
Disturbance Agents and Their Associated Effects on the Health of Interior Douglas-Fir Forests in the Central Rocky Mountains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
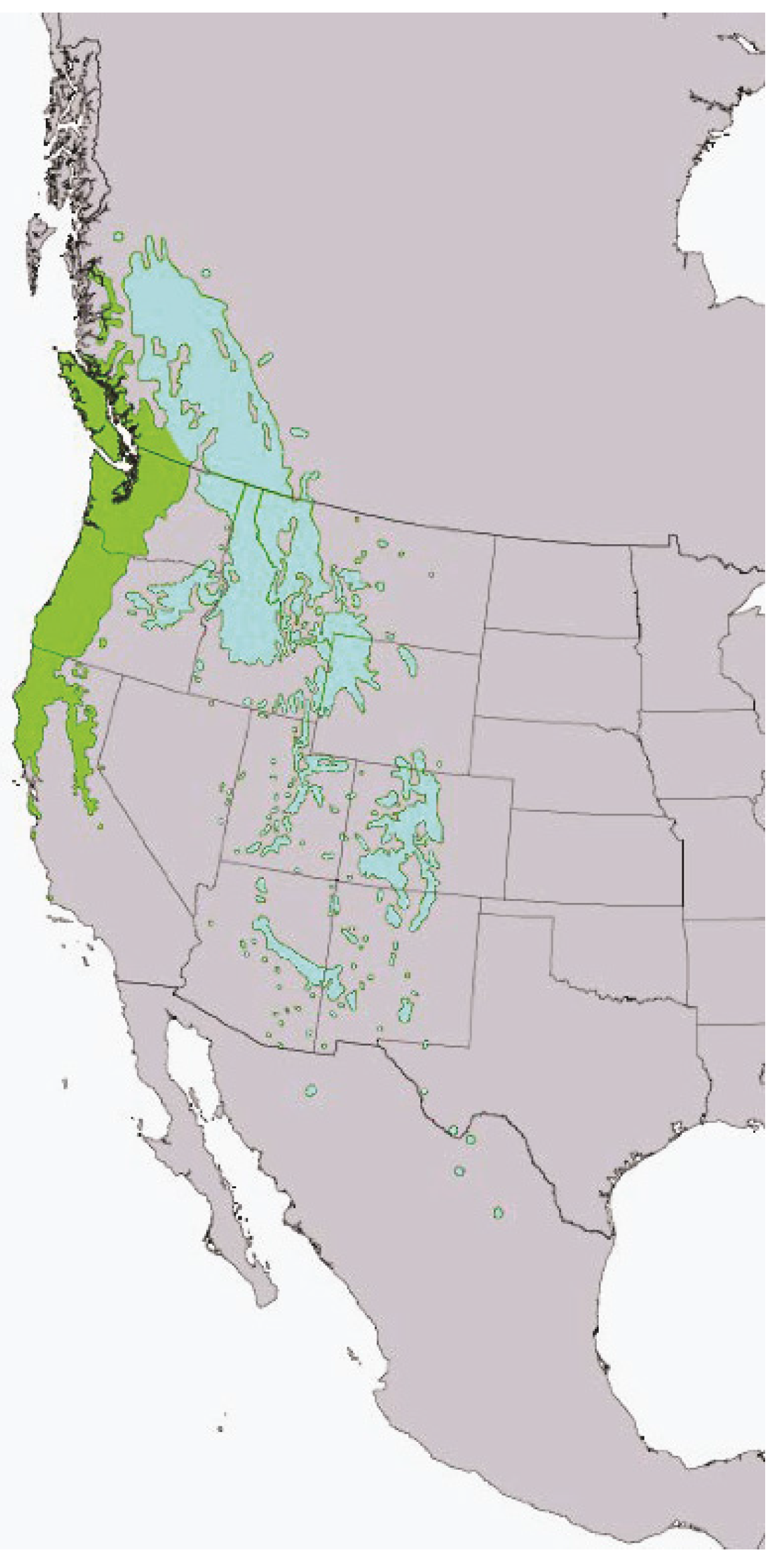
2. Interior Douglas-Fir Forests
3. Abiotic Disturbance Agents
3.1. Fire
3.1.1. Direct Fire Effects
3.1.2. Indirect Fire Effects
3.2. Additional Abiotic Disturbances
4. Biotic Disturbances
4.1. Douglas-Fir Beetle
4.2. Western Spruce Budworm
4.3. Douglas-Fir Dwarf Mistletoe
4.4. Root Diseases
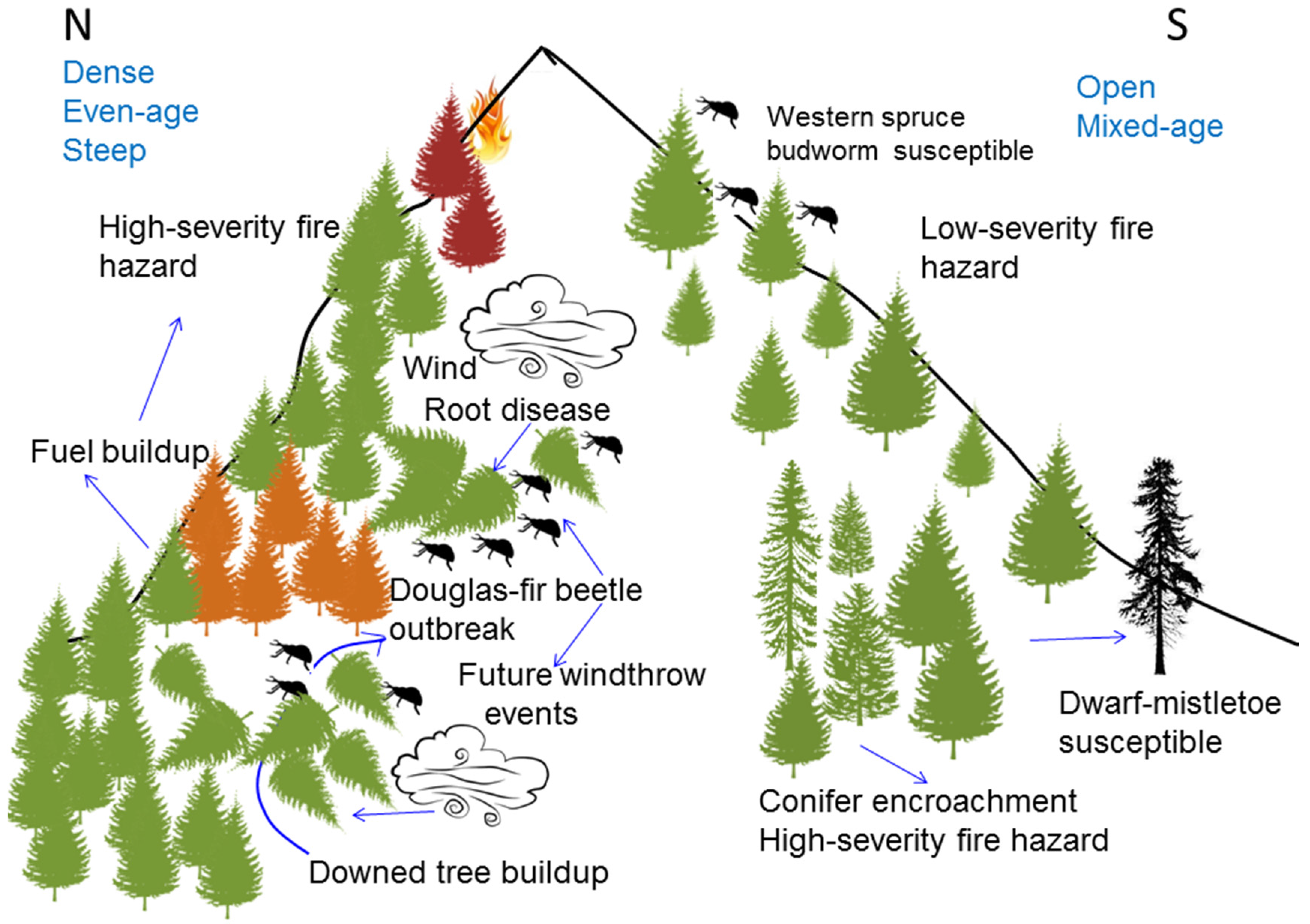
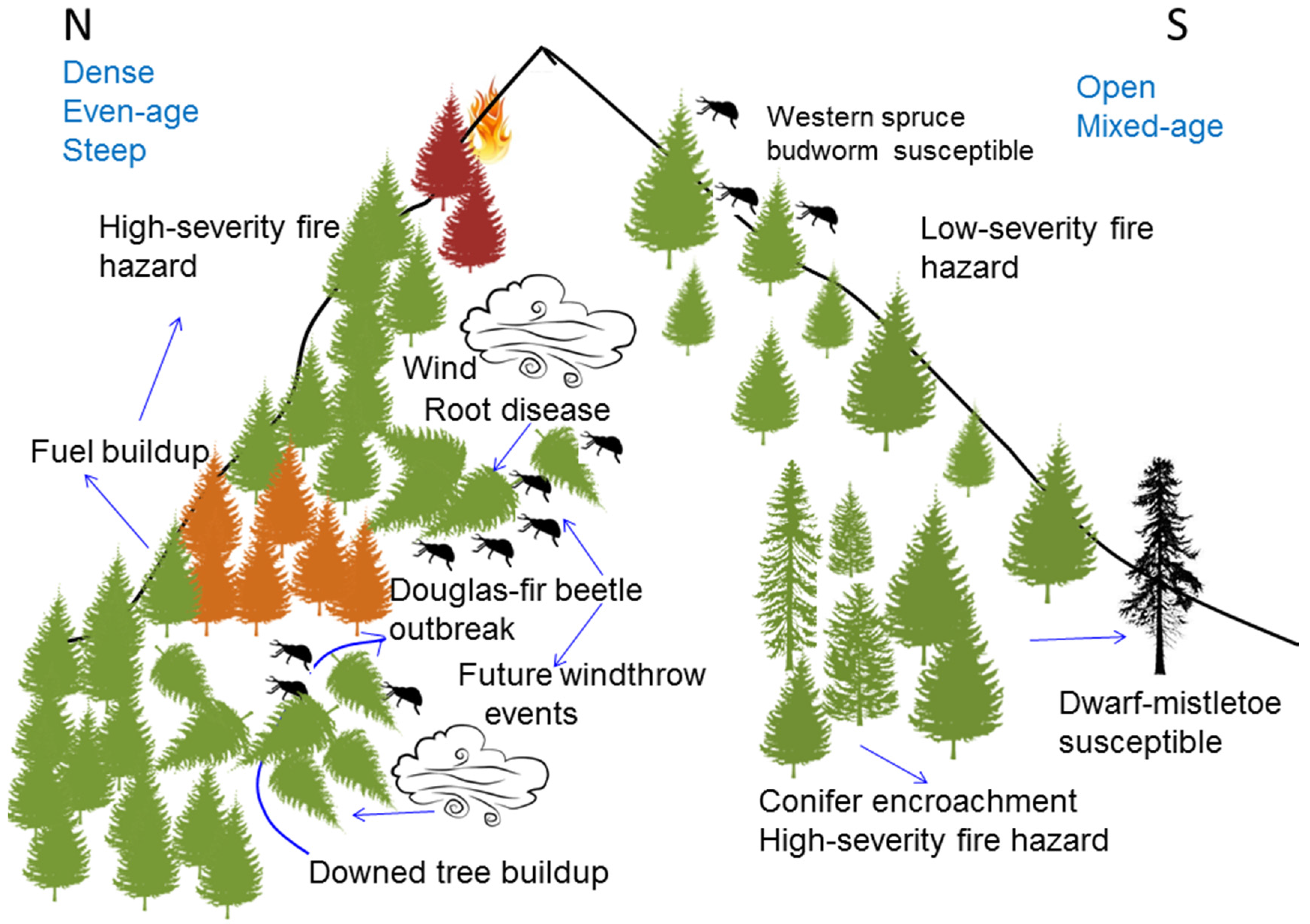
5. Disturbance Interactions
5.1. Fire and Douglas-Fir Beetle
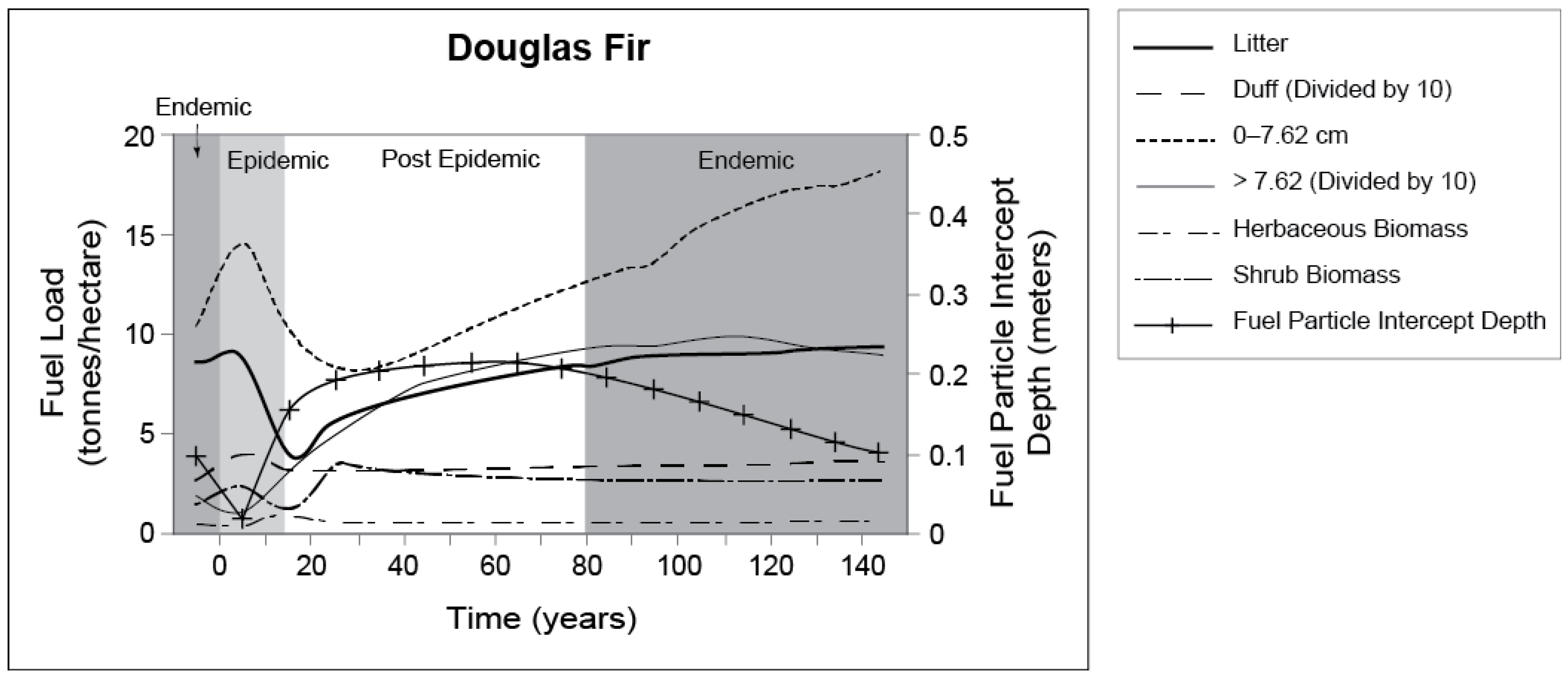
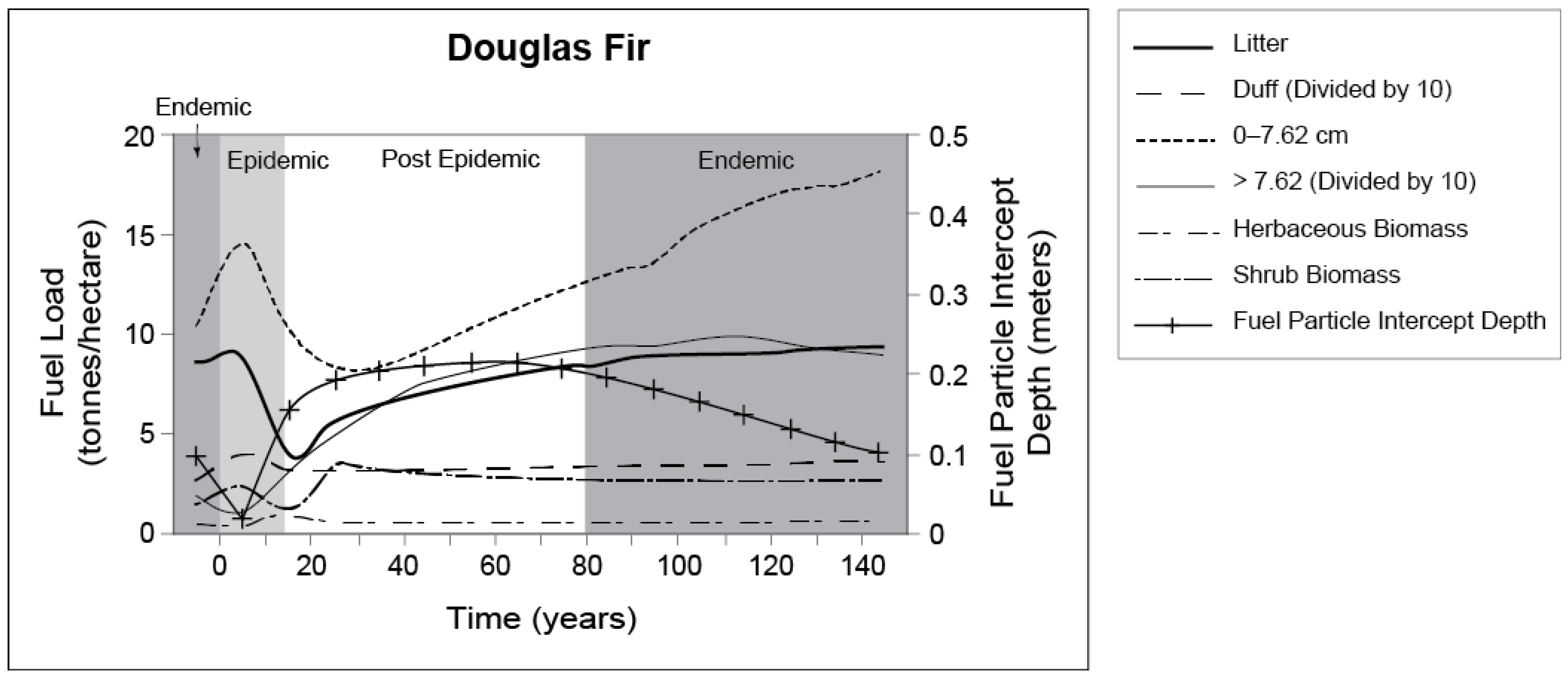
5.2. Douglas-Fir Beetle and Forest Fuel Changes
5.3. Western Spruce Budworm and Fire
5.4. Dwarf Mistletoe, Fire, and Other Disturbances
5.5. Western Spruce Budworm and Douglas-Fir Beetle
5.6. Root Disease, Windthrow, and Douglas-Fir Beetle
6. Anthropogenic Influences
6.1. Historic Human Mediated Disturbance
6.2. Forest Restoration
6.2.1. Pre-Fire Restoration Treatments
6.2.2. Fire Restoration Treatments
6.2.3. Post-Fire Restoration Treatments
7. Future Research Needs
- (1)
- Forest conditions including “stand structure, fuel characteristics, and fire regimes” have been greatly altered in part due to management practices that started in the 20th century, where “forest structure, fuel characteristics, and fire regimes of the mixed-conifer forests in the western United States have been dramatically altered” [248,249] (p. 22). One important issue is understanding the historic role and extent of fire in these ecosystems. The present accumulations of live and dead fuels have resulted in a shift toward higher-severity fire behavior. Although high-severity crown fires are not outside the historical range of variability for this fire regime type, the frequency and overall size of these types of fires appear to be increasing [66]. Additional fire reconstruction studies across various geographic scales can help elucidate the natural fire regimes of interior montane forests [231,250,251,252].
- (2)
- The inherent variability of interior Douglas-fir stands occurring on sites ranging from xeric to mesic and from lower-montane to subalpine zones necessitates that forest managers have a better understanding of how geographic locales influence forest fuel conditions. As stand composition and structure have shifted, so too have changes to fuel complexes.
- (3)
- Research is needed to determine how forest insects, particularly bark beetles and defoliators, affect fuels and fire behavior across a wider range of geographic locales and whether bark beetle alterations of surface and canopy fuels can elevate the potential for fire to spread into upper elevation forests.
- (4)
- Little is known about the combined effects of DFB and WSBW on interior Douglas-fir forests and, in turn, on ecosystem values across various spatial scales.
- (5)
- Information regarding climate change influences on the occurrence, timing, frequency, extent, and duration of disturbances at various temporal and spatial scales for interior Douglas-fir forests is lacking.
- (6)
- A greater understanding of DFB population dynamics in interior Douglas-fir stands is needed.
- (7)
- Douglas-fir dwarf mistletoe and DFB interactions are poorly understood.
- (8)
- Research on how snow avalanches and other abiotic disturbances affect interior Douglas-fir forest health is lacking.
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oliver, C.D. Forest development in North America following major disturbances. For. Ecol. Manag. 1981, 3, 153–168. [Google Scholar] [CrossRef]
- Delcourt, H.R.; Delcourt, P.A.; Webb, T., III. Dynamic plant ecology: The spectrum of vegetational change in space and time. Quat. Sci. Rev. 1983, 1, 153–175. [Google Scholar] [CrossRef]
- White, P.S.; Pickett, S.T.A. Natural disturbance and patch dynamics: An introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 3–13. [Google Scholar]
- Parsons, D.J.; DeBenedetti, S.H. Impact of fire suppression on a mixed-conifer forest. For. Ecol. Manag. 1979, 2, 21–33. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 894–908. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Cocke, A.E.; Fulé, P.Z.; Crouse, J.E. Forest change on a steep mountain gradient after extended fire exclusion: San Francisco Peaks, Arizona, USA. J. Appl. Ecol. 2005, 42, 814–823. [Google Scholar] [CrossRef]
- Hejl, S.J.; Hutto, R.L.; Preston, C.R.; Finch, D.M. Effects of silvicultuer treatments in the Rocky Mountains. In Ecology and Management of Neotropical Migratory Birds: A Synthesis and Review of Critical Issues, 1st ed.; Martin, T.E., Finch, D.M., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 220–244. [Google Scholar]
- Bentz, B. Bark beetle outbreaks in western North America. Causes and consequences. In Proceedings of the Bark Beetle Symposium, Snowbird, UT, USA, 15–17 November 2005.
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Hansen, M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Shaw, J.D.; Steed, B.E.; DeBlander, L.T. Forest inventory and analysis (FIA) annual inventory answers the question: What is happening to pinyon-juniper woodlands? J. For. 2005, 103, 280–285. [Google Scholar]
- Kleinman, S.J.; DeGomez, T.E.; Snider, G.B.; Williams, K.E. Large-scale pinyon ips (Ips confusus) outbreak in the southwestern United States tied with elevation and land cover. J. For. 2012, 110, 194–200. [Google Scholar] [CrossRef]
- Wulder, M.A.; White, J.C.; Bentz, B.; Alvarez, M.F.; Coops, N.C. Estimating the probability of mountain pine beetle red-attack damage. Remote Sens. Environ. 2006, 101, 150–166. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and carbon forest feeback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.J.; Hebertson, E.G.; Munson, A.S. Spruce beetle biology, ecology and management in the Rocky Mountains: An addendum to spruce beetle in the Rockies. Forests 2014, 5, 21–71. [Google Scholar] [CrossRef]
- Page, W.G.; Jenkins, M.J.; Runyon, J.B. Spruce beetle-induced changes to Engelmann spruce foliage flammability. For. Sci. 2014, 60, 691–702. [Google Scholar] [CrossRef]
- Schmitz, R.F.; Gibson, K.E. Douglas-Fir Beetle; Forest and Insect Disease Leaflet; USDA Forest Service: Washington, DC, USA, 1996; pp. 5–7.
- Negrón, J.F.; Anhold, J.A.; Munson, S.A. Within-stand spatial distribution of tree mortality caused by the Douglas-fir beetle (Coleoptera: Scolytidae). Environ. Entomol. 2001, 30, 215–224. [Google Scholar] [CrossRef]
- Kolb, T.E.; Wagner, M.R.; Covington, W.W. Forest health from different perspectives, forest health through silviculture. In Proceedings of the 1995 National Silviculture Workshop, Mescalaro, NM, USA, 8–11 May 1995; pp. 5–13.
- Healthy Forest Restoration Act. “HR 1904”. PL108-148; U.S. Department of Agriculture, 2003. Available online: http://www.fs.fed.us/emc/applit/includes/hfr2003.pdf (accessed on 2 March 2016).
- Coulson, R.N.; Stephens, F.M. Impacts of insects in forest landscapes: Implications for forest health management. In Invasive Forest Insects, Introduced Forest Trees, and Altered Ecosystems: Ecological Pest Management in Global Forests of a Changing World; Payne, T.D., Ed.; Springer-Verlag: New York, NY, USA, 2006; pp. 101–125. [Google Scholar]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, S.A.; Nebeker, T.E.; Negrón, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2003, 84, 362–371. [Google Scholar]
- Kulakowski, D.; Veblen, T.T.; Bebi, P. Effects of fire and spruce beetle outbreak legacies on the disturbance regime of a subalpine forest in Colorado. J. Biogeogr. 2003, 30, 1445–1456. [Google Scholar] [CrossRef]
- Kulakowski, D.; Veblen, T.T. Effects of prior disturbances on the extent and severity of wildfire in Colorado subalpine forests. Ecology 2007, 88, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Kulakowski, D.; Veblen, T.T. Multiple disturbance interactions and drought influence fire severity in Rocky Mountain subalpine forests. Ecology 2005, 86, 3018–3029. [Google Scholar] [CrossRef]
- Jorgensen, C.A.; Jenkins, M.J. Fuel complex alterations associated with spruce beetle-induced tree mortality in intermountain spruce-fir forests, USA. For. Sci. 2011, 57, 232–240. [Google Scholar]
- O’Connor, C.D.; Lynch, A.M.; Falk, D.A.; Swetnam, T.W. Post-fire forest dynamics and climate variability affect spatial and temporal properties of spruce beetle outbreaks on a Sky Island mountain range. For. Ecol. Manag. 2015, 336, 148–162. [Google Scholar] [CrossRef]
- Kulakowski, D.; Jarvis, D.; Veblen, T.T.; Smith, J. Stand replacing fires reduce susceptibility of lodgepole pine to mountain pine beetle outbreaks in Colorado. J. Biogeogr. 2012, 39, 2052–2060. [Google Scholar] [CrossRef]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia; British Columbia Ministry of Forests, 1991; p. 330. Available online: http://www.for.gov.bc.ca/hfd/pubs/Docs/Srs/SRseries.htm (accessed on 18 April 2015).
- Arno, S.F. Forest Regions of Montana; Research Paper INT-218; USDA Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1979; p. 39.
- Silen, R.R. Genetics of Douglas-Fir; Research Paper WO-35; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1978; p. 34.
- Simard, S.W. The foundational role of mycorrhizal networks in self-organization of interior Douglas-fir forests. For. Ecol. Manag. 2009, 258, 95–107. [Google Scholar] [CrossRef]
- Little, E.L., Jr. Atlas of United States Trees; Vol. 1, U.S. Department of Agriculture: Washington, DC, USA, 1971; pp. 1–9.
- Donato, D.C.; Harvey, B.J.; Romme, W.H.; Simard, M.; Turner, M.G. Bark beetle effects on fuel profiles across a range of stand structures in Douglas-fir forests of Greater Yellowstone. Ecol. Appl. 2013, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Fowells, H.A. Silvics of Forest Trees of the United States; No. 2171; USDA Forest Service Agriculture Handbook: Washington, DC, USA, 1965; pp. 546–556.
- Wright, J.W.; Kung, F.H.; Read, R.A.; Lemmien, W.A.; Bright, J.N. Genetic variation in Rocky Mountain Douglas-fir. Silv. Genet. 1971, 20, 54–60. [Google Scholar]
- Van Hooser, D.D.; Wadell, K.L.; Mills, J.R.; Tymcio, R.P. The interior Douglas-fir resource: Current status and projections to the year 2040. In Interior Douglas-Fir, the Species and Its Management, Proceedings of the Interior Douglas-fir: The Species and Its Management, Pullman, WA, USA, 27 February–1 March 1991; Baumgartner, D.M., Lotan, J.E., Eds.; Washington State University: Pullman, WA, USA, 1991; pp. 9–14. [Google Scholar]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina mountains Arizona. I. Ecological classification and distribution of species. J. Ariz. Acad. Sci. 1964, 3, 9–34. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina mountains, Arizona. V. biomass, production, and diversity along the elevation gradient. Ecology 1975, 56, 771–790. [Google Scholar] [CrossRef]
- Niering, W.A.; Lowe, C.H. Vegetation of the Santa Catalina Mountains, Arizona, USA: Community types and dynamics. Vegetatio 1984, 58, 3–28. [Google Scholar]
- Lavendar, D.P.; Hermann, R.K. Douglas-Fir: The Genus Pseudotsuga, 1st ed.; Oregon State University: Corvallis, OR, USA, 2014; pp. 19–24. [Google Scholar]
- Hermann, R.K.; Lavendar, D.P. Pseudotsuga menziesii (Mirb.) Franco Douglas-fir. In Silvics of North American Conifers; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 527–540. [Google Scholar]
- Kilgore, B.M. Fire in ecosystem distribution and structure: Western forests and scrublands. In Fire Regimes and Ecosystem Properties; General Technical Report WO-GTR-26; Mooney, H.A., Bonnickson, T.M., Christensen, N.L., Lotan, J.E., Reiners, W.A., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1981; pp. 58–89. [Google Scholar]
- Bailey, R.G. Descriptions of the Ecoregions of the United States; Miscellaneous Publication 1391; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1980; p. 77.
- Eyre, F.H. Forest Cover Types of the United States and Canada, 1st ed.; Society of American Foresters: Washington, DC, USA, 1980; p. 148. [Google Scholar]
- Harlow, W.M.; Harrrar, E.S.; White, F.M. Textbook of Dendrology: Covering the Important Forest Trees of the United States and Canada, 6th ed.; McGraw-Hill: New York, NY, USA, 1979; p. 510. [Google Scholar]
- Habeck, J.R.; Mutch, R.W. Fire dependent forests in the northern Rocky Mountains. Quat. Res. 1973, 3, 408–424. [Google Scholar] [CrossRef]
- Mauk, R.L.; Henderson, J.A. Coniferous Forest Habitat Types of Northern Utah; General Technical Report INT-170; U.S. Department of Agriculture, Forest Service, Intermountain Range and Experiment Station: Ogden, UT, USA, 1984.
- Youngblood, A.P.; Mauk, R.L. Coniferous Forest Habitat Types of Central and Southern Utah; General Technical Report INT-187; Department of Agriculture, Forest Service, Intermountain Range and Experiment Station: Ogden, UT, USA, 1985.
- Keegan, C.E.; Swanson, L.D.; Wichman, D.P.; VanHooser, D.D. Montana’s Forest Products Industry: A Descriptive analysis 1969–1988; Bureau of Business and Economic Research: Missoula, MT, USA, 1990; p. 52. [Google Scholar]
- Lanner, R.M. Trees of the Great Basin—A Natural History, 1st ed.; University of Nevada Press: Reno, NV, USA, 1984; p. 215. [Google Scholar]
- Lilleholm, R.J.; Kessler, W.B.; Merrill, K. Stand density index applied to timber and goshawk habitat objectives in Douglas-fir. Environ. Manag. 1993, 17, 773–779. [Google Scholar] [CrossRef]
- McKenzie, D.; Gedalof, Z.; Peterson, D.L.; Mote, P. Climate change, wildfire, and conservation. Conserv. Biol. 2004, 18, 890–902. [Google Scholar] [CrossRef]
- Hicke, J.A.; Johnson, M.C.; Hayes, J.L.; Preisler, H.K. Effects of bark beetle-caused tree mortality on wildfire. For. Ecol. Manag. 2012, 271, 81–90. [Google Scholar] [CrossRef]
- Jenkins, M.J.; Page, W.G.; Hebertson, E.G.; Alexander, M.E. Fuels and fire behavior dynamics in bark beetle attacked forests in western North America and implications for fire management. For. Ecol. Manag. 2012, 275, 23–34. [Google Scholar] [CrossRef]
- Pyne, S.J.; Andrews, P.L.; Laven, R.D. Introduction to Wildland Fire, 2nd ed.; Wiley: New York, NY, USA, 1996; p. 769. [Google Scholar]
- Brown, J.K. Introduction and fire regimes. In Wildland Fire in Ecosystems: Effects on Fire on Flora; Brown, J.K., Smith, J.K., Eds.; General Technical Report RMRS-42; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000. [Google Scholar]
- Morgan, P.; Hardy, C.C.; Swetnam, T.W.; Rollins, M.G.; Long, D.G. Mapping fire regimes across time and space: Understanding coarse and fine-scale fire patterns. Int. J. Wildland Fire 2001, 10, 329–324. [Google Scholar] [CrossRef]
- Halofsky, J.E.; Donato, D.C.; Hibbs, D.E.; Cambell, J.L.; Cannon, M.D.; Fontaine, J.B.; Thompson, J.R.; Anthony, R.G.; Bormann, B.T.; Kayes, L.J.; et al. Mixed-severity fire regimes: Lessons and hypotheses from Klamath-Siskiyou Ecoregion. Ecosphere 2011, 2, 1–19. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Van Wagner, C.E. Conditions for the start and spread of crown fire. Can. J. For. Res. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Schoennagel, T.; Veblen, T.T.; Romme, W. Interaction of fire, fuels, and climate across Rocky Mountain forests. BioScience 2004, 54, 661–676. [Google Scholar] [CrossRef]
- Chappell, C.B.; Agee, J.K. Fire severity and tree seedling establishment in Abies magnifica forests, southern Cascades, Oregon. Ecol. Appl. 1996, 6, 628–640. [Google Scholar] [CrossRef]
- Agee, J.K. The complex nature of mixed-severity fire regimes. In Mixed Severity Fire Regimes: Ecology and Management; Lagene, L., Zelnik, J., Cadwallader, S., Hughes, B., Eds.; Washington State University: Pullman, WA, USA; The Association for Fire Ecology: Spokane, WA, USA, 2004; pp. 1–10. [Google Scholar]
- Arno, S.F.; Parsons, D.J.; Keane, R.E. Mixed-Severity Fire Regimes in the Northern Rocky Mountains: Consequences of Fire Exclusion and Options for the Future; Research Paper RMRS-P-15-VOL-5; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; pp. 225–232.
- Brown, J.K. Fire regims and their relevance to ecosystem manaagment. In Proceedings of the Society of American Foresters National Convention, Anchorage, AK, USA, 18–22 September 1994; Society of American Foresters: Bethesda, MD, USA, 1995; pp. 171–178. [Google Scholar]
- Baker, W.L.; Veblen, T.T.; Sherriff, R.L. Fire, fuels, and restoration of ponderosa pine-Douglas-fir forests in the Rocky Mountains, USA. J. Biogeogr. 2007, 34, 251–269. [Google Scholar] [CrossRef]
- Jain, T.B.; Battaglia, M.A.; Han, H.S.; Graham, R.T.; Keyes, C.R.; Fried, J.S.; Sandquist, J.E. A Comprehensive Guide to Fuel Management Practices for Dry Mixed Conifer Forests in the Northwestern United States: Inventory and Model-Based Economic Analysis of Mechanical Fuel Treatments; Research Note RMRS-RN-64; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2014.
- Romme, W.H.; Floyd, M.L.; Hanna, D. Historical Range of Variability and Current Landscape Condition Analysis: South Central Highlands Section, Southwestern Colorado & Northwestern New Mexico; Colorado Restoration Institute, Colorado State University: Fort Collins, CO, USA; Region 2 U.S. Department of Agriculture, Forest Service: Golden, CO, USA, 2009; Available online: http://www.coloradoforestrestoration.org/CFRIpdfs/2009_HRVSouthCentral-Highlands.pdf (accessed on 1 March 2016).
- Wyant, J.G.; Omni, P.N.; Laven, R.D. Fire induced tree mortality in a Colorado ponderosa pine/Douglas-fir stand. For. Sci. 1986, 32, 49–59. [Google Scholar]
- Stephens, S.L.; Finney, M.A. Prescribed fire mortality of Sierra Nevada mixed conifer tree species: Effects of crown damage and forest floor combustion. For. Ecol. Manag. 2002, 162, 261–271. [Google Scholar] [CrossRef]
- Van Wagner, C.E. Height of crown scorch in forest fires. Can. J. For. Res. 1973, 3, 373–378. [Google Scholar] [CrossRef]
- Methven, I.R. Prescribed Fire, Crown Scorch and Mortality: Field and Laboratory Studies on Red and White Pine; Research Report PS-X-31; Canadian Forest Service, Petawawa Forest Experimental Station: Chalk River, ON, Canada, 1971; p. 10. [Google Scholar]
- Peterson, D.L. Crown scorch volume and scorch height: Estimates of post-fire tree condition. Can. J. For. Res. 1985, 15, 596–598. [Google Scholar] [CrossRef]
- Chambers, J.L.; Dougherty, P.M.; Hennessey, T.C. Fire: Its effects on growth and physiological processes in conifer forests. In Stress Physiology and Forest Productivity, 1st ed.; Hennessey, T.C., Dougherty, P.M., Kossuth, S.V., Johnson, J.D., Eds.; Martinaus Nijhoff: Hingham, MA, USA, 1986; pp. 171–191. [Google Scholar]
- Peterson, D.L.; Ryan, K.C. Modeling postfire conifer mortality for long-range planning. Environ. Manag. 1986, 10, 797–808. [Google Scholar] [CrossRef]
- Ryan, K.C.; Reinhardt, E.D. Predicting post fire severity in seven western conifers. Can. J. For. Res. 1988, 18, 1291–1297. [Google Scholar] [CrossRef]
- Peterson, D.L.; Arbaugh, M.J. Postfire survival in Douglas-fir and lodgepole pine: Comparing the effects of crown and bole damage. Can. J. For. Res. 1986, 16, 1175–1179. [Google Scholar] [CrossRef]
- Costa, J.J.; Oliveria, L.A.; Viegas, D.X.; Neto, L.P. On the temperature distribution inside a tree under fire conditions. Int. J. Wildland Fire 1991, 1, 87–96. [Google Scholar] [CrossRef]
- Hood, S.; Bentz, B. Predicting postfire Douglas-fir beetle attacks and tree mortality in the northern Rocky Mountains. Can. J. For. Res. 2007, 37, 1058–1069. [Google Scholar] [CrossRef]
- Hungerford, R.D.; Harrington, M.G.; Frandsen, W.H.; Ryan, K.C.; Niehoff, G.J. Influence of fire on factors that affect site productivity. In Proceedings of the Management and Productivity of Western-Montane Forest Soils, Boise, ID, USA, 10–12 April 1990; Harvey, A.E., Neuenschwander, L.F., Eds.; General Technical Report INT-280. U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1991; pp. 32–50. [Google Scholar]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on below ground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Egan, D. Protecting Old Trees from Prescribed Burning; Working Paper No. 24; Ecological Restoration Institute: Flagstaff, AZ, USA, 2011; p. 5. [Google Scholar]
- Swezy, D.M.; Agee, J.K. Prescribed fire effects on fine-root and tree mortality in old-growth ponderosa pine. Can. J. For. Res. 1991, 21, 626–634. [Google Scholar] [CrossRef]
- Stendell, E.; Horton, T.R.; Bruns, T.D. Early effects of prescribed fire on the structure of the fungal ectomycorrhizal community in a Sierra Nevada ponderosa pine forest. Mycol. Res. 1999, 103, 1353–1359. [Google Scholar] [CrossRef]
- Hart, S.C.; Classen, A.T.; Wright, R.J. Long-term interval burning alters fine root and mycorrhizal dynamics in a ponderosa pine forest. J. Appl. Ecol. 2005, 42, 752–761. [Google Scholar] [CrossRef]
- Rogstad, A. Recovering From Wildfire a Guide to Arizona’s Forest Owners. 2002. Available online: http://ag.arizona.edu/pubs/natresources/az1294/ (accessed on 1 March 2016).
- Gordon, W.S.; Jackson, R.B. Nutrient concentrations in fine roots. Ecology 2000, 81, 275–280. [Google Scholar] [CrossRef]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Gyssels, G.; Poesen, J.; Bochet, E.; Li, Y. Impact of plant roots on the resistance of soils to erosion by water: A review. Prog. Phys. Geogr. 2005, 29, 189–217. [Google Scholar] [CrossRef]
- Lertzman, K.; Riccius, P.; Fall, J. Does fire isolate elements of the landscape mosaic? Bull. Ecol. Soc. Am. 1997, 78, 92. [Google Scholar]
- Brown, J.K. The unnatural fuel build up issue. In Proceedings of the Symposium and Workshop on Wilderness Fire, Missoula, MT, USA, 15–18 November 1983; Lotan, J.E., Kilgore, B.M., Fischer, W.C., Mutch, R.W., Eds.; General Technical Report INT-182. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1983; pp. 127–128. [Google Scholar]
- Turner, M.G.; Romme, W.H. Landscape dynamics in crown fire ecosystems. Landsc. Ecol. 1994, 9, 59–77. [Google Scholar] [CrossRef]
- Agee, J.K. The landscape ecology of western forest fire regimes. Northwest Sci. 1998, 72, 24–34. [Google Scholar]
- Kaufmann, M.R.; Regan, C.M.; Brown, P.M. Heterogeneity in ponderosa pine/Douglas-fir forests: Age and size structure in unlogged and logged landscapes of central Colorado. Can. J. For. Res. 2000, 30, 698–711. [Google Scholar] [CrossRef]
- Covington, W.W.; Moore, M.M. Post settlement changes in natural fire regimes and forest structure: Ecological restoration of old-growth ponderosa pine forests. J. Sustain. For. 1994, 2, 153–181. [Google Scholar] [CrossRef]
- Heyerdahl, E.K.; Miller, R.F.; Parson, R.A. History of fire and Douglas-fir establishment in a savanna and sagebrush-grassland mosaic, southwestern Montana, USA. For. Ecol. Manag. 2006, 230, 107–118. [Google Scholar] [CrossRef]
- Daniels, L.D. Climate and fire: A case study of the Cariboo forest, British Columbia. In Proceedings of the Symposium on the Ecology and Management of Mixed Severity Fire Regimes Conference, Spokane, WA, USA, 17–19 November 2004; Taylor, L., Zelnik, J., Cadwallader, S., Hughes, B., Eds.; pp. 235–246.
- Schaetzl, R.F.; Burns, S.F.; Johnson, D.L.; Small, T.W. Tree uprotting: Review of impacts on forest ecology. Vegetatio 1989, 79, 65–176. [Google Scholar]
- Patten, R.S.; Knight, D. Snow avalanches and vegetation pattern in Cascade Canyon, Grand Teton National Park. Arct. Alp. Res. 1994, 26, 35–41. [Google Scholar] [CrossRef]
- Mitchel, S.J. A diagnostic framework for windthrow risk estimation. For. Chron. 1998, 74, 100–105. [Google Scholar] [CrossRef]
- Ulanova, N.G. The effects of windthrow on forests at different spatial scales. A review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Everham, E.M., III; Browkaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 114–175. [Google Scholar] [CrossRef]
- Mitchel, S.J. The windthrow triangle: A relative windthrow hazard assessment procedure for forest managers. For. Chron. 1995, 71, 446–450. [Google Scholar] [CrossRef]
- Beaty, S.W.; Stone, E.L. The variety of microsites created by tree falls. Can. J. For. Res. 1986, 16, 539–548. [Google Scholar] [CrossRef]
- Teich, M.; Bartlet, P.; Gret-Regamey, A.; Bebi, P. Snow avalanches in forested terrain: Influence of forest parameters, topography, and avalanche characteristics on runout distance. Arct. Antarct. Alp. Res. 2012, 44, 509–519. [Google Scholar] [CrossRef]
- Luckman, B.H. The geomorphic activity of snow avalanches. Geografiska Annaler. Series A. Phys. Geogr. 1977, 59, 31–48. [Google Scholar]
- McClung, D.; Schaerer, P. The Avalanche Handbook, 3rd ed.; The Mountainers Books: Seattle, WA, USA, 2006; p. 338. [Google Scholar]
- Burrows, C.L.; Burrows, V.L. Procedures for the study of snow avalanche chronology using growth layers of woody plants. Inst. Arct. Alp. Res. 1976, 23, 54. [Google Scholar]
- Hebertson, E.G. Snow Avalanche Disturbance in Intermountain Spruce-Fir Forests and Implications for the Spruce Bark Beetle (Coleoptera: Scolytidae). Ph.D. Thesis, Utah State University, Logan, UT, USA, 2004. [Google Scholar]
- Malanson, G.P.; Butler, D.R. Transverse pattern of vegetation on avalanche paths in the northern Rocky Mountains, Montana. Gt. Basin Nat. 1984, 44, 453–458. [Google Scholar]
- Oliver, C.D.; Adams, A.B.; Zasoski, R.J. Disturbance patterns and forest development in a recently deglaciated valley in northwestern Cascade range of Washington USA. Can. J. For. Res. 1984, 15, 221–232. [Google Scholar] [CrossRef]
- Butler, D.R.; Malanson, G.P. Non-equilibrium geomorphic processes and patterns on avalanche paths in the northern Rocky Mountains, U.S.A. Z. Geomorphol. 1990, 34, 257–270. [Google Scholar]
- Walsh, S.J.; Butler, D.R.; Allen, T.R.; Malanson, G.P. Influence of snow patterns and snow avalanches on the alpine treeline ecotone. J. Veg. Sci. 1994, 5, 657–672. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Donnegan, J. Climatic and human influences on fire regimes in ponderosa pine forests in the Colorado Front Range. Ecol. Appl. 2000, 10, 1178–1195. [Google Scholar] [CrossRef]
- Bebi, P.; Kulakowski, D.; Rixen, C. Snow avalanche disturbances in forest ecosystems-State of research and implications for management. For. Ecol Manag. 2009, 257, 1883–1892. [Google Scholar] [CrossRef]
- McGregor, M.D.; Furniss, M.M.; Oaks, R.D.; Gibson, K.E.; Meyer, H.E. MCH Pheromone for preventing Douglas-fir beetle infestation in windthrown trees. J. For. 1984, 82, 613–616. [Google Scholar]
- Blackford, D.C. Aspen Grove Trailhead Area Pleasant Grove RD, Uinta, NF; Report No. OFO-TR-05-15; U.S. Department of Agriculture, Forest Service Forest Health Protection: Ogden, UT, USA, 2005.
- Blackford, D.C.; Guyon, J. Recreation Sites Insect and Disease Activity Duchesne/Roosevelt RD, Ashley NF, Functional Assistance Trip Report; Report No. OFO-TR-06-05; U.S. Department of Agriculture, Forest Service, Forest Health Protection: Ogden, UT, USA, 2005.
- Hebertson, E.G. BLM Richfield Office-Henry Mountains Field Station Functional Assistance Visit; Report No. OFO-TR-06-03; U.S. Department of Agriculture, Forest Service, Forest Health Protection: Ogden, UT, USA, 2005.
- Raffa, K.F.; Berryman, A.A. Interacting selective pressure in conifer-bark beetle systems: A basis for reciprocal adaptations? Am. Nat. 1987, 129, 234–262. [Google Scholar] [CrossRef]
- Samman, S.; Logan, J. Assessment and Response to Bark Beetle Outbreaks in the Rocky Mountain Area; Report to Congress from Forest Health Protection, Washington Office, Forest Service, U.S. Department of Agriculture; General Technical Report RMRS-GTR-62; U.S. Department of Agricutlure Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; p. 46.
- Hardwood, W.G.; Rudinsky, J.A. The Flight and Olfactory Behavior of Checkered Beetles (Coleoptera: Cleridae) Predatory on the Douglas-Fir Beetle; Technical Bulletin 95; Agriculture Experiment Station, Oregon State University: Corvallis, OR, USA, 1966; p. 36. [Google Scholar]
- Jenkins, M.J.; Hebertson, E.; Page, W.G.; Jorgensen, C.A. Bark beetles, fuels, fires and implications for forest management in the Intermountain West. For. Ecol. Manag. 2008, 254, 16–34. [Google Scholar] [CrossRef]
- Anderson, M.N. Mechanisms of odor coding in coniferous bark beetles: From neuron to behavior and application. Psyche J. Entomol. 2012. [Google Scholar] [CrossRef]
- Furniss, M.M.; McGregor, M.D.; Foiles, M.W.; Patridge, M.D. Chronology and Characteristics of a Douglas-Fir Beetle [Dendroctonus pseudotsugae] Outbreak in Northern Idaho [Pseudotsuga menziesii]; General Technical Report, INT-59; U.S. Department of Agriculture, Forest Service, Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1979. Available online: http://agris.fao.org/agris-search/search.do?recordID=US7935406 (accessed on 1 March 2016).
- Negrón, J.F. Probability of infestation and extent of mortality associated with the Dogulas-fir beetle in Colorado Front Range. For. Ecol. Manag. 1998, 107, 71–85. [Google Scholar] [CrossRef]
- Rudinsky, J.A. Scolytid beetles associated with Douglas-fir: Response to terpenes. Science 1966, 152, 218–219. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.; Daterman, G. Using pheromone-baited traps to control the amount and distribution of tree mortality during outbreaks of the Douglas-fir beetle. For. Sci. 1997, 43, 65–70. [Google Scholar]
- Furniss, M.M. The Douglas-fir beetle in western forests a historical perspective—Part 1. Am. Entomol. 2014, 60, 84–96. [Google Scholar] [CrossRef]
- Lejeune, R.; McMullen, L.; Atkins, M. Influence of logging on Douglas fir beetle populations. For. Chron. 1961, 37, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Furniss, M.M. Susceptibility of fire-injured Douglas-fir to bark beetle attack in southern Idaho. J. For. 1965, 63, 8–11. [Google Scholar]
- Lewisohn, E.; Gijzen, M.; Croteau, R. Defense mechanisms of conifers. Differences in constitutive and wound-induced monoterepene biosynthesis among species. Plant Physiol. 1991, 96, 44–49. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Cates, R.G.; Alexander, H. Host resistance and susceptibility. In Bark Beetles in North American Conifers, A System for the Study of Evolutionary Biology, 1st ed.; Mitton, J.B., Sturgeon, K.B., Eds.; University of Texas Press: Austin, TX, USA, 1982; pp. 212–263. [Google Scholar]
- Goheen, D.J.; Hansen, E. Effects of pathogens and bark beetles on forests. In Beetle-Pathogen Interactions in Conifer Forests, 1st ed.; Schowalter, T.D., Filip, G.M., Eds.; Academic Press: London, UK, 1993; pp. 175–196. [Google Scholar]
- Mattson, W.; Haack, R. Role of drought in outbreaks of plant-eating insects. Bioscience 1987, 37, 110–118. [Google Scholar] [CrossRef]
- Cunningham, C.; Jenkins, M.; Roberts, D. Attack and brood production by the Douglas-fir beetle Dendroctonus pseudotsugae (Coleoptera: Scolytidae) in Douglas-fir, Pseudotsuga menziesii var. glauca (Pinaceae), following a wildfire. West. N. Am. Nat. 2005, 65, 70–79. [Google Scholar]
- McGregor, M.; Furniss, M.M.; Bousfield, W.E.; Almas, D.P.; Gravelle, P.J.; Oakes, R.D. Evaluation of the Douglas-Fir Beetle Infestation North Fork Clearwater River Drainage, Northern Idaho, 1970–73; Report No. 74-7; U.S. Department of Agriculture, Forest Service, Northern Region State and Private Forestry, Forest Pest Management: Missoula, MT, USA, 1974; p. 17.
- Fredericks, S.E.; Jenkins, M.J. Douglas-fir beetle {Dendroctonus pseudotsugae Hopkins, Coleoptera: Scolytidae} brood production on Douglas-fir defoliated by western spruce budworm (Choristoneura occidentalis Freeman, Lepidoptera: Tortricidae) in Logan Canyon, UT. Gt. Basin Nat. 1988, 48, 348–351. [Google Scholar]
- McMullen, L.H.; Atkins, M.D. On the flight and host selection of the Douglas-fir beetle, Dendroctonus pseudotsugae Hopk. (Coleoptera: Scolytidae). Can. Entomol. 1961, 94, 1309–1325. [Google Scholar] [CrossRef]
- Wright, L.C.; Berryman, A.A.; Wickman, B.E. Abundance of the fir engraver, Scolytus ventralis, and the Douglas-fir beetle, Dendroctonus pseudotsugae, following tree defoliation by the Douglas-fir tussock moth, Orgyia seudotsugata. Can. Entomol. 1984, 116, 293–305. [Google Scholar] [CrossRef]
- Edmonds, R.L.; Eglitis, A. The role of Douglas-fir and wood borers in the decomposition of and nutrient release from Douglas-fir logs. Can. J. For. Res. 1989, 19, 853–859. [Google Scholar] [CrossRef]
- Kegley, S. Douglas-Fir Beetle Management; Forest Health Protection and State Forestry Organizations. Chapter 4; U.S. Department of Agriculture, Forest Service. Availabel online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187396.pdf (archived on 16 May 2015).
- Coulson, R.N. Population dynamics of bark beetles. Annu. Rev. Entomol. 1979, 24, 417–447. [Google Scholar] [CrossRef]
- Negrón, J.F.; Schaupp, W.C.; Gibson, K.E.; Anhold, J.; Hansen, D.; Their, R.; Mocettini, P. Estimating extent of mortality associated with the Douglas-fir beetle in the central and northern Rockies. West. J. Appl. For. 1999, 14, 121–127. [Google Scholar]
- Atkins, M.D. Behavioral variation among scolytids in relation to their habitat. Can. Entomol. 1966, 98, 285–288. [Google Scholar] [CrossRef]
- Lawrence, R. Early Detection of Douglas-fir beetle infestation with subcanopy resolution hyperspectral imagery. West. J. Appl. For. 2003, 18, 202–206. [Google Scholar]
- Weatherby, J.C.; Thier, R.W. A preliminary Validation of Douglas-Fir Beetle Hazard Rating System. Mountain Home Ranger District, Boise National Forest, 1992; Forest Pest Management Report No. R4-93-05; U.S. Department of Agriculture, Forest Service: Boise, ID, USA, 1993; p. 7.
- Kulman, H.M. Effects of insect defoliation on growth and mortality of trees. Annu. Rev. Entomol. 1971, 16, 289–324. [Google Scholar] [CrossRef]
- Brookes, M.H.; Colbert, J.J.; Mitchell, R.G.; Stark, R.W. Western Spruce Budworm; Technical Bulletin 1694; U.S. Department of Agriculture, Forest Service, Cooperative State Research Service: Washington, DC, USA, 1987; p. 108.
- Fellin, D.G.; Dewey, J.E. Western Spruce Budworm; Forest Insect and Disease Leaflet 53; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1982; p. 10.
- Swetnam, T.W.; Lynch, A.M. Multicentury, regional scale patterns of western spruce budworm outbreaks. Ecol. Monogr. 1993, 63, 399–424. [Google Scholar] [CrossRef]
- Myers, J.H. Synchrony in outbreaks of forest Lepidoptera: A possible example of the moran effect. Ecology 1998, 79, 1111–1117. [Google Scholar] [CrossRef]
- Ryerson, D.E.; Swetnam, T.W.; Lynch, A.M. A tree-ring reconstruction of western spruce budworm outbreaks in the San Juan Mountains, Colorado, U.S.A. Can. J. For. Res. 2003, 33, 1010–1028. [Google Scholar] [CrossRef]
- Murdock, T.Q.; Taylor, S.W.; Flower, A.; Mehlenbacher, A.; Montenegro, A.; Zwiers, F.W.; Alfaro, R.; Spittlehouse, D.L. Pest outbreak distribution and forest management impacts in a changing climate in British Columbia. Environ. Sci. Policy 2013, 6, 75–89. [Google Scholar] [CrossRef]
- Cates, R.G.; Redak, R.; Henderson, C.B. Natural product defensive chemistry of Douglas-fir, western spruce budworm success and forest management practices. J. Appl. Entomol. 1983, 96, 173–182. [Google Scholar]
- Wulf, N.; Cates, R. Site and stand characteristics. In Western Spruce Budworm; Technical Bulletin 1694; Brookes, M.H., Campbell, R.W., Colbert, J.J., Mitchell, R.G., Stark, R.W., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1987; pp. 90–115. [Google Scholar]
- Carlson, C.E.; Fellin, D.G.; Schmidt, W.C.; O’Laughlin, J.; Pfister, R.D. The western spruce budworm in northern Rocky Mountain forests: A review of ecology, insecticidal treatments and silvicultural practices. In Proceedings of the Symposium Management of Second-Growth Forests: The State of Knowledge and Research Needs, Missoula, MT, USA, 14 May 1982; O’Laughlin, J., Pfister, R.D., Eds.; pp. 76–103.
- Hadley, K.; Veblen, T. Stand response to western spruce budworm and Douglas-fir bark beetle outbreaks, Colorado Front Range. Can. J. For. Res. 1993, 23, 479–491. [Google Scholar] [CrossRef]
- Dewey, J.E. Damage to Douglas-fir cones by Choristoneura occidentalis. J. Econ. Entomol. 1970, 63, 1804–1806. [Google Scholar] [CrossRef]
- Frank, C.J.; Jenkins, M.J. Impact of the western spruce budworm on buds, developing cones and seeds of Douglas-fir in west-central Idaho. Environ. Entomol. 1987, 16, 304–308. [Google Scholar] [CrossRef]
- Owens, J.N. The Reproductive Cycle of Douglas-Fir; Research Paper BC-P-8; Canadian Forest Service Pacific Forest Research Center: Victoria, BC, Canada, 1976; p. 23. [Google Scholar]
- Lowry, W. Apparent meteorological requirements for abundant cone crops in Douglas-fir. For. Sci. 1966, 12, 185–192. [Google Scholar]
- Fellin, D.G.; Shearer, R.C.; Carlson, C.E. Western spruce budworm in the northern Rocky Mountains. West. Wildlands Nat. Res. J. 1983, 9, 2–7. [Google Scholar]
- Swetnam, T.W.; Thompson, M.A.; Sutherland, E.K. Using Dendrochronology to Measure Radial Growth of Defoliated Trees; Handbook No. 639; U.S. Department of Agriculture, Forest Service, Cooperative State Research Service: Washington, DC, USA, 1988; p. 39.
- Swetnam, T.W.; Lynch, A.M. A tree-ring reconstruction of western spruce budworm history in the southern Rocky Mountains. For. Sci. 1989, 35, 962–986. [Google Scholar]
- Alfaro, R.I.; Van Sickle, G.A.; Thomson, A.J.; Wegwitz, E. Tree mortality and radial growth losses caused by the western spruce budworm in a Douglas-fir stand in British Columbia. Can. J. For. Res. 1982, 12, 780–787. [Google Scholar] [CrossRef]
- Hawksworth, F.G.; Wiens, D. Dwarf Mistletoes: Biology, Pathology, and Systematics; Agricultural Handbook 709; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1996; p. 410.
- Watson, D.M. Mistletoe—A keystone resource in forests and woodlands worldwide. Annu. Rev. Ecol. Syst. 2001, 32, 219–249. [Google Scholar] [CrossRef]
- Hoffman, J.T. Management Guide for Dwarf Mistletoe Arceuthobium spp.; U.S. Department of Agriculture, Forest Service, 2010. Available online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187427.pdf (accessed on 28 May 2015).
- Smith, R.H. Xylem Resin in the Resistance of the Pinaceae to Bark Beetles; General Technical Report PSW-1; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1972; p. 7.
- Pate, J.S. Mineral relationships of parasites and their hosts. In Parasitic Plants, 1st ed.; Press, M.C., Graves, J., Eds.; Chapman & Hall: London, UK, 1995; pp. 80–102. [Google Scholar]
- Shaw, D.C.; Watson, D.M.; Mathiasen, R.L. Comparison of dwarf mistletoes (Arceuthobium spp., Viscaceae) in the western United States with mistletoes (Amyema spp., Loranhaceae) in Australia-ecological analogs and reciprocal models for ecosystem management. Aust. J. Bot. 2004, 52, 481–498. [Google Scholar] [CrossRef]
- Lamont, B. Mineral nutrition of mistletoes. In Biology of Mistletoes, 1st ed.; Calder, D.M., Bernhardt, P., Eds.; The Academic Press: Sydney, Australia, 1983; p. 348. [Google Scholar]
- Tinnin, R.O.; Hawksworth, F.G.; Knutson, D.M. Witches’ broom formation in conifers infected by Arceuthobium spp.: An example of parasitic impact upon community dynamics. Am. Midl. Nat. 1982, 107, 351–359. [Google Scholar] [CrossRef]
- Kuijt, J. The Biology of Parasitic Flowering Plants, 1st ed.; University of California Press: Berkeley, CA, USA, 1969; p. 246. [Google Scholar]
- Hawksworth, F.G.; Wiens, D. Biology and Classification of Dwarf Mistletoes (Arceuthobium); Agriculture Handbook Book Number 401; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1972; p. 234. Available online: http://naldc.nal.usda.gov/naldc/download.xhtml?id=CAT87208731 (accessed on 23 February 2016).
- Geils, B.W.; Mathiasen, R.L. Intensification of dwarf mistletoe on Southwestern Douglas-fir. For. Sci. 1990, 36, 955–969. [Google Scholar]
- Sala, A.; Carey, E.V.; Callawy, R.M. Dwarf mistletoe affects whole-tree water relations of Douglas-fir and western larch primarily through changes in leaf to sapwood ratios. Oecologia 2001, 126, 42–52. [Google Scholar] [CrossRef]
- Hadfield, J.S.; Mathiasen, R.L.; Hawksworth, F.G. Douglas-Fir Dwarf Mistletoe; Forest Insect and Disease Leaflet 54; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2000; p. 9.
- Castello, J.D.; Leopold, D.J.; Smallidge, P.J. Pathogens, patterns, and processes in forest ecosystems. BioScience 1995, 45, 16–24. [Google Scholar] [CrossRef]
- Ayres, M.P.; Lombardero, M.J. Assessing the consequences of global change for forest disturbance from herbivores and pathogens. Sci. Total Environ. 2000, 262, 263–286. [Google Scholar] [CrossRef]
- Williams, R.E.; Shaw, C.G., III; Wargo, P.M.; Sites, W.H. Armillaria Root Disease; Forest Insect and Disease Leaflet 78; U.S. Department of Agriculture, Forest Service, Northern Area State and Private Forestry, 1986. Available online: http://www.na.fs.fed.us/spfo/pubs/fidls/armillaria/armillaria.htm?) (accessed on 23 February 2016)Forest Insect and Disease Leaflet 78.
- Wargo, P.M.; Shaw, C.G., III. Armillaria root rot: The puzzle is being solved. Plant Dis. 1985, 69, 826–832. [Google Scholar] [CrossRef]
- McDonald, G.I.; Martin, N.E.; Harvey, A.E. Armillaria in the Northern Rockies: Pathogenicity and Host Susceptibility on Pristine and Disturbed Sites; Research Note INT-371; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1987. Available online: https://archive.org/details/armillariainnort371mcdo (accessed on 23 February 2016).
- Shaw, C.G., III; Kile, G.A. (Eds.) Armillaria Root Disease; Agricultural Handbook No. 691; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1991.
- Wickman, B.E. Tree Mortality and Top-Kill Related to Defoliation by the Douglass-Fir Tussock Moth in the Blue Ridge Mountains Outbreak; Forest Research Paper PNW-223; U.S. Department of Agriculture, Forest Service Pacific Northwest Research Station: Portland, OR, USA, 1978; p. 39.
- Johnson, D.W. Forest Pest Training Manual; U.S. Department of Agriculture, Forest Service, Forest Pest Management, Rocky Mountain Region: Lakewood, CO, USA, 1985.
- Haggle, S.K. Managmement Guide for Armaliaria Root Disease; U.S. Department of Agriculture, Forest Service, Forest Health Protection and State Forestry Organizations, 2008. Available online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187208.pdf (accessed on 24 February 2016).
- Guillaumin, J.J.; Legrand, P. Armillaria root rots. In Infectious Forest Diseases, 1st ed.; Gonthier, P., Nicolotti, G., Eds.; CABI: Boston, MA, USA, 2013; pp. 159–178. [Google Scholar]
- Worral, J.J.; Sullivan, K.F.; Harrington, T.C.; Steimel, J.P. Incidence, host relations and population structure of Armillaria ostoyae in Colorado campgrounds. For. Ecol. Manag. 2004, 192, 191–206. [Google Scholar] [CrossRef]
- Amman, G.D. Bark beetle-fire associations in the Greater Yellowstone area. In Proceedings of the Fire and the Environment Symposium, Knoxville, TN, USA, 20–24 March 1991; Nordvin, S.C., Waldrop, T.A., Eds.; pp. 313–320.
- Amman, G.D.; Ryan, K.C. Insect Infestation of Fire-Injured Trees in the Greater Yellowstone Area; Research Note INT-398; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1991; p. 9.
- Ryan, K.C.; Amman, G.D. Interactions between fire-injured trees and insects in the greater Yellowstone area. In Plants and Their Environment, Proceedings of the First Biennial Scientific Conference on the Greater Yellowstone Ecosystem, Yellowstone National Park, WY, USA, 16–17 September 1991; Despain, D.G., Ed.; National Park Service: Denver, CO, USA, 1995; pp. 259–271. [Google Scholar]
- Rasmussen, L.; Amman, G.; Vandygriff, J.; Oakes, R.; Munson, S.; Gibson, K. Bark Beetle and Wood Borer Infestation in the Greater Yellowstone Area during Four Postfire Years; Research Paper INT-RP-487; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1996; p. 9.
- Dickinson, M.B.; Johnson, E.A. Fire effets on trees. In Forest Fires Behavior and Ecological Effects, 1st ed.; Johnson, E.A., Miyanishi, K., Eds.; Academic Press: New York, NY, USA, 2001; pp. 477–521. [Google Scholar]
- Stevens-Rumman, C.P.; Morgan, C.P.; Hoffman, C. Bark beetles and wildfires: How does forest recovery change with repeated disturbances in mixed conifer forests? Ecosphere 2015, 6, 1–17. [Google Scholar] [CrossRef]
- Furniss, M.M.; Livingston, R.L.; McGregor, M.D. Development of a stand susceptibility classification for Douglas-fir beetle. In Hazard-Rating Systems in Forest Insect Pest Management: Symposium Proceedings, Athens, GA, USA, 31 July–1 August 1980; Hedden, R.L., Barras, S., Coster, J.E., Eds.; U.S. Department of Agriculture, Forest Service: Washington, D.C., USA, 1981; pp. 115–128. [Google Scholar]
- Christiansen, E.; Waring, R.H.; Berryman, A.A. Resistance of conifers to bark beetle attack: Searching for general relationships. For. Ecol. Manag. 1987, 22, 89–106. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negrón, J.F.; Costello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caissie, R. Stand characteristics and downed woody debris accumulations associated with a mountain pine beetle (Dendroctonus ponderosae Hopkins) outbreak in Colorado. For. Ecol. Mang. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Collins, B.J.; Rhoades, C.C.; Battaglia, M.A.; Hubbard, R.M. The effects of bark beetle outbreaks on forest development, fuel loads and potential fire behavior in salvage logged and untreated lodgepole pine forests. For. Ecol. Manag. 2012, 284, 260–268. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Wildfire and spruce beetle outbreak: Simulation of interacting disturbances in the central Rocky Mountains. Ecoscience 2009, 16, 28–38. [Google Scholar] [CrossRef]
- Parker, T.J.; Clancy, K.M.; Mathiasen, R.L. Interactions among fire, insects and pathogens in coniferous forests of the interior western United States and Canada. Agric. For. Entomol. 2006, 8, 167–189. [Google Scholar] [CrossRef]
- Giunta, A. Douglas-Fir beetle mediated changes to fuel complexes, foliar moisture content, and terpenes in interior Douglas-fir forests of the central Rocky Mountains. Master’s Thesis, Utah State University, Logan, UT, USA, 2016. [Google Scholar]
- McMillin, J.D.; Allen., K.K. Impacts of Douglas-Fir Beetles on Overstory and Understory Conditions of Douglas-Fir Stands; General Technical Report R2-64; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Golden, CO, USA, 2000; p. 17.
- Von Arx, J.A. Ueber die Ascomycetengattungen Ceratostomella Sacc., Ophiostoma Syd. Und Rostrella Zimmerman. Antonie Leeuwenhoek 1952, 18, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Gruell, G.E. Fire and Vegetative Trends in Northern Rockies: Interpretations from 1871–1982 Photographs; General Technical Report INT-130; U.S. Department of Agriculture, Forest Service Intermountain Research Station: Missoula, MT, USA, 1983; p. 117.
- Gavin, D.; Flower, A.; Cohn, G.; Heyerdahl, E.; Parsons, R. Interactions of Insects, Fire and Climate on Fuel Loads and Fire Behavior in Mixed Conifer Forest; Final Report for JFSP Project 09-1-06-5; 2013. Available online: http://www.firescience.gov/projects/09-1-06-5/project/09-1-06-5_final_report.pdf (accessed on 27 May 2015).
- Black, S.H.; Kulakowski, D.; Noon, B.R.; DellaSalla, D.A. Do bark beetle outbreaks increase wildfire risks in the central U.S. Rocky Mountains? Implications from recent research. Nat. Areas J. 2013, 33, 59–65. [Google Scholar] [CrossRef]
- Flower, A.; Gavin, D.G.; Heyerdahl, E.K.; Parson, R.A.; Cohn, G.M. Western spruce budworm outbreaks did not increase fire risk over the last three centuries: A dendrochronological analysis of inter-disturbance synergism. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Hummel, S.; Agee, J.K. Western spruce budworm defoliation effects on forest structure and potential fire behavior. Northwest Sci. 2003, 77, 159–169. [Google Scholar]
- Burgan, R.E.; Rothermel, R.C. BEHAVE: Fire Behavior Prediction and Fuel Modeling System FUEL Subsystem; General Technical Report INT-167; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1984; p. 126.
- Koonce, A.L.; Roth, L.F. The effects of dwarf mistletoe on fuel in precommercial ponderosa pine stands. In Proceedings of the Eighth Conference on Fire and Forest Meteorology: Weather—The Drive Train Connecting the Solar Engine to Forest Ecosystems, Detroit, MI, USA, 29 April–2 May 1985; Donoghue, L.R., Martin, R.E., Eds.; Society of American Foresters: Bethesda, MD, USA, 1985; pp. 66–72. [Google Scholar]
- Dickman, A.; Cook, S. Fire and fungus in a mountain hemlock forest. Can. J. Bot. 1989, 67, 2005–2016. [Google Scholar] [CrossRef]
- Jain, T.B.; Battaglia, M.A.; Han, H.S.; Graham, R.T.; Keyes, C.R.; Fried, J.S.; Sandquist, J.E. A Comprehensive Guide to Fuel Management Practices for Dry Mixed Conifer Forests in the Northwestern United States; General Technical Report RMRS-GTR-292; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Golden, CO, USA, 2012; p. 331.
- Hagle, S.K.; Gibson, K.E.; Tunnock, S. Field Guide to Diseases and Insect Pests of Northern and Central Rocky Mountain Conifers; General Technical Report R1-03-08; U.S. Department of Agriculture, Forest Service, State and Private Forestry, Northern and Intermountain Regions: Missoula, MT, USA; Ogden, UT, USA, 2003; p. 197.
- Man, G. Major Forest and Insect Disease Conditions in the United States: 2011 Update; Forest Service Report FS-1000; U.S. Department of Agriculture: Washington, DC, USA, 2012; pp. 14–16.
- Jenkins, M.L. Major Forest and Insect Disease Conditions in the United States: 2013; Forest Service Report FS-1054; U.S. Department of Agriculture, Forest Service, Forest Health Protection: Washington, DC, USA, 2015; pp. 18–20.
- Lessard, E.D.; Schmid, J.M. Emergence, attack densities, and host relationships for the Douglas-fir beetle, (Dendroctonus pseudotsugae Hopkins) in northern Colorado. Gt. Basin Nat. 1990, 50, 333–338. [Google Scholar]
- Schmid, J.M.; Mata, S.A. Natural Variability of Specific Forest Insect Populations and Their Associated Effects in Colorado; General Technical Report RM-GTR-275; U.S Department of Agriculture, Forest Service: Washington, DC, USA, 1996.
- Berryman, A.A.; Wright, L.C. Defoliation, tree condition, and bark beetles. In The Douglas-Fir Tussock Moth a Synthesis; Stark, M.H., Campell, R.W., Eds.; Technical Bulletin 1585; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1978; pp. 81–87. [Google Scholar]
- Singh, P. Research and management strategies for major tree diseases in Canada: Synthesis part 1. For. Chron. 1993, 69, 151–162. [Google Scholar] [CrossRef]
- Huber, D.P.W.; Borden, J.H. Angiosperm bark volatiles disrupt response of Douglas-fir beetle, Dendroctonus pseudotsugae, to attractant-baited traps. J. Chem. Ecol. 2001, 27, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Kay, C.E. Aboriginal overkill and native burning: Implications for modem ecosystem management. West. J. Appl. For. 1995, 10, 121–126. [Google Scholar]
- Keane, R.E.; Ryan, K.C.; Veblen, T.T.; Allen, C.D.; Logan, J.A.; Hawkes, B. The cascading effects of fire exclusion in Rocky Mountain ecosystems. In Rocky Mountain Futures: An ecological Perspective, 1st ed.; Baron, J.S., Ed.; Island Press: Washington, DC, USA, 2002; pp. 133–152. [Google Scholar]
- Baker, W.L. Indians and fire in the Rocky Mountains: The wilderness hypothesis reviewed. In Fire Native Peoples, and the Natural Landscape, 1st ed.; Vale, T.R., Ed.; Island Press: Washington, DC, USA, 2002; pp. 41–77. [Google Scholar]
- Peterson, C.S.; Speth, L.E. A History of the Wasatch-Cache National Forest. Utah State University, 1980. Available online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5053310.pdf (accessed on 1 March 2016). [Google Scholar]
- Alexander, T.G.; Fish, R.J. The Forest Service in Utah. Available online: http://www.uen.org/utah_history_encyclopedia/f/FOREST_SERVICE.html (accessed on 1 March 2016).
- Owston, P.W.; Stein, W.I. Pseudotsuga Carr., Douglas-fir. In Seeds of Woody Plants in the United States; Agriculture Handbook 450; Schopmeyer, C.S., Ed.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1974; pp. 674–683. [Google Scholar]
- Noss, R.F.; Franklin, J.F.; Baker, W.L.; Schoennagel, T.; Moyle, P.B. Managing fire-prone forests in the western United States. Front. Ecol. Environ. 2006, 4, 481–487. [Google Scholar] [CrossRef]
- Landres, P.; Morgan, P.; Swanson, F. Overview of the use of natural variability concepts in managing ecological systems. Ecol. Appl. 1999, 9, 1179–1188. [Google Scholar]
- Graham, R.T.; Harvey, A.E.; Jain, T.B.; Tonn, J.R. Effects of Thinning and Similar Stand Treatments on Fire Behavior in Western Forests; General Technical Report PNW-GTR-463; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1999.
- Rippy, R.C.; Stewart, J.E.; Zambino, P.J.; Klopfenstein, N.B.; Tirocke, J.M.; Kim, M.S.; Thies, W.G. Root Diseases in Coniferous Forests of the Inland West: Potential Implications of Fuels Treatments; General Technical Report, RMRS-GTR-141; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005.
- Edmonds, R.L.; Agee, J.K.; Gara, R.I. Forest Health and Protection, 1st ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2000; pp. 359–397. [Google Scholar]
- Fernandes, P.M.; Hermínio, S.B. A review of prescribed burning effectiveness in fire hazard reduction. Int. J. Wildland Fire 2003, 12, 117–128. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Mitchell, R.G.; Filip, G.M. Historical and Current Roles of Insects and Pathogens in Eastern Oregon and Washington Forested Landscapes; General Technical Report, PNW-GTR-327; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1994.
- Arno, S.F. Forest fire history in the northern Rockies. J. For. 1980, 78, 460–465. [Google Scholar]
- McIver, J.D.; Starr, L. A literature review on the environmental effects of postfire logging. West. J. Appl. For. 2001, 16, 159–168. [Google Scholar]
- Potts, D.F.; Peterson, D.L.; Zuuring, H.R. Watershed Modeling for Fire Management Planning in the Northern Rocky Mountains; Research Paper PSW-177; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkely, CA, USA, 1985.
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Post-wildfire logging hinders regeneration and increases fire risk. Science 2006. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2016, 53, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.T.; McCaffrey, S.; Jain, T.B. Science Basis for Changing Forest Structure to Modify Wildfire Behavior and Severity; General Technical Report RMRS-120; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 2004.
- Stephens, S.L.; Moghaddas, J.J. Experimental fuel treatment impacts on forest structure, potential fire behavior, and predicted tree mortality in a California mixed conifer forest. For. Ecol. Manag. 2005, 215, 21–36. [Google Scholar] [CrossRef]
- Allen, C.D.; Betancourt, J.L.; Swetnam, T.W. Landscape changes in the southwestern United States: Techniques, long-term data sets, and trends. In Perspectives on the Land-Use History of North America: A Context for Understanding Our Changing Environment; Sisk, T.D., Ed.; U.S. Geological Survey: Reston, VA, USA; pp. 71–84.
- Caprio, A.C.; Graber, D.M. Returning fire to the mountains: Can we successfully restore the ecological role of pre-Euroamerican fire regimes to the Sierra Nevada. In Proceedings of the Wilderness Science in a Time of Change Conference, Missoula, MT, USA, 23–27 May 1999; Cole, D.N., McCool, S.F., Borrie, W.T., O’Laughlin, J., Eds.; U.S. Department of Agriculture, Forest Service: Proceedings RMRS-P-15-VOL-5. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000; pp. 223–241. [Google Scholar]
- Heyerdahl, E.K.; Lertzman, K.; Wong, C.M. Mixed-severity fire regimes in dry forests of southern interior British Columbia, Canada. Can. J. For. Res. 2012, 42, 88–98. [Google Scholar] [CrossRef]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giunta, A.D.; Jenkins, M.J.; Hebertson, E.G.; Munson, A.S. Disturbance Agents and Their Associated Effects on the Health of Interior Douglas-Fir Forests in the Central Rocky Mountains. Forests 2016, 7, 80. https://doi.org/10.3390/f7040080
Giunta AD, Jenkins MJ, Hebertson EG, Munson AS. Disturbance Agents and Their Associated Effects on the Health of Interior Douglas-Fir Forests in the Central Rocky Mountains. Forests. 2016; 7(4):80. https://doi.org/10.3390/f7040080
Chicago/Turabian StyleGiunta, Andrew D., Michael J. Jenkins, Elizabeth G. Hebertson, and Allen S. Munson. 2016. "Disturbance Agents and Their Associated Effects on the Health of Interior Douglas-Fir Forests in the Central Rocky Mountains" Forests 7, no. 4: 80. https://doi.org/10.3390/f7040080
APA StyleGiunta, A. D., Jenkins, M. J., Hebertson, E. G., & Munson, A. S. (2016). Disturbance Agents and Their Associated Effects on the Health of Interior Douglas-Fir Forests in the Central Rocky Mountains. Forests, 7(4), 80. https://doi.org/10.3390/f7040080