Google Scholar]
- Körner, C.; Basler, D. Phenology Under Global Warming. Science 2010, 327, 1461–1462. [Google Scholar] [CrossRef] [PubMed]
- Tollenaar, M.; Daynard, T.B.; Hunter, R.B. Effect of Temperature on Rate of Leaf Appearance and Flowering Date in Maize. Crop Sci. 1979, 19, 363–366. [Google Scholar] [CrossRef]
- Wang, E.; Engel, T. Simulation of Phenological Development of Wheat Crops. Agric. Syst. 1998, 58, 1–24. [Google Scholar] [CrossRef]
- Bannayan, M.; Hoogenboom, G.; Crout, N.M.J. Photothermal Impact on Maize Performance: A Simulation Approach. Ecol. Model. 2004, 180, 277–290. [Google Scholar] [CrossRef]
- Hickin, R.P.; Vittum, M.T. The Importance of Soil and Air Temperature in Spring Phenoclimatic Modelling. Int. J. Biometeorol. 1976, 20, 200–206. [Google Scholar] [CrossRef]
- Cutforth, H.W.; Shaykewich, C.F. A Temperature Response Function for Corn Development. Agric. For. Meteorol. 1990, 50, 159–171. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Breiman, L. Bagging Predictors. Mach. Learn. 1996, 24, 123–140. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, W.; Gao, S.; Zhang, K.; Wang, J.; Huang, N. Estimating Deciduous Broadleaf Forest Gross Primary Productivity by Remote Sensing Data Using a Random Forest Regression Model. J. Appl. Rem. Sens. 2019, 13, 1. [Google Scholar] [CrossRef]
- Chang, X.; Xing, Y.; Gong, W.; Yang, C.; Guo, Z.; Wang, D.; Wang, J.; Yang, H.; Xue, G.; Yang, S. Evaluating Gross Primary Productivity over 9 ChinaFlux Sites Based on Random Forest Regression Models, Remote Sensing, and Eddy Covariance Data. Sci. Total Environ. 2023, 875, 162601. [Google Scholar] [CrossRef]
- Scientific Data Curation Team Metadata Record for: Global Terrestrial Carbon Fluxes of 1999–2019 Estimated by Upscaling Eddy Covariance Data with a Random Forest 2020, 5018 Bytes. Available online: https://pubmed.ncbi.nlm.nih.gov/32973132/ (accessed on 20 November 2023).
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning; Springer Series in Statistics; Springer: New York, NY, USA, 2009; ISBN 978-0-387-84857-0. [Google Scholar]
- Ichii, K.; Ueyama, M.; Kondo, M.; Saigusa, N.; Kim, J.; Alberto, M.C.; Ardö, J.; Euskirchen, E.S.; Kang, M.; Hirano, T.; et al. New Data-driven Estimation of Terrestrial CO 2 Fluxes in Asia Using a Standardized Database of Eddy Covariance Measurements, Remote Sensing Data, and Support Vector Regression. JGR Biogeosci. 2017, 122, 767–795. [Google Scholar] [CrossRef]
- Yang, F.; Ichii, K.; White, M.A.; Hashimoto, H.; Michaelis, A.R.; Votava, P.; Zhu, A.-X.; Huete, A.; Running, S.W.; Nemani, R.R. Developing a Continental-Scale Measure of Gross Primary Production by Combining MODIS and AmeriFlux Data through Support Vector Machine Approach. Remote Sens. Environ. 2007, 110, 109–122. [Google Scholar] [CrossRef]
- Yu, T.; Zhang, Q.; Sun, R. Comparison of Machine Learning Methods to Up-Scale Gross Primary Production. Remote Sens. 2021, 13, 2448. [Google Scholar] [CrossRef]
- Yang, F.; White, M.A.; Michaelis, A.R.; Ichii, K.; Hashimoto, H.; Votava, P.; Zhu, A.-X.; Nemani, R.R. Prediction of Continental-Scale Evapotranspiration by Combining MODIS and AmeriFlux Data Through Support Vector Machine. IEEE Trans. Geosci. Remote Sens. 2006, 44, 3452–3461. [Google Scholar] [CrossRef]
- Gu, J.; Wang, Z.; Kuen, J.; Ma, L.; Shahroudy, A.; Shuai, B.; Liu, T.; Wang, X.; Wang, G.; Cai, J.; et al. Recent Advances in Convolutional Neural Networks. Pattern Recognit. 2018, 77, 354–377. [Google Scholar] [CrossRef]
- Oquab, M.; Bottou, L.; Laptev, I.; Sivic, J. Learning and Transferring Mid-Level Image Representations Using Convolutional Neural Networks. In Proceedings of the 2014 IEEE Conference on Computer Vision and Pattern Recognition, Columbus, OH, USA, 23–28 June 2014; pp. 1717–1724. [Google Scholar]
- Pan, S.J.; Yang, Q. A Survey on Transfer Learning. IEEE Trans. Knowl. Data Eng. 2010, 22, 1345–1359. [Google Scholar] [CrossRef]
- Huang, G.-B.; Zhu, Q.-Y.; Siew, C.-K. Extreme Learning Machine: Theory and Applications. Neurocomputing 2006, 70, 489–501. [Google Scholar] [CrossRef]
- Wang, J.; Lu, S.; Wang, S.-H.; Zhang, Y.-D. A Review on Extreme Learning Machine. Multimed. Tools Appl. 2022, 81, 41611–41660. [Google Scholar] [CrossRef]
- Xiao, J.; Zhuang, Q.; Baldocchi, D.D.; Law, B.E.; Richardson, A.D.; Chen, J.; Oren, R.; Starr, G.; Noormets, A.; Ma, S.; et al. Estimation of Net Ecosystem Carbon Exchange for the Conterminous United States by Combining MODIS and AmeriFlux Data. Agric. For. Meteorol. 2008, 148, 1827–1847. [Google Scholar] [CrossRef]
- Duan, Z.; Yang, Y.; Zhou, S.; Gao, Z.; Zong, L.; Fan, S.; Yin, J. Estimating Gross Primary Productivity (GPP) over Rice–Wheat-Rotation Croplands by Using the Random Forest Model and Eddy Covariance Measurements: Upscaling and Comparison with the MODIS Product. Remote Sens. 2021, 13, 4229. [Google Scholar] [CrossRef]
- Tramontana, G.; Ichii, K.; Camps-Valls, G.; Tomelleri, E.; Papale, D. Uncertainty Analysis of Gross Primary Production Upscaling Using Random Forests, Remote Sensing and Eddy Covariance Data. Remote Sens. Environ. 2015, 168, 360–373. [Google Scholar] [CrossRef]
- Wang, Q.; Yue, C.; Li, X.; Liao, P.; Li, X. Enhancing Robustness of Monthly Streamflow Forecasting Model Using Embedded-Feature Selection Algorithm Based on Improved Gray Wolf Optimizer. J. Hydrol. 2023, 617, 128995. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Huang, C.; Qiao, N. An NDVI-Based Vegetation Phenology Is Improved to Be More Consistent with Photosynthesis Dynamics through Applying a Light Use Efficiency Model over Boreal High-Latitude Forests. Remote Sens. 2017, 9, 695. [Google Scholar] [CrossRef]
- You, Y.; Wang, S.; Pan, N.; Ma, Y.; Liu, W. Growth Stage-Dependent Responses of Carbon Fixation Process of Alpine Grasslands to Climate Change over the Tibetan Plateau, China. Agric. For. Meteorol. 2020, 291, 108085. [Google Scholar] [CrossRef]
- Min, J.; Lee, Y. Bankruptcy Prediction Using Support Vector Machine with Optimal Choice of Kernel Function Parameters. Expert Syst. Appl. 2005, 28, 603–614. [Google Scholar] [CrossRef]
- Wang, H.; Shao, W.; Hu, Y.; Cao, W.; Zhang, Y. Assessment of Six Machine Learning Methods for Predicting Gross Primary Productivity in Grassland. Remote Sens. 2023, 15, 3475. [Google Scholar] [CrossRef]
- Zhou, H.; Yue, X.; Lei, Y.; Zhang, T.; Tian, C.; Ma, Y.; Cao, Y. Responses of Gross Primary Productivity to Diffuse Radiation at Global FLUXNET Sites. Atmos. Environ. 2021, 244, 117905. [Google Scholar] [CrossRef]
- Camps-Valls, G.; Campos-Taberner, M.; Moreno-Martínez, Á.; Walther, S.; Duveiller, G.; Cescatti, A.; Mahecha, M.D.; Muñoz-Marí, J.; García-Haro, F.J.; Guanter, L.; et al. A Unified Vegetation Index for Quantifying the Terrestrial Biosphere. Sci. Adv. 2021, 7, eabc7447. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Huang, G.; Xu, J.; Tang, J. Extreme Learning Machines: New Trends and Applications. Sci. China Inf. Sci. 2015, 58, 1–16. [Google Scholar] [CrossRef]
- Peng, Y.; Gitelson, A.A.; Sakamoto, T. Remote Estimation of Gross Primary Productivity in Crops Using MODIS 250m Data. Remote Sens. Environ. 2013, 128, 186–196. [Google Scholar] [CrossRef]
- Liu, J.; Zuo, Y.; Wang, N.; Yuan, F.; Zhu, X.; Zhang, L.; Zhang, J.; Sun, Y.; Guo, Z.; Guo, Y.; et al. Comparative Analysis of Two Machine Learning Algorithms in Predicting Site-Level Net Ecosystem Exchange in Major Biomes. Remote Sens. 2021, 13, 2242. [Google Scholar] [CrossRef]
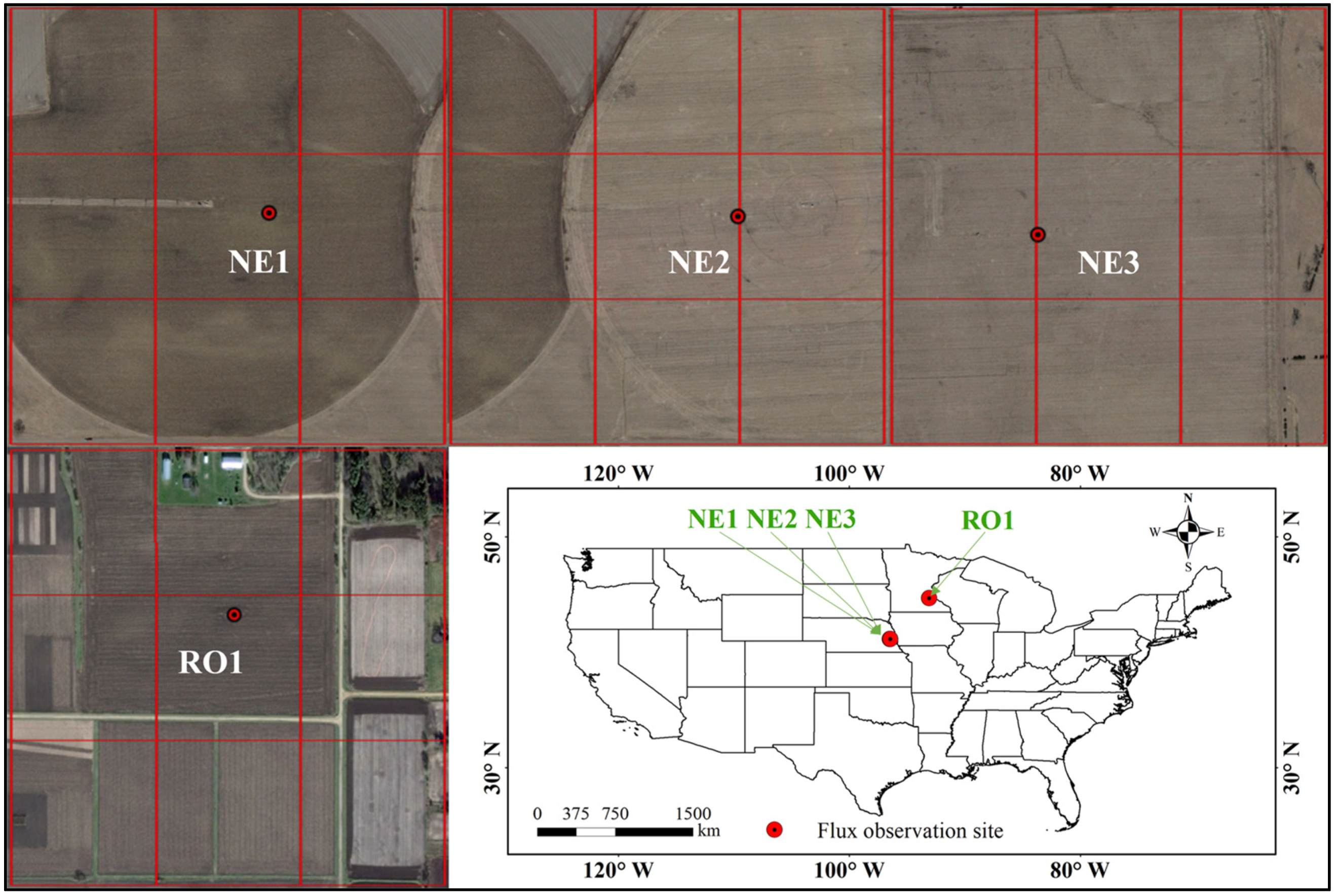
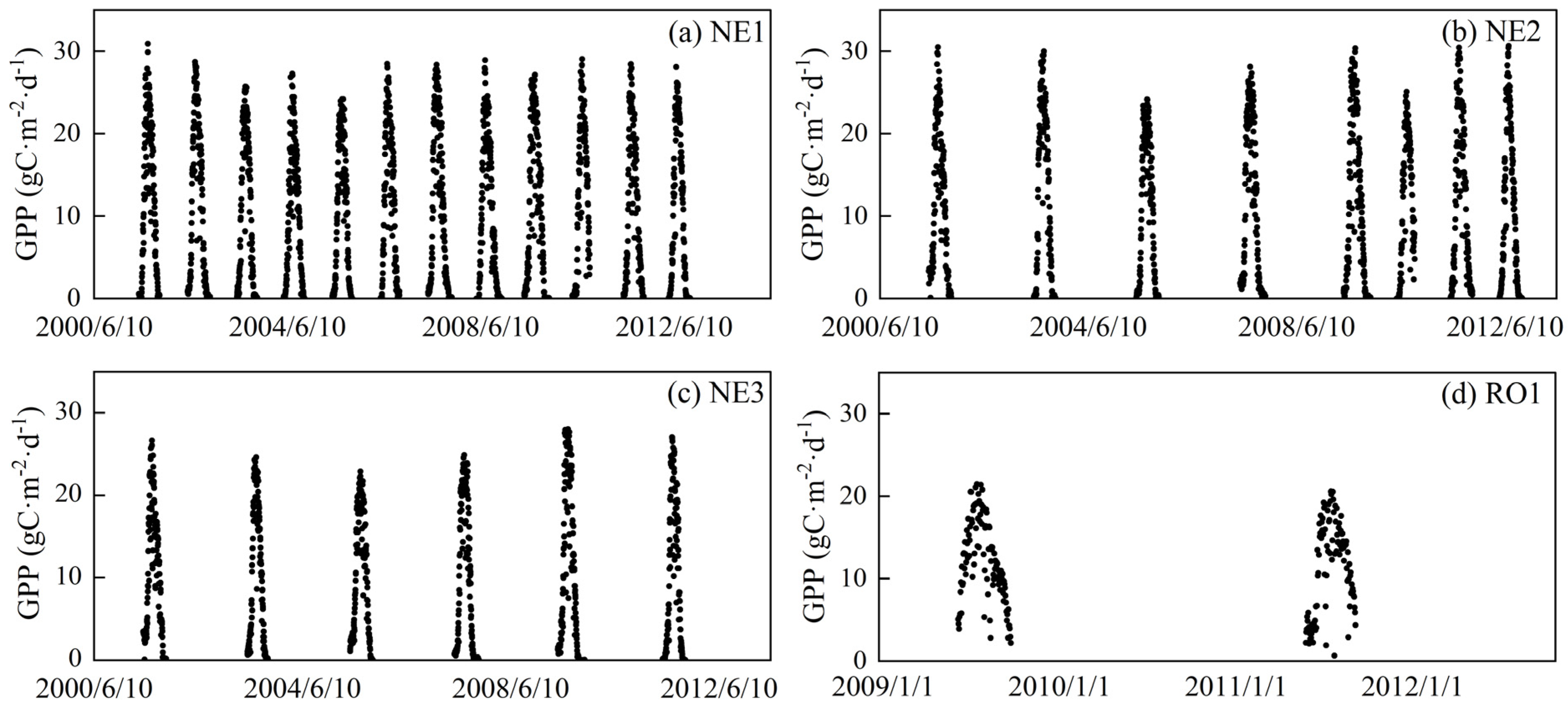
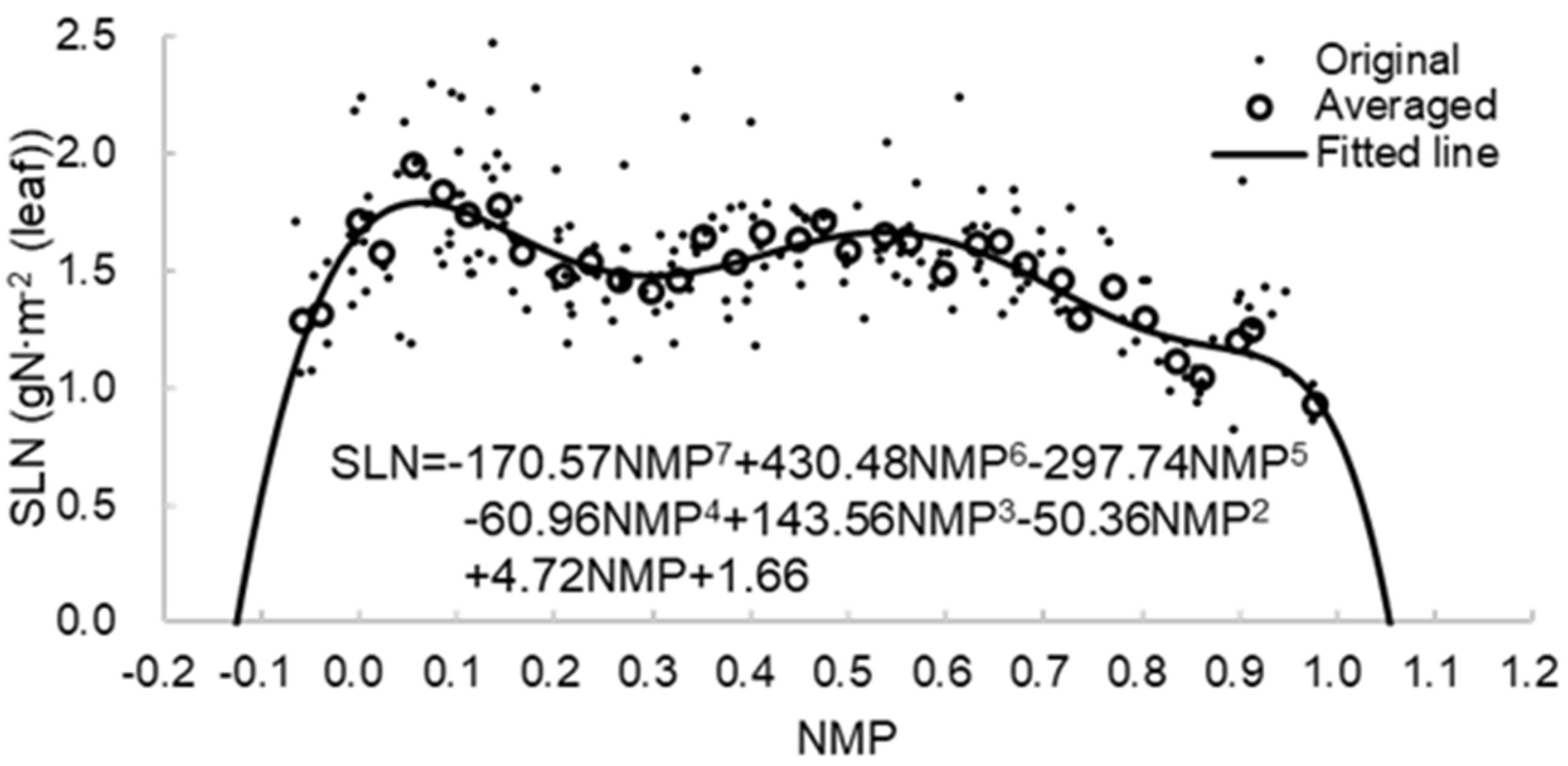
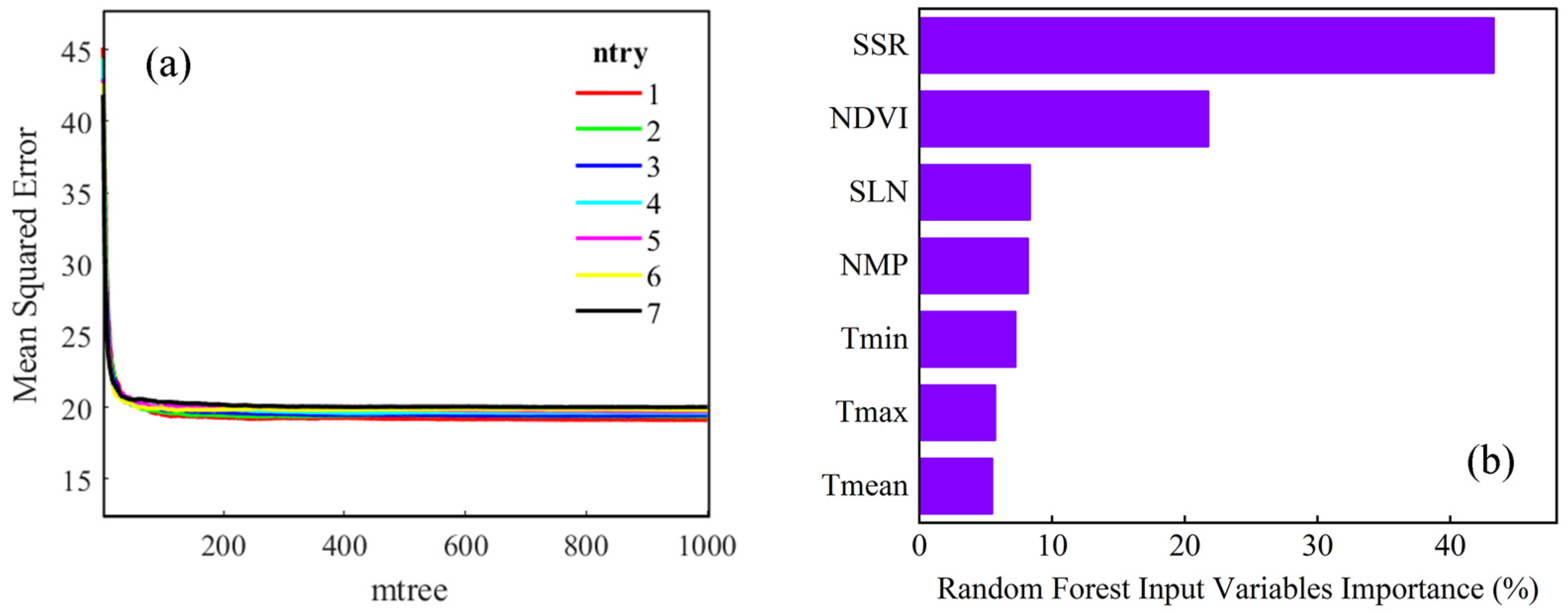

| Site | Longitude (°W) | Latitude (°N) | Available Year | Data Size |
|---|---|---|---|---|
| NE1 | −96.4766 | 41.1651 | 2001, 2002, 2003, 2004, 2005, 2006, 2007, 2008, 2009, 2010, 2011, 2012 | 1945 |
| NE2 | −96.4701 | 41.1649 | 2001, 2003, 2005, 2007, 2009~2012 | 1276 |
| NE3 | −96.4397 | 41.1797 | 2001, 2003, 2005, 2007, 2009, 2011 | 987 |
| RO1 | −93.0898 | 44.7143 | 2009, 2011 | 214 |
| Symbol | Input Variable Combination |
|---|---|
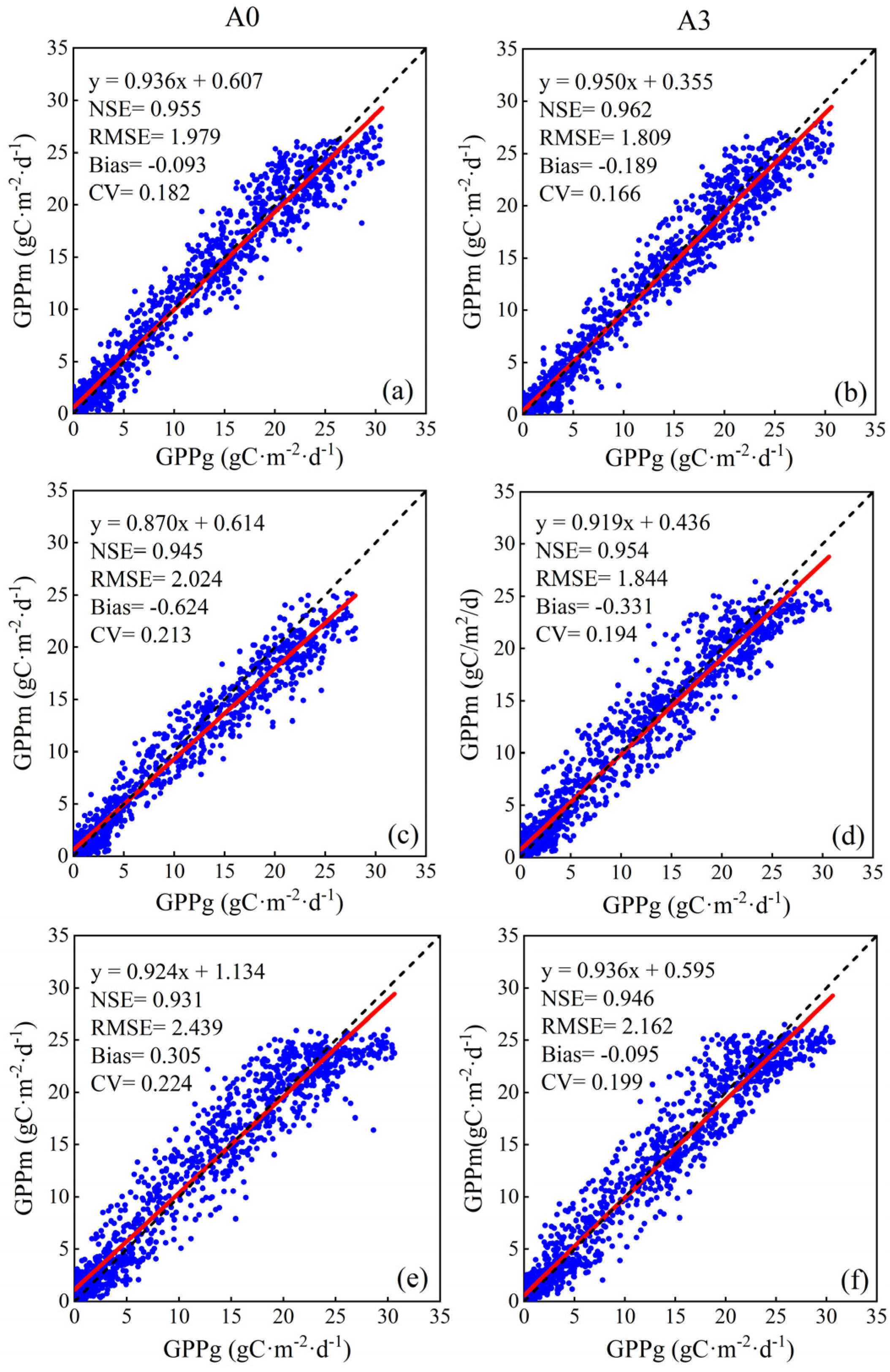
| A0 | NDVI + Tmean + Tmin + Tmax + SSR |
| A1 | NDVI + Tmean + Tmin + Tmax + SSR + SLN |
| A2 | NDVI + Tmean + Tmin + Tmax + SSR + NMP |
| A3 | NDVI + Tmean + Tmin + Tmax + SSR + SLN + NMP |
| NSE | RMSE | Bias | CV | ||||||
|---|---|---|---|---|---|---|---|---|---|
| μ | σ | μ | σ | μ | σ | μ | σ | ||
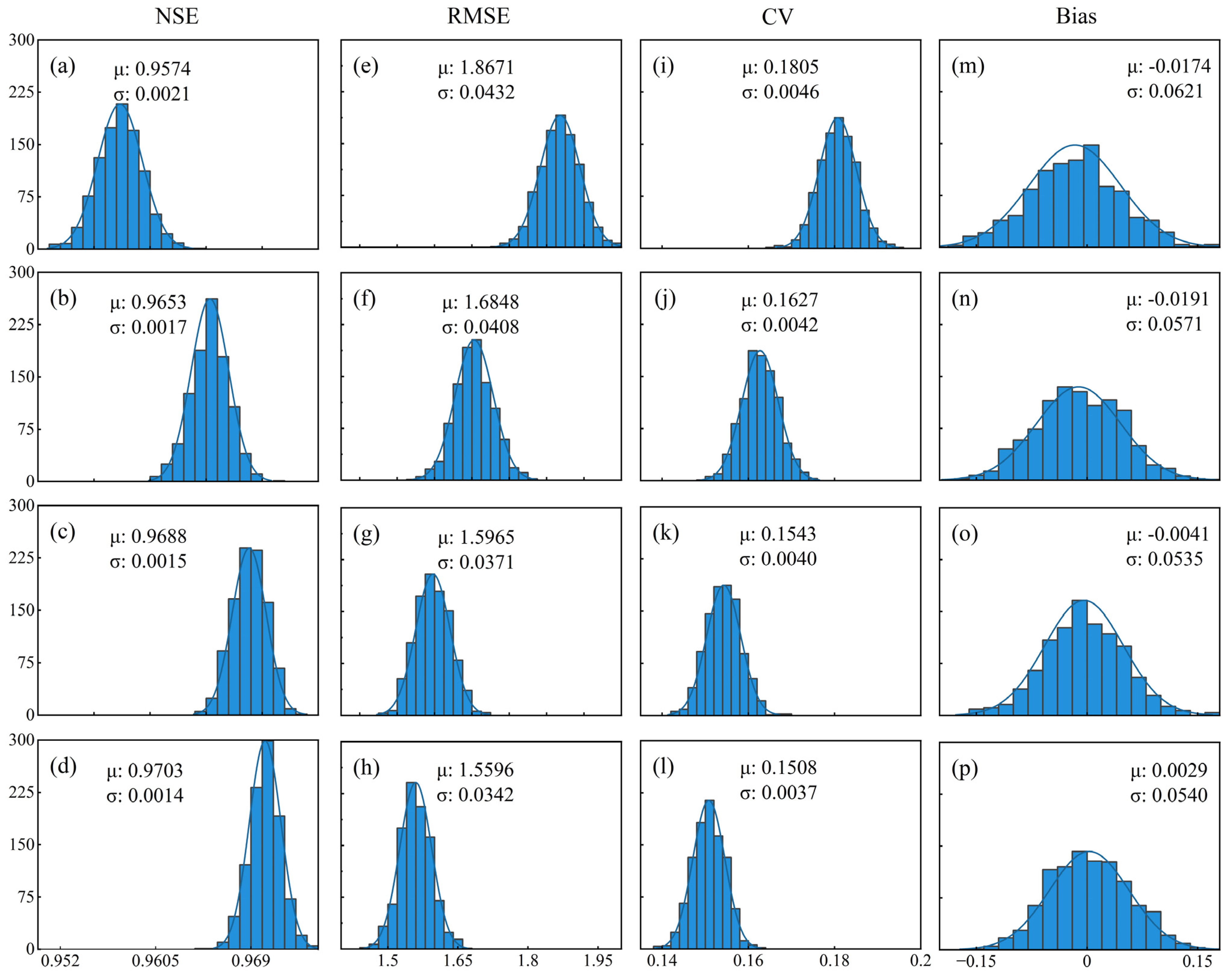
| RF | A0 | 0.9574 | 0.0021 | 1.8671 | 0.0432 | −0.0174 | 0.0621 | 0.1805 | 0.0046 |
| A1 | 0.9653 | 0.0017 | 1.6848 | 0.0408 | −0.0115 | 0.0571 | 0.1627 | 0.0042 | |
| A2 | 0.9688 | 0.0015 | 1.5965 | 0.0371 | −0.0041 | 0.0535 | 0.1543 | 0.0040 | |
| A3 | 0.9703 | 0.0014 | 1.5596 | 0.0342 | 0.0029 | 0.0539 | 0.1508 | 0.0037 | |
| SVM | A0 | 0.9589 | 0.0019 | 1.8357 | 0.0401 | −0.0521 | 0.0651 | 0.1772 | 0.0043 |
| A1 | 0.9668 | 0.0016 | 1.6480 | 0.0378 | −0.0139 | 0.0556 | 0.1594 | 0.0038 | |
| A2 | 0.9699 | 0.0014 | 1.5703 | 0.0353 | 0.0064 | 0.0562 | 0.1517 | 0.0039 | |
| A3 | 0.9706 | 0.0014 | 1.5509 | 0.0363 | 0.0163 | 0.0569 | 0.1470 | 0.0038 | |
| CNN | A0 | 0.9529 | 0.0059 | 1.9610 | 0.1151 | −0.1513 | 0.1848 | 0.1897 | 0.0112 |
| A1 | 0.9553 | 0.0029 | 1.9103 | 0.0637 | −0.0454 | 0.1859 | 0.1847 | 0.0064 | |
| A2 | 0.9609 | 0.0029 | 1.7872 | 0.0647 | −0.0231 | 0.1535 | 0.1729 | 0.0066 | |
| A3 | 0.9597 | 0.0022 | 1.8152 | 0.0490 | −0.0031 | 0.1763 | 0.1755 | 0.0051 | |
| ELM | A0 | 0.9578 | 0.0019 | 1.8595 | 0.0390 | −0.0004 | 0.0631 | 0.1795 | 0.0041 |
| A1 | 0.9644 | 0.0016 | 1.7069 | 0.0374 | 0.0004 | 0.0567 | 0.1650 | 0.0038 | |
| A2 | 0.9681 | 0.0014 | 1.6146 | 0.0338 | 0.0014 | 0.8541 | 0.1560 | 0.0037 | |
| A3 | 0.9674 | 0.0015 | 1.6321 | 0.0363 | 0.0013 | 0.0542 | 0.1579 | 0.0039 | |
| RMSE | URMSE | ||
|---|---|---|---|
| RF | A0 | 2.837 | 2.771 |
| A3 | 2.654 | 2.213 | |
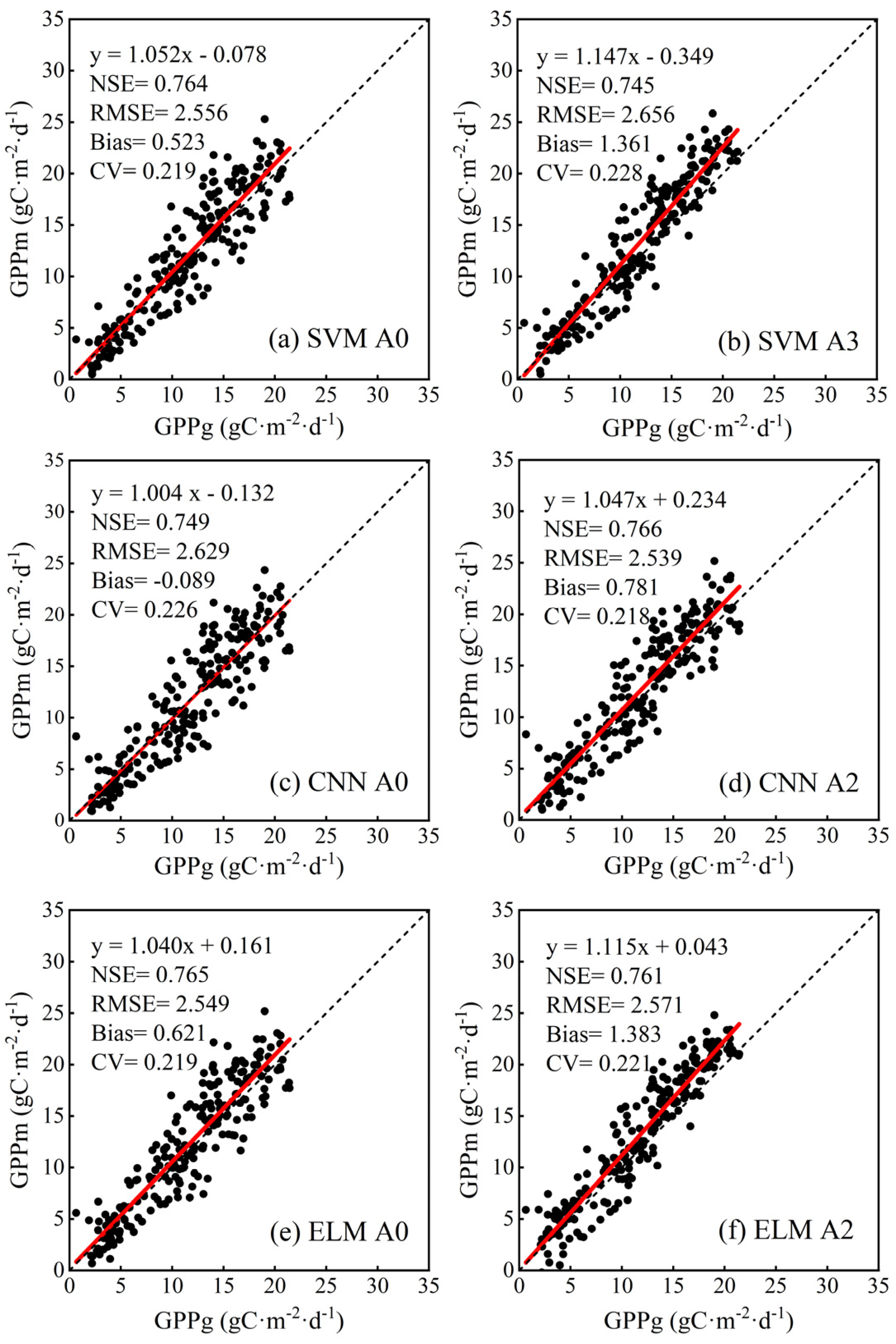
| SVM | A0 | 2.556 | 2.502 |
| A3 | 2.656 | 2.280 | |
| CNN | A0 | 2.629 | 2.627 |
| A2 | 2.539 | 2.417 | |
| ELM | A0 | 2.549 | 2.472 |
| A2 | 2.571 | 2.168 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.; Hu, S.; Zeng, L.; Meng, K.; Liao, Z.; Wang, K. Estimation of Daily Maize Gross Primary Productivity by Considering Specific Leaf Nitrogen and Phenology via Machine Learning Methods. Remote Sens. 2024, 16, 341. https://doi.org/10.3390/rs16020341
Hu C, Hu S, Zeng L, Meng K, Liao Z, Wang K. Estimation of Daily Maize Gross Primary Productivity by Considering Specific Leaf Nitrogen and Phenology via Machine Learning Methods. Remote Sensing. 2024; 16(2):341. https://doi.org/10.3390/rs16020341
Chicago/Turabian StyleHu, Cenhanyi, Shun Hu, Linglin Zeng, Keyu Meng, Zilong Liao, and Kuang Wang. 2024. "Estimation of Daily Maize Gross Primary Productivity by Considering Specific Leaf Nitrogen and Phenology via Machine Learning Methods" Remote Sensing 16, no. 2: 341. https://doi.org/10.3390/rs16020341
APA StyleHu, C., Hu, S., Zeng, L., Meng, K., Liao, Z., & Wang, K. (2024). Estimation of Daily Maize Gross Primary Productivity by Considering Specific Leaf Nitrogen and Phenology via Machine Learning Methods. Remote Sensing, 16(2), 341. https://doi.org/10.3390/rs16020341