
Topographic Analysis of Intertidal Polychaete Reefs (Sabellaria alveolata) at a Very High Spatial Resolution


Abstract
1. Introduction
2. Materials and Methods
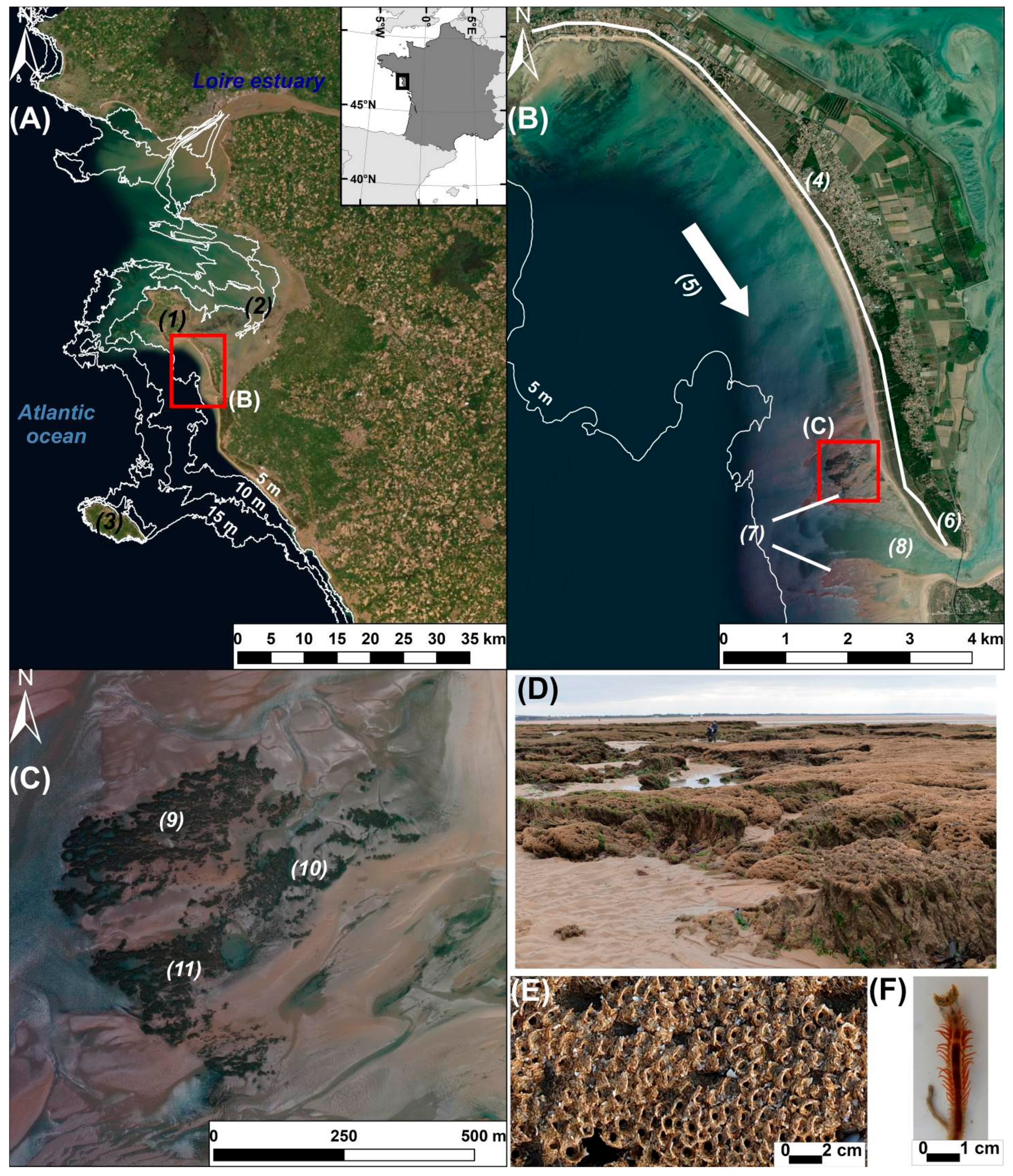
2.1. Study Area
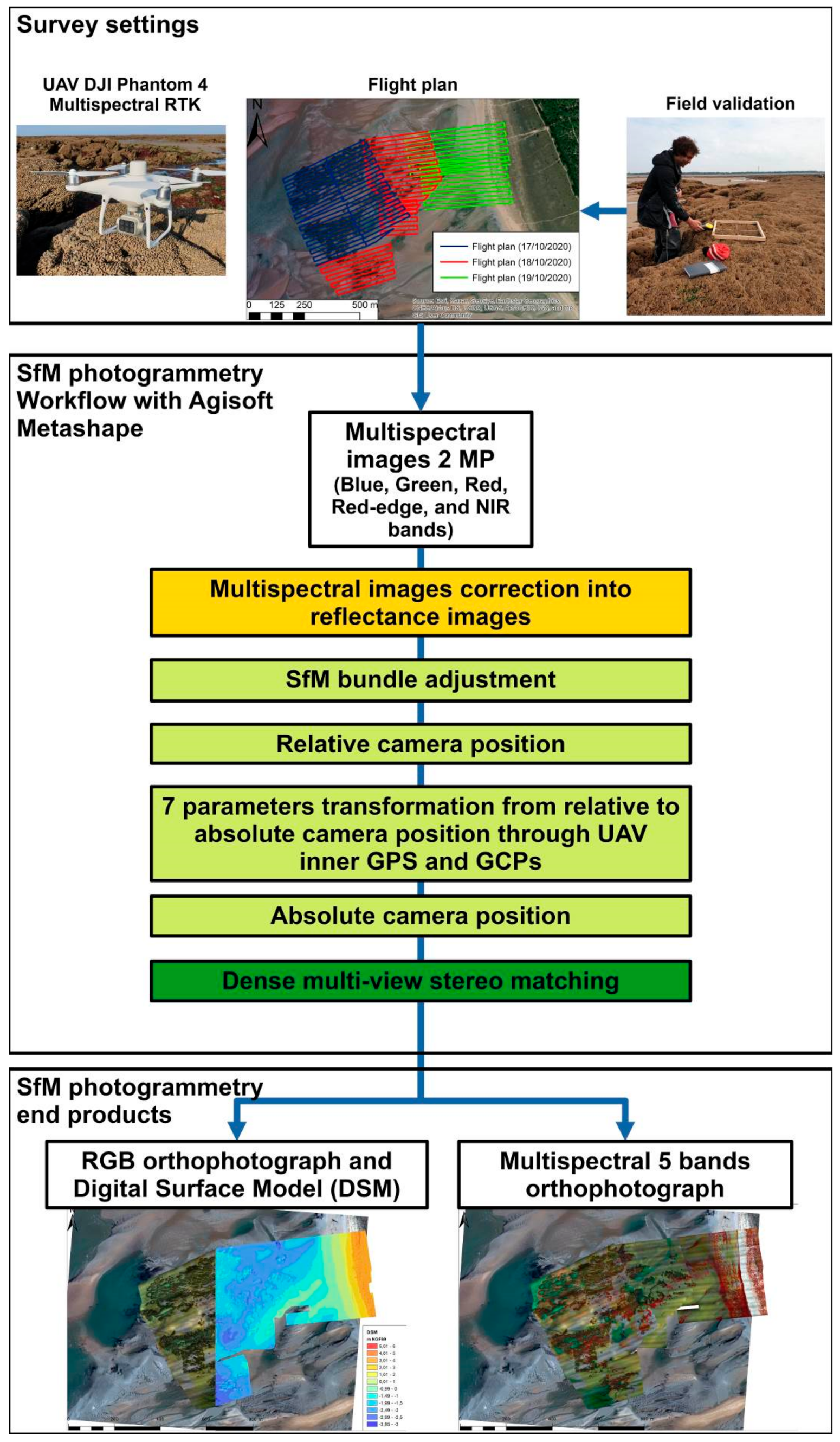
2.2. Survey Settings

2.3. SfM Photogrammetry Workflow
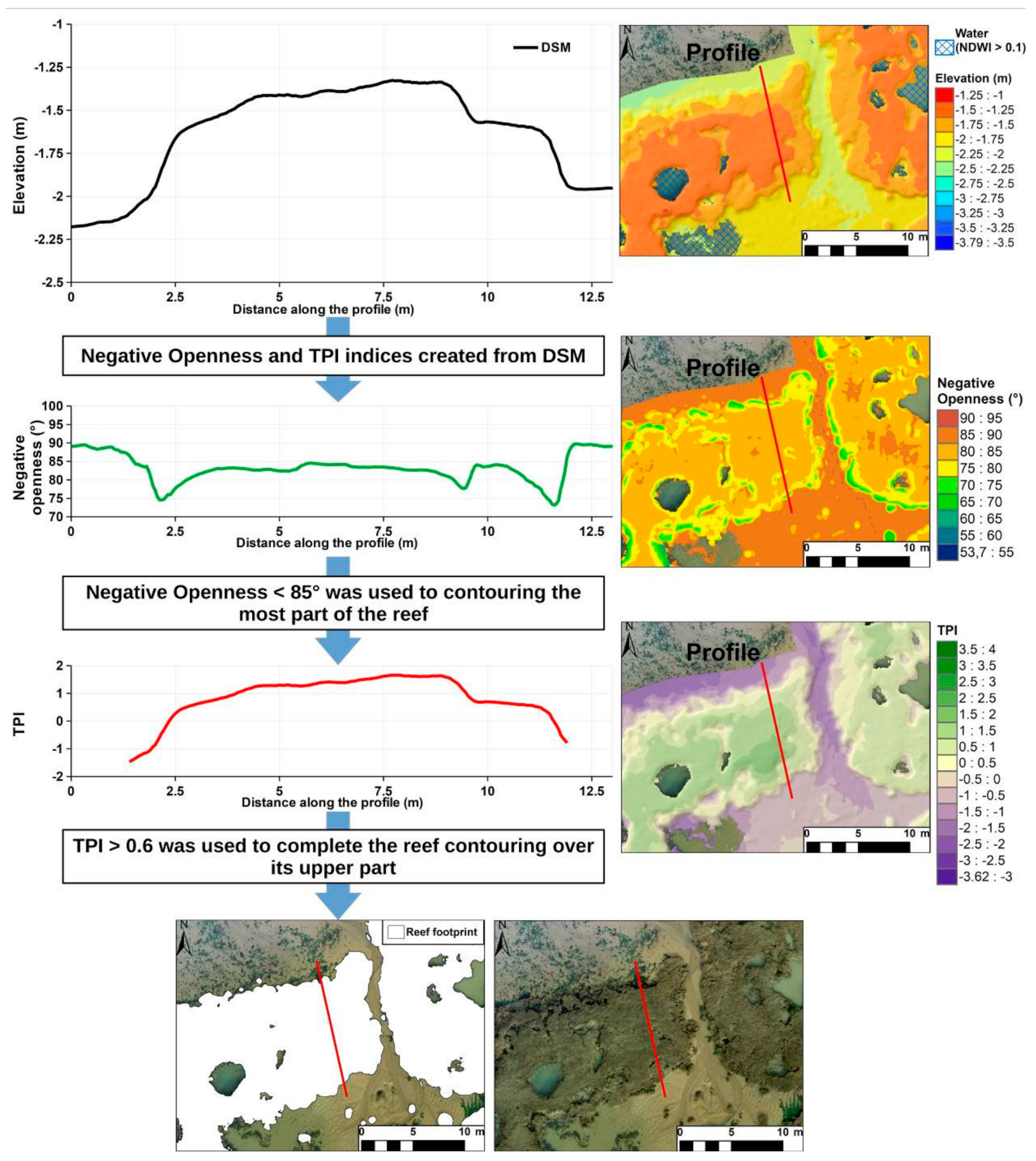
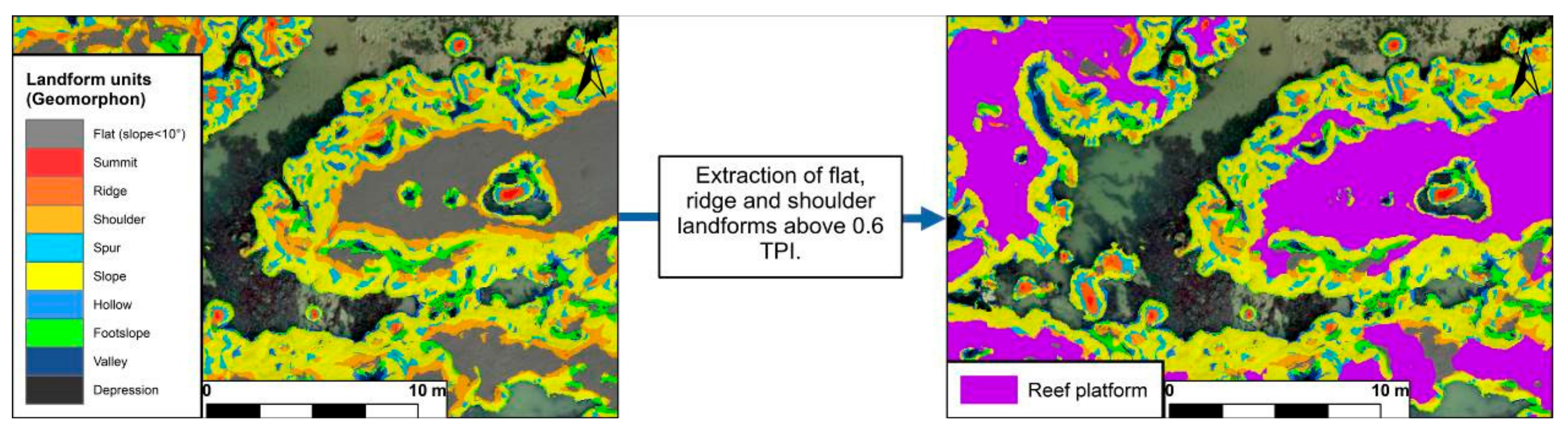
2.4. Reef Contouring Using Topography from DSM
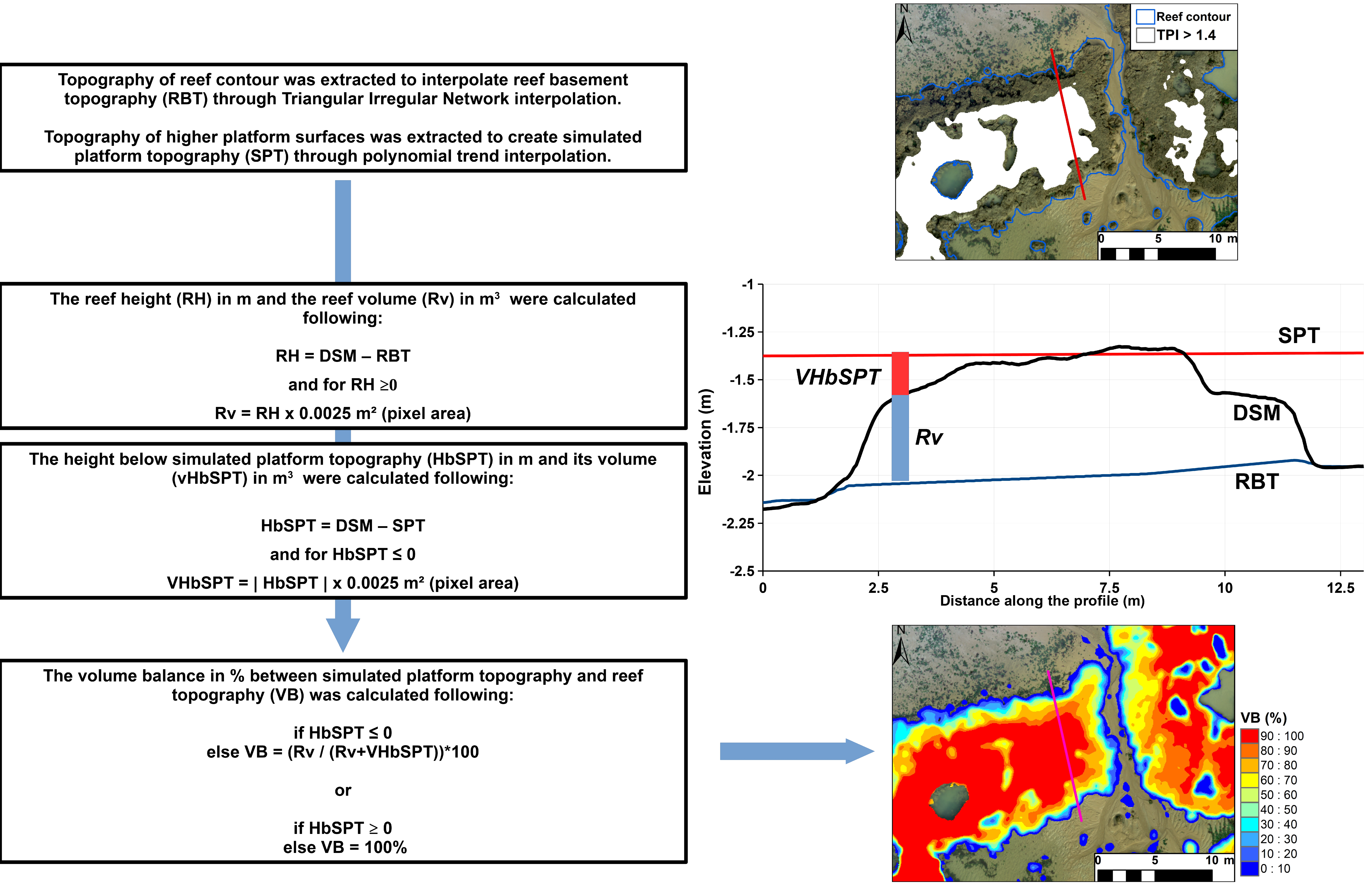
2.5. Reef Morphology Assessment
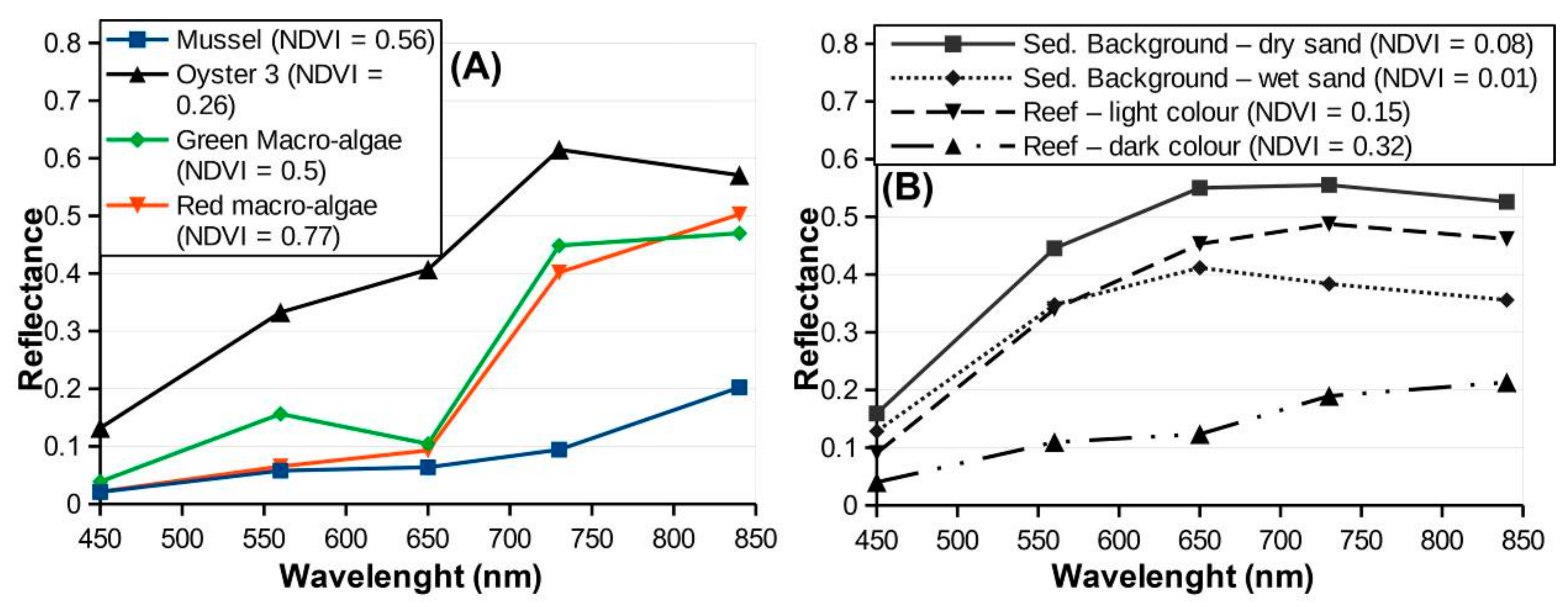
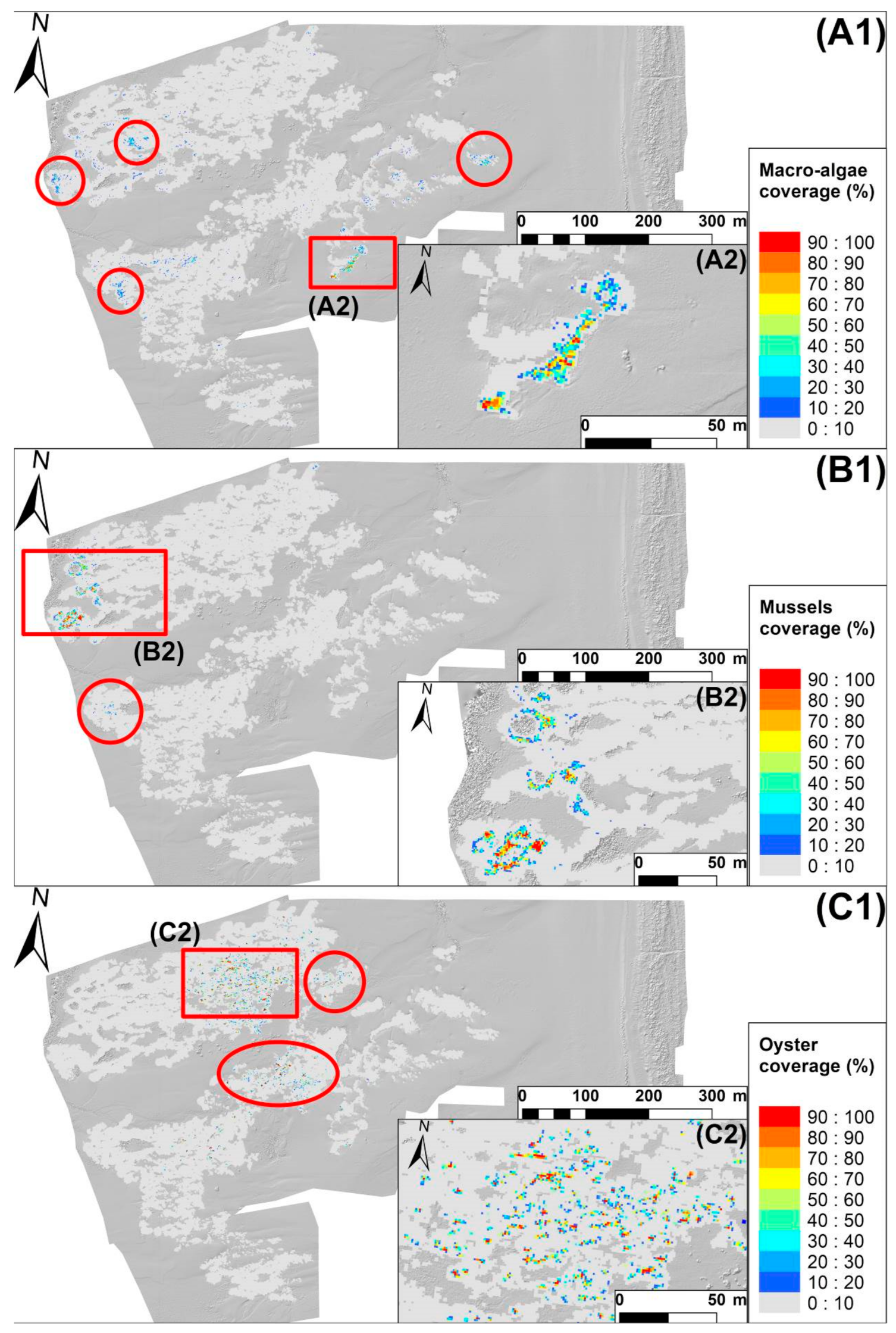
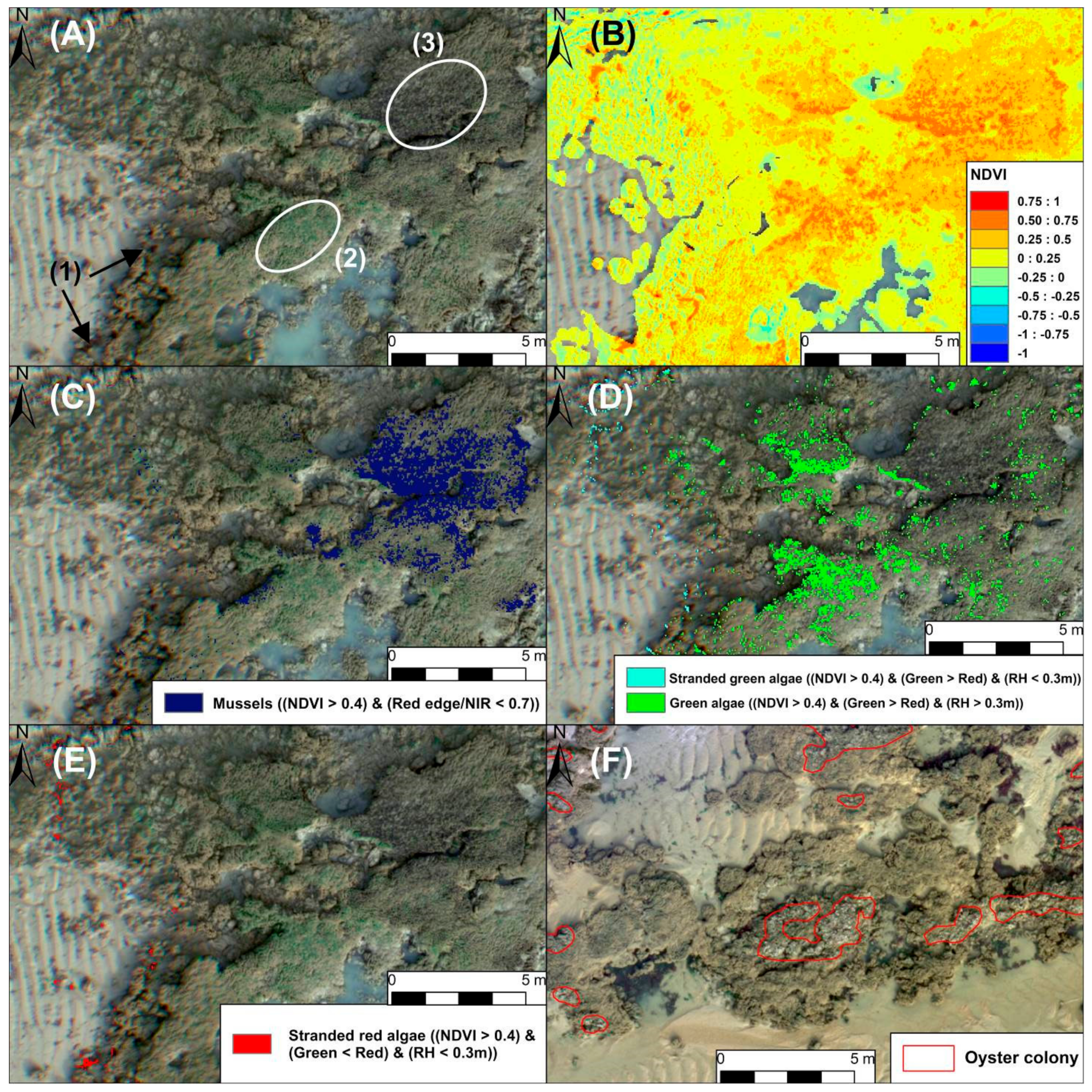
2.6. Reef Epibionts Mapping
3. Results
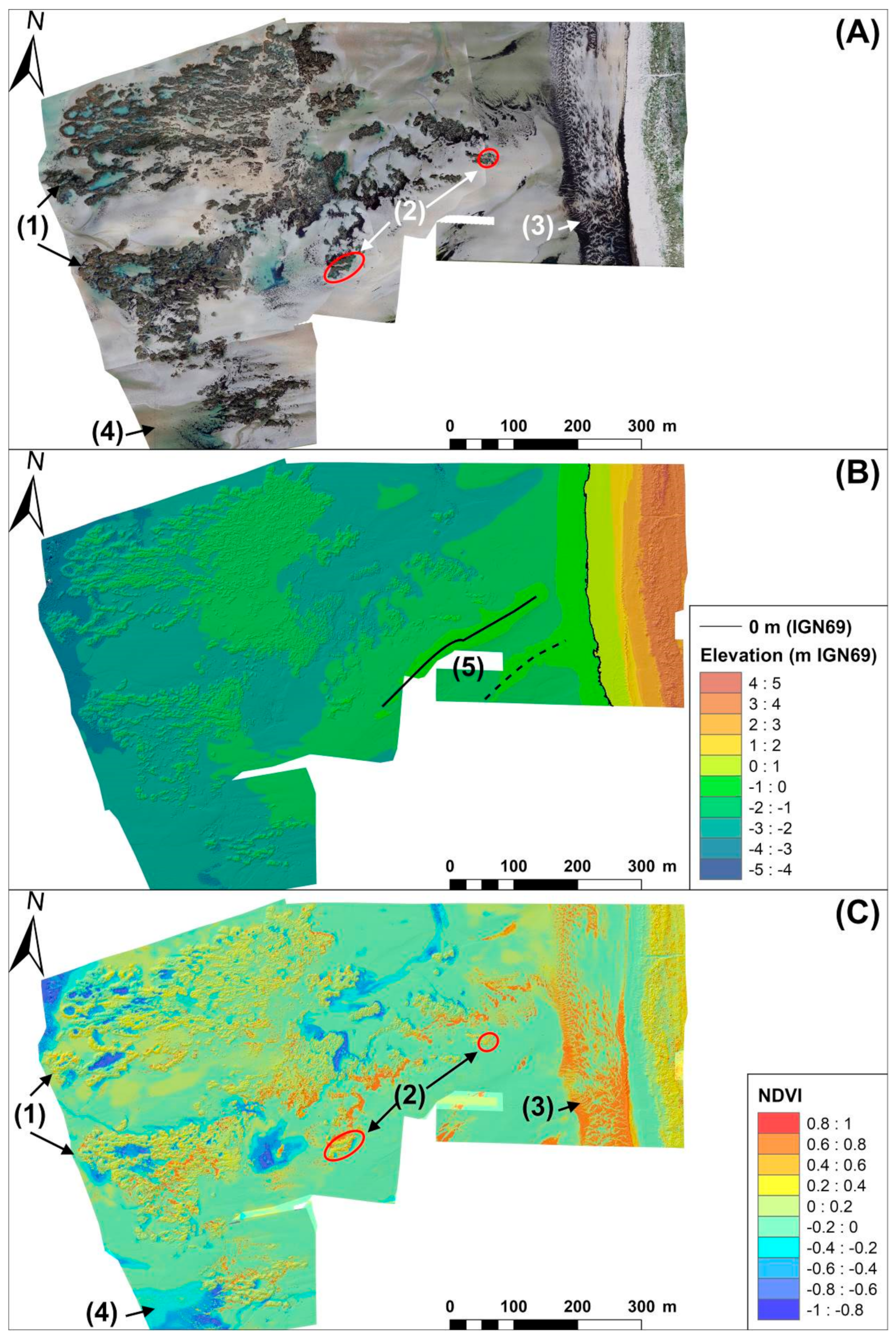
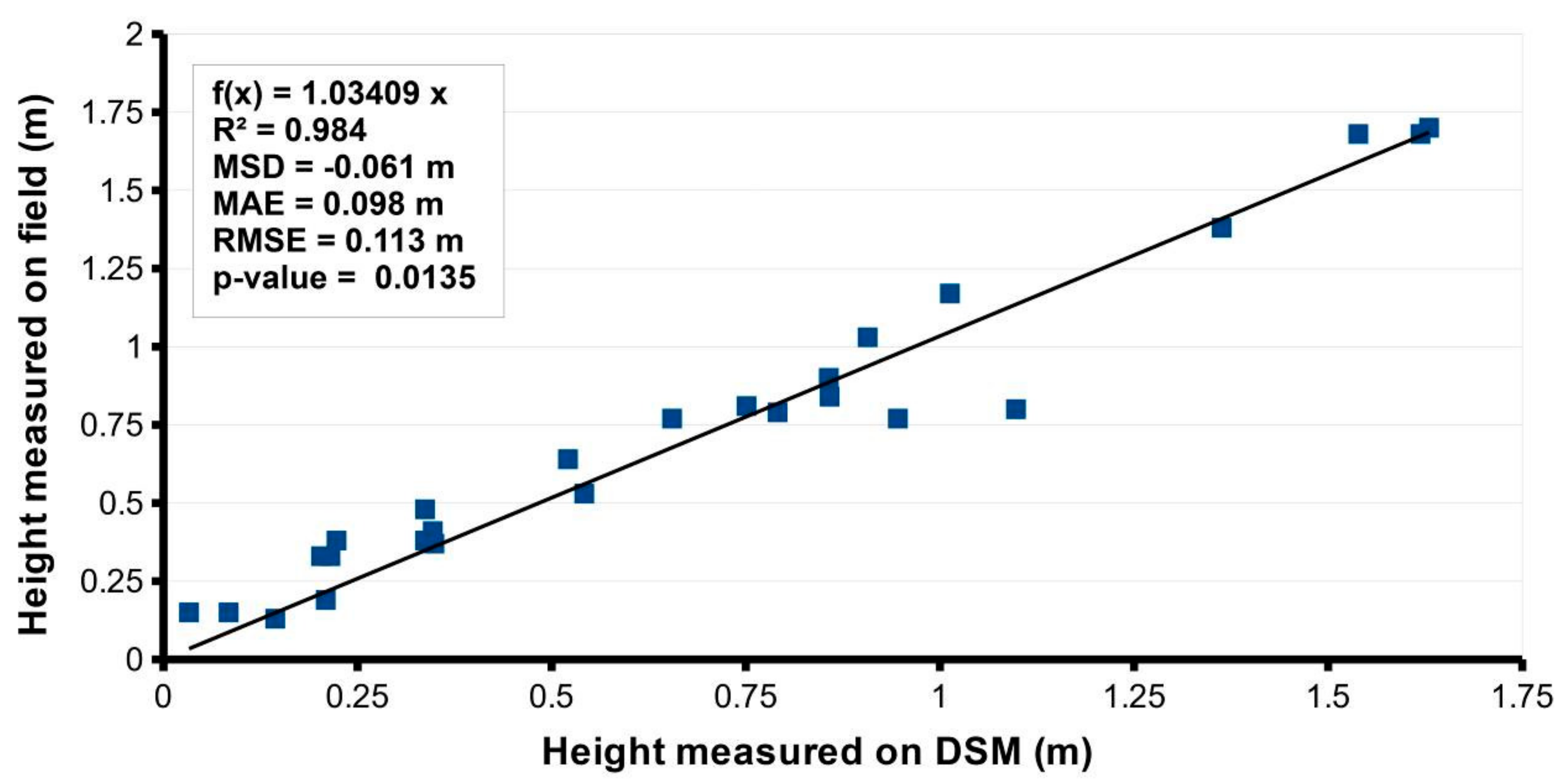
3.1. Multispectral and DSM Accuracy Assessments
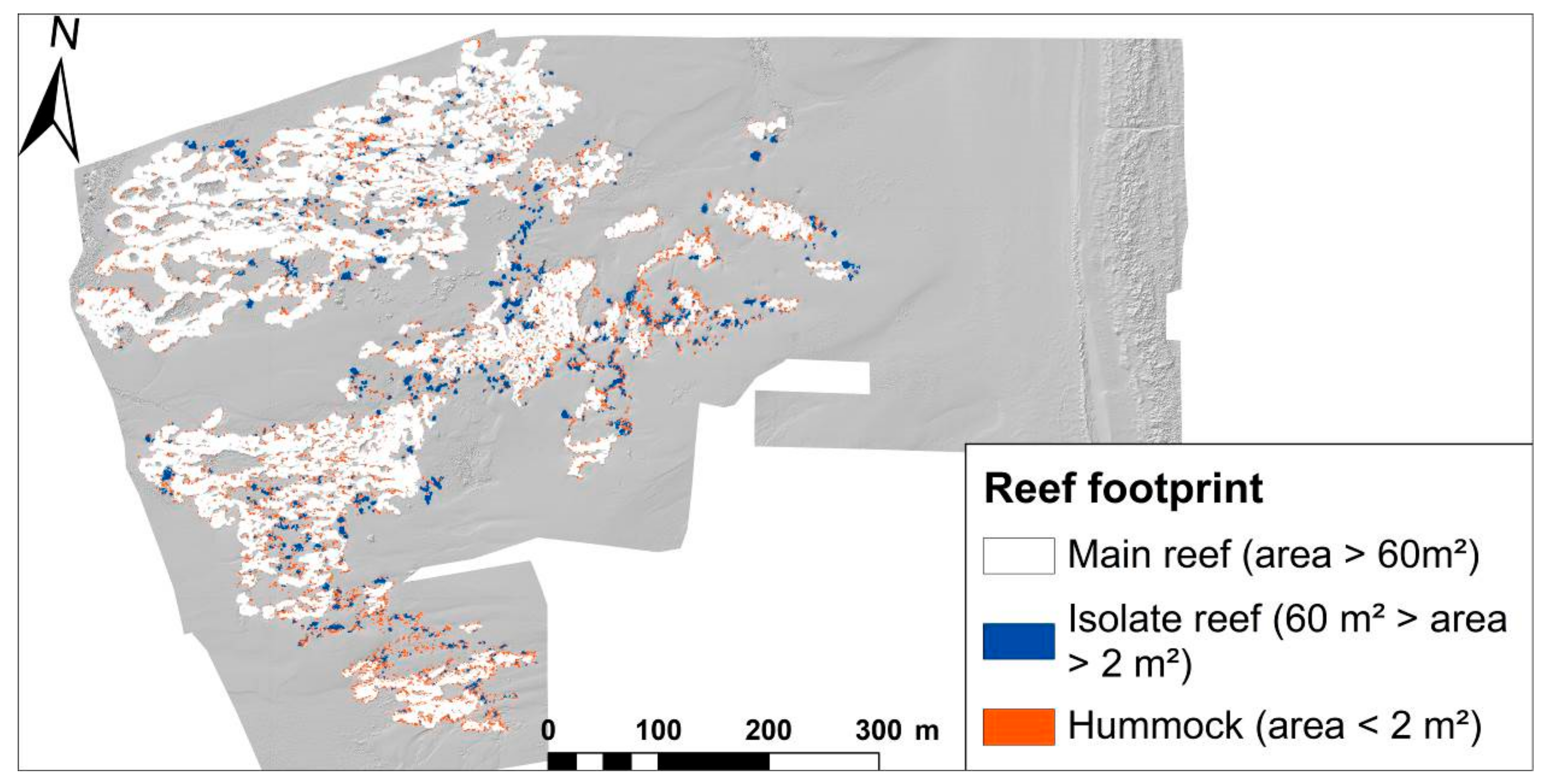
3.2. Reef Contouring


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SfM Photogrammetry End-Product | |
|---|---|
| Number of images per band | 4716 |
| Dense point cloud density | 361 points/m2 |
| Raw multispectral resolution (cm/pixel) | 2.6 cm/pixel |
| Raw DSM resolution (cm/pixel) | 5.26 cm/pixel |
| Field validation | |
| Field Validation number used | 29 |
| Vertical accuracy MSD | −6.1 cm |
| Vertical accuracy MAE | 9.8 cm |
| Vertical accuracy RMSE | 11.3 cm |


3.3. Reef Morphology
3.4. Epibiont Mapping

4. Discussion
4.1. Accuracy and Constraints of UAV Remote Sensing of Reefs
4.2. Identification of the Reef Footprint
4.3. Topographic Analysis
4.4. Epibiont Mapping
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





References
- Curd, A.; Pernet, F.; Corporeau, C.; Delisle, L.; Firth, L.B.; Nunes, F.L.D.; Dubois, S.F. Connecting organic to mineral: How the physiological state of an ecosystem-engineer is linked to its habitat structure. Ecol. Indic. 2019, 98, 49–60. [Google Scholar] [CrossRef]
- Curd, A.; Cordier, C.; Firth, L.B.; Bush, L.; Gruet, Y.; Le Mao, P.; Blaze, J.A.; Board, C.; Bordeyne, F.; Burrows, M.T.; et al. A broad-scale long-term dataset of Sabellaria alveolata distribution and abundance curated through the REEHAB (REEf HABitat) Project. Seanoe 2020, 2. [Google Scholar] [CrossRef]
- Nicoletti, L.; Lattanzi, L.; La Porta, B.; La Valle, P.; Gambi, M.; Tomassetti, P.; Tucci, P.; Chimenz Gusso, C. Sabellaria reefs from the Latium coast (central Tyrrhenian Sea). Biol. Mar. Mediterr. 2001, 8, 252–258. [Google Scholar]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Chapter Three—Mediterranean Bioconstructions along the Italian Coast. In Advances in Marine Biology; Sheppard, C., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 79, pp. 61–136. [Google Scholar]
- Cole, V.J.; Chapman, M.G. Patterns of distribution of annelids: Taxonomic and spatial inconsistencies between two biogeographic provinces and across multiple spatial scales. Mar. Ecol. Prog. Ser. 2007, 346, 235–241. [Google Scholar] [CrossRef]
- Dubois, S.; Retière, C.; Olivier, F. Biodiversity associated with Sabellaria alveolata (Polychaeta: Sabellariidae) reefs: Effects of human disturbances. J. Mar. Biol. Assoc. UK 2002, 82, 817–826. [Google Scholar] [CrossRef]
- Jones, A.G.; Dubois, S.F.; Desroy, N.; Fournier, J. Interplay between abiotic factors and species assemblages mediated by the ecosystem engineer Sabellaria alveolata (Annelida: Polychaeta). Estuar. Coast. Shelf Sci. 2018, 200, 1–18. [Google Scholar] [CrossRef]
- Gruet, Y. Recherche sur l’écologie des “récifs” d’Hermelles édifiés par l’Annélide Polychète Sabellaria alveolata (Linné). Ph.D. Thesis, Université de Nantes, Nantes, France, 1982. [Google Scholar]
- Bernier, P.; Gruet, Y. Environnement littoral, sédimentation et biodiversité de l’Estran. Île de Noirmoutier; Documents des Laboratoires de Géologie: Lyon, France, 2011; pp. 1–163. [Google Scholar]
- Dubois, S.; Barillé, L.; Cognie, B.; Beninger, P.G. Particle capture and processing mechanisms in Sabellaria alveolata (Polychaeta: Sabellariidae). Mar. Ecol. Prog. Ser. 2005, 301, 159–171. [Google Scholar] [CrossRef]
- Le Cam, J.B.; Fournier, J.; Etienne, S.; Couden, J. The strength of biogenic sand reefs: Visco-elastic behaviour of cement secreted by the tube building polychaete Sabellaria alveolata, Linnaeus, 1767. Estuar. Coast. Shelf Sci. 2011, 91, 333–339. [Google Scholar] [CrossRef]
- Gruet, Y. Aspects morphologiques et dynamiques de constructions de l’annélide polychète Sabellaria alveolata (Linné). Rev. des Trav. l’Institut des pêches Marit. 1972, 36, 131–161. [Google Scholar]
- Desroy, N.; Dubois, S.F.; Fournier, J.; Ricquiers, L.; Le Mao, P.; Guerin, L.; Gerla, D.; Rougerie, M.; Legendre, A. The conservation status of Sabellaria alveolata (L.) (Polychaeta: Sabellariidae) reefs in the Bay of Mont-Saint-Michel. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 462–471. [Google Scholar] [CrossRef]
- Naylor, L.A.; Viles, H.A. A temperate reef builder: An evaluation of the growth, morphology and composition of Sabellaria alveolata (L.) colonies on carbonate platforms in South Wales. Geol. Soc. Spec. Publ. 2000, 178, 9–19. [Google Scholar] [CrossRef]
- Lisco, S.N.; Acquafredda, P.; Gallicchio, S.; Sabato, L.; Bonifazi, A.; Cardone, F.; Corriero, G.; Gravina, M.F.; Pierri, C.; Moretti, M. The sedimentary dynamics of Sabellaria alveolata bioconstructions (Ostia, Tyrrhenian Sea, central Italy). J. Palaeogeogr. 2020, 9, 2. [Google Scholar] [CrossRef]
- Dubois, S.; Commito, J.; Olivier, F.; Retière, C. Effects of epibionts on Sabellaria alveolata (L.) biogenic reefs and their associated fauna in the Bay of Mont Saint-Michel. Estuar. Coast. Shelf Sci. 2006, 68, 635–646. [Google Scholar] [CrossRef]
- Schlund, E.; Basuyaux, O.; Lecornu, B.; Pezy, J.; Baffreau, A.; Dauvin, J.C. Macrofauna associated with temporary sabellaria alveolata reefs on the west coast of Cotentin (France). Springerplus 2016, 5, 1–21. [Google Scholar] [CrossRef]
- Jones, A.G.; Denis, L.; Fournier, J.; Desroy, N.; Duong, G.; Dubois, S.F. Linking multiple facets of biodiversity and ecosystem functions in a coastal reef habitat. Mar. Environ. Res. 2020, 162, 105092. [Google Scholar] [CrossRef]
- Muller, A.; Poitrimol, C.; Nunes, F.L.D.; Boyé, A.; Curd, A.; Desroy, N.; Firth, L.B.; Bush, L.; Davies, A.J.; Lima, F.P.; et al. Musical Chairs on Temperate Reefs: Species Turnover and Replacement within Functional Groups Explain Regional Diversity Variation in Assemblages Associated with Honeycomb Worms. Front. Mar. Sci. 2021, 8, 1–18. [Google Scholar] [CrossRef]
- Bonifazi, A.; Lezzi, M.; Ventura, D.; Lisco, S.; Cardone, F.; Gravina, M.F. Macrofaunal biodiversity associated with different developmental phases of a threatened Mediterranean Sabellaria alveolata (Linnaeus, 1767) reef. Mar. Environ. Res. 2019, 145, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Plicanti, A.; Domínguez, R.; Dubois, S.F.; Bertocci, I. Human impacts on biogenic habitats: Effects of experimental trampling on Sabellaria alveolata (Linnaeus, 1767) reefs. J. Exp. Mar. Bio. Ecol. 2016, 478, 34–44. [Google Scholar] [CrossRef]
- Noernberg, M.A.; Fournier, J.; Dubois, S.; Populus, J. Using airborne laser altimetry to estimate Sabellaria alveolata (Polychaeta: Sabellariidae) reefs volume in tidal flat environments. Estuar. Coast. Shelf Sci. 2010, 90, 93–102. [Google Scholar] [CrossRef][Green Version]
- Gruet, Y.; Baudet, J. Les introductions d’espèces d’invertébrés marins. Les Biocénoses Mar. Littorales Françaises des Côtes Atl. 1997, 28, 242–250. [Google Scholar]
- Poursanidis, D.; Traganos, D.; Reinartz, P.; Chrysoulakis, N. On the use of Sentinel-2 for coastal habitat mapping and satellite-derived bathymetry estimation using downscaled coastal aerosol band. Int. J. Appl. Earth Obs. Geoinf. 2019, 80, 58–70. [Google Scholar] [CrossRef]
- Marchand, Y.; Cazoulat, R. Biological reef survey using spot satellite data classification by cellular automata method—Bay of Mont Saint-Michel (France). Comput. Geosci. 2003, 29, 413–421. [Google Scholar] [CrossRef]
- Le Bris, A.; Rosa, P.; Lerouxel, A.; Cognie, B.; Gernez, P.; Launeau, P.; Robin, M.; Barillé, L. Hyperspectral remote sensing of wild oyster reefs. Estuar. Coast. Shelf Sci. 2016, 172, 1–12. [Google Scholar] [CrossRef]
- Bajjouk, T.; Jauzein, C.; Drumetz, L.; Dalla Mura, M.; Duval, A.; Dubois, S.F. Hyperspectral and Lidar: Complementary Tools to Identify Benthic Features and Assess the Ecological Status of Sabellaria alveolata Reefs. Front. Mar. Sci. 2020, 7, 1–16. [Google Scholar] [CrossRef]
- Pan, Z.; Glennie, C.; Fernandez-Diaz, J.C.; Starek, M. Comparison of bathymetry and seagrass mapping with hyperspectral imagery and airborne bathymetric lidar in a shallow estuarine environment. Int. J. Remote Sens. 2016, 37, 516–536. [Google Scholar] [CrossRef]
- Kotilainen, A.; Kaskela, A. Comparison of airborne LiDAR and shipboard acoustic data in complex shallow water environments: Filling in the white ribbon zone. Mar. Geol. 2017, 385, 250–259. [Google Scholar] [CrossRef]
- Duffy, J.P.; Pratt, L.; Anderson, K.; Land, P.E.; Shutler, J.D. Spatial assessment of intertidal seagrass meadows using optical imaging systems and a lightweight drone. Estuar. Coast. Shelf Sci. 2018, 200, 169–180. [Google Scholar] [CrossRef]
- Fairley, I.; Mendzil, A.; Togneri, M.; Reeve, D.E. The use of unmanned aerial systems to map intertidal sediment. Remote Sens. 2018, 10, 1918. [Google Scholar] [CrossRef]
- Windle, A.E.; Poulin, S.K.; Johnston, D.W.; Ridge, J.T. Rapid and accurate monitoring of intertidal Oyster Reef Habitat using unoccupied aircraft systems and structure from motion. Remote Sens. 2019, 11, 394. [Google Scholar] [CrossRef]
- Fallati, L.; Saponari, L.; Savini, A.; Marchese, F.; Corselli, C.; Galli, P. Multi-temporal UAV data and object-based image analysis (OBIA) for estimation of substrate changes in a post-bleaching scenario on a Maldivian reef. Remote Sens. 2020, 12, 2093. [Google Scholar] [CrossRef]
- Donnarumma, L.; D’Argenio, A.; Sandulli, R.; Russo, G.F.; Chemello, R. Unmanned aerial vehicle technology to assess the state of threatened biogenic formations: The vermetid reefs of mediterranean intertidal rocky coasts. Estuar. Coast. Shelf Sci. 2021, 251, 107228. [Google Scholar] [CrossRef]
- Ventura, D.; Bonifazi, A.; Gravina, M.F.; Belluscio, A.; Ardizzone, G. Mapping and classification of ecologically sensitive marine habitats using unmanned aerial vehicle (UAV) imagery and Object-Based Image Analysis (OBIA). Remote Sens. 2018, 10, 1331. [Google Scholar] [CrossRef]
- Collin, A.; Dubois, S.; James, D.; Houet, T. Improving intertidal reef mapping using UAV surface, red edge, and near-infrared data. Drones 2019, 3, 67. [Google Scholar] [CrossRef]
- Collin, A.; Dubois, S.; Ramambason, C.; Etienne, S. Very high-resolution mapping of emerging biogenic reefs using airborne optical imagery and neural network: The honeycomb worm (Sabellaria alveolata) case study. Int. J. Remote Sens. 2018, 39, 5660–5675. [Google Scholar] [CrossRef]
- D’Urban Jackson, T.; Williams, G.J.; Walker-Springett, G.; Davies, A.J. Three-dimensional digital mapping of ecosystems: A new era in spatial ecology. Proc. R. Soc. B Biol. Sci. 2020, 287, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lecours, V.; Espriella, M. Can multiscale roughness help computer-assisted identification of coastal habitats in Florida? In Proceedings of the Geomorphometry 2020 Conference, Perugia, Italy, 22–26 June 2020; pp. 111–114. [Google Scholar]
- Ventura, D.; Dubois, S.F.; Bonifazi, A.; Jona Lasinio, G.; Seminara, M.; Gravina, M.F.; Ardizzone, G. Integration of close-range underwater photogrammetry with inspection and mesh processing software: A novel approach for quantifying ecological dynamics of temperate biogenic reefs. Remote Sens. Ecol. Conserv. 2020, 7, 169–186. [Google Scholar] [CrossRef]
- Guisan, A.; Weiss, S.B.; Weiss, A.D. GLM versus CCA Spatial Modeling of Plant Species Distribution Author(s): Reviewed work (s): GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999, 143, 107–122. [Google Scholar] [CrossRef]
- Yokoyama, R.; Shirasawa, M.; Pike, R.J. Visualizing topography by openness: A new application of image processing to digital elevation models. Photogramm. Eng. Remote Sens. 2002, 68, 257–265. [Google Scholar]
- Jasiewicz, J.; Stepinski, T.F. Geomorphons—A pattern recognition approach to classification and mapping of landforms. Geomorphology 2013, 182, 147–156. [Google Scholar] [CrossRef]
- De Reu, J.; Bourgeois, J.; Bats, M.; Zwertvaegher, A.; Gelorini, V.; De Smedt, P.; Chu, W.; Antrop, M.; De Maeyer, P.; Finke, P.; et al. Application of the topographic position index to heterogeneous landscapes. Geomorphology 2013, 186, 39–49. [Google Scholar] [CrossRef]
- Dubois, S.; Laurent, B.; Retière, C. Efficiency of particle retention and clearance rate in the polychaete Sabellaria alveolata L. C. R. Biol. 2003, 326, 413–421. [Google Scholar] [CrossRef]
- Le Mauff, B. Dynamique hydro-sédimentaire du goulet de Fromentine et des plages adjacentes jusqu’au Pays-de-Monts. Ph.D. Thesis, University of Nantes, Nantes, France, 2018. [Google Scholar]
- Gernez, P.; Doxaran, D.; Barillé, L. Shellfish aquaculture from Space: Potential of Sentinel2 to monitor tide-driven changes in turbidity, chlorophyll concentration and oyster physiological response at the scale of an oyster farm. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Schwartz, M. Encyclopedia of Coastal Science; Schwartz, M.L., Ed.; Encyclopedia of Earth Sciences Series; Springer: New York, NY, USA, 2005; ISBN 1402019033. [Google Scholar]
- Lucieer, A.; de Jong, S.M.; Turner, D. Mapping landslide displacements using Structure from Motion (SfM) and image correlation of multi-temporal UAV photography. Prog. Phys. Geogr. 2014, 38, 97–116. [Google Scholar] [CrossRef]
- Turner, D.; Lucieer, A.; Wallace, L. Direct georeferencing of ultrahigh-resolution UAV imagery. IEEE Trans. Geosci. Remote Sens. 2014, 52, 2738–2745. [Google Scholar] [CrossRef]
- DJI. P4 Multispectral Image Processing Guide P4 Multispectral Image Processing Guide. 2020. Available online: https://dl.djicdn.com/downloads/p4-multispectral/20200717/P4_Multispectral_Image_Processing_Guide_EN.pdf (accessed on 10 November 2021).
- Furukawa, Y.; Ponce, J. Accurate, Dense, and Robust Multiview Stereopsis. IEEE Trans. Pattern Anal. Mach. Intell. 2010, 32, 1362–1376. [Google Scholar] [CrossRef]
- Brunier, G.; Michaud, E.; Fleury, J.; Anthony, E.J.; Morvan, S.; Gardel, A. Assessing the relationship between macro-faunal burrowing activity and mudflat geomorphology from UAV-based Structure-from-Motion photogrammetry. Remote Sens. Environ. 2020, 241, 111717. [Google Scholar] [CrossRef]
- Brunier, G.; Fleury, J.; Anthony, E.J.; Gardel, A.; Dussouillez, P. Close-range airborne Structure-from-Motion Photogrammetry for high-resolution beach morphometric surveys: Examples from an embayed rotating beach. Geomorphology 2016, 261, 76–88. [Google Scholar] [CrossRef]
- James, M.R.; Robson, S. Mitigating systematic error in topographic models derived from UAV and ground-based image networks. Earth Surf. Process. Landf. 2014, 39, 1413–1420. [Google Scholar] [CrossRef]
- James, M.R.; Robson, S. Straightforward reconstruction of 3D surfaces and topography with a camera: Accuracy and geoscience application. J. Geophys. Res. Earth Surf. 2012, 117. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model Dev. Discuss. 2015, 8, 2271–2312. [Google Scholar] [CrossRef]
- Jaud, M.; Grasso, F.; Le Dantec, N.; Verney, R.; Delacourt, C.; Ammann, J.; Deloffre, J.; Grandjean, P. Potential of UAVs for Monitoring Mudflat Morphodynamics (Application to the Seine Estuary, France). ISPRS Int. J. Geo-Inf. 2016, 5, 50. [Google Scholar] [CrossRef]
- McFeeters, S.K. The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Liao, W.H. Region description using extended local ternary patterns. In Proceedings of the International Conference on Pattern Recognition, Istanbul, Turkey, 23–26 August 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 1003–1006. [Google Scholar]
- Fisher, P. Improved modeling of elevation error with Geostatistics. Geoinformatica 1998, 2, 215–233. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; Progress Report RSC 1978-1; 1973, 93p. Available online: https://core.ac.uk/download/pdf/42887948.pdf (accessed on 10 November 2021).
- Hunt, E.R., Jr.; Daughtry, C.S.T.; Eitel, J.U.H.; Long, D.S. Remote Sensing Leaf Chlorophyll Content Using a Visible Band Index. Agron. J. 2011, 103, 1090–1099. [Google Scholar] [CrossRef]
- Fitzgerald, R.W.; Lees, B.G. Assessing the classification accuracy of multisource remote sensing data. Remote Sens. Environ. 1994, 47, 362–368. [Google Scholar] [CrossRef]
- Štroner, M.; Urban, R.; Seidl, J.; Reindl, T.; Brouček, J. Photogrammetry using UAV-mounted GNSS RTK: Georeferencing strategies without GCPs. Remote Sens. 2021, 13, 1336. [Google Scholar] [CrossRef]
- Štroner, M.; Urban, R.; Reindl, T.; Seidl, J.; Brouček, J. Evaluation of the georeferencing accuracy of a photogrammetric model using a quadrocopter with onboard GNSS RTK. Sensors 2020, 20, 2318. [Google Scholar] [CrossRef] [PubMed]
- Forlani, G.; Dall’Asta, E.; Diotri, F.; di Cella, U.M.; Roncella, R.; Santise, M. Quality assessment of DSMs produced from UAV flights georeferenced with on-board RTK positioning. Remote Sens. 2018, 10, 311. [Google Scholar] [CrossRef]
- Taddia, Y.; Stecchi, F.; Pellegrinelli, A. Using dji phantom 4 rtk drone for topographic mapping of coastal areas. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci.-ISPRS Arch. 2019, 42, 625–630. [Google Scholar] [CrossRef]
- Taddia, Y.; Stecchi, F.; Pellegrinelli, A. Coastal mapping using dji phantom 4 RTK in post-processing kinematic mode. Drones 2020, 4, 9. [Google Scholar] [CrossRef]
- Passalacqua, P.; Belmont, P.; Staley, D.M.; Simley, J.D.; Arrowsmith, J.R.; Bode, C.A.; Crosby, C.; DeLong, S.B.; Glenn, N.F.; Kelly, S.A.; et al. Analyzing high resolution topography for advancing the understanding of mass and energy transfer through landscapes: A review. Earth-Sci. Rev. 2015, 148, 174–193. [Google Scholar] [CrossRef]
- Laurent, B.; Mouget, J.-L.; Méléder, V.; Rosa, P.; Jesus, B. Spectral response of benthic diatoms with different sediment backgrounds. Remote Sens. Environ. 2011, 115, 1034–1042. [Google Scholar] [CrossRef]
- Le Hir, P.; Roberts, W.; Cazaillet, O.; Christie, M.; Bassoullet, P.; Bacher, C. Characterization of intertidal flat hydrodynamics. Cont. Shelf Res. 2000, 20, 1433–1459. [Google Scholar] [CrossRef]
- Pianca, C.; Holman, R.; Siegle, E. Mobility of meso-scale morphology on a microtidal ebb delta measured using remote sensing. Mar. Geol. 2014, 357, 334–343. [Google Scholar] [CrossRef]
- Balouin, Y.; Howa, H.; Michel, D. Swash platform morphology in the ebb-tidal delta of the Barra Nova inlet, South Portugal. J. Coast. Res. 2001, 17, 784–791. [Google Scholar]
- Gruet, Y. Spatio-temporal Changes of Sabellarian Reefs Built by the Sedentary Polychaete Sabellaria alveolata (Linné). Mar. Ecol. 1986, 7, 303–319. [Google Scholar] [CrossRef]
- Schmidt, J.; Hewitt, A. Fuzzy land element classification from DTMs based on geometry and terrain position. Geoderma 2004, 121, 243–256. [Google Scholar] [CrossRef]
- Janowski, L.; Wroblewski, R.; Dworniczak, J.; Kolakowski, M.; Rogowska, K.; Wojcik, M.; Gajewski, J. Offshore benthic habitat mapping based on object-based image analysis and geomorphometric approach. A case study from the Slupsk Bank, Southern Baltic Sea. Sci. Total Environ. 2021, 801, 149712. [Google Scholar] [CrossRef] [PubMed]
- Gallant, J.C.; Dowling, T.I. A multiresolution index of valley bottom flatness for mapping depositional areas. Water Resour. Res. 2003, 39. [Google Scholar] [CrossRef]
- Doughty, L.C.; Cavanaugh, C.K. Mapping Coastal Wetland Biomass from High Resolution Unmanned Aerial Vehicle (UAV) Imagery. Remote Sens. 2019, 11, 540. [Google Scholar] [CrossRef]
- Rossiter, T.; Furey, T.; McCarthy, T.; Stengel, D.B. UAV-mounted hyperspectral mapping of intertidal macroalgae. Estuar. Coast. Shelf Sci. 2020, 242, 106789. [Google Scholar] [CrossRef]
- Barillé, L.; Le Bris, A.; Méléder, V.; Launeau, P.; Robin, M.; Louvrou, I.; Ribeiro, L. Photosynthetic epibionts and endobionts of Pacific oyster shells from oyster reefs in rocky versus mudflat shores. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef]
- Chand, S.; Bollard, B. Low altitude spatial assessment and monitoring of intertidal seagrass meadows beyond the visible spectrum using a remotely piloted aircraft system. Estuar. Coast. Shelf Sci. 2021, 255, 107299. [Google Scholar] [CrossRef]
- Espriella, M.C.; Lecours, V.; Frederick, P.C.; Camp, E.V.; Wilkinson, B. Quantifying intertidal habitat relative coverage in a Florida estuary using UAS imagery and GEOBIA. Remote Sens. 2020, 12, 677. [Google Scholar] [CrossRef]
- Nunes, F.L.D.; Van Wormhoudt, A.; Faroni-Perez, L.; Fournier, J. Phylogeography of the reef-building polychaetes of the genus Phragmatopoma in the western Atlantic Region. J. Biogeogr. 2017, 44, 1612–1625. [Google Scholar] [CrossRef]


| CLASS | Mussels | Green Macroalgae on Reef | Starnded Green Macroalgae | Stranded Red Macroalgae | Sum User | Accuracy User (%) |
|---|---|---|---|---|---|---|
| Mussels | 636 | 0 | 0 | 0 | 636 | 100 |
| Green macroalgae on reef | 0 | 930 | 0 | 0 | 930 | 100 |
| Stranded green macroalgae | 0 | 213 | 375 | 0 | 588 | 63.77 |
| Stranded red macroalgae | 2 | 25 | 55 | 401 | 483 | 83.02 |
| Unclassified | 30 | 12 | 13 | 0 | 0 | 0 |
| Sum Producer | 638 | 1168 | 430 | 401 | ||
| Accuracy Producer (%) | 99.68 | 79.62 | 87.21 | 100 | ||
| Overall Accuracy (%) | 88.8 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunier, G.; Oiry, S.; Gruet, Y.; Dubois, S.F.; Barillé, L. Topographic Analysis of Intertidal Polychaete Reefs (Sabellaria alveolata) at a Very High Spatial Resolution. Remote Sens. 2022, 14, 307. https://doi.org/10.3390/rs14020307
Brunier G, Oiry S, Gruet Y, Dubois SF, Barillé L. Topographic Analysis of Intertidal Polychaete Reefs (Sabellaria alveolata) at a Very High Spatial Resolution. Remote Sensing. 2022; 14(2):307. https://doi.org/10.3390/rs14020307
Chicago/Turabian StyleBrunier, Guillaume, Simon Oiry, Yves Gruet, Stanislas F. Dubois, and Laurent Barillé. 2022. "Topographic Analysis of Intertidal Polychaete Reefs (Sabellaria alveolata) at a Very High Spatial Resolution" Remote Sensing 14, no. 2: 307. https://doi.org/10.3390/rs14020307
APA StyleBrunier, G., Oiry, S., Gruet, Y., Dubois, S. F., & Barillé, L. (2022). Topographic Analysis of Intertidal Polychaete Reefs (Sabellaria alveolata) at a Very High Spatial Resolution. Remote Sensing, 14(2), 307. https://doi.org/10.3390/rs14020307