Abstract
An abrupt onset stimulus was presented while the participants' eyes were in motion. Because of saccadic suppression, participants did not perceive the visual transient that normally accompanies the sudden appearance of a stimulus. In contrast to the typical finding that the presentation of an abrupt onset captures attention and interferes with the parti cipants' responses, we found that an intra-saccadic abrupt onset does not capture attention: It has no effect beyond that of increasing the set-size of the search array by one item. This finding favours the local transient account of attentional capture over the novel object hy pothesis.
Introduction
Contrary to subjective experience, we are driven to a considerable extent by our environment. A prime example of this is attentional capture. When an object suddenly appears, we often cannot help but look at it (Theeuwes, Kramer, Hahn, & Irwin, 1998). And even if we do manage to avoid directing our gaze towards a novel object, it invariably attracts our attention (Posner, 1980). This involuntary shift of attention can be measured easily in an experimental setting: If a suddenly appearing stimulus (an 'abrupt onset') is not relevant to the task at hand, its appearance will disrupt performance (Theeuwes, 1994). Conversely, if an abrupt onset happens to be task-relevant, even if just by chance, performance will improve (Yantis & Jonides, 1984).
In the present study, we investigate the effect of an abrupt onset that is presented during a saccadic eye movement. Visual perception is greatly impaired during saccadic eye movements, a phenomenon that is generally referred to as saccadic suppression (Matin, 1974). The origin of saccadic suppression is a topic of debate, centred around two main hypotheses (for a recent review, see Castet, 2010). The traditional, and widely accepted, view holds that contrast sensitivity is reduced during eye movements, particularly for low spatial frequencies (Burr & Ross, 1982; Volkmann, Riggs, White, & Moore, 1978). However, an alternative view holds that saccadic suppression is, at least in part, due to temporal masking (Campbell & Wurtz, 1978; Castet, 2010). According to the masking account, we are not aware of the intra-saccadic percept, because it is masked by the post-saccadic image. However, although the origin of saccadic suppression is topic of debate, its perceptual effect is clear: Visual events that occur while the eyes are in motion are not perceived (except under rare circumstances, cf. Castet & Masson, 2000). For the purpose of the present experiment, saccadic suppression is therefore a useful tool, because it allows us to present an abrupt onset, while preventing participants from perceiving the exact moment of its appearance (the ‘visual transient’).
Whether or not an intra-saccadic abrupt onset captures attention speaks to the mechanism that underlies attentional capture. Some authors have suggested that novel objects capture attention by virtue of being novel perceptual entities (analogous to object-files, cf. Kahneman, Treisman, & Gibbs, 1992), regardless of whether their appearance is accompanied by a visual transient (the novel object hypothesis). Support for this view comes mostly from experiments in which stimuli were equiluminant with the display background (Yantis & Hillstrom, 1994; Yantis & Jonides, 1996; also see Davoli, Suszko, & Abrams, 2007). These experiments have shown that a luminance increment is not required for objects to capture attention, in apparent support of the novel object hypothesis. With regard to the present study, the novel object hypothesis predicts that intra-saccadic abrupt onsets capture attention, because they are novel perceptual objects even though the visual transient of their appearance is not perceived.
Others have argued that attentional capture is solely driven by visual transients (the local transient hypothesis; Franconeri, Hollingworth, & Simons, 2005; Hollingworth, Simons, & Franconeri, 2010; Jonides & Yantis, 1988). Evidence that favours the local transient hypothesis over the novel object hypothesis comes predominantly from paradigms in which a novel object appears without being accompanied by a unique visual transient. For example, in a cleverly designed experiment, Franconeri and colleagues (2005) used a contracting annulus that briefly occluded a search array by moving over it. When the search array re-emerged from underneath the annulus, it included a novel object. Their crucial finding was that the novel object did not capture attention when its appearance was hidden from view by the annulus (but see Chua, 2009).
There appears to be an implicit, but important assumption that underlies much of the debate between the novel object and local transient accounts of attentional capture: the assumption that a luminance increment is the only transient that matters (e.g., Jonides & Yantis, 1988; Yantis & Hillstrom, 1994; Yantis & Jonides, 1996). A different, more contemporary view, which underlies most models of visual attention (Itti & Koch, 2001; Itti, Koch, & Niebur, 1998), is that distinct features (orientation, colour, etc.) are processed in parallel (Koch & Ullman, 1985). In this view, a change from one orientation to another, or from one colour to another, constitutes a visual transient, just like a luminance increment. Therefore, in order to adequately test the novel object account of attentional capture, one would need to present a novel object without a visual transient of any kind, not just without a luminance increment. In our view, the only way to achieve this is by presenting a novel object during an eye movement, while visual perception is greatly impaired (Matin, 1974).
At present, evidence favours the view that intra-saccadically presented stimuli do not capture attention. This, in turn, supports the local transient account of attentional capture. Crucial in this regard are the classic studies on changes blindness (Rensink, O’Regan, & Clark, 1997; Grimes, 1996; for similar findings, see Mack & Rock, 1998; Bridgeman, Hendry, & Stark, 1975). In one variation of the change blindness paradigm, two images are presented in alternation (Brockmole & Henderson, 2005a, 2005b; Grimes, 1996). Both images are identical, except for the presence of a particular object. For example, a tree may be present in one image, but not the other. In the experimental condition, the moment at which the additional object appears is time-locked to the onset of a saccadic eye movement. In the control condition, the object appears during fixation. The typical finding is that people are very poor at detecting the novel object when it appears during an eye movement, whereas detection is exceedingly efficient when the object appears during fixation. The explanation is that normally a suddenly appearing object constitutes a visual transient that captures our attention. But when the change occurs during saccadic suppression, the visual transient is not perceived, and the change no longer captures our attention.
However, it is important to clearly define the limits of what we can conclude from these findings. Change blindness experiments elegantly show that intra-saccadic abrupt onsets have, at most, a small effect when studied in the context of natural scenes. But they do not show that intra-saccadic abrupt onsets have no effect at all, even when using a paradigm that has been specifically designed to elicit a robust effect of attentional capture.
More specifically, the scenes that were used in previous studies contained many objects, which were not organised in any obvious pattern (Brockmole & Henderson, 2005a, 2005b). This made it difficult for participants to infer whether a new object had appeared. In contrast, in the current experiment we initially presented only three objects, with the novel intra-saccadically presented object being the fourth. Furthermore, the display was highly structured. The small set size and high degree of structure made it possible for participants to retain the entire search display in trans-saccadic/ visual working memory, which has an estimated capacity of about four objects (Irwin, 1992; Luck & Vogel, 1997; Prime, Tsotsos, Keith, & Crawford, 2007). Consequently, compared to previous studies, it was very easy for participants to infer the appearance of the abrupt onset. The crucial question is whether attentional capture is reinstated under these circumstances.
Another possibility is that intra-saccadic abrupt onsets capture attention only when they occur in a retinotopic frame of reference. This may seem like a quixotic hypothesis, but recent studies have shown that, immediately after an eye movement, attentional effects are, in some cases, predominantly retinotopic (Golomb, Chun, & Mazer, 2008; Golomb, Marino, Chun, & Mazer, 2011; Mathôt & Theeuwes, 2010a, 2010b; but see Pertzov, Zohary, & Avidan, 2010). Presumably, this is because every eye movement is followed by a brief 'window of instability' during which the visual system has not yet fully updated its retinotopic representation (Morris, Kubischik, Hoffmann, Krekelberg, & Bremmer, 2012; for a review see Mathôt & Theeuwes, 2011a).
Therefore, with the present study we aim to answer the following questions: First, do intra-saccadic abrupt onsets completely fail to capture attention, even in a paradigm that has been designed to elicit a robust effect of attentional capture? And second, in which reference frame do intra-saccadic abrupt onsets capture attention, if they capture attention at all?
Experiment 1
The aim of Experiment 1 was to investigate whether a suddenly appearing stimulus captures attention when the onset of the stimulus occurs during an eye movement (an intra-saccadic abrupt onset). We used a variation of the attentional capture paradigm, introduced by Theeuwes (1994; for a recent review, see Theeuwes, 2010).
In a typical attentional capture paradigm, participants report the orientation of a line-segment in a unique placeholder (e.g., a single red circle among multiple green circles). Initially, all potential target line-segments are masked. On a proportion of trials, an additional stimulus (an abrupt onset) is presented simultaneously with the unmasking of the target (i.e., the moment when the target becomes visible). Empirically, the typical result is that participants are slower to respond when an abrupt onset is presented. Theoretically, the interpretation is that the abrupt onset captures the participants' attention, diverting attention away from the target stimulus, thus delaying their response.
In our variation of this task, the unmasking of the search array and the presentation of the onset were timelocked to the onset of a saccadic eye movement. Furthermore, there were three conditions, in order to determine the reference frame of the attentional capture effect, if any.
In the spatiotopic condition, the masked and unmasked search arrays were presented at the same location on the display. This allowed us to investigate whether an intra-saccadic abrupt onset captures attention when it occurs in spatiotopic, or world centred, coordinates.
In the retinotopic condition, the masked and unmasked search arrays were presented in the same retinal coordinates: The search array moved with the eyes. This allowed us to investigate the effect of an abrupt onset in retinotopic, or eye centred, coordinates.
Finally, in the both condition, the abrupt onset was both a retinotopic and a spatiotopic visual event. This allowed us to investigate whether the effects of intra-saccadic abrupt onsets (if any) in the retinopic and spatiotopic conditions would be additive, or interact in some way.
To avoid the possibility of overloading trans-saccadic/ visual working memory, which has an estimated capacity of about 4 items (Irwin, 1992; Luck & Vogel, 1997; Prime et al., 2007), we used a set-size of 3 (not including the abrupt onset, cf. Theeuwes, 1994).
The experimental script and participant data are available from the following location:
Methods
8 observers, including one of the authors (SM), participated in the experiment. All participants were between the ages of 21 and 39, and reported normal or corrected visual acuity. Eye movements were recorded using an Eyelink 1000 (SR Research, Missisauga, Canada, ON), a video based eye tracker sampling at 1000Hz. The experiment was created using OpenSesame (Mathôt, Schreij, & Theeuwes, in press). Stimuli were presented on a 22” CRT monitor, with a resolution of 1024x768px and a refresh rate of 100Hz.
A schematic example trial is shown in Figure 1a. Before the start of each trial, a white fixation dot (r=0.25°) was presented against a dark background. This fixation dot was presented 9.4° to the left or the right of the display centre. After 750ms, which gave participants sufficient time to (re)fixate, a drift correction procedure was executed. Drift correction was triggered when a sustained fixation was detected and did not require a manual response from the participant.
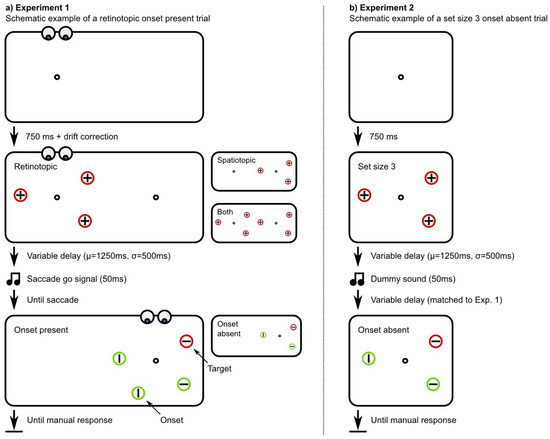
Figure 1.
a) A schematic example trial of Experiment 1 in the retinotopic, onset present condition. The target is defined as the linesegment in the uniquely coloured circle. The abrupt onset appears at a previously unoccupied location, either in retinotopic coordinates (eye centred; as shown here), spatiototopic coordinates (world centred), or both. b) A schematic example trial of Experiment 2 in the set size 3, onset absent condition.
Next, the trial proper started with the presentation of a masked search array and a saccade target that was identical to the fixation dot. The saccade target was always presented at the mirror location of the fixation dot. The masked search array consisted of three unfilled circular placeholders (r=1.06°). All placeholders were of the same colour, which could be either red or green. All placeholders contained a cross, consisting of a white vertical and horizontal line-segment (1.25°). All placeholders were presented at randomly selected locations, 5.3° from the fixation dot and/ or saccade target (see below), and spaced apart by multiples of 60° angular.
In the spatiotopic condition, the masked search array was presented around the saccade target. In the retinotopic condition, the masked search array was presented around the fixation dot. In the both condition, two identical masked search arrays were presented simultaneously around both the fixation dot and the saccade target (i.e. six placeholders were presented in total).
The participants were instructed to keep fixating on the fixation dot until after a random interval (µ=1250ms, σ=500ms; minimum=500ms) an auditory go-signal was presented (50ms, 440Hz, sine wave). Participants were instructed to make a saccade to the saccade target when they heard the go-signal, but to avoid anticipatory responses1.
As soon as a saccade was detected, defined as the moment at which the horizontal gaze position deviated more than 1.9° from the fixation dot (µ=906ms, σ=219ms; unfiltered across all participants), the (unmasked) search array was presented. All placeholders, except the one that contained the target line-segment, changed colour (i.e., from red to green or vice versa). In all placeholders one of the line-segments disappeared, so that a single horizontal or vertical line-segment remained. There was always a single, unmasked search array, which was presented around the saccade target. The retinotopic, spatiotopic, and both conditions differed only in the location and number of the pre-saccadic, masked search array(s).
In the onset present condition, an additional distractor, identical to the other distractors, appeared at a randomly selected location that was previously unoccupied. In the onset absent condition, no additional distractor was presented.
Participants reported the orientation of the target linesegment (i.e., the line-segment in the uniquely coloured placeholder) as quickly as possible. If the target line-segment was horizontal they pressed the left button on a serial response box (Psychology Software Tools, Sharpsburg, PA, USA). The right button was pressed on a vertical target line-segment.
Onset presence (absent, present), condition (spatiotopic, retinotopic, both), and initial fixation condition (left, right) were mixed within blocks. Target colour (red, green) was fully randomized. The experiment consisted of 36 practice trials, followed by 288 experimental trials.
Results
Trials in which gaze deviated more than 3° from the expected point of gaze (10.3%), trials in which saccade latency was below 50ms (-) or above 1000ms (19.4%)1, and trials in which response time was below 100ms (0.1%) or above 2000ms (0.4%) were discarded.
The mean error rate across valid trials was 10%. Across valid and correct trials, the mean response time was 743ms, and the mean saccade latency was 840ms.
An alpha level of .05 was used for all statistical tests. F-values are reported when larger than 1.
A repeated measures analysis of variance (ANOVA) was conducted with onset presence (absent, present) and condition (spatiotopic, retinotopic, both) as within-subject factors and mean correct response time as dependent variable (Figure 2a). This revealed an effect of onset presence, F(1,7) = 7.1, p < .05, such that response times were faster when an onset was presented than when no onset was presented.
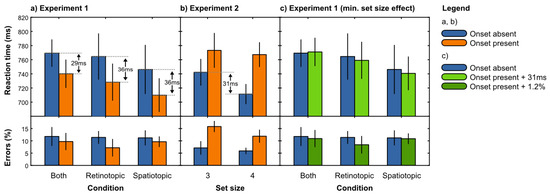
Figure 2.
a) Results of Experiment 1. Participants were faster to respond if an intra-saccadic abrupt onset was presented. This 'paradoxical facilitation' was found in all three conditions (retinotopic, spatiotopic, and both). b) Results of Experiment 2.Participants were slower to respond if an abrupt onset was presented during fixation, reflecting the typical pattern of attentional capture. Importantly, if no abrupt onset was presented, participants were faster in the set size 4 condition than in the set size 3 condition. c) The set size effect of Experiment 2 fully accounted for the paradoxical facilitation of Experiment 1.
A similar repeated measures ANOVA using error rate as dependent variable revealed no significant effects. Tentatively, there was a trend towards a reduced error rate in the presence of a distractor, F(1,7) = 3.2, analogous to the effect that was found in the response times.
Discussion
The results of Experiment 1 were unanticipated: The presentation of an abrupt onset caused facilitation, as measured by decreased response times and, tentatively, decreased error rates. This effect, which was present across all three conditions, contrasts with the typical finding that an abrupt onset causes interference. A possible explanation for this paradoxical facilitation was investigated in Experiment 2.
Experiment 2
The aim of Experiment 2 was twofold. Firstly, we wanted to verify our methodology by replicating the typical finding that an abrupt onset causes interference. To do this, we created an experiment that was similar to Experiment 1, but did not involve an eye movement.
Secondly, we wanted to investigate the possibility that the paradoxical facilitation observed in Experiment 1 could be explained by a set-size effect. If the visual system disregards all (or most) changes that occur during a saccade, we might expect the visual system to process the post-saccadic search array simply as it is found 'on arrival'. If this is the case, in Experiment 1 presenting an intra-saccadic abrupt onset was equivalent to increasing the set-size from 3 to 4.
At first glance you might expect that an effect of setsize, if any, should be such that an increased set-size leads to increased response times (cf. the classic study by Treisman & Gelade, 1980). But previous research has shown that this is not always the case. Under particular circumstances, increasing the number of non-targets will cause the target element to become more salient, resulting in faster response times (Green, 1991, 1992; Sagi & Julesz, 1987). This is particularly the case when the target is defined as the unique stimulus, which is the case in the present experiments. Simply put, a red circle is more unique, and therefore more conspicuous, among three green circles than among two green circles.
To test whether a set-size effect could account for the paradoxical facilitation found in Experiment 1, we included set-sizes of 3 and 4 (not including the abrupt onset) in Experiment 2.
The experimental script and participant data are available from the following location:
Method
The method was identical to that of Experiment 1, with the following exceptions.
8 observers, including one of the authors (SM), participated in the experiment. All participants were between the ages of 18 and 45, and reported normal or corrected visual acuity. Eye movements were not monitored. Stimuli were presented on a 19” CRT monitor, with a resolution of 1024x768px and a refresh rate of 120Hz.
A schematic example trial is shown in Figure 1b. The fixation dot was presented at the centre of the display, and both the masked and unmasked search array were presented centrally, around the fixation dot. In order to match the paradigm as closely as possible to that of Experiment 1, a dummy sound, identical to the saccade go-signal of Experiment 1, was presented. In addition, the saccade latencies of Experiment 1 were 'played back': On each trial, a saccade latency was randomly selected from the valid trials of Experiment 1 and used for the delay interval after the dummy sound.
A set-size of 4 was included in addition to the original set-size of 3 (set-sizes do not include the abrupt onset).
Participants reported a horizontal target line-segment by pressing the 'z'-key on a standard keyboard, and a vertical line-segment by pressing the slash-key.
Onset presence (absent, present) and set-size (3, 4) were mixed within blocks. The experiment consisted of 12 practice trials, followed by 192 experimental trials.
Results
Trials in which response time was below 100ms (-) or above 2000ms (0.6%) were discarded.
The average error rate across valid trials was 10%. Across valid and correct trials, the mean response time was 747ms.
A repeated measures ANOVA was conducted with onset presence (absent, present) and set-size (3, 4) as within-subject factors and mean correct response time as dependent variable (Figure 2b). This revealed an effect of onset presence, F(1,7) = 6.7, p < .05, reflecting the typical distractor interference effect. Tentatively, there were trends towards an effect of set-size, F(1,7) = 3.2, and a set-size by onset presence interaction, F(1,7) = 3.1.
A similar repeated measures ANOVA using error rate as dependent variable revealed an effect of onset presence, F(1,7) = 16.4, p < .01, again reflecting a distractor interference effect. Tentatively, there was a trend towards an effect of set-size, F(1,7) = 4.4.
To get a better estimate of the pure set-size effect in the absence of an abrupt onset, we performed a two-tailed paired samples t-test between set-sizes 3 and 4 using only the distractor absent trials, with mean correct response time as dependent variable. This revealed that responses were slower for set-size 3 than for set-size 4, t(7) = 4.4, p < .005. A similar t-test using error rate as dependent variable showed no effect.
Discussion
In Experiment 2 we replicated the typical finding that an abrupt onset stimulus leads to increased response times and error rates, thus boosting our confidence in the overall methodology and results of Experiment 1.
Furthermore, the results show that participants respond faster in the set-size 4 condition than in the set-size 3 condition, at least when no onset stimulus is presented. This confirms the existence of a set-size effect (Green, 1991, 1992; Sagi & Julesz, 1987).
In the next section, we will examine whether the setsize effect found in Experiment 2 can account for the paradoxical facilitation observed in Experiment 1.
Cross-Experimental Analysis
Although it is clear that the paradoxical facilitation (Exp. 1) can be explained at least in part by a set-size effect (Exp. 2), it is not obvious that these two effects completely cancel each other out. To investigate this more rigorously, we conducted a cross-experimental analysis.
First, we derived the magnitude of the set-size effect from the onset absent trials of Experiment 2. We subtracted the mean correct response time in the set-size 3 condition from that in the set-size 4 condition, which gave us a estimated effect magnitude of 31ms. We did the same for the mean error rate (even though there was no significant set-size effect in the error data) and obtained an effect magnitude of 1.2%.
Next, we added the estimated effect magnitude to the response times and error rates of the onset-present trials of Experiment 1 (Figure 2c). Using the 'set-size corrected' data, we performed the same repeated measures ANOVA as before, with onset presence (absent, present) and condition (spatiotopic, retinotopic, both) as within-subject factors and mean correct response time as dependent variable. This revealed no effects, nor did a similar analysis using mean error rate as dependent variable. However, to avoid drawing conclusions from a null-result, we conducted a complementary Bayesian analysis.
We determined the size of the attentional capture effect in the set-size corrected data, by taking the difference in mean response time between onset present and onset absent trials, collapsed over the three conditions. This gave us the following estimate: M = -1.6ms, SE = 13.3ms. Next, we chose realistic lower and upper bounds for the effect. We set the lower bound to 0ms, since very small yet reliable capture effects have been reported (e.g., ~10ms reported by Mulckhuyse, Talsma, & Theeuwes, 2007). We set the upper bound to 100ms, since very large capture effects tend to be in this range (e.g., ~85ms for small separation trials in Exp. 2 of Mathôt & Theeuwes, 2011b). Using these parameters, and assuming a uniform prior distribution, we determined the Bayes factor (cf. Dienes, 2011), Bf = 0.15. Following Jeffreys (1961; reproduced in Wetzels et al., 2011), this constitutes substantial evidence for H0.
In summary, there is substantial evidence that the paradoxical facilitation (Exp. 1) is fully accounted for by a set-size effect (Exp. 2).
General Discussion
In the present study we pitted the novel object account of attentional capture (Chua, 2009; Davoli et al., 2007; Yantis & Hillstrom, 1994; Yantis & Jonides, 1996) against the local transient account (Franconeri et al., 2005; Hollingworth et al., 2010; Jonides & Yantis, 1988). We presented a stimulus during a saccadic eye movement. Because visual perception is greatly impaired while the eyes are in motion (Matin, 1974), this manipulation masked the visual transient that normally accompanies the appearance of a stimulus. Phrased differently, in our experiments participants did not perceive the exact moment at which the stimulus appeared.
The main finding is that an intra-saccadically presented stimulus does not capture attention—it has no effect beyond that of increasing the set size of the search display by one. By most definitions, a suddenly appearing stimulus is a novel perceptual entity, regardless of whether its appearance is masked by saccadic suppression. Therefore, this result strongly favours the local transient account, which postulates that a visual transient is required for attentional capture to occur, over the novel object account.
The present study complements previous research in a number of important ways. Franconeri and colleagues (2005) found that a novel stimulus does not capture attention when it emerges from underneath an occluder. Based on this finding they concluded that novel objects do not capture attention without a visual transient, in line with our own views. However, the presence of a moving occluder might have had unanticipated side-effects, for example because it is likely to capture attention itself (Franconeri & Simons, 2003). In contrast, our manipulation, which was one of timing, did not interfere with the search display in a similar way. Our results therefore offer important corroborative evidence for the crucial role of visual transients in attentional capture.
Furthermore, we used a sparse and structured display, which consisted of three to four items. This is a critical departure from previous studies on intra-saccadically presented stimuli, which have generally used unstructured and complex natural scenes (e.g., Brockmole & Henderson, 2005a, 2005b). It has been shown that the capacity of visual working memory is about 4 items (Luck & Vogel, 1997). Given the involuntary allocation of attention to the target of an upcoming eye movement (e.g., Deubel & Schneider, 1996; Godijn & Pratt, 2002), and the tight coupling between attention and working memory (Awh & Jonides, 2001), it is conceivable that the effective capacity of working memory is somewhat reduced in paradigms that require an eye movement, such as the one used here. Yet, previous studies have consistently shown that such impairment, if any, is negligible, and that transsaccadic memory (i.e. working memory across saccades) has a comparable capacity of 3 to 4 items (Irwin, 1992; Prime et al., 2007; for a recent review, see Prime, Vesia, & Crawford, 2011). Therefore, in our experiments the entire search array could be retained in memory. The finding that, even under these circumstances, an intra-saccadically presented novel object does not capture attention, strongly suggests that a visual transient is indeed required for attentional capture to occur: Previous failures to find attentional capture by intra-saccadically presented stimuli cannot be attributed to an overload of working memory.
In defence of the novel object account of attentional capture, one could argue that the perceptual effect of a saccade is not unlike that of a sudden visual disruption (Castet, 2010). In that sense, our strategy of presenting a sudden onset during a saccade is comparable to previous experiments, in which a sudden onset was presented during a visual disruption (Davoli et al., 2007; Franconeri et al., 2005; Hollingworth et al., 2010). Consequently, one could argue that the present experiment was not a fair test of the novel object hypothesis, because object representations might not be robust to the disruptions caused by saccadic eye movements.
However, the crucial distinction between the local transient and novel object hypothesis is the level at which attentional capture is assumed to operate. The local transient account assumes that attentional capture is triggered by a sudden change in some low level feature of the visual input, such as a luminance increment or colour change. In contrast, the novel object hypothesis postulates the existence of perceptual entities (analogous to object-files, cf. Kahneman et al., 1992), which are, in some sense, detached from low level visual input. The term 'perceptual entity' is not clearly defined, but it is presumed to reflect some “relatively high-order mechanism” (Yantis & Hillstrom, 1994, p. 106) that “bridges over the discontinuities produced by (...) saccades” (Kahneman et al., 1992, p. 178). Given these definitions, the results of the present study constitute clear evidence against the novel object hypothesis.
The present results also speak to the mechanisms that underlie visual stability—our ability to integrate visual information from one fixation to the next. In a classic essay, MacKay (1972) suggested that eye movements can be viewed as questions. Before every eye movement we 'ask' whether, after the eye movement, all stimuli will be where we expect them to be. The answer is provided by the image that is seen after the eye movement. If the answer does not match the question, a change is detected (this general idea, often referred to as 'state feedback control', is also increasingly applied outside of the domain of vision, see e.g. Hickok, 2012). Crucially, if a stimulus is presented only after the eyes have set in motion, as we have done here, it will not be part of the 'question'. Consequently, the change is not automatically detected, and we have to resort to higher level processes (Brockmole & Henderson, 2005a). Using more contemporary terminology, visual stability relies on a sparse representation that includes only the objects that were in the focus of attention prior to the eye movement (Irwin, 1991, 1996; Prime et al., 2007; for a review, see Mathôt & Theeuwes, 2011b). Because an intra-saccadic abrupt onset appears at a previously unattended location and does not capture attention by itself, its sudden appearance has little effect on our subjective sense of visual stability and on our behaviour. This is demonstrated particularly saliently by the finding that presenting an intra-saccadic abrupt onset is equivalent to increasing the set size by one item: Apparently, after the eye movement the image is evaluated more or less anew, without regard to whether the abrupt onset was present before the eye movement or not.
Finally, we have shown that the lack of attentional capture by intra-saccadically presented objects cannot be explained by the fact that just after an eye movement the visual system has not yet fully updated its retinotopic representation (Golomb et al., 2008; Mathôt & Theeuwes, 2010a, 2010b; Morris et al., 2012): Retinotopically matched abrupt onsets do not capture attention any more than spatiotopically matched abrupt onsets, or a combination of both.
In summary, with a paradigm designed to elicit a robust effect of attentional capture, the present study confirms that intra-saccadically presented stimuli do not capture attention (Brockmole & Henderson, 2005a, 2005b; Grimes, 1996). It is all about the transient.
Acknowledgements
This research was funded by a grant from NWO (Netherlands Organization for Scientific Research), grant 463-06-014 to Jan Theeuwes. We would like to thank Donatas Jonikaitis for pointing out the potential importance of a set-size effect.
Notes
| 1 | Our instruction emphasized saccadic accuracy over speed. This resulted in the exclusion of a substantial proportion of trials (19.4%) in which participants executed exceedingly slow eye movements (>1000ms). However, there is no reason to suspect that this has systematically influenced the pattern of results highlighted here. |
References
- Awh, E., and J. Jonides. 2001. Overlapping mechanisms of attention and spatial working memory. Trends in Cognitive Sciences 5, 3: 119–126. [Google Scholar] [CrossRef] [PubMed]
- Bridgeman, B., D. Hendry, and L. Stark. 1975. Failure to detect displacement of the visual world during saccadic eye movements. Vision Research 15, 6: 719–722. [Google Scholar] [CrossRef] [PubMed]
- Brockmole, J. R., and J. M. Henderson. 2005a. Prioritization of new objects in real-world scenes: Evidence from eye movements. Journal of Experimental Psychology: Human Perception and Performance 31, 5: 857–868. [Google Scholar] [CrossRef] [PubMed]
- Brockmole, J. R., and J. M. Henderson. 2005b. Object appearance, disappearance, and attention prioritization in real-world scenes. Psychonomic bulletin & review 12, 6: 1061–1067. [Google Scholar]
- Burr, D. C., and J. Ross. 1982. Contrast sensitivity at high velocities. Vision research 22, 4: 479–484. [Google Scholar] [CrossRef]
- Campbell, F. W., and R. H. Wurtz. 1978. Saccadic omission: Why we do not see a grey-out during a saccadic eye movement. Vision Research 18, 10: 1297–1303. [Google Scholar] [CrossRef]
- Castet, E. 2010. Edited by U. J. Ilg and G. S. Masson. Perception of intra-saccadic motion. In Dynamics of Visual Motion Processing. Springer Science: pp. 213–238. [Google Scholar]
- Castet, E., and G. S. Masson. 2000. Motion perception during saccadic eye movements. Nature Neuroscience 3, 2: 177–183. [Google Scholar] [CrossRef]
- Chua, F. K. 2009. A new object captures attention—But only when you know it’s new. Attention, Perception, & Psychophysics 71, 4: 699–711. [Google Scholar] [CrossRef]
- Davoli, C. C., J. W. Suszko, and R. A. Abrams. 2007. New objects can capture attention without a unique luminance transient. Psychonomic Bulletin & Review 14, 2: 338–343. [Google Scholar] [CrossRef]
- Deubel, H., and W. X. Schneider. 1996. Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Research 36, 12: 1827–1837. [Google Scholar] [CrossRef]
- Dienes, Z. 2011. Bayesian versus orthodox statistics: Which side are you on? Perspectives on Psychological Science 6, 3: 274–290. [Google Scholar] [CrossRef]
- Franconeri, S. L., A. Hollingworth, and D. J. Simons. 2005. Do new objects capture attention? Psychological Science 16, 4: 275–281. [Google Scholar] [CrossRef] [PubMed]
- Franconeri, S. L., and D. J. Simons. 2003. Moving and looming stimuli capture attention. Attention, Perception, & Psychophysics 65, 7: 999–1010. [Google Scholar]
- Godijn, R., and J. Pratt. 2002. Endogenous saccades are preceded by shifts of visual attention: Evidence from cross-saccadic priming effects. Acta Psychologica 110, 1: 83–102. [Google Scholar] [CrossRef] [PubMed]
- Golomb, J. D., M. M. Chun, and J. A. Mazer. 2008. The native coordinate system of spatial attention is retinotopic. Journal of Neuroscience 28, 42: 10654–10662. [Google Scholar] [CrossRef] [PubMed]
- Golomb, J. D., A. C. Marino, M. M. Chun, and J. A. Mazer. 2011. Attention doesn’t slide: Spatiotopic updating after eye movements instantiates a new, discrete attentional locus. Attention, Perception, & Psychophysics 73: 1–8. [Google Scholar]
- Green, M. 1991. Visual search, visual streams, and visual architectures. Attention, Perception, & Psychophysics 50, 4: 388–403. [Google Scholar]
- Green, M. 1992. Visual search: Detection, identification, and localization. Perception 21: 765. [Google Scholar] [CrossRef]
- Grimes, J. 1996. On the failure to detect changes in scenes across saccades. Perception 5: 89–110. [Google Scholar]
- Hickok, G. 2012. Computational neuroanatomy of speech production. Nature Reviews Neuroscience 13, 2: 135–145. [Google Scholar] [CrossRef]
- Hollingworth, A., D. J. Simons, and S. L. Franconeri. 2010. New objects do not capture attention without a sensory transient. Attention, Perception & Psychophysics 72, 5: 1298–1310. [Google Scholar] [CrossRef]
- Irwin, D. E. 1991. Information integration across saccadic eye movements. Cognitive Psychology 23, 3: 420–456. [Google Scholar] [CrossRef]
- Irwin, D. E. 1992. Memory for position and identity across eye movements. Journal of Experimental Psychology: Learning, Memory, and Cognition 18, 2: 307–317. [Google Scholar] [CrossRef]
- Irwin, D. E. 1996. Integrating information across saccadic eye movements. Current Directions in Psychological Science 5, 3: 94–100. [Google Scholar] [CrossRef]
- Itti, L., and C. Koch. 2001. Computational modelling of visual attention. Nature Reviews Neuroscience 2, 3: 194–204. [Google Scholar] [CrossRef] [PubMed]
- Itti, L., C. Koch, and E. Niebur. 1998. A model of saliency-based visual attention for rapid scene analysis. IEEE Transactions on Pattern Analysis and Machine Intelligence 20, 11: 1254–1259. [Google Scholar] [CrossRef]
- Jeffreys, H. 1961. Theory of Probability. Oxford University Press: Oxford, United Kingdom. [Google Scholar]
- Jonides, J., and S. Yantis. 1988. Uniqueness of abrupt visual in capturing attention. Perception & Psychophysics 43, 4: 346–354. [Google Scholar]
- Kahneman, D., A. Treisman, and B. J. Gibbs. 1992. The reviewing of object files: Object-specific integration of information. Cognitive psychology 24, 2: 175–219. [Google Scholar] [CrossRef]
- Koch, C., and S. Ullman. 1985. Shifts in selective visual attention: Towards the underlying neural circuitry. Human Neurobiology 4, 4: 219–227. [Google Scholar]
- Luck, S. J., and E. K. Vogel. 1997. The capacity of visual working memory for features and conjunctions. Nature 390: 279–281. [Google Scholar] [CrossRef]
- Mack, A., and I. Rock. 1998. Inattentional blindness: Perception without attention. MIT Press: Cambridge, MA. [Google Scholar]
- MacKay, D. M. 1972. Visual stability. Investigative Ophthalmology 11: 518–524. [Google Scholar]
- Mathôt, S., D. Schreij, and J. Theeuwes. in press. OpenSesame: An open-source, graphical experiment builder for the social sciences. In Behavior Research Methods. [Google Scholar]
- Mathôt, S., and J. Theeuwes. 2010a. Gradual remapping results in early retinotopic and late spatiotopic inhibition of return. Psychological Science 21, 12: 1793–1798. [Google Scholar] [CrossRef]
- Mathôt, S., and J. Theeuwes. 2010b. Evidence for the predictive remapping of visual attention. Experimental Brain Research 200, 1: 117–122. [Google Scholar] [CrossRef] [PubMed]
- Mathôt, S., and J. Theeuwes. 2011a. Visual attention and stability. Philosophical Transactions of the Royal Society B: Biological Sciences 366: 516–527. [Google Scholar] [CrossRef]
- Mathôt, S., and J. Theeuwes. 2011b. Mantra: An open method for object and movement tracking. Behavior Research Methods 43, 4: 1182–1193. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matin, E. 1974. Saccadic suppression: A review and an analysis. Psychological Bulletin 81, 12: 899–917. [Google Scholar] [CrossRef] [PubMed]
- Morris, A. P., M. Kubischik, K.-P. Hoffmann, B. Krekelberg, and F. Bremmer. 2012. Dynamics of eyeposition signals in the dorsal visual system. Current Biology 22, 3: 173–179. [Google Scholar] [CrossRef]
- Mulckhuyse, M., D. Talsma, and J. Theeuwes. 2007. Grabbing attention without knowing: Automatic capture of attention by subliminal spatial cues. Visual Cognition 15, 7: 779–788. [Google Scholar] [CrossRef]
- Pertzov, Y., E. Zohary, and G. Avidan. 2010. Rapid formation of spatiotopic representations as revealed by inhibition of return. Journal of Neuroscience 30, 26: 8882–8887. [Google Scholar] [CrossRef]
- Posner, M. I. 1980. Orienting of attention. Quarterly Journal of Experimental Psychology 32, 1: 3–25. [Google Scholar] [CrossRef]
- Prime, S. L., L. Tsotsos, G. Keith, and J. Crawford. 2007. Visual memory capacity in transsaccadic integration. Experimental Brain Research 180, 4: 609–628. [Google Scholar] [CrossRef]
- Prime, S. L., M. Vesia, and J. D. Crawford. 2011. Cortical mechanisms for trans-saccadic memory and integration of multiple object features. Philosophical Transactions of the Royal Society B: Biological Sciences 366, 1564: 540–553. [Google Scholar] [CrossRef] [PubMed]
- Rensink, R. A., J. K. O’Regan, and J. J. Clark. 1997. The need for attention to perceive changes in scenes. Psychological Science 8, 5: 368–373. [Google Scholar] [CrossRef]
- Sagi, D., and B. Julesz. 1987. Short-range limitation on detection of feature differences. Spatial Vision 2, 1: 39–49. [Google Scholar]
- Theeuwes, J. 1994. Stimulus-driven capture and attentional set: Selective search for color and visual abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance 20: 799. [Google Scholar] [CrossRef]
- Theeuwes, J. 2010. Top-down and bottom-up control of visual selection. Acta Psychologica 135: 77–99. [Google Scholar] [CrossRef]
- Theeuwes, J., A. F. Kramer, S. Hahn, and D. E. Irwin. 1998. Our eyes do not always go where we want them to go: Capture of the eyes by new objects. Psychological Science 9, 5: 379–385. [Google Scholar] [CrossRef]
- Treisman, A. M., and G. Gelade. 1980. A feature-integration theory of attention. Cognitive Psychology 12, 1: 97–136. [Google Scholar] [CrossRef]
- Volkmann, F. C., L. A. Riggs, K. D. White, and R. K. Moore. 1978. Contrast sensitivity during saccadic eye movements. Vision Research 18, 9: 1193–1199. [Google Scholar] [CrossRef]
- Wetzels, R., D. Matzke, M. D. Lee, J. N. Rouder, G. J. Iverson, and E.-J. Wagenmakers. 2011. Statistical evidence in experimental psychology. Perspectives on Psychological Science 6, 3: 291–298. [Google Scholar] [CrossRef]
- Yantis, S., and A. P. Hillstrom. 1994. Stimulus-driven attentional capture: Evidence from equiluminant visual objects. Journal of Experimental Psychology: Human Perception and Performance 20, 1: 95–107. [Google Scholar] [CrossRef] [PubMed]
- Yantis, S., and J. Jonides. 1984. Abrupt visual onsets and selective attention: Evidence from visual search. [Google Scholar]
- Journal of Experimental Psychology: Human Perception and Performance 10, 5: 601–621.
- Yantis, S., and J. Jonides. 1996. Attentional capture by abrupt onsets: New perceptual objects or visual masking? Journal of Experimental Psychology: Human Perception and Performance 22, 6: 1505–1513. [Google Scholar] [CrossRef] [PubMed]


© 2012 by the authors. This article is licensed under a Creative Commons Attribution 4.0 International License.