
Satellite and High-Spatio-Temporal Resolution Data Collected by Southern Elephant Seals Allow an Unprecedented 3D View of the Argentine Continental Shelf
 , , , and
, , , and
Abstract
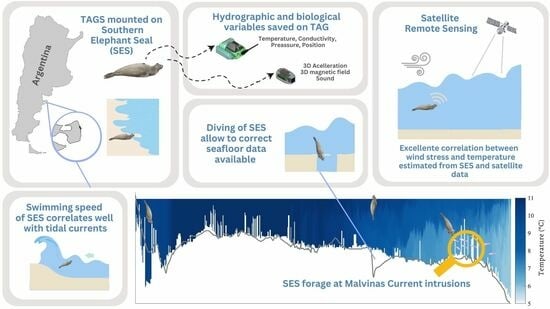
:
1. Introduction
2. Materials and Methods
2.1. In-Situ Data
2.2. Satellite Data
2.3. Reanalysis Data
2.4. Bathymetry Data
2.5. Tidal Model
2.6. Methodology
3. Results
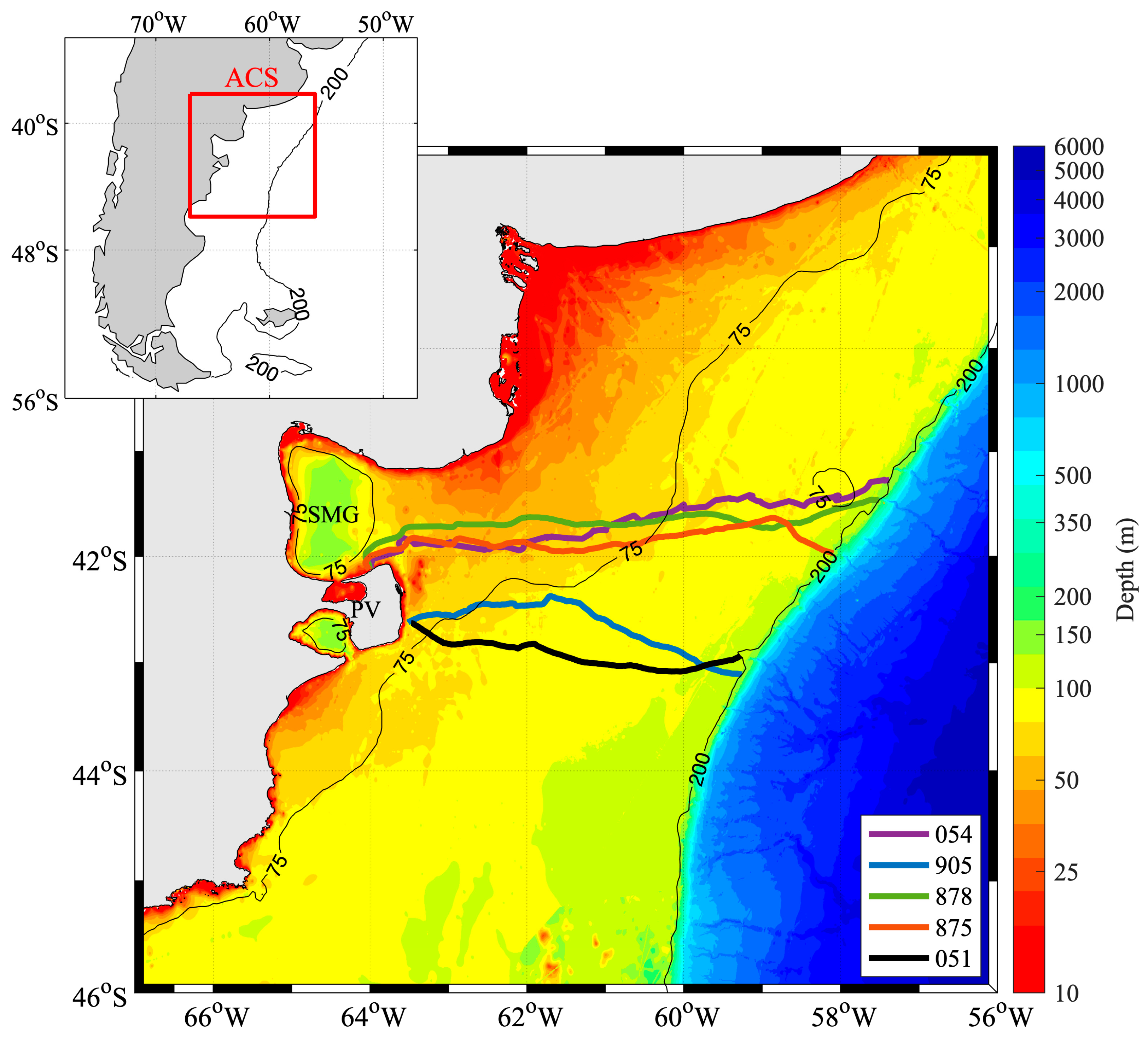
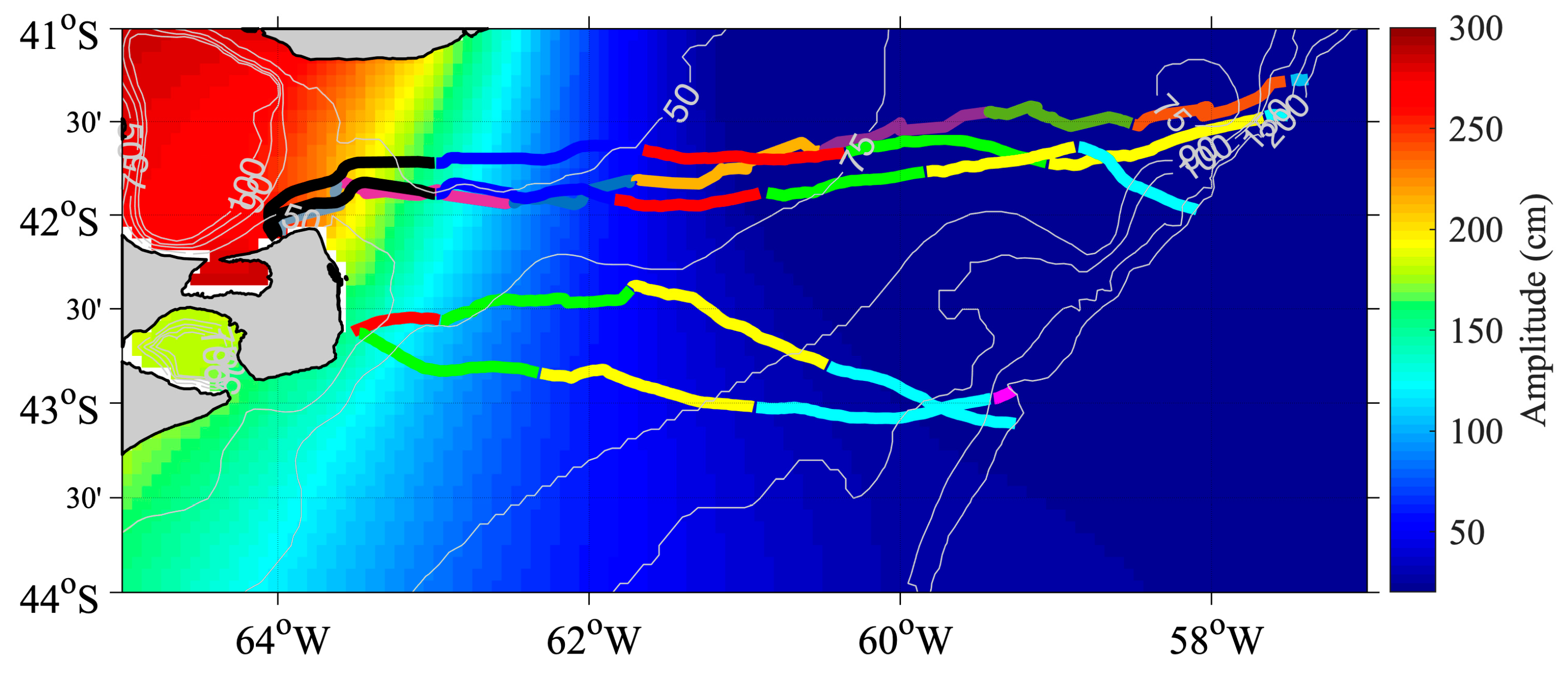
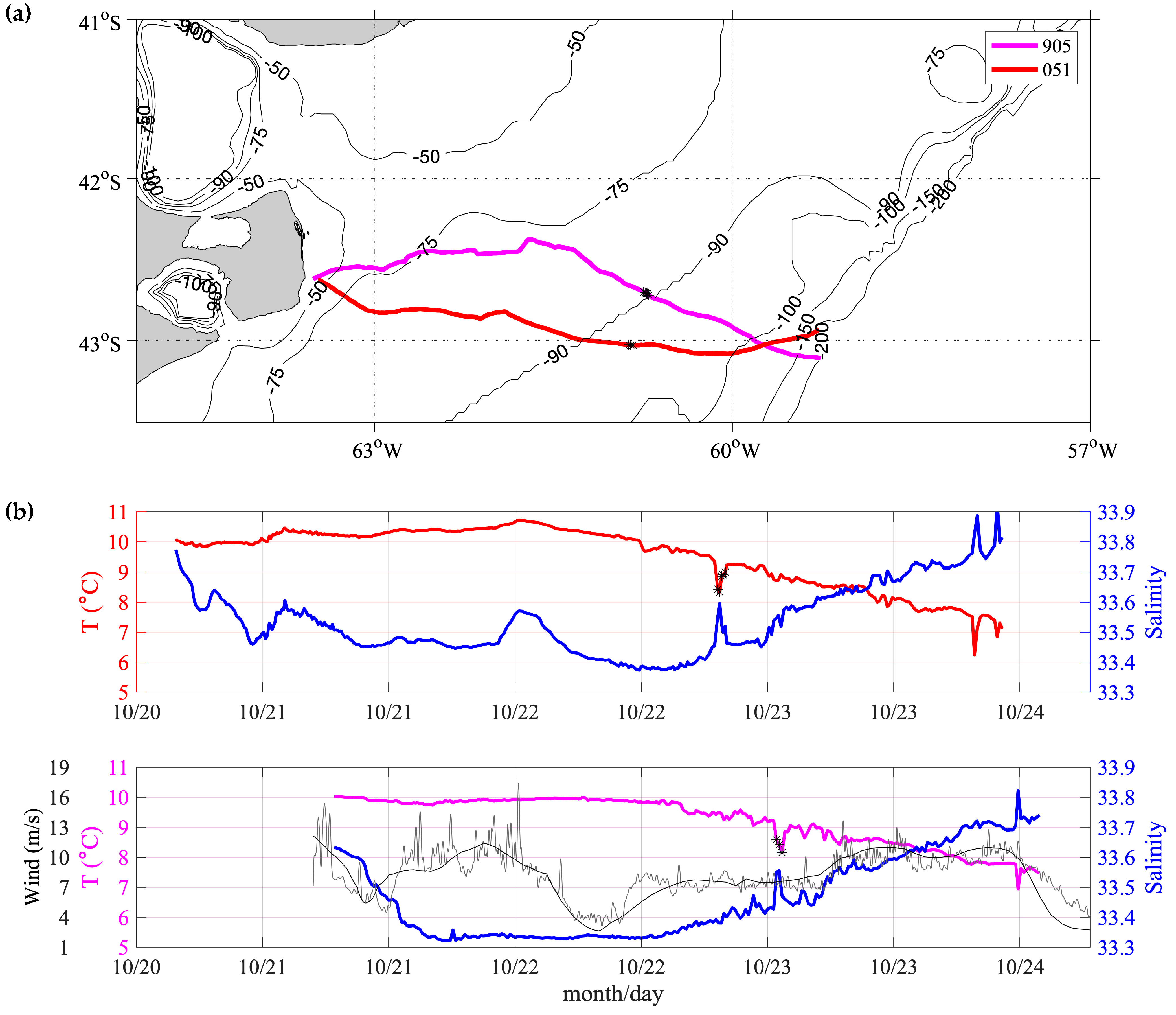
3.1. SESs Trajectories
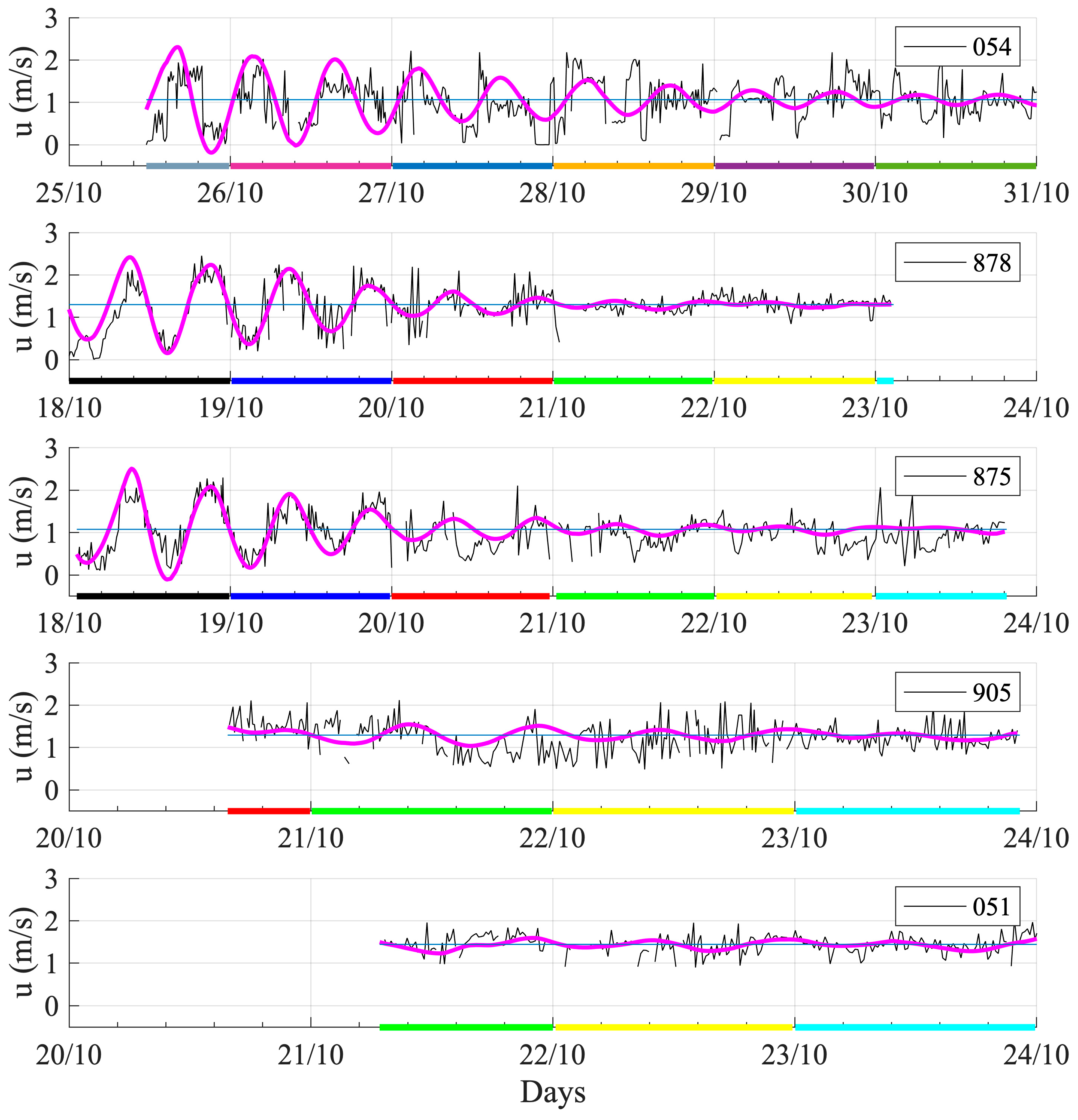
3.2. SESs Speeds
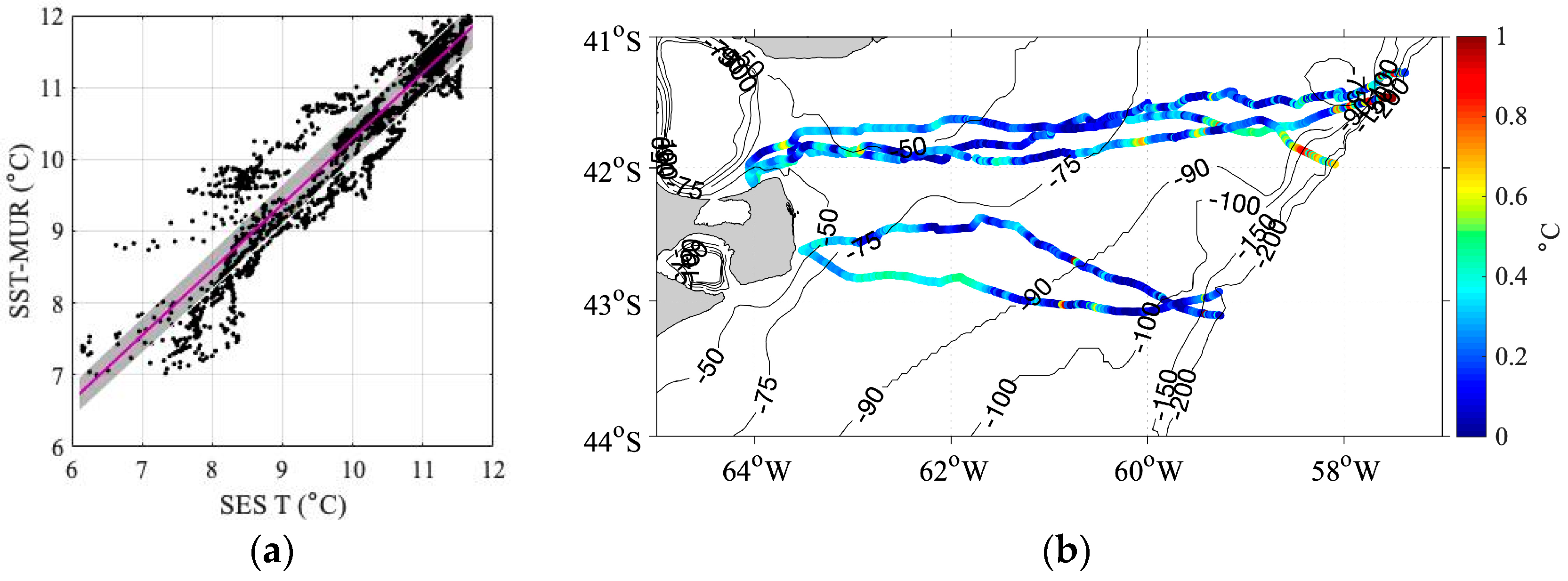
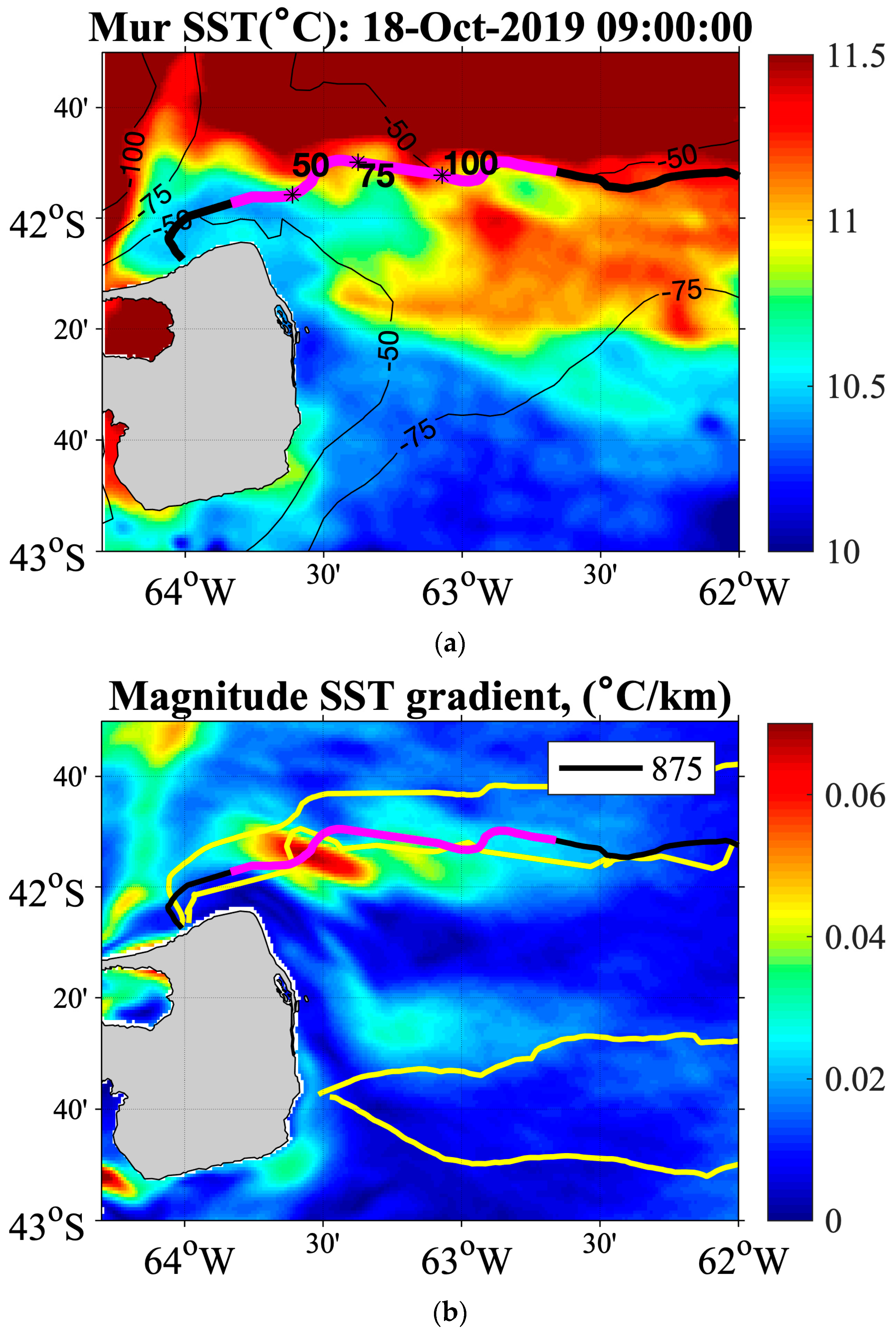
3.3. Comparison between Near-Surface SES Temperatures and Satellite SST
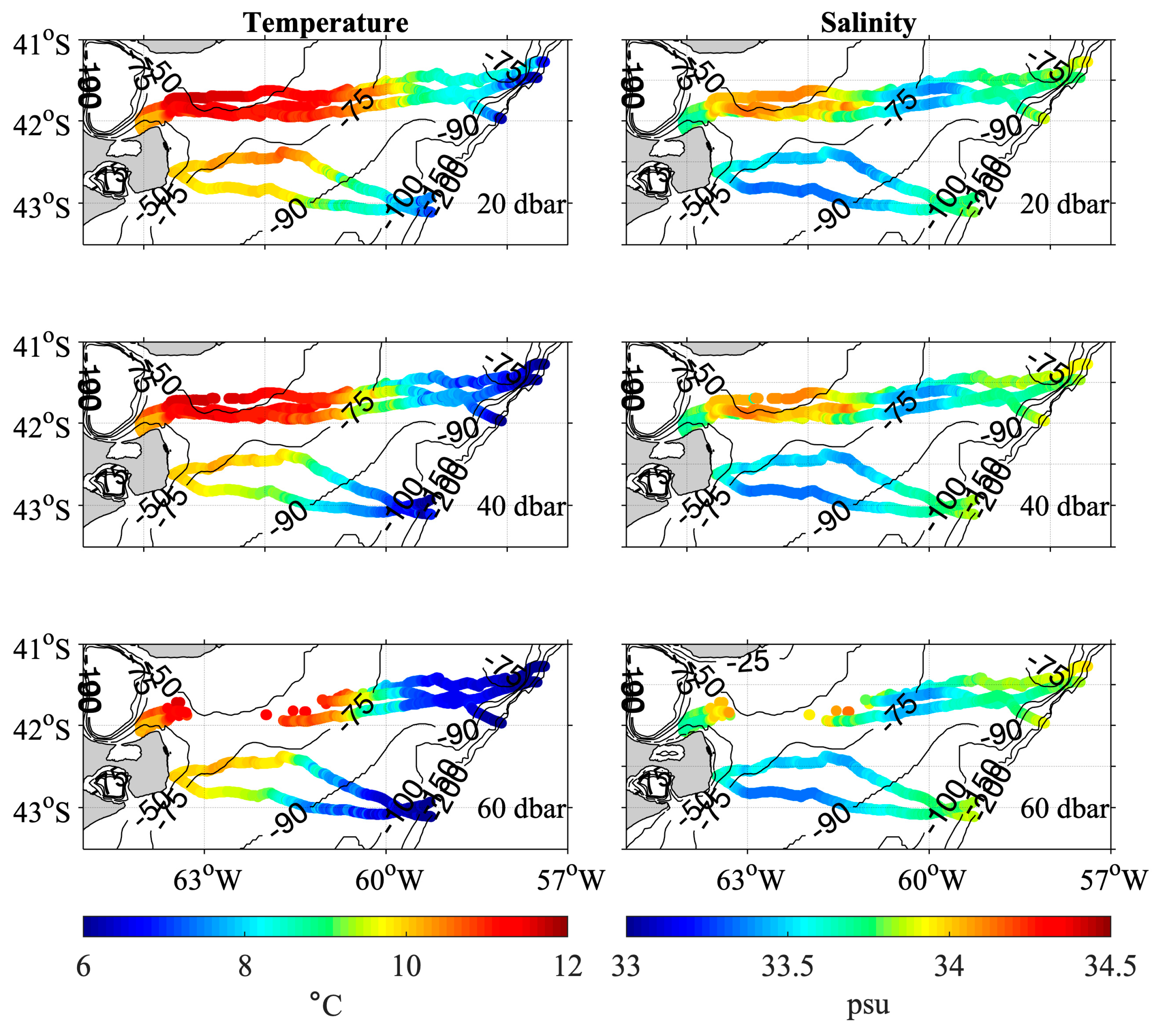
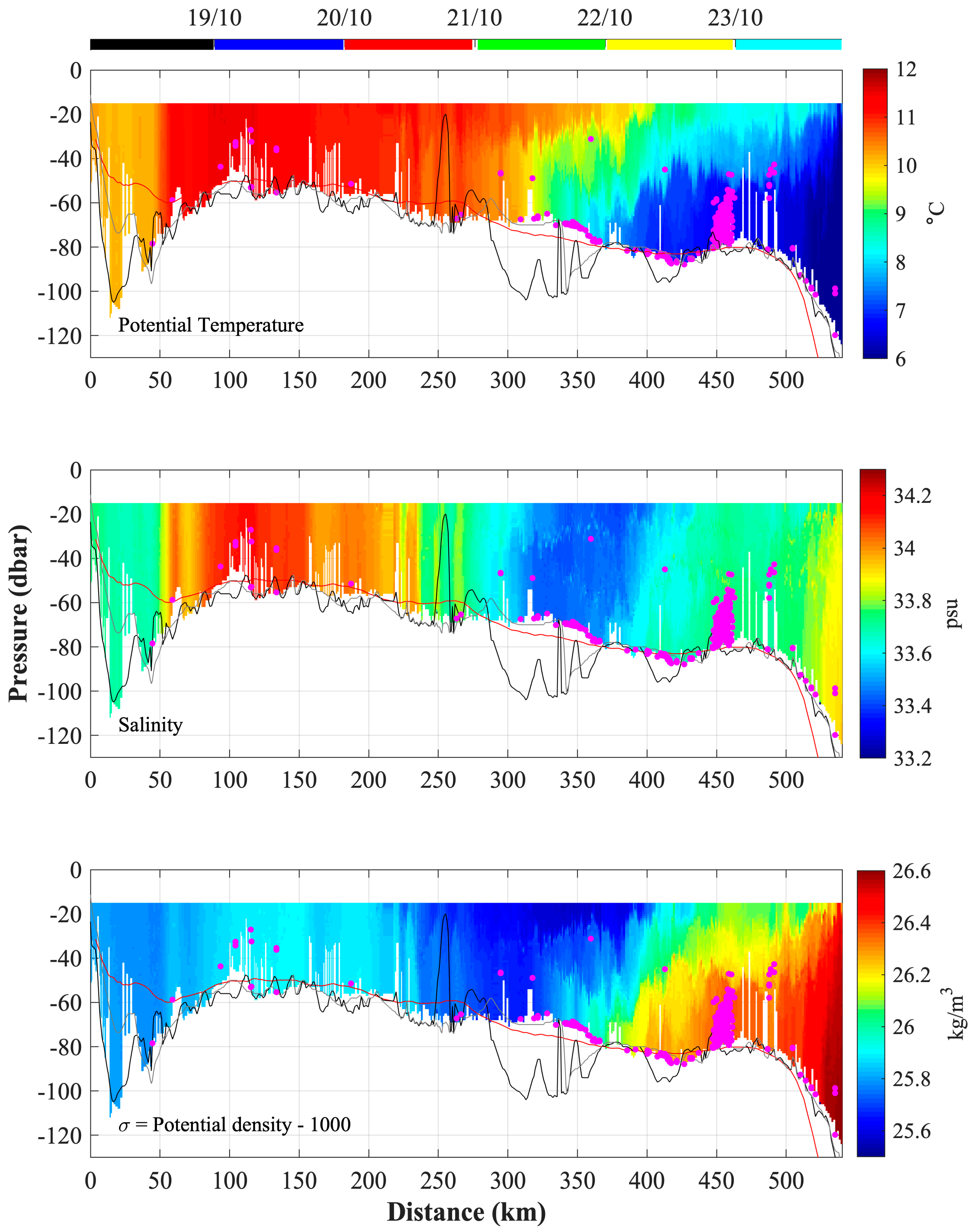
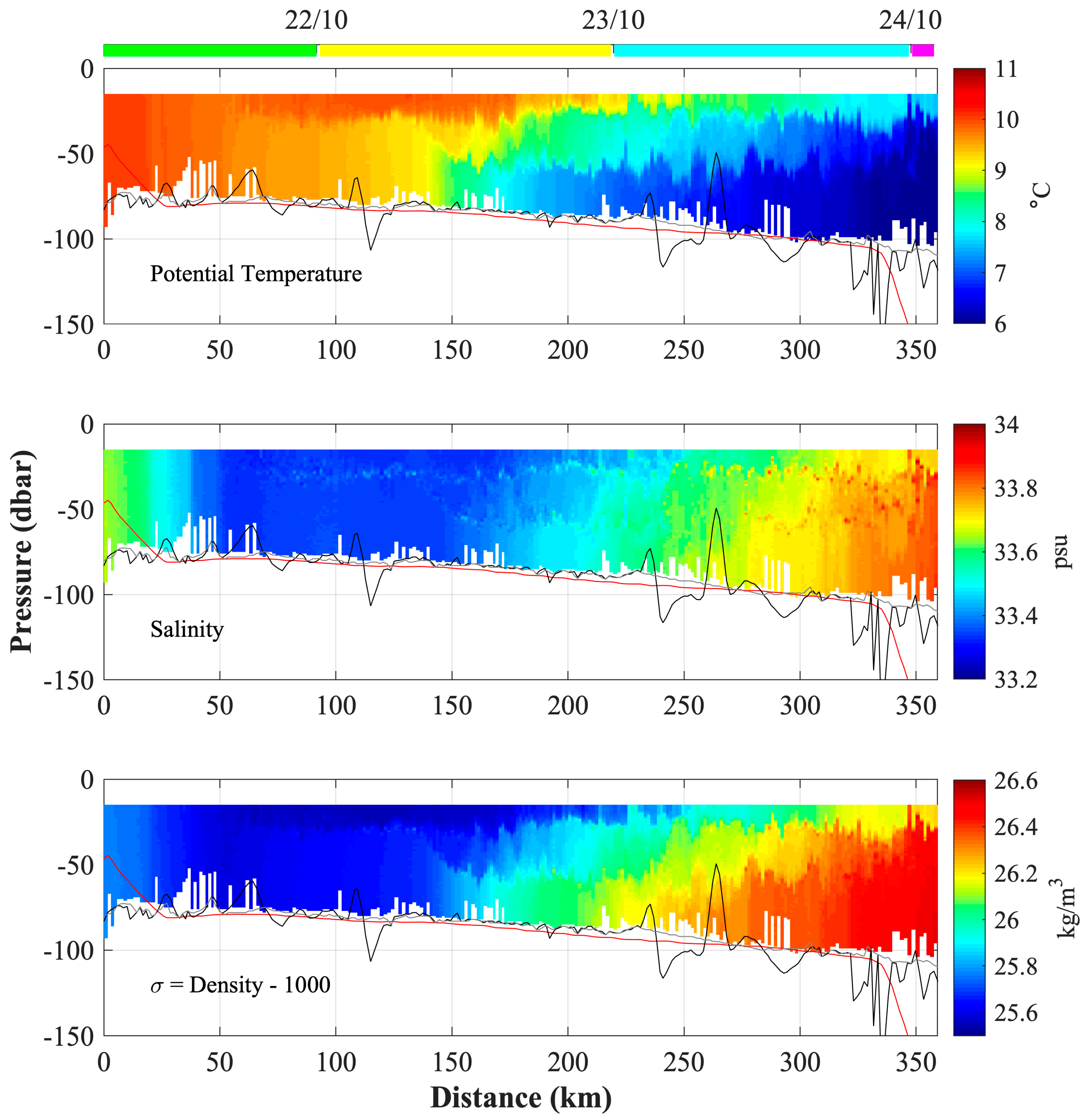
3.4. Analysis of the Temperature and Salinity Spatial Distributions
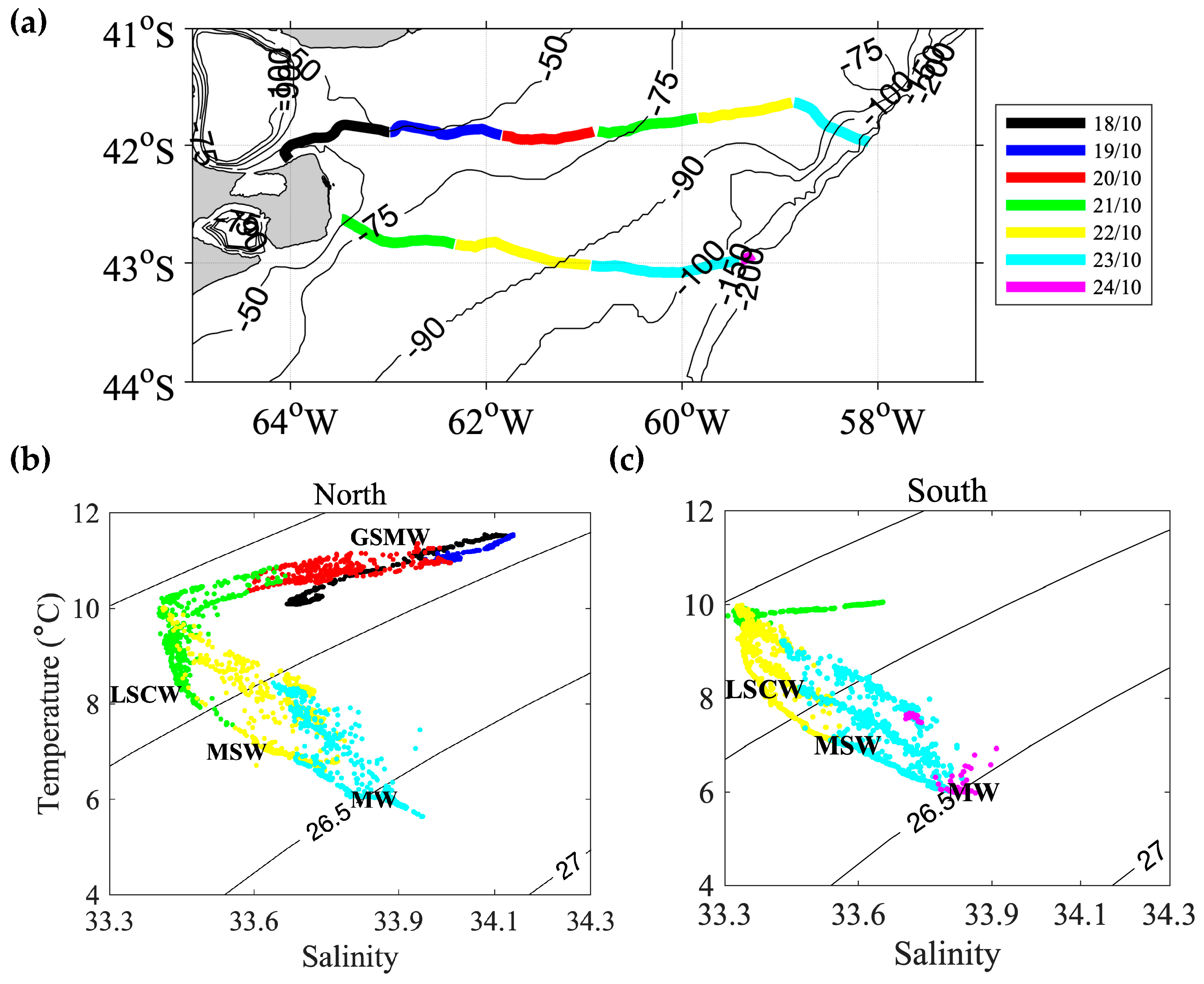
3.5. Water Mass Characterization
3.6. Vertical Profiles of Temperature, Salinity, and Density
3.6.1. Northern Trajectory
3.6.2. Southern Trajectory
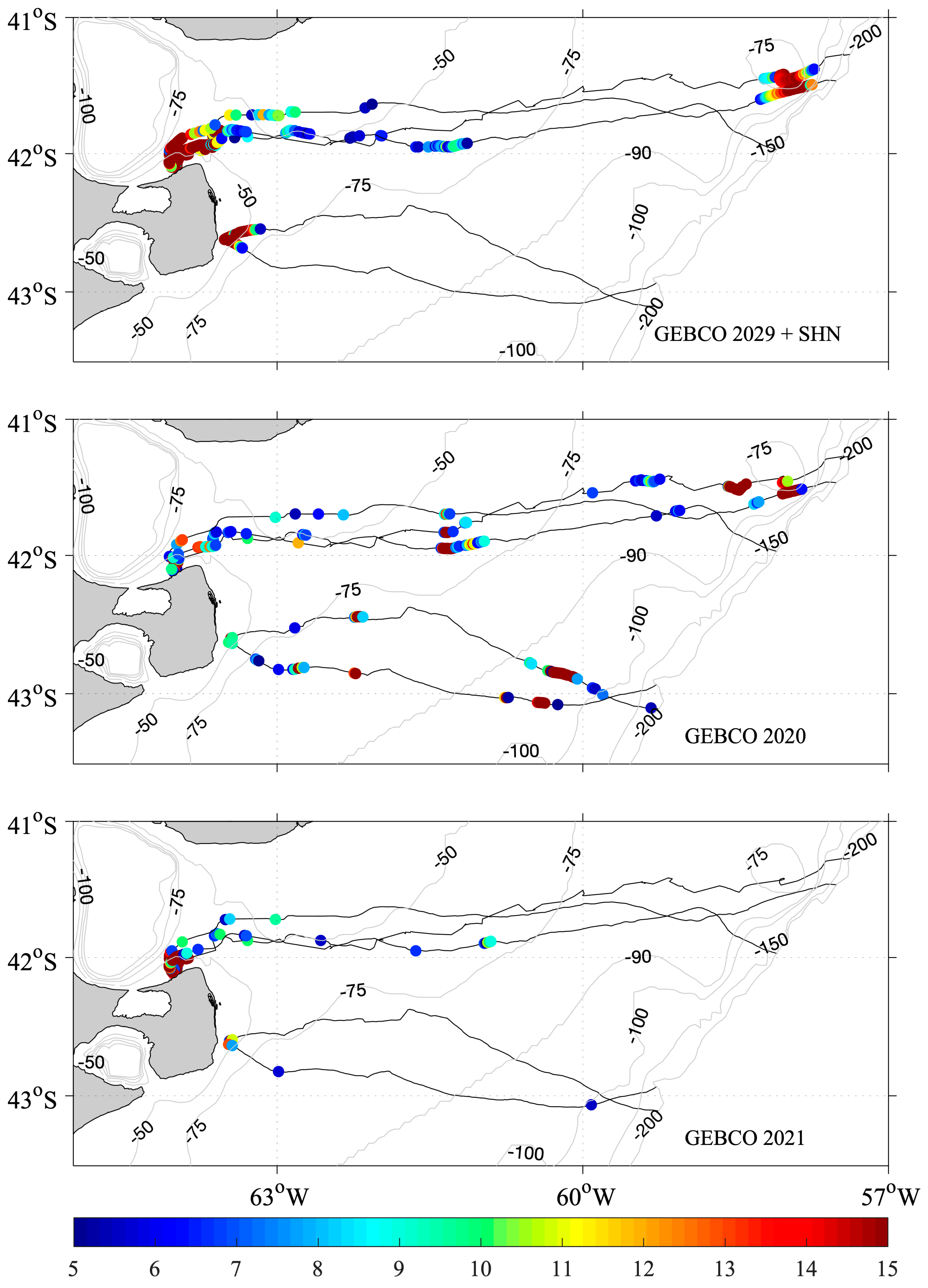
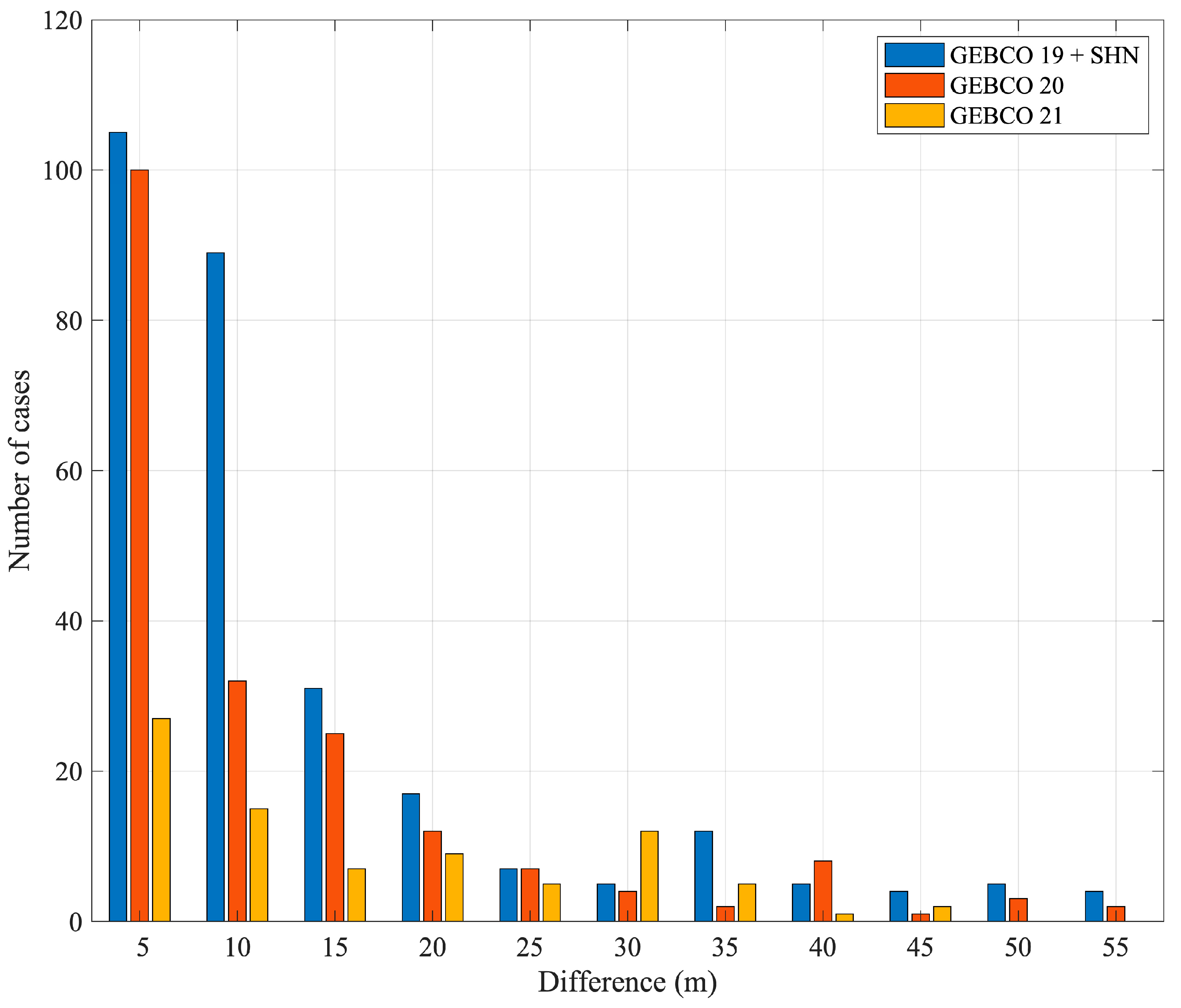
3.7. Comparison between Depths Reached by SESs and GEBCO Bathymetric Charts
4. Summary and Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harcourt, R.; Sequeira, A.M.M.; Zhang, X.; Roquet, F.; Komatsu, K.; Heupel, M.; McMahon, C.; Whoriskey, F.; Meekan, M.; Carroll, G.; et al. Animal-Borne Telemetry: An Integral Component of the Ocean Observing Toolkit. Front. Mar. Sci. 2019, 6, 326. [Google Scholar] [CrossRef]
- Chapman, C.C.; Lea, M.-A.; Meyer, A.; Sallée, J.-B.; Hindell, M. Defining Southern Ocean fronts and their influence on biological and physical processes in a changing climate. Nat. Clim. Chang. 2020, 10, 209–219. [Google Scholar] [CrossRef]
- Treasure, A.M.; Roquet, F.; Ansorge, I.J.; Bester, M.N.; Boehme, L.; Bornemann, H.; Charrassin, J.-B.; Chevallier, D.; Costa, D.P.; Fedak, M.A.; et al. Marine Mammals Exploring the Oceans Pole to Pole: A Review of the MEOP Consortium. Oceanography 2017, 30, 132–138. [Google Scholar] [CrossRef]
- McIntyre, T.; de Bruyn, P.J.N.; Ansorge, I.J.; Bester, M.N.; Bornemann, H.; Plötz, J.; Tosh, C.A. A lifetime at depth: Vertical distribution of southern elephant seals in the water column. Polar Biol. 2010, 33, 1037–1048. [Google Scholar] [CrossRef]
- Boehme, L.; Lovell, P.; Biuw, M.; Roquet, F.; Nicholson, J.; Thorpe, S.E.; Meredith, M.P.; Fedak, M. Technical Note: Animal-borne CTD-Satellite Relay Data Loggers for real-time oceanographic data collection. Ocean Sci. 2009, 5, 685–695. Available online: www.ocean-sci.net/5/685/2009/ (accessed on 30 June 2021). [CrossRef]
- Roquet, F.; Park, Y.-H.; Guinet, C.; Bailleul, F.; Charrassin, J.-B. Observations of the Fawn Trough Current over the Kerguelen Plateau from instrumented elephant seals. J. Mar. Syst. 2009, 78, 377–393. [Google Scholar] [CrossRef]
- Hindell, M.A.; McMahon, C.R.; Jonsen, I.; Harcourt, R.; Arce, F.; Guinet, C. Inter- and intrasex habitat partitioning in the highly dimorphic southern elephant seal. Ecol. Evol. 2021, 11, 1620–1633. [Google Scholar] [CrossRef]
- Bailleul, F.; Charrassin, J.-B.; Monestiez, P.; Roquet, F.; Biuw, M.; Guinet, C. Successful foraging zones of southern elephant seals from the Kerguelen Islands in relation to oceanographic conditions. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 2169–2181. [Google Scholar] [CrossRef]
- Dragon, A.-C.; Monestiez, P.; Bar-Hen, A.; Guinet, C. Linking foraging behaviour to physical oceanographic structures: Southern elephant seals and mesoscale eddies east of Kerguelen Islands. Prog. Oceanogr. 2010, 87, 61–71. [Google Scholar] [CrossRef]
- Hindell, M.A.; McMahon, C.R.; Bester, M.N.; Boehme, L.; Costa, D.; Fedak, M.A.; Guinet, C.; Herraiz-Borreguero, L.; Harcourt, R.G.; Huckstadt, L.; et al. Circumpolar habitat use in the southern elephant seal: Implications for foraging success and population trajectories. Ecosphere 2016, 7, e01213. [Google Scholar] [CrossRef]
- Campagna, C.; Piola, A.R.; Marin, M.R.; Lewis, M.; Fernández, T. Southern elephant seal trajectories, fronts and eddies in the Brazil/Malvinas Confluence. Deep Sea Res. Part I Oceanogr. Res. Pap. 2006, 53, 1907–1924. [Google Scholar] [CrossRef]
- Campagna, J.; Lewis, M.N.; González Carman, V.; Campagna, C.; Guinet, C.; Johnson, M.; Randall WDavis Rodríguez, D.H.; Hindell, M.A. Ontogenetic niche partitioning in southern elephant seals from Argentine Patagonia. Mar. Mammal Sci. 2020, 37, 631–651. [Google Scholar] [CrossRef]
- Aubone, N.; Saraceno, M.; Alberto, M.T.; Campagna, J.; Le Ster, L.; Picard, B.; Hindell, M.; Campagna, C.; Guinet, C. Physical changes recorded by a deep diving seal on the Patagonian slope drive large ecological changes. J. Mar. Syst. 2021, 223, 103612. [Google Scholar] [CrossRef]
- Campagna, C.; LE Boeuf, B.J.; Blackwell, S.B.; Crocker, D.E.; Quintana, F. Diving behaviour and foraging location of female southern elephant seals from Patagonia. J. Zool. 1995, 236, 55–71. [Google Scholar] [CrossRef]
- McGovern, K.; Rodríguez, D.; Lewis, M.; Eder, E.; Piola, A.; Davis, R. Habitat associations of post-breeding female southern elephant seals (Mirounga leonina) from Península Valdés, Argentina. Deep Sea Res. Part I Oceanogr. Res. Pap. 2022, 185, 103789. [Google Scholar] [CrossRef]
- Campagna, C.; Lewis, M. Growth and Distribution of A Southern Elephant Seal Colony. Mar. Mammal Sci. 1992, 8, 387–396. [Google Scholar] [CrossRef]
- Laws, R.M. History and present status of southern elephant seal populations. In Elephant Seals: Population ecology, Behavior and Physiology; Le Boeuf, B.J., Laws, R.M., Eds.; University of California Press: Berkeley, CA, USA, 1994; pp. 49–65. [Google Scholar]
- Campagna, C.; Piola, A.R.; Marin, M.R.; Lewis, M.; Zajaczkovski, U.; Fernández, T. Deep divers in shallow seas: Southern elephant seals on the Patagonian shelf. Deep Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1792–1814. [Google Scholar] [CrossRef]
- Piola, A.R.; Avellaneda, N.M.; Guerrero, R.A.; Jardón, F.P.; Palma, E.D.; Romero, S.I. Malvinas-slope water intrusions on the northern Patagonia continental shelf. Ocean Sci. 2010, 6, 345–359. [Google Scholar] [CrossRef]
- Glorioso, P.D. Temperature distribution related to shelf-sea fronts on the Patagonian Shelf. Cont. Shelf Res. 1987, 7, 27–34. [Google Scholar] [CrossRef]
- Sabatini, M.; Martos, P. SCIENTIA MARINA Mesozooplankton features in a frontal area off northern Patagonia (Argentina) during spring 1995 and 1998. Sci. Mar. 1995, 66, 215–232. [Google Scholar] [CrossRef]
- Romero, S.I.; Piola, A.R.; Charo, M.; Garcia, C.A.E. Chlorophyll-a variability off Patagonia based on SeaWiFS data. J. Geophys. Res. Ocean 2006, 111. [Google Scholar] [CrossRef]
- Rivas, A.L.; Pisoni, J.P. Identification, characteristics and seasonal evolution of surface thermal fronts in the Argentinean Continental Shelf. J. Mar. Syst. 2010, 79, 134–143. [Google Scholar] [CrossRef]
- Bogazzi, E.; Baldoni, A.; Rivas, A.; Martos, P.; Reta, R.; Orensanz, J.M.; Lasta, M.; Dell’Arciprete, P.; Werner, F. Spatial correspondence between areas of concentration of Patagonian scallop (Zygochlamys patagonica) and frontal systems in the southwestern Atlantic. Fish. Oceanogr. 2005, 14, 359–376. [Google Scholar] [CrossRef]
- Franco, B.C.; Palma, E.D.; Tonini, M.H. Benthic-pelagic uncoupling between the Northern Patagonian Frontal System and Patagonian scallop beds. Estuar. Coast. Shelf Sci. 2015, 153, 145–155. [Google Scholar] [CrossRef]
- Tonini, M.H.; Palma, E.D.; Pisoni, J.P. Modeling the seasonal circulation and connectivity in the North Patagonian Gulfs, Argentina. Estuar. Coast. Shelf Sci. 2022, 271, 107868. [Google Scholar] [CrossRef]
- Lucas, A.J.; Guerrero, R.A.; Mianzán, H.W.; Acha, E.M.; Lasta, C.A. Coastal oceanographic regimes of the Northern Argentine Continental Shelf (34–43°S). Estuar. Coast. Shelf Sci. 2005, 65, 405–420. [Google Scholar] [CrossRef]
- Piola, A.R.; Scasso, L.M. Circulación en el golfo San Matías. Geoacta 1988, 15, 33–51. [Google Scholar]
- Bianchi, A.A.; Bianucci, L.; Piola, A.R.; Pino, D.R.; Schloss, I.; Poisson, A.; Balestrini, C.F. Vertical stratification and air-sea CO2 fluxes in the Patagonian shelf. J. Geophys. Res. Ocean 2005, 110, 1–10. [Google Scholar] [CrossRef]
- Lusquiños, A.J.; Valdés, A.J. Aportes al Conocimiento de las Masas de Agua del Atlántico Sudoccidental; Rep. H659; Serv. de Hidrografía Nav.: Buenos Aires, Argentina, 1971; p. 48. [Google Scholar]
- Saraceno, M.; Martín, J.; Moreira, D.; Pisoni, J.P.; Tonini, M.H. Physical Changes in the Patagonian Shelf. In Global Change in Atlantic Coastal Patagonian Ecosystems: A Journey Through Time; Helbling, E.W., Narvarte, M.A., González, R.A., Villafañe, V.E., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2022; pp. 43–71. [Google Scholar] [CrossRef]
- Photopoulou, T.; Fedak, M.A.; Matthiopoulos, J.; McConnell, B.; Lovell, P. The generalized data management and collection protocol for Conductivity-Temperature-Depth Satellite Relay Data Loggers. Anim. Biotelemetry 2015, 3, 21. [Google Scholar] [CrossRef]
- Roquet, F.; Boehme, L.; Block, B.; Charrasin, J.B.; Costa, D.; Guinet, C.; Harcourt, R.G.; Hindell, M.A.; Hückstädt, L.A.; McMahon, C.R.; et al. Ocean Observations Using Tagged Animals. Oceanography 2017, 30, 139. [Google Scholar] [CrossRef]
- Goulet, P.; Guinet, C.; Swift, R.; Madsen, P.T.; Johnson, M. A miniature biomimetic sonar and movement tag to study the biotic environment and predator-prey interactions in aquatic animals. Deep Sea Res. Part I Oceanogr. Res. Pap. 2019, 148, 1–11. [Google Scholar] [CrossRef]
- Guinet, C.; Vacquié-Garcia, J.; Picard, B.; Bessigneul, G.; Lebras, Y.; Dragon, A.; Viviant, M.; Arnould, J.; Bailleul, F. Southern elephant seal foraging success in relation to temperature and light conditions: Insight into prey distribution. Mar. Ecol. Prog. Ser. 2014, 499, 285–301. [Google Scholar] [CrossRef]
- Vacquié-Garcia, J.; Guinet, C.; Laurent, C.; Bailleul, F. Delineation of the southern elephant seal’s main foraging environments defined by temperature and light conditions. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 113, 145–153. [Google Scholar] [CrossRef]
- Merchant, N.D.; Fristrup, K.M.; Johnson, M.P.; Tyack, P.L.; Witt, M.J.; Blondel, P.; Parks, S.E. Measuring acoustic habitats. Methods Ecol. Evol. 2015, 6, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Lopez, R.; Malardé, J.-P.; Danès, P.; Gaspar, P. Improving Argos Doppler location using multiple-model smoothing. Anim. Biotelemetry 2015, 3, 32. [Google Scholar] [CrossRef]
- Dragon, A.-C.; Bar-Hen, A.; Monestiez, P.; Guinet, C. Horizontal and vertical movements as predictors of foraging success in a marine predator. Mar. Ecol. Prog. Ser. 2012, 447, 243–257. [Google Scholar] [CrossRef]
- Lowther, A.D.; Lydersen, C.; Fedak, M.A.; Lovell, P.; Kovacs, K.M. The Argos-CLS Kalman Filter: Error Structures and State-Space Modelling Relative to Fastloc GPS Data. PLoS ONE 2015, 10, e0124754. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.G.; Petersen, S.L.; Peters, G. GPS tracking a marine predator: The effects of precision, resolution and sampling rate on foraging tracks of African Penguins. Mar. Biol. 2004, 145, 215–223. [Google Scholar] [CrossRef]
- Le Bras, Y.; Jouma’a, J.; Picard, B.; Guinet, C. How Elephant Seals (Mirounga leonina) Adjust Their Fine Scale Horizontal Movement and Diving Behaviour in Relation to Prey Encounter Rate. PLoS ONE 2016, 11, e0167226. [Google Scholar] [CrossRef]
- Siegelman, L.; Roquet, F.; Mensah, V.; Rivière, P.; Pauthenet, E.; Picard, B.; Guinet, C. Correction and Accuracy of High- and Low-Resolution CTD Data from Animal-Borne Instruments. J. Atmos. Ocean. Technol. 2019, 36, 745–760. [Google Scholar] [CrossRef]
- Campagna, C.; Fedak, M.A.; McConnell, B.J. Post-Breeding Distribution and Diving Behavior of Adult Male Southern Elephant Seals from Patagonia. J. Mammal. 1999, 80, 1341–1352. Available online: https://academic.oup.com/jmammal/article/80/4/1341/851974 (accessed on 20 April 2022). [CrossRef]
- Roquet, F.; Charrassin, J.-B.; Marchand, S.; Boehme, L.; Fedak, M.; Reverdin, G.; Guinet, C. Delayed-Mode Calibration of Hydrographic Data Obtained from Animal-Borne Satellite Relay Data Loggers. J. Atmos. Ocean. Technol. 2011, 28, 787–801. [Google Scholar] [CrossRef]
- McMahon, C.R.; Field, I.C.; Bradshaw, C.J.; White, G.C.; Hindell, M.A. Tracking and data–logging devices attached to elephant seals do not affect individual mass gain or survival. J. Exp. Mar. Biol. Ecol. 2008, 360, 71–77. [Google Scholar] [CrossRef]
- Chin, T.M. Multi-Resolution Variational Analysis (MRVA): High-resolution data fusion over global surface. AGU Fall Meet. Abstr. 2012, 2012, NG31A-1574. [Google Scholar]
- Valla, D.; Piola, A.R. Evidence of upwelling events at the northern Patagonian shelf break. J. Geophys. Res. Oceans 2015, 120, 7635–7656. [Google Scholar] [CrossRef]
- Lago, L.S.; Saraceno, M.; Martos, P.; Guerrero, R.A.; Piola, A.R.; Paniagua, G.F.; Ferrari, R.; Artana, C.I.; Provost, C. On the Wind Contribution to the Variability of Ocean Currents Over Wide Continental Shelves: A Case Study on the Northern Argentine Continental Shelf. J. Geophys. Res. Oceans 2019, 124, 7457–7472. [Google Scholar] [CrossRef]
- Hersbach, H.; Bell, B.; Berrisford, P.; Horányi, A.; Muñoz-Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Schepers, D.; Simmons, A.; et al. ERA5 Hourly Data on Single Levels from 1940 to Present. Copernicus Climate Change Service (c3s) Climate Data Store (CDS); European Commission: Luxembourg, 2018; p. 224. [Google Scholar] [CrossRef]
- ECMWF. IFS Documentation CY41r2—Part I: Observations; ECMWF: Reading, UK, 2016; p. 223. [Google Scholar]
- ECMWF. IFS Documentation CY41r2—Part II: Data Assimilation; ECMWF: Reading, UK, 2016. [Google Scholar]
- Pisoni, J.P.; Rivas, A.L.; Tonini, M.H. Coastal upwelling in the San Jorge Gulf (Southwestern Atlantic) from remote sensing, modelling and hydrographic data. Estuar. Coast. Shelf Sci. 2020, 245, 106919. [Google Scholar] [CrossRef]
- Dinápoli, M.G.; Simionato, C.G.; Moreira, D. Nonlinear Interaction Between the Tide and the Storm Surge with the Current due to the River Flow in the Río de la Plata. Estuaries Coasts 2021, 44, 939–959. [Google Scholar] [CrossRef]
- Egbert, G.D.; Erofeeva, S.Y. Efficient Inverse Modeling of Barotropic Ocean Tides. J. Atmos. Ocean. Technol. 2002, 19, 183–204. [Google Scholar] [CrossRef]
- Lago, L.S.; Saraceno, M.; Ruiz-Etcheverry, L.A.; Passaro, M.; Oreiro, F.A.; Donofrio, E.E.; Gonzalez, R.A. Improved Sea Surface Height From Satellite Altimetry in Coastal Zones: A Case Study in Southern Patagonia. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2017, 10, 3493–3503. [Google Scholar] [CrossRef]
- McDougall, T.J.; Barker, P.M. Getting Started with TEOS-10 and the Gibbs Seawater (GSW) Oceanographic Toolbox; SCOR/IAPSO WG: Arnhem, The Netherlands, 2011. [Google Scholar]
- Simpson, J.H. Simpson_1981s.pdf. Phil. Trans. R. Soc. Lond. 1981, 302, 531–546. [Google Scholar]
- Kahl, L.C.; Bianchi, A.A.; Osiroff, A.P.; Pino, D.R.; Piola, A.R. Distribution of sea-air CO2 fluxes in the Patagonian Sea: Seasonal, biological and thermal effects. Cont. Shelf Res. 2017, 143, 18–28. [Google Scholar] [CrossRef]
- Sabatini, M.; Akselman, R.; Reta, R.; Negri, R.; Lutz, V.; Silva, R.; Segura, V.; Gil, M.; Santinelli, N.; Sastre, A.; et al. Spring plankton communities in the southern Patagonian shelf: Hydrography, mesozooplankton patterns and trophic relationships. J. Mar. Syst. 2012, 94, 33–51. [Google Scholar] [CrossRef]
- Palma, E.D.; Matano, R.P. A numerical study of the Magellan Plume. J. Geophys. Res. Ocean. 2012, 117. [Google Scholar] [CrossRef]
- Tonini, M.H.; Palma, E.D.; Piola, A.R. A numerical study of gyres, thermal fronts and seasonal circulation in austral semi-enclosed gulfs. Cont. Shelf Res. 2013, 65, 97–110. [Google Scholar] [CrossRef]
- Brandhorst, W.; Castello, J.P. Evaluación de los Recursos de Anchoita (Engraulis anchoita) Frente a la Argentina y Uruguay; Proyecto de Desarrollo Pesquero; Ser. Inf. Tee. Publicación: Mar del Plata, Argentina, 1971. [Google Scholar]
- Pisoni, J.P. Los Sistemas Frontales y la Circulación en las Inmediaciones de los Golfos Norpatagónicos. Ph.D. Thesis, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, Buenos Aires, Argentina, 2012. Available online: https://hdl.handle.net/20.500.12110/tesis_n5193_Pisoni (accessed on 12 July 2022).
- Cazau, D.; Bonnel, J.; Jouma’a, J.; le Bras, Y.; Guinet, C. Measuring the Marine Soundscape of the Indian Ocean with Southern Elephant Seals Used as Acoustic Gliders of Opportunity. J. Atmos. Ocean. Technol. 2017, 34, 207–223. [Google Scholar] [CrossRef]
- Franco, B.C. Procesos Acoplados Bento-Pelágicos Relacionados con el Establecimiento y Deriva Larval de la Vieira Patagónica (Zygochlamys patagónica) en el Océano Atlántico Sudoeste. Ph.D. Thesis, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, Buenos Aires, Argentina, 2013. Available online: https://hdl.handle.net/20.500.12110/tesis_n5466_Franco (accessed on 15 March 2023).
- McMahon, C.R.; Hindell, M.A.; Charrassin, J.B.; Coleman, R.; Guinet, C.; Harcourt, R.; Labrousse, S.; Raymond, B.; Sumner, M.; Ribeiro, N. Southern Ocean pinnipeds provide bathymetric insights on the East Antarctic continental shelf. Commun. Earth Environ. 2023, 4, 266. [Google Scholar] [CrossRef]
- Franco, B.C.; Ruiz-Etcheverry, L.A.; Marrari, M.; Piola, A.R.; Matano, R.P. Climate Change Impacts on the Patagonian Shelf Break Front. Geophys. Res. Lett. 2022, 49, e2021GL096513. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Id SESs | Head | Back |
|---|---|---|
| 054 | CTD SMRU | |
| 875 | Dtag-4 sonar | CTD SMRU |
| 878 | Dtag-4 sonar | CTD SMRU |
| 905 | Dtag-4 sonar | CTD SMRU |
| 051 | Dtag-4- Acoustic | CTD SMRU |
| Tag Name | Variables Measured | Range | Accuracy | Precision | Frequency | Range |
|---|---|---|---|---|---|---|
| CTD | Temperature | −5 to 35 °C | 0.0005 °C | 0.0001 °C | 0.5 Hz | −5 to 35 °C |
| Geolocation (ARGOS) | 5km | |||||
| Salinity | 0 to 50 | 0.01 | 0.002 | 0.5Hz | 0 to 50 | |
| Pressure | 0 to 2000 dBar | 2 dbar | 0.05 dBar | 0.5Hz | 0 to 2000 dBar | |
| SPOT | Geolocation (ARGOS) | 5 km | ||||
| Sonar | Pressure | 0 to 2000 dBar | 2 dbar | 0.05 dBar | 50 Hz | 0 to 2000 dBar |
| Triaxial acceleration | 0.03 ms -2 | 200 Hz | ||||
| Magnetic field | 0.5 µT | 50 Hz | ||||
| Geolocation (GPS) | 50 m | |||||
| Dtag-4 | Pressure | 0 to 2000 dBar | 2 dbar | 0.05 dBar | 50 Hz | 0 to 2000 dBar |
| Triaxial acceleration | 0.03 ms -2 | 200 Hz | ||||
| Magnetic field | 0.5 µT | 50 Hz | ||||
| Geolocation (GPS) | 50 m | |||||
| Sound (Acoustic sensor) | 20kHz |
| SESs ID | Date and Time of Departure | Date and Time of Arrival at the 200 m Isobath | Mean Number of Dives per Day within the ACS | Number of Days to Cross the ACS | Distance Traveled (km) | Mean Distance Traveled per Day (km) |
|---|---|---|---|---|---|---|
| 054 | 25-October 11:29:54 | 01-November 03:08:00 | 92.8 | 6.7 | 667.2 | 100.3 |
| 878 | 17-October 23:32:56 | 23-October 02:41:46 | 85.4 | 5.1 | 586.8 | 114.4 |
| 875 | 18-October 01:09:10 | 23-October 19:31:48 | 83.1 | 5.8 | 540.4 | 93.7 |
| 905 | 20-October 15:44:22 | 23-October 22:19:10 | 109.0 | 3.4 | 381.1 | 116.4 |
| 051 | 21-October 06:48:48 | 24-October 01:52:00 | 92.7 | 2.8 | 359.4 | 128.6 |
| Mean | 92.6 ± 10.2 | 4.7 ± 1.6 | 507.0 ± 133.0 | 110.1 ± 13.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez, M.M.; Ruiz-Etcheverry, L.A.; Saraceno, M.; Gros-Martial, A.; Campagna, J.; Picard, B.; Guinet, C. Satellite and High-Spatio-Temporal Resolution Data Collected by Southern Elephant Seals Allow an Unprecedented 3D View of the Argentine Continental Shelf. Remote Sens. 2023, 15, 5604. https://doi.org/10.3390/rs15235604
Martinez MM, Ruiz-Etcheverry LA, Saraceno M, Gros-Martial A, Campagna J, Picard B, Guinet C. Satellite and High-Spatio-Temporal Resolution Data Collected by Southern Elephant Seals Allow an Unprecedented 3D View of the Argentine Continental Shelf. Remote Sensing. 2023; 15(23):5604. https://doi.org/10.3390/rs15235604
Chicago/Turabian StyleMartinez, Melina M., Laura A. Ruiz-Etcheverry, Martin Saraceno, Anatole Gros-Martial, Julieta Campagna, Baptiste Picard, and Christophe Guinet. 2023. "Satellite and High-Spatio-Temporal Resolution Data Collected by Southern Elephant Seals Allow an Unprecedented 3D View of the Argentine Continental Shelf" Remote Sensing 15, no. 23: 5604. https://doi.org/10.3390/rs15235604
APA StyleMartinez, M. M., Ruiz-Etcheverry, L. A., Saraceno, M., Gros-Martial, A., Campagna, J., Picard, B., & Guinet, C. (2023). Satellite and High-Spatio-Temporal Resolution Data Collected by Southern Elephant Seals Allow an Unprecedented 3D View of the Argentine Continental Shelf. Remote Sensing, 15(23), 5604. https://doi.org/10.3390/rs15235604