Viruses of Polar Aquatic Environments

{kind=link}
Abstract
:1. Introduction
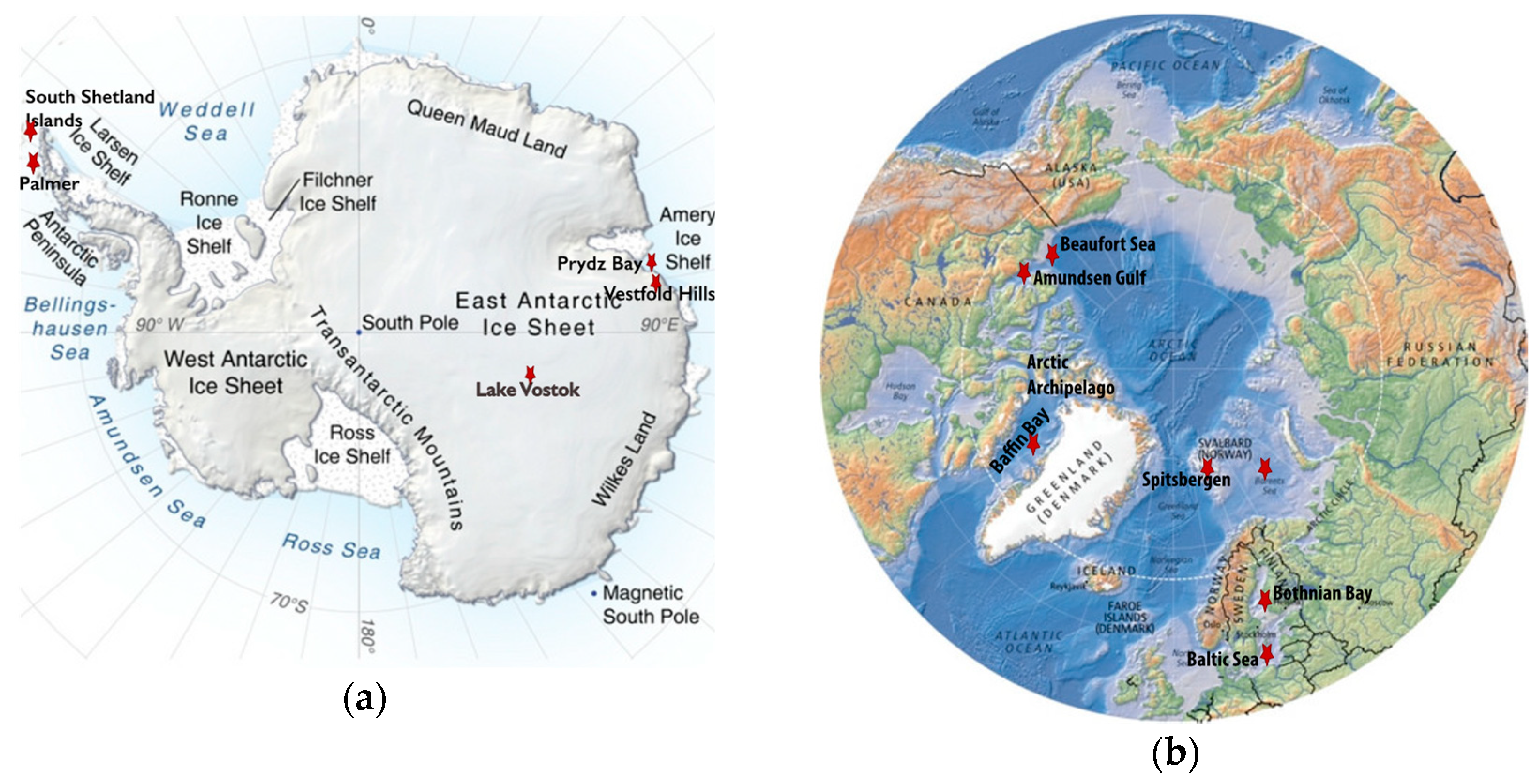
2. Antarctica
2.1. DNA Viruses in Antarctica
2.2. RNA Viruses in Antarctica
3. Viruses in the Arctic
3.1. DNA Viruses in the Arctic
3.2. RNA Viruses in the Arctic
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Boetius, A.; Anesio, A.M.; Deming, J.W.; Mikucki, J.A.; Rapp, J.Z. Microbial ecology of the cryosphere: Sea ice and glacial habitats. Nat. Rev. Microbiol. 2015, 13, 677. [Google Scholar] [CrossRef] [PubMed]
- Rampelotto, P.H. Polar microbiology: Recent advances and future perspectives. Biology 2014, 3, 81–84. [Google Scholar] [CrossRef]
- Laybourn-Parry, J. No place too cold. Science 2009, 324, 1521–1522. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R. Microbial ecology of antarctic aquatic systems. Nat. Rev. Microbiol. 2015, 13, 691. [Google Scholar] [CrossRef] [PubMed]
- Anesio, A.M.; Bellas, C.M. Are low temperature habitats hot spots of microbial evolution driven by viruses? Trends Microbiol. 2011, 19, 52–57. [Google Scholar] [CrossRef]
- Anesio, A.M.; Laybourn-Parry, J. Glaciers and ice sheets as a biome. Trends Ecol. Evol. 2012, 27, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Ghiglione, J.-F.; Galand, P.E.; Pommier, T.; Pedrós-Alió, C.; Maas, E.W.; Bakker, K.; Bertilson, S.; Kirchman, D.L.; Lovejoy, C.; Yager, P.L.; et al. Pole-to-pole biogeography of surface and deep marine bacterial communities. Proc. Natl. Acad. Sci. USA 2012, 109, 17633–17638. [Google Scholar] [CrossRef]
- Herron, E.M.; Dewey, J.F.; Pitman, I.W.C. Plate tectonics model for the evolution of the arctic. Geology 1974, 2, 377–380. [Google Scholar] [CrossRef]
- Gaina, C.; Medvedev, S.; Torsvik, T.H.; Koulakov, I.; Werner, S.C. 4d arctic: A glimpse into the structure and evolution of the arctic in the light of new geophysical maps, plate tectonics and tomographic models. Surv. Geophys. 2014, 35, 1095–1122. [Google Scholar] [CrossRef]
- Kanao, M.; Suvorov, V.D.; Toda, S.; Tsuboi, S. Seismicity, structure and tectonics in the arctic region. Geosci. Front. 2015, 6, 665–677. [Google Scholar] [CrossRef]
- Adie, R.J. The geology of antarctica. In Antarctic Research: The Matthew Fontaine Maury Memorial Symposium; John Wiley & Sons: Hoboken, NJ, USA, 1962. [Google Scholar]
- Amy, J.B.; Joseph, A.M.; Joseph, D.P.; Phillipe, C.; Fabien, J.; Wade, H.J. Microbial diversity in a pacific ocean transect from the arctic to antarctic circles. Aquat. Microbial Ecol. 2005, 41, 91–102. [Google Scholar]
- Suttle, C.A. Marine viruses—Major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801. [Google Scholar] [CrossRef] [PubMed]
- Middelboe, M.; Brussaard, C.P.D. Marine viruses: Key players in marine ecosystems. Viruses 2017, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Bergh, Ø.; BØrsheim, K.Y.; Bratbak, G.; Heldal, M. High abundance of viruses found in aquatic environments. Nature 1989, 340, 467. [Google Scholar] [CrossRef] [PubMed]
- Roux, S.; Hallam, S.J.; Woyke, T.; Sullivan, M.B. Viral dark matter and virus–host interactions resolved from publicly available microbial genomes. eLife 2015, 4, e08490. [Google Scholar] [CrossRef] [PubMed]
- López-Bueno, A.; Tamames, J.; Velázquez, D.; Moya, A.; Quesada, A.; Alcamí, A. High diversity of the viral community from an antarctic lake. Science 2009, 326, 858–861. [Google Scholar] [CrossRef] [PubMed]
- Säwström, C.; Lisle, J.; Anesio, A.M.; Priscu, J.C.; Laybourn-Parry, J.J.E. Bacteriophage in polar inland waters. Extremophiles 2008, 12, 167–175. [Google Scholar] [CrossRef]
- Williams, T.J.; Long, E.; Evans, F.; DeMaere, M.Z.; Lauro, F.M.; Raftery, M.J.; Ducklow, H.; Grzymski, J.J.; Murray, A.E.; Cavicchioli, R. A metaproteomic assessment of winter and summer bacterioplankton from antarctic peninsula coastal surface waters. ISME J. 2012, 6, 1883–1900. [Google Scholar] [CrossRef]
- Schofield, O.; Saba, G.; Coleman, K.; Carvalho, F.; Couto, N.; Ducklow, H.; Finkel, Z.; Irwin, A.; Kahl, A.; Miles, T.; et al. Decadal variability in coastal phytoplankton community composition in a changing west antarctic peninsula. Deep Sea Res. Part I: Oceanogr. Res. Pap. 2017, 124, 42–54. [Google Scholar] [CrossRef]
- Brum, J.R.; Hurwitz, B.L.; Schofield, O.; Ducklow, H.W.; Sullivan, M.B. Seasonal time bombs: Dominant temperate viruses affect southern ocean microbial dynamics. ISME J. 2016, 10, 437–449. [Google Scholar] [CrossRef]
- Li, Z.; Cassar, N.; Huang, K.; Ducklow, H.; Schofield, O. Interannual variability in net community production at the western antarctic peninsula region (1997–2014). J. Geophys. Res. Oceans 2016, 121, 4748–4762. [Google Scholar] [CrossRef]
- Saba, G.K.; Fraser, W.R.; Saba, V.S.; Iannuzzi, R.A.; Coleman, K.E.; Doney, S.C.; Ducklow, H.W.; Martinson, D.G.; Miles, T.N.; Patterson-Fraser, D.L.; et al. Winter and spring controls on the summer food web of the coastal west antarctic peninsula. Nat. Commun. 2014, 5, 4318. [Google Scholar] [CrossRef] [PubMed]
- Grzymski, J.J.; Riesenfeld, C.S.; Williams, T.J.; Dussaq, A.M.; Ducklow, H.; Erickson, M.; Cavicchioli, R.; Murray, A.E. A metagenomic assessment of winter and summer bacterioplankton from antarctica peninsula coastal surface waters. ISME J. 2012, 6, 1901. [Google Scholar] [CrossRef] [PubMed]
- Laybourn-Parry, J.; Pearce, D.A. The biodiversity and ecology of antarctic lakes: Models for evolution. Philos. Trans. Royal Soc. 2007, 362, 2273–2289. [Google Scholar] [CrossRef] [PubMed]
- Tschitschko, B.; Williams, T.J.; Allen, M.A.; Zhong, L.; Raftery, M.J.; Cavicchioli, R. Ecophysiological distinctions of haloarchaea from a hypersaline antarctic lake as determined by metaproteomics. Appl. Environ. Microbiol. 2016, 82, 3165–3173. [Google Scholar] [CrossRef] [PubMed]
- Chenard, C.; Lauro, F.M. Exploring the viral ecology of high latitude aquatic systems. In Microbial Ecology of Extreme Environments; Springer International Publishing: Berlin, Germany, 2017; pp. 185–200. [Google Scholar]
- Rastrojo, A.; Alcamí, A. Viruses in polar lake and soil ecosystems. Adv. Virus Res. 2018, 101, 39–54. [Google Scholar]
- Aguirre de Cárcer, D.; López-Bueno, A.; Alonso-Lobo, J.M.; Quesada, A.; Alcamí, A. Metagenomic analysis of lacustrine viral diversity along a latitudinal transect of the antarctic peninsula. FEMS Microbiol. Ecol. 2016, 92, fiw074. [Google Scholar] [CrossRef]
- López-Bueno, A.; Rastrojo, A.; Peiró, R.; Arenas, M.; Alcamí, A. Ecological connectivity shapes quasispecies structure of rna viruses in an antarctic lake. Mol. Ecol. 2015, 24, 4812–4825. [Google Scholar] [CrossRef]
- Aguirre de Cárcer, D.; López-Bueno, A.; Pearce, D.A.; Alcamí, A. Biodiversity and distribution of polar freshwater DNA viruses. Sci. Adv. 2015, 1, e1400127. [Google Scholar] [CrossRef]
- Tschitschko, B.; Williams, T.J.; Allen, M.A.; Páez-Espino, D.; Kyrpides, N.; Zhong, L.; Raftery, M.J.; Cavicchioli, R. Antarctic archaea–virus interactions: Metaproteome-led analysis of invasion, evasion and adaptation. ISME J. 2015, 9, 2094. [Google Scholar] [CrossRef]
- Lauro, F.M.; DeMaere, M.Z.; Yau, S.; Brown, M.V.; Ng, C.; Wilkins, D.; Raftery, M.J.; Gibson, J.A.E.; Andrews-Pfannkoch, C.; Lewis, M.; et al. An integrative study of a meromictic lake ecosystem in antarctica. ISME J. 2010, 5, 879. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.; Lauro, F.M.; DeMaere, M.Z.; Brown, M.V.; Thomas, T.; Raftery, M.J.; Andrews-Pfannkoch, C.; Lewis, M.; Hoffman, J.M.; Gibson, J.A.; et al. Virophage control of antarctic algal host–virus dynamics. Proc. Natl. Acad. Sci. USA 2011, 108, 6163–6168. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.A.; Culley, A.I.; Schvarcz, C.R.; Steward, G.F. RNA viruses as major contributors to antarctic virioplankton. Environ. Microbiol. 2016, 18, 3714–3727. [Google Scholar] [CrossRef] [PubMed]
- Brum, J.R.; Ignacio-Espinoza, J.C.; Roux, S.; Doulcier, G.; Acinas, S.G.; Alberti, A.; Chaffron, S.; Cruaud, C.; de Vargas, C.; Gasol, J.M.; et al. Patterns and ecological drivers of ocean viral communities. Science 2015, 348, 1261498. [Google Scholar] [CrossRef] [PubMed]
- Toro, M.; Camacho, A.; Rochera Cordellat, C.; Rico, E.; Bañón, M.; Fernández-Valiente, E.; Marco, E.; Justel, A.; Avendaño, M.; Ariosa, Y.; et al. Limnological characteristics of the freshwater ecosystems of byers peninsula, livingston island, in maritime antarctica. Polar Biol. 2007, 30, 635–649. [Google Scholar] [CrossRef]
- Angly, F.E.; Felts, B.; Breitbart, M.; Salamon, P.; Edwards, R.A.; Carlson, C.; Chan, A.M.; Haynes, M.; Kelley, S.; Liu, H.; et al. The marine viromes of four oceanic regions. PLoS Biol. 2006, 4, e368. [Google Scholar] [CrossRef] [PubMed]
- Derelle, E.; Ferraz, C.; Escande, M.-L.; Eychenié, S.; Cooke, R.; Piganeau, G.; Desdevises, Y.; Bellec, L.; Moreau, H.; Grimsley, N. Life-cycle and genome of otv5, a large DNA virus of the pelagic marine unicellular green alga ostreococcus tauri. PLoS ONE 2008, 3, e2250. [Google Scholar] [CrossRef]
- Ducklow, H.; Clarke, A.; Dickhut, R.; Doney, S.C.; Geisz, H.; Huang, K.; Martinson, D.G.; Meredith, M.P.; Moeller, H.V.; Montes-Hugo, M.; et al. The marine system of the western antarctic peninsula. In Antarctica Ecosystems: An Extreme Environment in a Changing World; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- La Scola, B.; Desnues, C.; Pagnier, I.; Robert, C.; Barrassi, L.; Fournous, G.; Merchat, M.; Suzan-Monti, M.; Forterre, P.; Koonin, E.; et al. The virophage as a unique parasite of the giant mimivirus. Nature 2008, 455, 100. [Google Scholar] [CrossRef]
- Ng, C.; DeMaere, M.Z.; Williams, T.J.; Lauro, F.M.; Raftery, M.; Gibson, J.A.E.; Andrews-Pfannkoch, C.; Lewis, M.; Hoffman, J.M.; Thomas, T.; et al. Metaproteogenomic analysis of a dominant green sulfur bacterium from ace lake, antarctica. ISME J. 2010, 4, 1002. [Google Scholar] [CrossRef]
- Madan, N.J.; Marshall, W.A.; Laybourn-Parry, J. Virus and microbial loop dynamics over an annual cycle in three contrasting antarctic lakes. Freshwater Biol. 2005, 50, 1291–1300. [Google Scholar] [CrossRef]
- Laybourn-Parry, J.; Bell, E.M. Ace lake: Three decades of research on a meromictic, antarctic lake. Polar Biol. 2014, 37, 1685–1699. [Google Scholar] [CrossRef]
- Seshadri, R.; Kravitz, S.A.; Smarr, L.; Gilna, P.; Frazier, M. Camera: A community resource for metagenomics. PLoS Biol. 2007, 5, e75. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, W.; Yan, S.; Xiao, J.; Zhang, Y.; Li, B.; Pan, Y.; Wang, Y. Diversity of virophages in metagenomic data sets. J. Virol. 2013, 87, 4225–4236. [Google Scholar] [CrossRef]
- Fischer, M.G.; Suttle, C.A. A virophage at the origin of large DNA transposons. Science 2011, 332, 231. [Google Scholar] [CrossRef]
- Yutin, N.; Shevchenko, S.; Kapitonov, V.; Krupovic, M.; Koonin, E.V. A novel group of diverse polinton-like viruses discovered by metagenome analysis. BMC Biol. 2015, 13, 95. [Google Scholar] [CrossRef]
- Ferris, J.M.; Burton, H.R.J.H. The annual cycle of heat content and mechanical stability of hypersaline deep lake, vestfold hills, antarctica. In Biology of the Vestfold Hills, Antarctica: Proceedings of the Symposium, Hobart, August 1984; Springer: Berlin, Germany, 1988; Volume 165, pp. 115–128. [Google Scholar]
- Krupovič, M.; Forterre, P.; Bamford, D.H. Comparative analysis of the mosaic genomes of tailed archaeal viruses and proviruses suggests common themes for virion architecture and assembly with tailed viruses of bacteria. J. Mol. Biol. 2010, 397, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Gowing, M.M. Large viruses and infected microeukaryotes in ross sea summer pack ice habitats. J. Mar. Biol. 2003, 142, 1029–1040. [Google Scholar] [CrossRef]
- Luhtanen, A.-M.; Eronen-Rasimus, E.; Oksanen, H.M.; Tison, J.-L.; Delille, B.; Dieckmann, G.S.; Rintala, J.-M.; Bamford, D.H. The first known virus isolates from antarctic sea ice have complex infection patterns. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef]
- Gowing, M.M.; Riggs, B.; Garrison, D.L.; Gibson, A.H.; Jeffries, M. Large viruses in ross sea late autumn pack ice habitats. Mar. Ecol. Prog. Ser. 2002, 241, 1–11. [Google Scholar] [CrossRef]
- Paterson, H.; Laybourn-Parry, J.J.P.B. Antarctic sea ice viral dynamics over an annual cycle. Polar Biol. 2012, 35, 491–497. [Google Scholar] [CrossRef]
- De Corte, D.; Sintes, E.; Yokokawa, T.; Herndl, G.J. Changes in viral and bacterial communities during the ice-melting season in the coastal arctic (kongsfjorden, ny- lesund). Environ. Microbiol. 2011, 13, 1827–1841. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Matthews, B.; Suttle, C.A. Effects of environmental variation and spatial distance on bacteria, archaea and viruses in sub-polar and arctic waters. ISME J. 2013, 7, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Venger, M.P.; Kopylov, A.I.; Zabotkina, E.A.; Makarevich, P.R. The influence of viruses on bacterioplankton of the offshore and coastal parts of the barents sea. Russ. J. Mar. Biol. 2016, 42, 26–35. [Google Scholar] [CrossRef]
- Shirokolobova, T.I.; Zhichkin, A.P.; Venger, M.P.; Vodopyanova, V.V.; Moissev, D.V. Bacteria and viruses of the ice-free aquatic area of teh barents sea at the beginning of polar night. Dokl. Biol. Sci. 2016, 469, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Collins, R.E.; Deming, J.W.J.P.B. Abundant dissolved genetic material in arctic sea ice part ii: Viral dynamics during autumn freeze-up. Polar Biol. 2011, 34, 1831–1841. [Google Scholar] [CrossRef]
- Simon, N.; Foulon, E.; Grulois, D.; Six, C.; Desdevises, Y.; Latimier, M.; Le Gall, F.; Tragin, M.; Houdan, A.; Derelle, E.; et al. Revision of the genus micromonas manton et parke (chlorophyta, mamiellophyceae), of the type species m. Pusilla (butcher) manton & parke and of the species m. Commoda van baren, bachy and worden and description of two new species based on the genetic and phenotypic characterization of cultured isolates. Protist 2017, 168, 612–635. [Google Scholar] [PubMed]
- Hingamp, P.; Grimsley, N.; Acinas, S.G.; Clerissi, C.; Subirana, L.; Poulain, J.; Ferrera, I.; Sarmento, H.; Villar, E.; Lima-Mendez, G.; et al. Exploring nucleo-cytoplasmic large DNA viruses in tara oceans microbial metagenomes. ISME J. 2013, 7, 1678. [Google Scholar] [CrossRef]
- Maat, D.S.; Biggs, T.; Evans, C.; van Bleijswijk, J.D.L.; van der Wel, N.N.; Dutilh, B.E.; Brussaard, C.P.D. Characterization and temperature dependence of arctic micromonas polaris viruses. Viruses 2017, 9, 134. [Google Scholar] [CrossRef]
- Wells, L.E.; Deming, J. Characterization of a cold-active bacteriophage on two psychrophilic marine hosts. Aqua. Microb. Ecol. 2006, 45, 15–29. [Google Scholar] [CrossRef]
- Payet, J.; Suttle, C.A. Viral infection of bacteria and phytoplankton in the arctic ocean as viewed through the lens of fingerprint analysis. Aqua. Microb. Ecol. 2014, 72, 47–61. [Google Scholar] [CrossRef]
- Rohwer, F.; Edwards, R. The phage proteomic tree: A genome-based taxonomy for phage. J. Bacteriol. 2002, 184, 4529. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.-C.; Chen, X.-L.; Shen, Q.-T.; Zhao, D.-L.; Tang, B.-L.; Su, H.-N.; Wu, Z.-Y.; Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; et al. Filamentous phages prevalent in pseudoalteromonas spp. Confer properties advantageous to host survival in arctic sea ice. ISME J. 2014, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Rakonjac, J.; Das, B.; Derda, R. Editorial: Filamentous bacteriophage in bio/nano/technology, bacterial pathogenesis and ecology. Front. Microb. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Riemann, L.; Holmfeldt, K.; Titelman, J. Importance of viral lysis and dissolved DNA for bacterioplankton activity in a p-limited estuary, northern baltic sea. J. Microb. Ecol. 2009, 57, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Tangherlini, M.; Dell’Anno, A. From virus isolation to metagenome generation for investigating viral diversity in deep-sea sediments. Sci. Rep. 2017, 7, 8355. [Google Scholar] [CrossRef]
- Zeigler Allen, L.; McCrow, J.P.; Ininbergs, K.; Dupont, C.L.; Badger, J.H.; Hoffman, J.M.; Ekman, M.; Allen, A.E.; Bergman, B.; Venter, J.C. The baltic sea virome: Diversity and transcriptional activity of DNA and rna viruses. mSystems 2017, 2. [Google Scholar] [CrossRef]
- Luhtanen, A.M.; Eronen-Rasimus, E.; Kaartokallio, H.; Rintala, J.M.; Autio, R.; Roine, E. Isolation and characterization of phage–host systems from the baltic sea ice. Extremophiles 2014, 18, 121–130. [Google Scholar] [CrossRef]
- Senčilo, A.; Luhtanen, A.-M.; Saarijärvi, M.; Bamford, D.H.; Roine, E. Cold-active bacteriophages from the baltic sea ice have diverse genomes and virus–host interactions. Environ. Microb. 2014, 17, 3628–3641. [Google Scholar] [CrossRef]
- Krishnamurthy, S.R.; Wang, D. Origins and challenges of viral dark matter. Virus Res. 2017, 239, 136–142. [Google Scholar] [CrossRef]
- Simmonds, P.; Adams, M.J.; Benkő, M.; Breitbart, M.; Brister, J.R.; Carstens, E.B.; Davison, A.J.; Delwart, E.; Gorbalenya, A.E.; Harrach, B.; et al. Virus taxonomy in the age of metagenomics. Nat. Rev. Microb. 2017, 15, 161. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Nute, M.G.; Warnow, T.; Pop, M. Misunderstood parameter of ncbi blast impacts the correctness of bioinformatics workflows. Bioinformatics 2018, bty833. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Altschul, S.F.; Pop, M. Outlier detection in blast hits. Algorithm. Mol. Biol. 2018, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Culley, A.I.; Lang, A.S.; Suttle, C.A. Metagenomic analysis of coastal RNA virus communities. Science 2006, 312, 1795–1798. [Google Scholar] [CrossRef] [PubMed]
- Urayama, S.-I.; Takaki, Y.; Nishi, S.; Yoshida-Takashima, Y.; Deguchi, S.; Takai, K.; Nunoura, T. Unveiling the rna virosphere associated with marine microorganisms. Mol. Ecol. Res. 2018, 18, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Steward, G.F.; Culley, A.I.; Mueller, J.A.; Wood-Charlson, E.M.; Belcaid, M.; Poisson, G. Are we missing half of the viruses in the ocean? ISME J. 2012, 7, 672. [Google Scholar] [CrossRef] [PubMed]
- Kalanetra, K.M.; Bano, N.; Hollibaugh, J.T. Ammonia-oxidizing archaea in the arctic ocean and antarctic coastal waters. Environ. Microb. 2009, 11, 2434–2445. [Google Scholar] [CrossRef]
- Galand, P.E.; Casamayor, E.O.; Kirchman, D.L.; Potvin, M.; Lovejoy, C. Unique archaeal assemblages in the arctic ocean unveiled by massively parallel tag sequencing. ISME J. 2009, 3, 860. [Google Scholar] [CrossRef]
- Wang, H.; Yu, Y.; Liu, T.; Pan, Y.; Yan, S.; Wang, Y. Diversity of putative archaeal rna viruses in metagenomic datasets of a yellowstone acidic hot spring. SpringerPlus 2015, 4, 189. [Google Scholar] [CrossRef]
- Shtarkman, Y.M.; Koçer, Z.A.; Edgar, R.; Veerapaneni, R.S.; D’Elia, T.; Morris, P.F.; Rogers, S.O. Subglacial lake vostok (antarctica) accretion ice contains a diverse set of sequences from aquatic, marine and sediment-inhabiting bacteria and eukarya. PLoS ONE 2013, 8, e67221. [Google Scholar] [CrossRef]
- Siegert Martin, J.; Priscu John, C.; Alekhina Irina, A.; Wadham Jemma, L.; Lyons, W.B. Antarctic subglacial lake exploration: First results and future plans. Philos. Trans. Royal Soc. A 2016, 374, 20140466. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yau, S.; Seth-Pasricha, M. Viruses of Polar Aquatic Environments. Viruses 2019, 11, 189. https://doi.org/10.3390/v11020189
Yau S, Seth-Pasricha M. Viruses of Polar Aquatic Environments. Viruses. 2019; 11(2):189. https://doi.org/10.3390/v11020189
Chicago/Turabian StyleYau, Sheree, and Mansha Seth-Pasricha. 2019. "Viruses of Polar Aquatic Environments" Viruses 11, no. 2: 189. https://doi.org/10.3390/v11020189
APA StyleYau, S., & Seth-Pasricha, M. (2019). Viruses of Polar Aquatic Environments. Viruses, 11(2), 189. https://doi.org/10.3390/v11020189