
Genetic and Phylogenetic Characterization of Tataguine and Witwatersrand Viruses and Other Orthobunyaviruses of the Anopheles A, Capim, Guamá, Koongol, Mapputta, Tete, and Turlock Serogroups


 and
and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orthobunyavirus Serogroup Virus Name (Virus Abbreviation) Isolate Designation | Year of Virus Isolation | Country of Virus Isolation (Current Country Name) | Virus Source (Species) | Signs of Human Infection | Reference(s) | New GenBank Accession Numbers |
|---|---|---|---|---|---|---|
| Anopheles A serogroup | ||||||
| Lukuni virus (LUKV) | ||||||
| TRVL 10076 | 1955 | Trinidad and Tobago Crown Colony of the British Empire (Trinidad and Tobago) | Mosquitoes (Aedes (Ochlerotatus) scapularis) | NR | [33] | KP792670-72 |
| Capim serogroup | ||||||
| Capim virus (CAPV) | ||||||
| BeAn 8582 | 1958 | Brazil | Woolly opossum (Caluromys philander) | NR | [34] | KT160026-28 |
| Guajará virus (GJAV) | ||||||
| BeAn 10615 | 1959 | Brazil | Swiss laboratory mouse, sentinel | NR | [34,35] | KP792661-63 |
| Guamá serogroup | ||||||
| Bimiti virus (BIMV) | ||||||
| TRVL 8362 | 1955 | Trinidad and Tobago Crown Colony of the British Empire (Trinidad and Tobago) | Mosquitoes (Culex (Melanoconion) spissipes) | NR | [36,37] | KP792655-57 |
| Catú virus (CATUV) | ||||||
| BeH 151 | 1955 | Brazil | Mosquitoes (Culex (Melanoconion) spissipes) | Fever, myalgia | [38] | KP792658-60 |
| Guamá virus (GMAV) | ||||||
| BeAn 277 | 1955 | Brazil | Tufted capuchin (Cebus apella), sentinel | Fever, myalgia | [38] | KP792664-66 |
| Mahogany Hammock virus (MHV) | ||||||
| Fe4-2a | 1964 | USA | Mosquitoes (Culex (Melanoconion) sp.) | NR | [39] | KP835518-20 |
| Moju virus (MOJUV) | ||||||
| BeAr 12590 | 1959 | Brazil | Mosquitoes (Culex sp.) | NR | [34] | KP792673-75 |
| Koongol serogroup | ||||||
| Koongol virus (KOOV) | ||||||
| MRM31 | 1960 | Australia | Mosquitoes (Culex (Culex) annulirostris) | NR | [40] | KP792667-69 |
| Mapputta serogroup | ||||||
| Mapputta virus (MAPV) | ||||||
| MRM186 | 1960 | Australia | Mosquitoes (Anopheles (Cellia) meraukensis) | NR | [40] | KP792694-96 |
| Trubanaman virus (TRUV) | ||||||
| MRM3630 | 1966 | Australia | Mosquitoes (Anopheles (Cellia) annulipes) | Arthritis, rash | [41,42] | KP792682-84 |
| Tete serogroup | ||||||
| Bahig virus (BAHV) | ||||||
| EgB 90 | 1966 | Egypt | Eurasian golden oriole (Oriolus oriolus) | NR | [43] | KP792652-54 |
| Matruh virus (MTRV) | ||||||
| An-1047 | 1961 | United Arab Republic (Egypt) | Lesser whitethroat (Sylvia curruca) | NR | [43] | KP792691-93 |
| Tete virus (TETEV) | ||||||
| SAAn 3518 | 1959 | Union of South Africa (South Africa) | Village weaver (Ploceus cucullatus) | NR | [44] | KP792679-81 |
| Turlock serogroup | ||||||
| Umbre virus (UMBV) | ||||||
| IG1424 | 1955 | India | Mosquitoes (Culex (Oculeomyia) bitaeniorhynchus) | NR | [44] | KP792685-87 |
| Unassigned | ||||||
| Tataguine virus (TATV) | ||||||
| Ib-H 9963 | 1968 | Nigeria | Human (Homo sapiens) | Fever, myalgia | [45] | KP792676-78 |
| Witwatersrand virus (WITV) | ||||||
| SAAr 1062 | 1958 | Union of South Africa (South Africa) | Mosquitoes (Culex (Eumelanomyia) rubinotus) | NR | [46] | KP792688-90 |
2. Materials and Methods
2.1. Viral Genomic RNA Isolation and Library Preparation
2.2. Bioinformatic and Phylogenetic Analyses
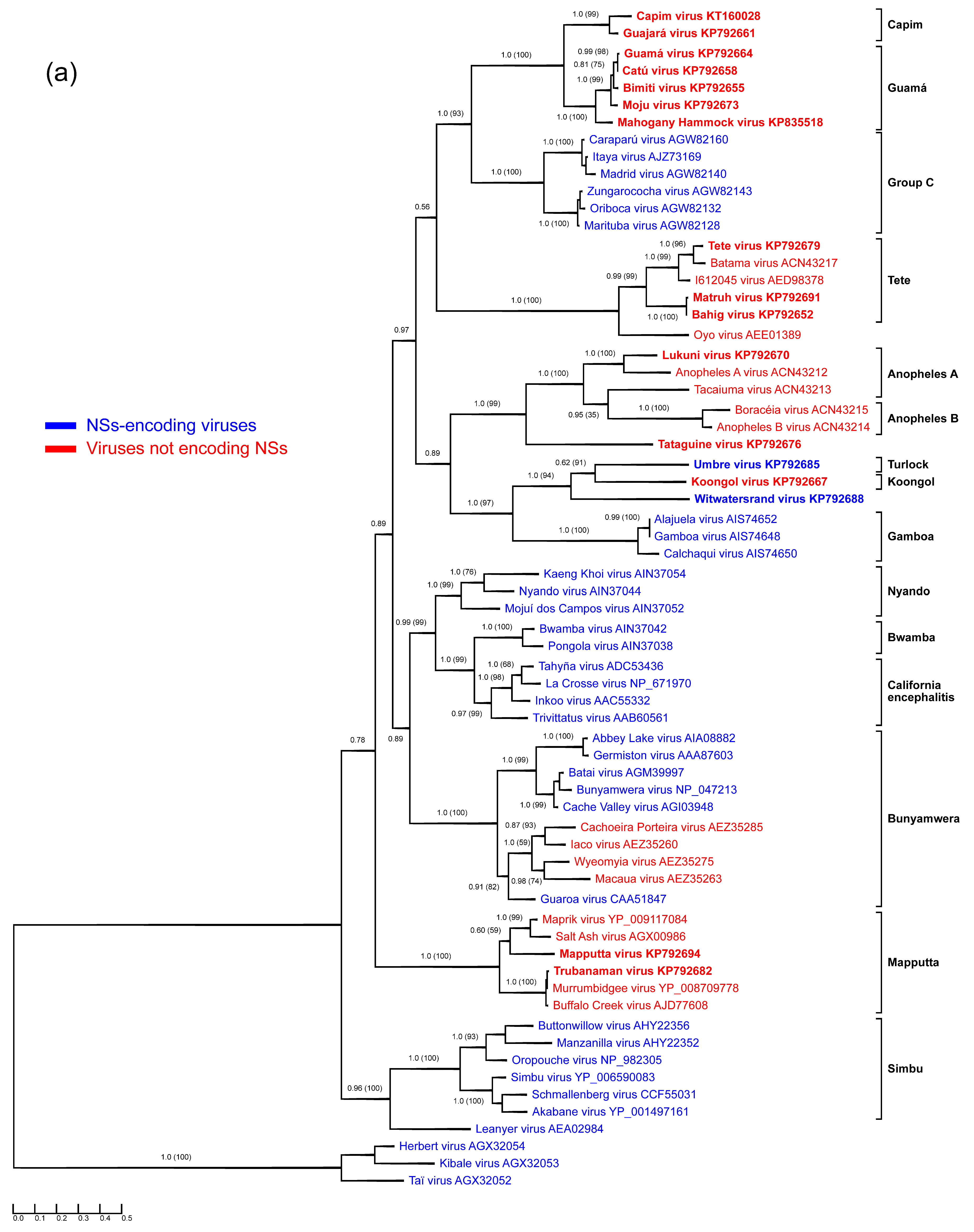
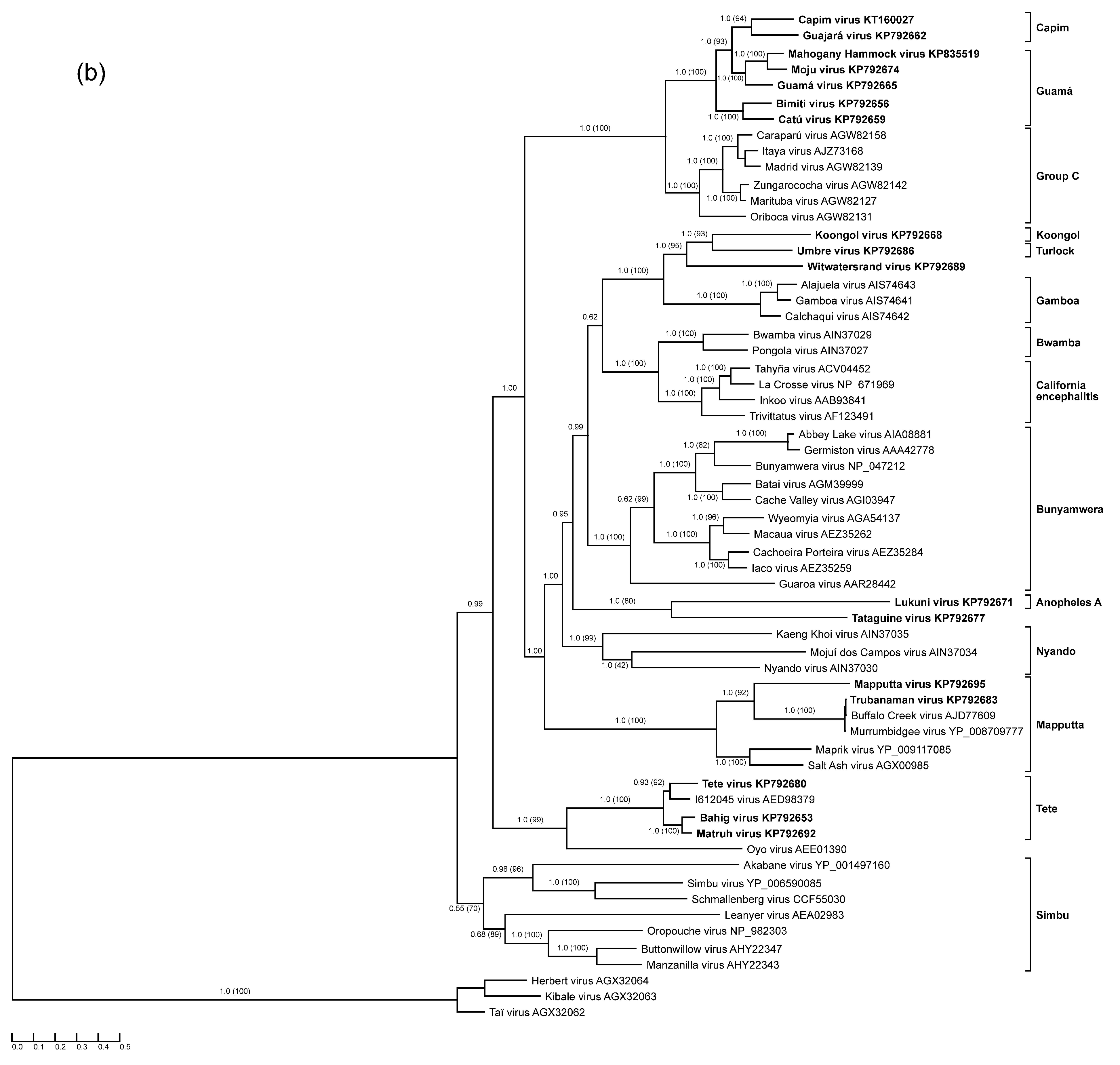
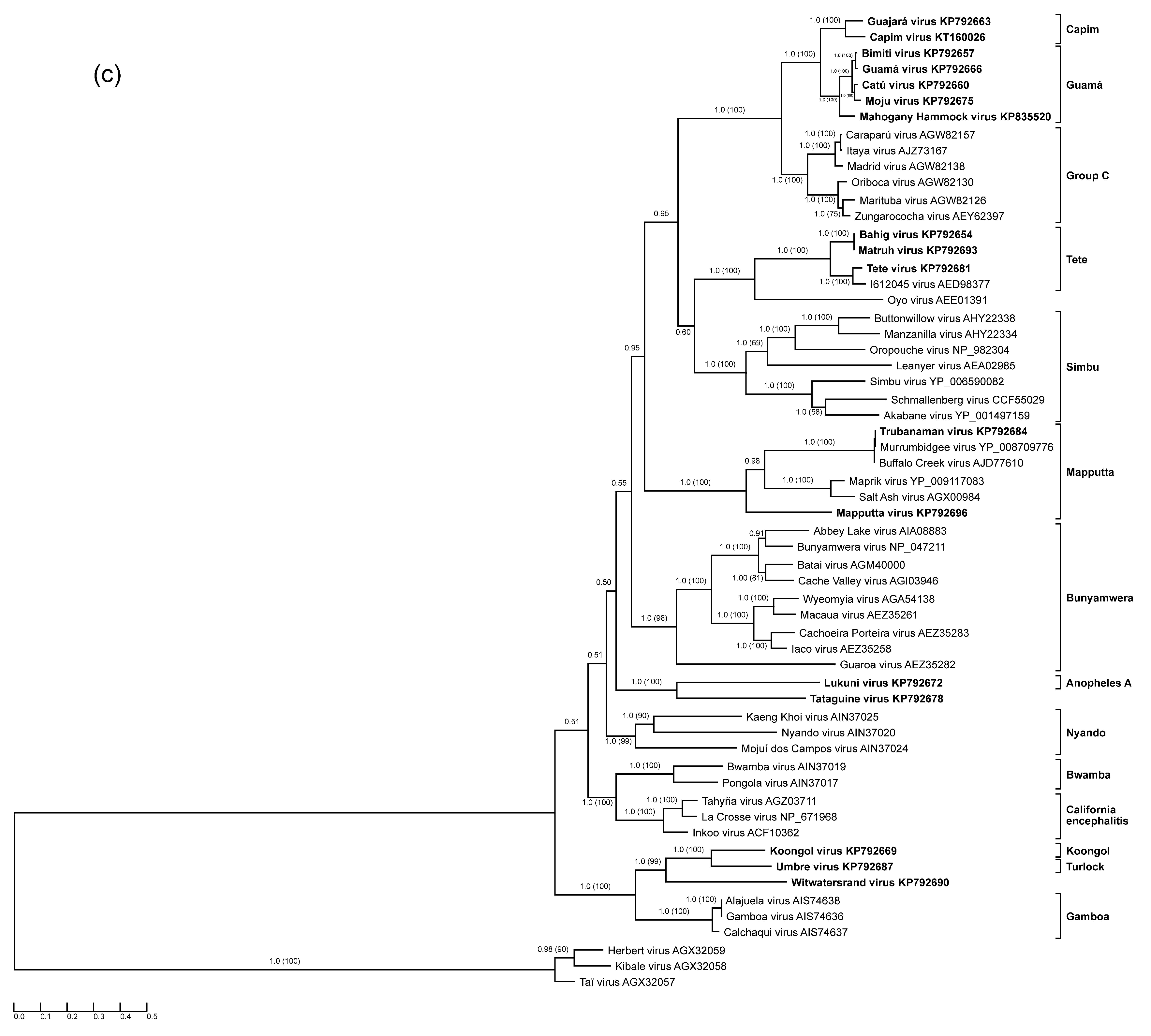
3. Results
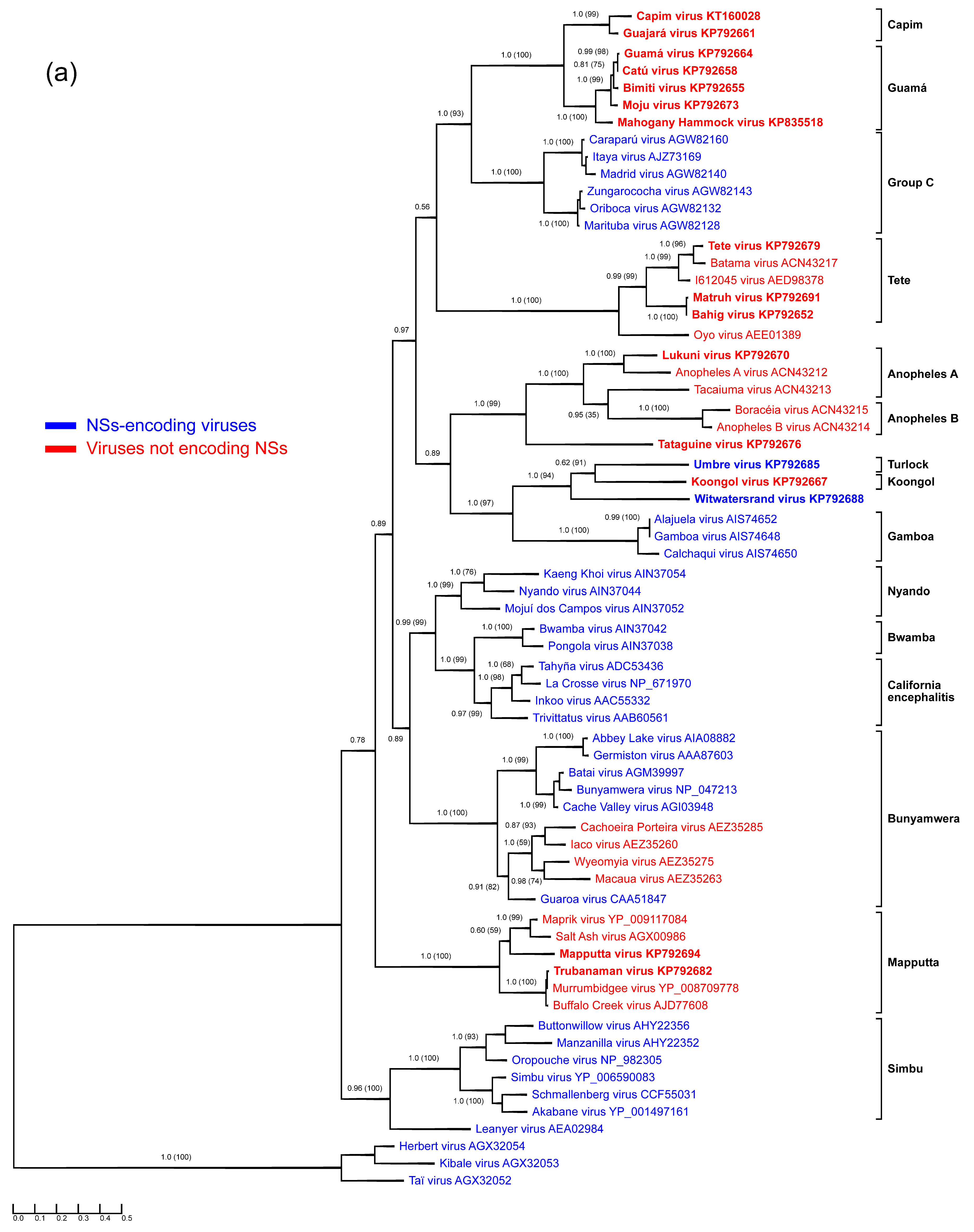
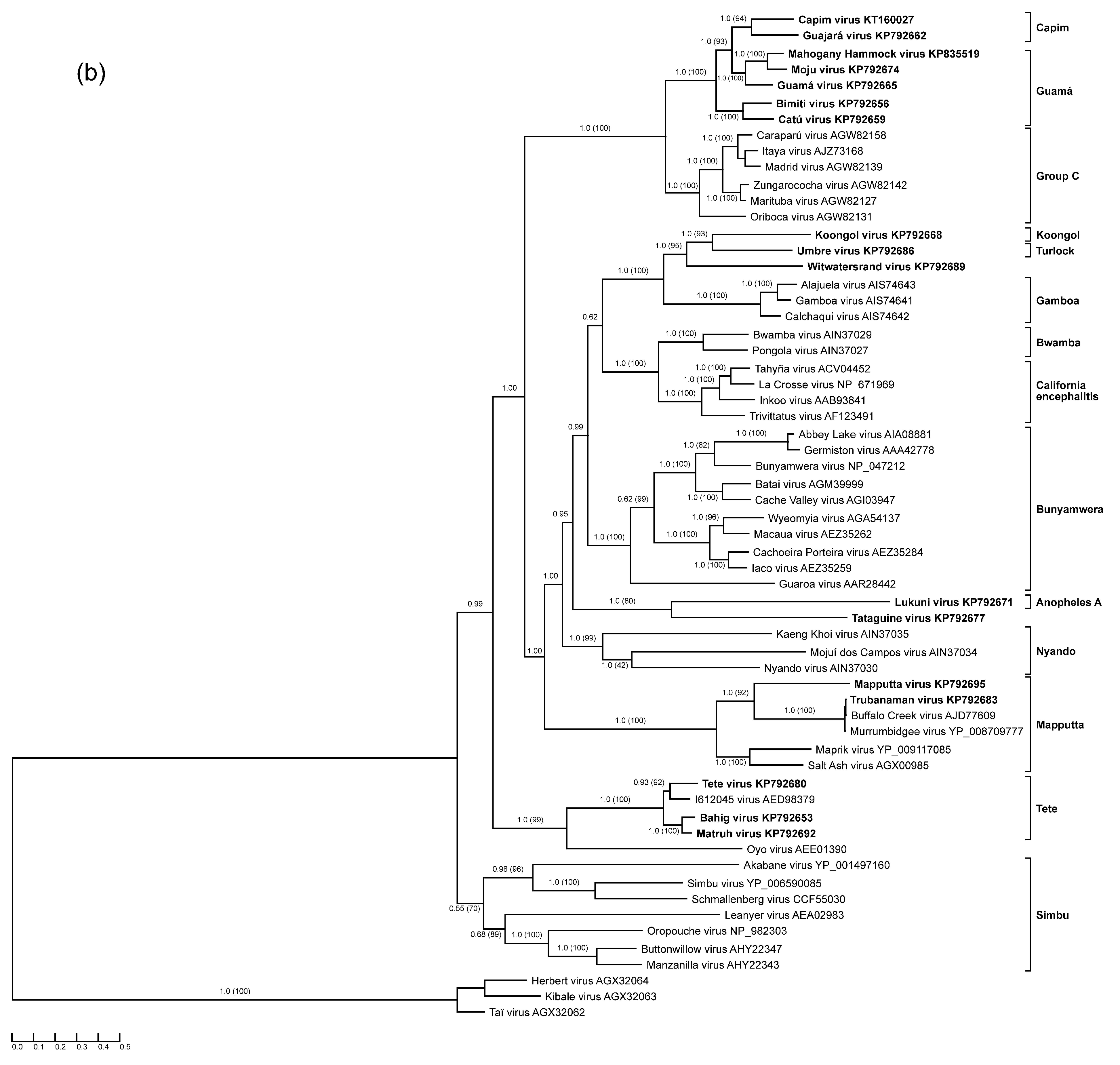
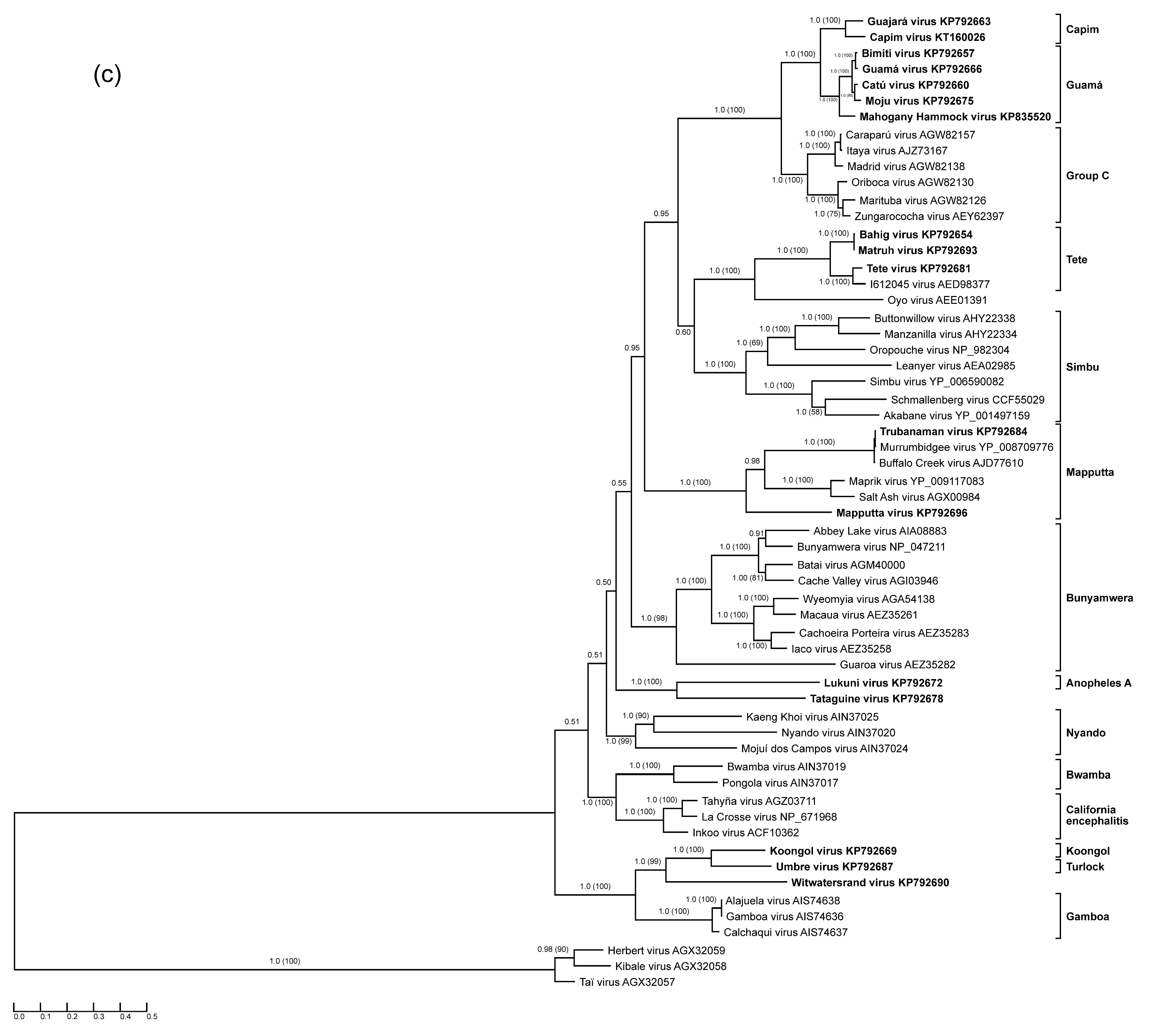
3.1. Anopheles A Serogroup
3.2. Guamá and Capim Serogroups
3.3. Mapputta Serogroup
3.4. Tete Serogroup
3.5. Koongol and Turlock Serogroups
3.6. Tataguine and Witwatersrand Viruses (Unassigned Bunyaviruses)
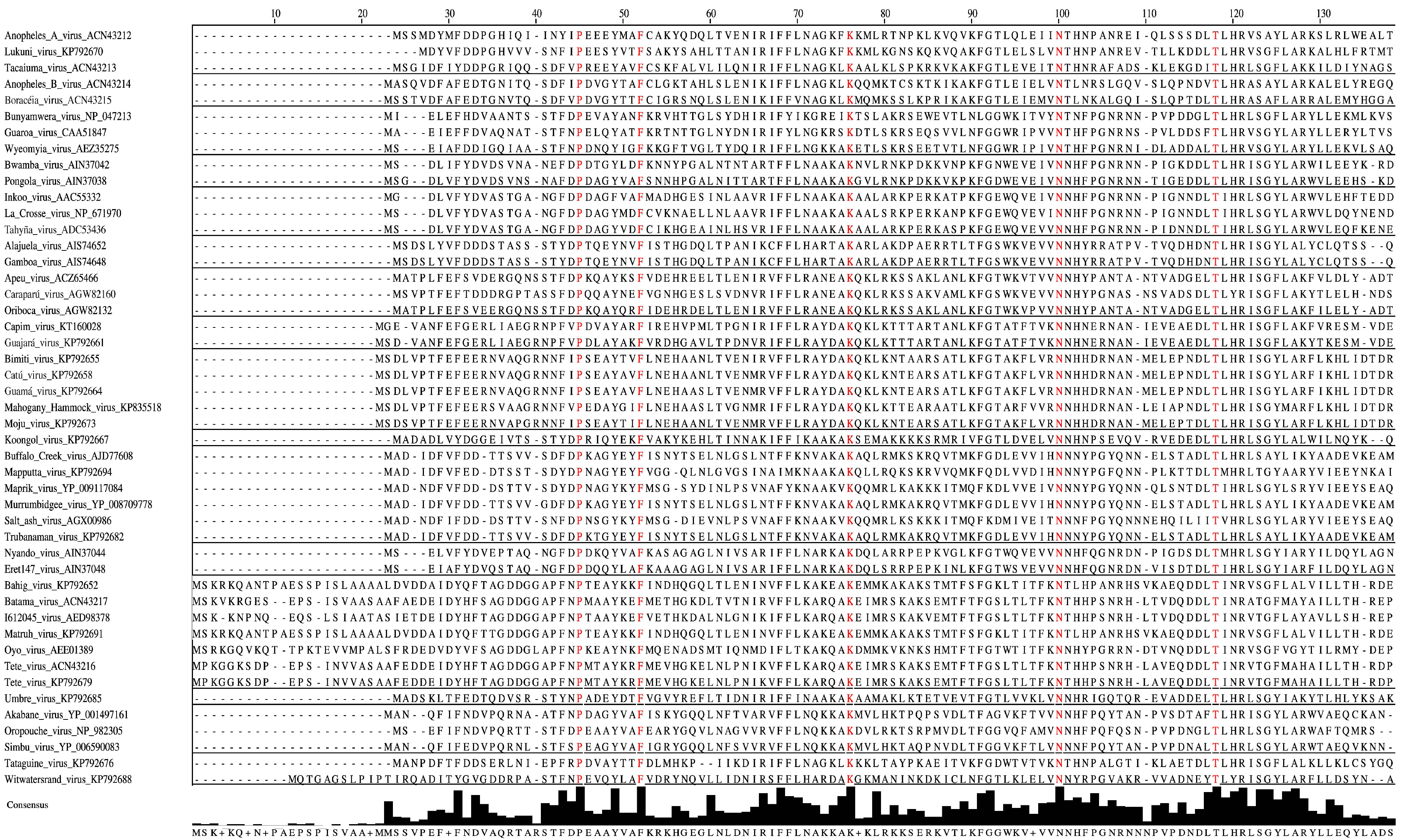
3.7. Characteristics of S Segments: N and NSs proteins
| Orthobunyavirus Serogroup Virus Name (Virus Abbreviation) | N | NSs | Glycoprotein Precursor (# of Cysteines) | Gn | NSm | Gc | L |
|---|---|---|---|---|---|---|---|
| Anopheles A serogroup | |||||||
| Lukuni virus (LUKV) | 242 | - | 1408 (71) | 286 | 168 | 940 | 2241 |
| Capim serogroup | |||||||
| Capim virus (CAPV) | 235 | - | 1430 (74) | 284 | 188 | 947 | 2252 |
| Guajará virus (GJAV) | 235 | - | 1435 (73) | 286 | 188 | 948 | 2252 |
| Guamá serogroup | |||||||
| Bimiti virus (BIMV) | 237 | - | 1443 (73) | 284 | 191 | 954 | 2250 |
| Catú virus (CATUV) | 237 | - | 1440 (72) | 284 | 191 | 952 | 2250 |
| Guamá virus (GMAV) | 237 | - | 1439 (78) | 284 | 187 | 952 | 2250 |
| Mahogany Hammock virus (MHV) | 237 | - | 1436 (75) | 284 | 189 | 946 | 2250 |
| Moju virus (MOJUV) | 237 | - | 1435 (75) | 284 | 189 | 946 | 2250 |
| Koongol serogroup | |||||||
| Koongol virus (KOOV) | 234 | - | 1105 (57) | 284 | 38 | 777 | 2270 |
| Mapputta serogroup | |||||||
| Mapputta virus (MAPV) | 236 | - | 1370 (77) | 288 | 161 | 910 | 2241 |
| Trubanaman virus (TRUV) | 237 | - | 1371 (71) | 286 | 164 | 908 | 2242 |
| Tete serogroup | |||||||
| Bahig virus (BAHV) | 258 | - | 1433 (69) | 286 | 179 | 955 | 2280 |
| Matruh virus (MTRV) | 258 | - | 1433 (67) | 286 | 179 | 955 | 2280 |
| Tete virus (TETEV) | 258 | - | 1432 (68) | 286 | 178 | 955 | 2281 |
| Turlock serogroup | |||||||
| Umbre virus (UMBV) | 237 | 79 | 1466 (72) | 284 | 176 | 991 | 2293 |
| Unassigned | |||||||
| Tataguine virus (TATV) | 239 | - | 1446 (73) | 287 | 171 | 976 | 2246 |
| Witwatersrand virus (WITV) | 245 | 111 | 1448 (70) | 285 | 173 | 974 | 2288 |
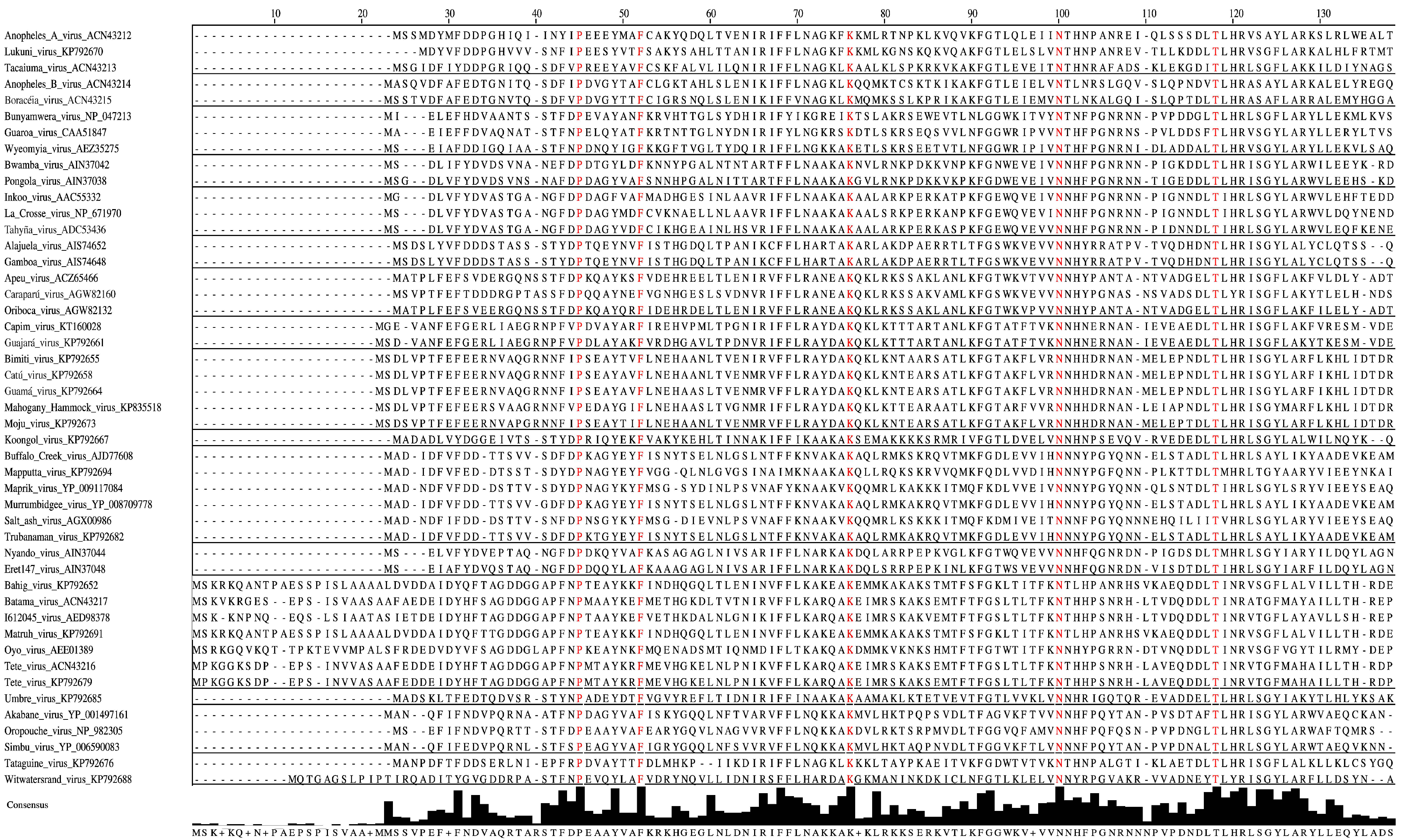

3.8. Characterization of M segments: Glycoprotein Precursor Polyprotein and Gn and Gc Proteins Cleavage Products
3.9. Characterization of L Segments: RNA-Dependent RNA Polymerase
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Whitehouse, C.A.; Kuhn, J.H.; Wada, J.; Ergunay, K. Family Bunyaviridae. In Global Virology I—Identifying and Investigating Viral Diseases; Shapshak, P., Sinnott, J., Somboonwit, C., Kuhn, J.H., Eds.; Springer: New York, USA, 2015; pp. 199–246. [Google Scholar]
- Plyusnin, A.; Beaty, B.J.; Elliott, R.M.; Goldbach, R.; Kormelink, R.; Lundkvist, Å.; Schmaljohn, C.S.; Tesh, R.B. Family Bunyaviridae. In Virus Taxonomy–Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier/Academic Press: London, UK, 2011; pp. 725–741. [Google Scholar]
- Elliott, R.M.; Schmaljohn, C.S. Bunyaviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott/The Williams & Wilkins Company: Philadelphia, PA, USA, 2013; pp. 1244–1282. [Google Scholar]
- Kuhn, J.H.; Peters, C.J. Arthropod-borne and rodent-borne virus infections. In Harrison’s Principles of Internal Medicine, 19th ed.; Kasper, D.L., Fauci, A.S., Hauser, S.L., Longo, D.L., Jameson, J.L., Loscalzo, J., Eds.; McGraw-Hill Education: Columbus, OH, USA, 2015; Volume 2, pp. 1304–1323. [Google Scholar]
- Marklewitz, M.; Zirkel, F.; Rwego, I.B.; Heidemann, H.; Trippner, P.; Kurth, A.; Kallies, R.; Briese, T.; Lipkin, W.I.; Drosten, C.; et al. Discovery of a unique novel clade of mosquito-associated bunyaviruses. J. Virol. 2013, 87, 12850–12865. [Google Scholar] [CrossRef] [PubMed]
- Marklewitz, M.; Zirkel, F.; Kurth, A.; Drosten, C.; Junglen, S. Evolutionary and phenotypic analysis of live virus isolates suggests arthropod origin of a pathogenic rna virus family. Proc. Natl. Acad. Sci. USA 2015, 112, 7536–7541. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M. Orthobunyaviruses: Recent genetic and structural insights. Nat. Rev. Microbiol. 2014, 12, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M.; Blakqori, G. Molecular Biology of Orthobunyaviruses. In Bunyaviridae: Molecular and Cellular Biology; Plyusnin, A., Elliott, R.M., Eds.; Caister Academic Press: Norfolk, UK, 2011; pp. 1–40. [Google Scholar]
- Calisher, C.H. Taxonomy, classification, and geographic distribution of california serogroup bunyaviruses. Prog. Clin. Biol. Res. 1983, 123, 1–16. [Google Scholar] [PubMed]
- Calisher, C.H.; Coimbra, T.L.; Lopez Ode, S.; Muth, D.J.; Sacchetta Lde, A.; Francy, D.B.; Lazuick, J.S.; Cropp, C.B. Identification of new guama and group c serogroup bunyaviruses and an ungrouped virus from southern brazil. Am. J. Trop. Med. Hyg. 1983, 32, 424–431. [Google Scholar] [PubMed]
- Calisher, C.H.; Gutierrez, E.; Francy, D.B.; Alava, A.; Muth, D.J.; Lazuick, J.S. Identification of hitherto unrecognized arboviruses from ecuador: Members of serogroups b, c, bunyamwera, patois, and minatitlan. Am. J. Trop. Med. Hyg. 1983, 32, 877–885. [Google Scholar] [PubMed]
- Calisher, C.H.; Lazuick, J.S.; Wolff, K.L.; Muth, D.J. Antigenic relationships among turlock serogroup bunyaviruses as determined by neutralization tests. Acta Virol. 1984, 28, 148–151. [Google Scholar] [PubMed]
- Kinney, R.M.; Calisher, C.H. Antigenic relationships among simbu serogroup (bunyaviridae) viruses. Am. J. Trop. Med. Hyg. 1981, 30, 1307–1318. [Google Scholar] [PubMed]
- Travassos da Rosa, A.P.; Tesh, R.B.; Pinheiro, F.P.; Travassos da Rosa, J.F.; Peterson, N.E. Characterization of eight new phlebotomus fever serogroup arboviruses (bunyaviridae: Phlebovirus) from the amazon region of brazil. Am. J. Trop. Med. Hyg. 1983, 32, 1164–1171. [Google Scholar] [PubMed]
- Bowen, M.D.; Jackson, A.O.; Bruns, T.D.; Hacker, D.L.; Hardy, J.L. Determination and Comparative Analysis of the Small Rna Genomic Sequences of California Encephalitis, Jamestown Canyon, Jerry Slough, Melao, Keystone and Trivittatus Viruses (Bunyaviridae, Genus Bunyavirus, California Serogroup). J. Gen. Virol. 1995, 76, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Dunn, E.F.; Pritlove, D.C.; Elliott, R.M. The s rna genome segments of batai, cache valley, guaroa, kairi, lumbo, main drain and northway bunyaviruses: Sequence determination and analysis. J. Gen. Virol. 1994, 75, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.R.; Travassos da Rosa, A.P.; Weaver, S.C.; Tesh, R.B.; Vasconcelos, P.F. Molecular epidemiology of group c viruses (Bunyaviridae, orthobunyavirus) isolated in the americas. J. Virol. 2005, 79, 10561–10570. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.F.; Wang, H.; Nunes, M.; Vasconcelos, P.F.; Weaver, S.C.; Shope, R.E.; Watts, D.M.; Tesh, R.B.; Barrett, A.D. Nucleotide sequences and phylogeny of the nucleocapsid gene of oropouche virus. J. Gen. Virol. 2000, 81, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Groseth, A.; Wollenberg, K.R.; Mampilli, V.; Shupert, T.; Weisend, C.; Guevara, C.; Kochel, T.J.; Tesh, R.B.; Ebihara, H. Spatiotemporal analysis of guaroa virus diversity, evolution, and spread in south america. Emerg. Infect. Dis. 2015, 21, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Hontz, R.D.; Guevara, C.; Halsey, E.S.; Silvas, J.; Santiago, F.W.; Widen, S.G.; Wood, T.G.; Casanova, W.; Vasilakis, N.; Watts, D.M.; et al. Itaya virus, a novel orthobunyavirus associated with human febrile illness, peru. Emerg. Infect. Dis. 2015, 21. [Google Scholar] [CrossRef] [PubMed]
- Tilston-Lunel, N.L.; Hughes, J.; Acrani, G.O.; da Silva, D.E.; Azevedo, R.S.; Rodrigues, S.G.; Vasconcelos, P.F.; Nunes, M.R.; Elliott, R.M. Genetic analysis of members of the species Oropouche virus and identification of a novel M segment sequence. J. Gen. Virol. 2015, 96, 1636–1650. [Google Scholar] [CrossRef] [PubMed]
- Van Eeden, C.; Harders, F.; Kortekaas, J.; Bossers, A.; Venter, M. Genomic and phylogenetic characterization of Shuni virus. Arch. Virol. 2014, 159, 2883–2892. [Google Scholar] [CrossRef] [PubMed]
- Ladner, J.T.; Savji, N.; Lofts, L.; Travassos da Rosa, A.; Wiley, M.R.; Gestole, M.C.; Rosen, G.E.; Guzman, H.; Vasconcelos, P.F.; Nunes, M.R.; et al. Genomic and phylogenetic characterization of viruses included in the Manzanilla and Oropouche species complexes of the genus Orthobunyavirus, family Bunyaviridae. J. Gen. Virol. 2014, 95, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Gauci, P.J.; McAllister, J.; Mitchell, I.R.; Boyle, D.B.; Bulach, D.M.; Weir, R.P.; Melville, L.F.; Gubala, A.J. Genomic characterisation of three Mapputta group viruses, a serogroup of Australian and Papua New Guinean bunyaviruses associated with human disease. PLoS One 2015, 10, e0116561. [Google Scholar] [CrossRef] [PubMed]
- Groseth, A.; Mampilli, V.; Weisend, C.; Dahlstrom, E.; Porcella, S.F.; Russell, B.J.; Tesh, R.B.; Ebihara, H. Molecular characterization of human pathogenic bunyaviruses of the Nyando and Bwamba/Pongola virus groups leads to the genetic identification of Mojui dos Campos and Kaeng Khoi virus. PLoS Negl. Trop. Dis. 2014, 8, e3147. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.R.T.; Chiang, J.O.; de Lima, C.P.S.; Martins, L.C.; Aragão Dias, A.; Cardoso, J.F.; Silva, S.P.; da Silva, D.E.A.; Oliveira, L.F.; Vasconcelos, J.M.; et al. New genome sequences of Gamboa viruses (family Bunyaviridae, genus Orthobunyavirus) isolated in Panama and Argentina. Genome Announc. 2014, 2, e00940-14. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.S.; Medeiros, D.B.; Rodrigues, S.G.; Martins, L.C.; de Lima, C.P.; de Oliveira, L.F.; de Vasconcelos, J.M.; da Silva, D.E.; Cardoso, J.F.; da Silva, S.P.; et al. Pacui Virus, Rio Preto da Eva Virus, and Tapirape Virus, three distinct viruses within the family Bunyaviridae. Genome Announc. 2014, 2, e00923-14. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, G.; Yang, Y.; Dang, R.; Zhao, T. Genome sequence of Abbey Lake virus, a novel orthobunyavirus isolated from china. Genome Announc. 2014, 2, e00433-14. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Lazuick, J.S.; Muth, D.J.; de Souza Lopes, O.; Crane, G.T.; Elbel, R.E.; Shope, R.E. Antigenic relationships among Tacaiuma complex viruses of the Anopheles A serogroup (Bunyaviridae). Bull. Pan Am. Health Organ. 1980, 14, 386–391. [Google Scholar] [PubMed]
- Da Rosa, J.F.S.; da Rosa, A.P.D.A.; Degallier, N.; Vasconcelos, P.F.D.C. Caracterização e relacionamento antigênico de três novos Bunyavirus no grupo Anopheles A (Bunyaviridae) dos arbovirus. Rev. Saúd. Públ. 1992, 26, 173–178. (In Portuguese) [Google Scholar] [CrossRef]
- Calisher, C.H.; Sasso, D.R.; Maness, K.S.; Gheorghiu, V.N.; Shope, R.E. Relationships of anopheles a group arboviruses. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1973, 143, 465–468. [Google Scholar] [CrossRef]
- Nichol, S.T.; Beaty, B.J.; Elliott, R.M.; Goldbach, R.; Plyusnin, A.; Schmaljohn, C.S.; Tesh, R.B. Family Bunyaviridae. In Virus Taxonomy—Eighth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Eds.; Elsevier/Academic Press: London, UK, 2005; pp. 695–716. [Google Scholar]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Bushbush, Ieri and Lukuni viruses, three unrelated new agents isolated from trinidadian forest mosquitoes. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1967, 125, 45–50. [Google Scholar] [CrossRef]
- Woodall, J.P. Virus Research in Amazonia. In Atas do Simpósio sôbre a Biota Amazônica; Conselho Nacional de Pesquitas: Rio de Janeiro, Brazil, 1967; Volume 6, Patologia,f; pp. 31–63. (In Portuguese) [Google Scholar]
- Karabatsos, N. International Catalogue of Arboviruses, Including Certain Other Viruses of Vertebrates, 3rd ed.; American Society of Tropical Medicine and Hygiene for the Subcommittee on Information Exchange of the American Committee on Arthropod-borne Viruses: San Antonio, TX, USA, 1985. [Google Scholar]
- Spence, L.; Anderson, C.R.; Aitken, T.H.G.; Downs, W.G. Bimiti virus, a new agent isolated from Trinidadian mosquitoes. Am. J. Trop. Med. Hyg. 1962, 11, 414–417. [Google Scholar] [PubMed]
- Whitman, L.; Casals, J. The guamá group: A new serological group of hitherto underscribed viruses. Immunological studies. Am. J. Trop. Med. Hyg. 1961, 10, 259–263. [Google Scholar] [PubMed]
- Causey, O.R.; Causey, C.E.; Maroja, O.M.; Macedo, D.G. The isolation of arthropod-borne viruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Am. J. Trop. Med. Hyg. 1961, 10, 227–249. [Google Scholar] [PubMed]
- Coleman, P.H.; Ryder, S.; Work, T.H. Mahogany Hammock virus, a new guama group arbovirus from the florida everglades. Am. J. Epidemiol. 1969, 89, 217–221. [Google Scholar] [PubMed]
- Doherty, R.L.; Carley, J.G.; Mackerras, M.J.; Marks, E.N. Studies of arthropod-borne virus infections in Queensland. III. Isolation and characterization of virus strains from wild-caught mosquitoes in North Queensland. Aust. J. Exp. Biol. Med. Sci. 1963, 41, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Doherty, R.L.; Whitehead, R.H.; Wetters, E.J.; Gorman, B.M. Studies of the epidemiology of arthropod-borne virus infections at Mitchell River Mission, Cape York Peninsula, North Queensland. II. Arbovirus infections of mosquitoes, man and domestic fowls, 1963–1966. Trans. R. Soc. Trop. Med. Hyg. 1968, 62, 430–438. [Google Scholar] [CrossRef]
- Boughton, C.R.; Hawkes, R.A.; Naim, H.M. Arbovirus infection in humans in NSW: Seroprevalence and pathogenicity of certain Australian bunyaviruses. Aust. N. Z. J. Med. 1990, 20, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.C.; Tukei, P.M.; Lule, M.; Mujomba, E.; Mukuye, A. Virology: Identification studies, An 1047–61. East Afr. Virus Res. Inst. Rep. 1966, 16, 24. [Google Scholar]
- Taylor, R.M. Catalogue of Arthropod-Borne Viruses of the World; US Government Printing Office, Public Health Service Pub. No. 1760: Washington, DC, USA, 1967.
- Fagbami, A.H.; Monath, T.P.; Murphy, F.A. Tataguine virus infection in mice and hamsters. Microbios 1980, 27, 89–96. [Google Scholar] [PubMed]
- McIntosh, B.M.; Kokernot, R.H.; Paterson, H.E. Witwatersrand virus: An apparently new virus isolated from culicine mosquitoes. S. Afr. J. Med. Sci. 1960, 23, 33–37. [Google Scholar]
- Shope, R.E.; Woodall, J.P.; Travassos da Rosa, A.P. The Epidemiology of Diseases Caused by Viruses in Groups C and Guama (Bunyaviridae). In The Arboviruses: Epidemiology and Ecology; Monath, T.P., Ed.; CRC Press: Boca Raton, Florida, 1988; Volume 3, pp. 38–51. [Google Scholar]
- Doherty, R.L.; Wetters, E.J.; Gorman, B.M.; Whitehead, R.H. Arbovirus infection in Western Queensland: serological studies, 1963–1969. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 740–747. [Google Scholar] [CrossRef]
- Doherty, R.L.; Whitehead, R.H.; Wetters, E.J.; Gorman, B.M.; Carley, J.G. A survey of antibody to 10 arboviruses (Koongol group, Mapputta group and ungrouped) isolated in Queensland. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 748–753. [Google Scholar] [CrossRef]
- Sanderson, C.J. A serologic survey of Queensland cattle for evidence of arbovirus infections. Am. J. Trop. Med. Hyg. 1969, 18, 433–439. [Google Scholar] [PubMed]
- Newton, S.E.; Short, N.J.; Irving, A.M.; Dalgarno, L. The Mapputta group of arboviruses: Ultrastructural and molecular studies which place the group in the Bunyavirus genus of the family Bunyaviridae. Aust. J. Exp. Biol. Med. Sci. 1983, 61, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Weir, R. Classification and Identification of Viruses Isolated from Mosquitoes in the Northern Territory, 1982–1992, Using a Range of Techniques. Ph.D. Thesis, University of Sydney, Sydney, Australia, 2002. [Google Scholar]
- Coffey, L.L.; Page, B.L.; Greninger, A.L.; Herring, B.L.; Russell, R.C.; Doggett, S.L.; Haniotis, J.; Wang, C.; Deng, X.; Delwart, E.L. Enhanced arbovirus surveillance with deep sequencing: Identification of novel rhabdoviruses and bunyaviruses in Australian mosquitoes. Virology 2014, 448, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Balducci, M.; Verani, P.; Lopes, M.C.; Gregoric, B. Isolation in Italy of Bahig and Matruh viruses (Tete group) from migrating birds. Ann. Microbiol. (Paris) 1973, 124B, 231–237. [Google Scholar]
- Converse, J.D.; Hoogstraal, H.; Moussa, M.I.; Stek, M., Jr.; Kaiser, M.N. Bahig virus (Tete group) in naturally- and transovarially-infected Hyalomma marginatum ticks from Egypt and Italy. Arch Gesamte Virusforsch 1974, 46, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, Z.; Rudolf, I. Tick-borne viruses in Europe. Parasitol. Res. 2012, 111, 9–36. [Google Scholar] [CrossRef] [PubMed]
- Brès, P.; Williams, M.C.; Chambon, L. Isolement au Sénégal d’un nouveau prototype d’arbovirus, la souche “Tataguine” (IPD/A 252). Ann. Inst. Pasteur 1966, 111, 585–591. (In French) [Google Scholar]
- Fagbami, A.H. Studies on transmission of Tataguine virus by Culex (pipens) fatigans mosquitoes. Afr. J. Med. Med. Sci. 1979, 8, 31–33. [Google Scholar] [PubMed]
- Fagbami, A.H.; Monath, T.P.; Tomori, O.; Lee, V.H.; Fabiyi, A. Studies on Tataguine infection in Nigeria. Trop. Geogr. Med. 1972, 24, 298–302. [Google Scholar] [PubMed]
- Salaun, J.J.; Rickenbach, A.; Brès, P.; Germain, M.; Eouzan, J.P.; Ferrara, L. Isolement au Cameroun de trois souches de virus Tataguine. Bull. Soc. Pathol. Exot. Ses Filiales 1968, 61, 557–564. (In French) [Google Scholar]
- Saluzzo, J.F.; Gonzalez, J.P.; Hervé, J.P.; Georges, A.J. Enquête sérologique sur la prévalence de certains arbovirus dans la population humaine du sud-est de la République Centrafricaine en 1979. Bull. Soc. Pathol. Exot. Ses Filiales 1981, 74, 490–499. (In French) [Google Scholar]
- Causey, O.R.; Kemp, G.E.; Madbouly, M.H.; Lee, V.H. Arbovirus surveillance in Nigeria, 1964–1967. Bull. Soc. Pathol. Exot. Ses Filiales 1969, 62, 249–253. [Google Scholar]
- Moore, D.L.; Causey, O.R.; Carey, D.E.; Reddy, S.; Cooke, A.R.; Akinkugbe, F.M.; David-West, T.S.; Kemp, G.E. Arthropod-borne viral infections of man in Nigeria, 1964–1970. Ann. Trop. Med. Parasitol. 1975, 69, 49–64. [Google Scholar] [PubMed]
- Fagbami, A.H.; Tomori, O. Tataguine virus isolations from humans in Nigeria, 1971–1975. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 788. [Google Scholar] [CrossRef]
- Digoutte, J.P.; Brès, P.; Nguyen Trung Luong, P.; Durand, B. Isolement du virus Tataguine à partir de deux fièvres exanthématiques. Bull. Soc. Pathol. Exot. Ses Filiales 1969, 62, 72–80. (In French) [Google Scholar]
- Bishop, D.H.; Shope, R.E. Bunyaviridae. In Comprehensive Virology; Fraenkel-Conrat, H., Wagner, R.R., Eds.; Plenum Press: New York, NY, USA, 1979; Volume 14. [Google Scholar]
- Center for Genomic Regulation. M-Coffee Aligns DNA, RNA or Proteins by Combining the Output of Popular Aligners. Available Online: http://tcoffee.crg.cat/apps/tcoffee/do:mcoffee (accessed on 11 September 2015).
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of best-fit models of protein evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Center for Biological Sequence Analysis. TMHMM Server v. 2.0. Prediction of Transmembrane Helices. Available online: http://www.cbs.dtu.dk/services/TMHMM-2.0/ (accessed on 14 April 2015).
- Center for Biological Sequence Analysis. NetNGlyc 1.0 Server. Available online: http://www.cbs.dtu.dk/services/NetNGlyc/ (accessed on 14 April 2015).
- Mohamed, M.; McLees, A.; Elliott, R.M. Viruses in the Anopheles A, Anopheles B, and Tete serogroups in the Orthobunyavirus genus (family Bunyaviridae) do not encode an NSs protein. J. Virol. 2009, 83, 7612–7618. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.D.; Mishra, A.C.; Mourya, D.T. Molecular characterization of Umbre virus (Bunyaviridae). Virol. J. 2008, 5, 115. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Eifan, S.A.; Elliott, R.M. Mutational analysis of the Bunyamwera orthobunyavirus nucleocapsid protein gene. J. Virol. 2009, 83, 11307–11317. [Google Scholar] [CrossRef] [PubMed]
- Fazakerley, J.K.; Gonzalez-Scarano, F.; Strickler, J.; Dietzschold, B.; Karush, F.; Nathanson, N. Organization of the middle RNA segment of snowshoe hare bunyavirus. Virology 1988, 167, 422–432. [Google Scholar] [CrossRef]
- Plassmeyer, M.L.; Soldan, S.S.; Stachelek, K.M.; Roth, S.M.; Martin-Garcia, J.; Gonzalez-Scarano, F. Mutagenesis of the La Crosse Virus glycoprotein supports a role for Gc (1066–1087) as the fusion peptide. Virology 2007, 358, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Grady, L.J.; Sanders, M.L.; Campbell, W.P. The sequence of the M RNA of an isolate of La Crosse virus. J. Gen. Virol. 1987, 68, 3057–3071. [Google Scholar] [CrossRef] [PubMed]
- Lees, J.F.; Pringle, C.R.; Elliott, R.M. Nucleotide sequence of the Bunyamwera virus M RNA segment: Conservation of structural features in the bunyavirus glycoprotein gene product. Virology 1986, 148, 1–14. [Google Scholar] [CrossRef]
- Pardigon, N.; Vialat, P.; Gerbaud, S.; Girard, M.; Bouloy, M. Nucleotide-sequence of the M-segment of Germiston virus - comparison of the M-gene product of several bunyaviruses. Virus Res. 1988, 11, 73–85. [Google Scholar] [CrossRef]
- Braakman, I.; van Anken, E. Folding of viral envelope glycoproteins in the endoplasmic reticulum. Traffic 2000, 1, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Brauburger, K.; Elliott, R.M. Role of N-linked glycans on Bunyamwera virus glycoproteins in intracellular trafficking, protein folding, and virus infectivity. J. Virol. 2005, 79, 13725–13734. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Elliott, R.M. Mutagenesis of the L protein encoded by Bunyamwera virus and production of monospecific antibodies. J. Gen. Virol. 1992, 73, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.A.; Harrison, A.K.; Whitfield, S.G. Bunyaviridae: Morphologic and morphogenetic similarities of Bunyamwera serologic supergroup viruses and several other arthropod-borne viruses. Intervirology 1973, 1, 297–316. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, J.S.; Casals, J.; Chumakov, M.P.; Gaidamovich, S.Y.; Hannoun, C.; Holmes, I.H.; Horzinek, M.C.; Mussgay, M.; Oker-Blom, N.; Russell, P.K. Bunyaviruses and Bunyaviridae. Intervirology 1975, 6, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, J.S.; Casals, J.; Chumakov, M.P.; Gaidamovich, S.Y.; Hannoun, C.; Holmes, I.H.; Horzinek, M.C.; Mussgay, M.; Russell, P.K. Bunyaviruses and Bunyaviridae. Intervirology 1974, 2, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.H.; Calisher, C.H.; Casals, J.; Chumakov, M.P.; Gaidamovich, S.Y.; Hannoun, C.; Lvov, D.K.; Marshall, I.D.; Oker-Blom, N.; Pettersson, R.F.; et al. Bunyaviridae. Intervirology 1980, 14, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, M.J.; Bruenn, J.A.; Hay, J.; Czechowski, D.; Taylor, D.J. Discovery and evolution of bunyavirids in arctic phantom midges and ancient bunyavirid-like sequences in insect genomes. J. Virol. 2014, 88, 8783–8794. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Kolodziejek, J.; Rudolf, I.; Bercic, R.; Nowotny, N.; Hubalek, Z. Partial genetic characterization of Sedlec virus (Orthobunyavirus, Bunyaviridae). Infect. Genet. Evol. 2013, 19, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; da Rosa, A.T.; Savji, N.; Sze, W.; Wick, I.; Guzman, H.; Hutchison, S.; Tesh, R.; Lipkin, W.I. Aguacate virus, a new antigenic complex of the genus Phlebovirus (family Bunyaviridae). J. Gen. Virol. 2011, 92, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Savji, N.; Travassos da Rosa, A.; Desai, A.; Sanchez-Seco, M.P.; Guzman, H.; Lipkin, W.I.; Tesh, R. Characterization of the Salehabad virus species complex of the genus Phlebovirus (Bunyaviridae). J. Gen. Virol. 2013, 94, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Savji, N.; Travassos da Rosa, A.; Guzman, H.; Yu, X.; Desai, A.; Rosen, G.E.; Hutchison, S.; Lipkin, W.I.; Tesh, R. Characterization of the Uukuniemi virus group (Phlebovirus: Bunyaviridae): Evidence for seven distinct species. J. Virol. 2013, 87, 3187–3195. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Tesh, R.; Travassos da Rosa, A.; Savji, N.; Sze, W.; Jain, K.; Serge, R.; Guzman, H.; Guevara, C.; Nunes, M.R.; et al. Characterization of the Candiru antigenic complex (Bunyaviridae: Phlebovirus), a highly diverse and reassorting group of viruses affecting humans in tropical America. J. Virol. 2011, 85, 3811–3820. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Tesh, R.B.; Savji, N.; Travassos da Rosa, A.P.; Guzman, H.; Bussetti, A.V.; Desai, A.; Ladner, J.; Sanchez-Seco, M.; Lipkin, W.I. Characterization of the Sandfly fever Naples species complex and description of a new Karimabad species complex (genus Phlebovirus, family Bunyaviridae). J. Gen. Virol. 2014, 95, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Wiley, M.R.; Travassos da Rosa, A.P.; Guzman, H.; Quiroz, E.; Savji, N.; Carrera, J.P.; Bussetti, A.V.; Ladner, J.T.; Lipkin, W.I.; et al. Characterization of the Punta Toro species complex (genus Phlebovirus, family Bunyaviridae). J. Gen. Virol. 2015, 96, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, K.; Weisend, C.; Kajihara, M.; Matysiak, C.; Williamson, B.N.; Simuunza, M.; Mweene, A.S.; Takada, A.; Tesh, R.B.; Ebihara, H. Comprehensive molecular detection of tick-borne phleboviruses leads to the retrospective identification of taxonomically unassigned bunyaviruses and the discovery of a novel member of the genus phlebovirus. J. Virol. 2015, 89, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, K.; Weisend, C.; Travassos da Rosa, A.P.; Anzick, S.L.; Dahlstrom, E.; Porcella, S.F.; Dorward, D.W.; Yu, X.J.; Tesh, R.B.; Ebihara, H. Characterization of the Bhanja serogroup viruses (Bunyaviridae): A novel species of the genus Phlebovirus and its relationship with other emerging tick-borne phleboviruses. J. Virol. 2013, 87, 3719–3728. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Williams, S.H.; Sameroff, S.; Sanchez Leon, M.; Jain, K.; Lipkin, W.I. Virome Analysis of Amblyomma americanum, Dermacentor variabilis, and Ixodes scapularis Ticks Reveals Novel Highly Divergent Vertebrate and Invertebrate Viruses. J. Virol. 2014, 88, 11480–11492. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-X.; Shi, M.; Tian, J.-H.; Lin, X.-D.; Kang, Y.-J.; Chen, L.-J.; Qin, X.-C.; Xu, J.; Holmes, E.C.; Zhang, Y.-Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- L’vov, D.K.; Al’khovskii, S.V.; Shchelkanov, M.Y.; Shchetinin, A.M.; Deriabin, P.G.; Gitel’man, A.K.; Aristova, V.A.; Botikov, A.G. [Taxonomic status of the Burana virus (BURV) (Bunyaviridae, Nairovirus, Tamdy group) isolated from the ticks Haemaphysalis punctata Canestrini et Fanzago, 1877 and Haem. concinna Koch, 1844 (Ixodidae, Haemaphysalinae) in Kyrgyzstan]. Vopr. Virusol. 2014, 59, 10–15. (In Russian) [Google Scholar] [PubMed]
- L’vov, D.K.; Al’khovskii, S.V.; Shchelkanov, M.Y.; Shchetinin, A.M.; Deriabin, P.G.; Samokhvalov, E.I.; Gitel’man, A.K.; Botikov, A.G. [Genetic characterization of the Caspiy virus (CASV) (Bunyaviridae, Nairovirus) isolated from the Laridae (Vigors, 1825) and Sternidae (Bonaparte, 1838) birds and the Argasidae (Koch, 1844) ticks Ornithodoros capensis Neumann, 1901, in Western and Eastern coasts of the Caspian Sea]. Vopr. Virusol. 2014, 59, 24–29. (In Russian) [Google Scholar] [PubMed]
- Al’khovskii, S.V.; L’Vov, D.K.; Shchelkanov, M.; Shchetinin, A.M.; Deriabin, P.G.; Gitel’man, A.K.; Botikov, A.G.; Samokhvalov, E.I.; Zakarian, V.A. [Taxonomic status of the Artashat virus (ARTSV) (Bunyaviridae, Nairovirus) isolated from the ticks Ornithodoros alactagalis Issaakjan, 1936 and O. verrucosus Olenev, Sassuchin et Fenuk, 1934 (Argasidae Koch, 1844) collected in Transcaucasia]. Vopr. Virusol. 2014, 59, 24–28. (In Russian) [Google Scholar] [PubMed]
- Al’khovskii, S.V.; L’Vov, D.K.; Shchelkanov, M.; Shchetinin, A.M.; Deriabin, P.G.; Samokhvalov, E.I.; Gitel’man, A.K.; Botikov, A.G. [The taxonomy of the Issyk-Kul virus (ISKV, Bunyaviridae, Nairovirus), the etiologic agent of the Issyk-Kul fever isolated from bats (Vespertilionidae) and ticks Argas (Carios) vespertilionis (Latreille, 1796)]. Vopr. Virusol. 2013, 58, 11–15. (In Russian) [Google Scholar] [PubMed]
- Ishii, A.; Ueno, K.; Orba, Y.; Sasaki, M.; Moonga, L.; Hang’ombe, B.M.; Mweene, A.S.; Umemura, T.; Ito, K.; Hall, W.W.; Sawa, H. A nairovirus isolated from African bats causes haemorrhagic gastroenteritis and severe hepatic disease in mice. Nat. Commun. 2014, 5, 5651. [Google Scholar] [CrossRef] [PubMed]
- Cowley, J.A.; McCulloch, R.J.; Spann, K.M.; Cadogan, L.C.; Walker, P.J. Preliminary Molecular and Biological Characterization of Mourilyan Virus (MoV): A New Bunya-Related Virus of Penaeid Prawns. In Diseases in Asian Aquaculture V. Proceedings of the 5th Symposium on Diseases in Asian Aquaculture; Walker, P.J., Lester, R.G., Bondad-Reantaso, M.G., Eds.; Fish Health Section, Asian Fisheries Society: Manila, The Philippines, 2005; pp. 113–124. [Google Scholar]
- Mielke-Ehret, N.; Mühlbach, H.-P. Emaravirus: A novel genus of multipartite, negative strand RNA plant viruses. Viruses 2012, 4, 1515–1536. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; Garcia, M.L.; Goodin, M.; Sasaya, T.; Haenni, A.L. Negative-strand RNA viruses: The plant-infecting counterparts. Virus Res. 2011, 162, 184–202. [Google Scholar] [CrossRef] [PubMed]
- Briese, T.; Calisher, C.H.; Higgs, S. Viruses of the family Bunyaviridae: Are all available isolates reassortants? Virology 2013, 446, 207–216. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shchetinin, A.M.; Lvov, D.K.; Deriabin, P.G.; Botikov, A.G.; Gitelman, A.K.; Kuhn, J.H.; Alkhovsky, S.V. Genetic and Phylogenetic Characterization of Tataguine and Witwatersrand Viruses and Other Orthobunyaviruses of the Anopheles A, Capim, Guamá, Koongol, Mapputta, Tete, and Turlock Serogroups. Viruses 2015, 7, 5987-6008. https://doi.org/10.3390/v7112918
Shchetinin AM, Lvov DK, Deriabin PG, Botikov AG, Gitelman AK, Kuhn JH, Alkhovsky SV. Genetic and Phylogenetic Characterization of Tataguine and Witwatersrand Viruses and Other Orthobunyaviruses of the Anopheles A, Capim, Guamá, Koongol, Mapputta, Tete, and Turlock Serogroups. Viruses. 2015; 7(11):5987-6008. https://doi.org/10.3390/v7112918
Chicago/Turabian StyleShchetinin, Alexey M., Dmitry K. Lvov, Petr G. Deriabin, Andrey G. Botikov, Asya K. Gitelman, Jens H. Kuhn, and Sergey V. Alkhovsky. 2015. "Genetic and Phylogenetic Characterization of Tataguine and Witwatersrand Viruses and Other Orthobunyaviruses of the Anopheles A, Capim, Guamá, Koongol, Mapputta, Tete, and Turlock Serogroups" Viruses 7, no. 11: 5987-6008. https://doi.org/10.3390/v7112918
APA StyleShchetinin, A. M., Lvov, D. K., Deriabin, P. G., Botikov, A. G., Gitelman, A. K., Kuhn, J. H., & Alkhovsky, S. V. (2015). Genetic and Phylogenetic Characterization of Tataguine and Witwatersrand Viruses and Other Orthobunyaviruses of the Anopheles A, Capim, Guamá, Koongol, Mapputta, Tete, and Turlock Serogroups. Viruses, 7(11), 5987-6008. https://doi.org/10.3390/v7112918