In Silico Analysis of Relationship between Proteins from Plastid Genome of Red Alga Palmaria sp. (Japan) and Angiotensin I Converting Enzyme Inhibitory Peptides



Abstract

1. Introduction
2. Results and Discussion
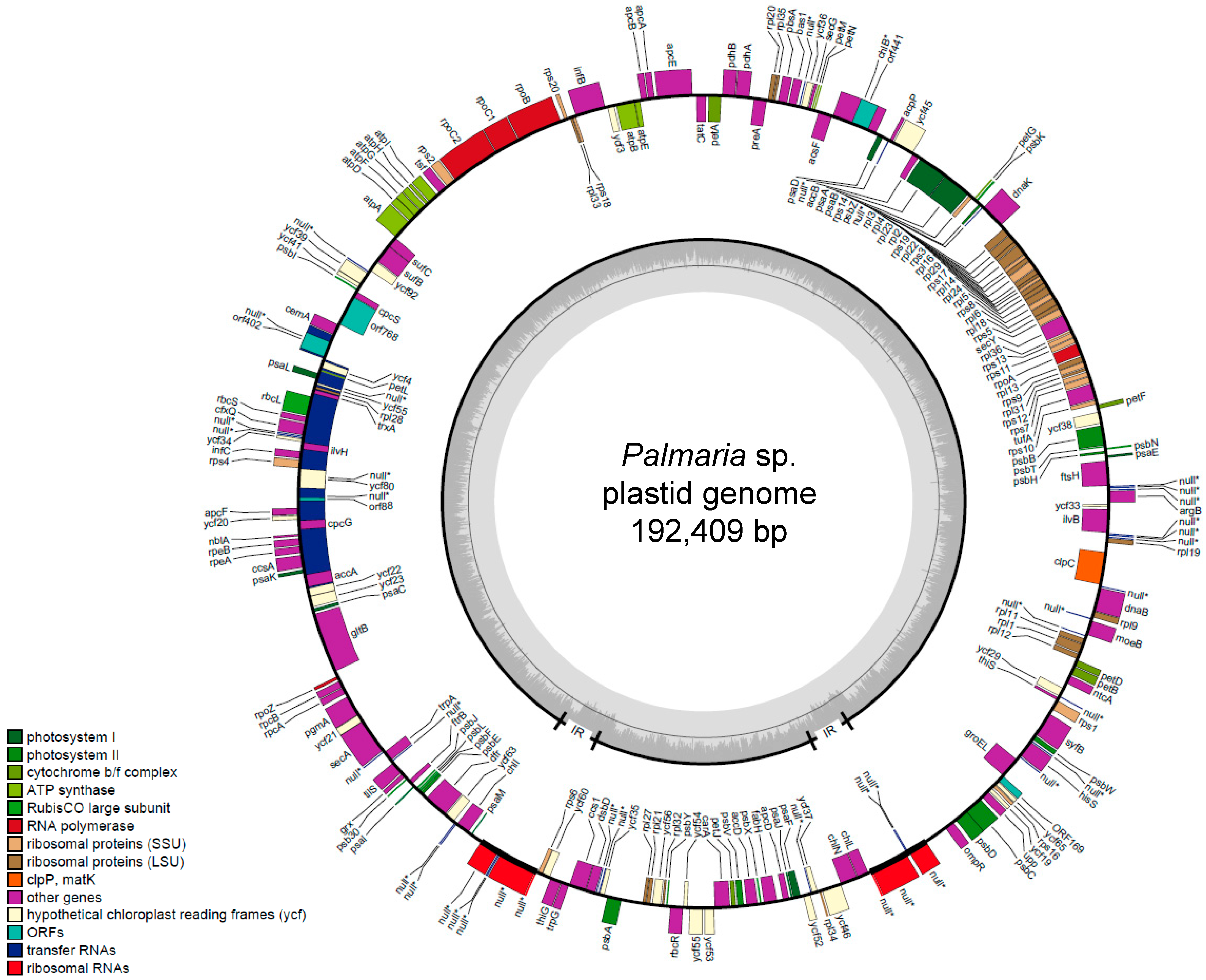
2.1. General Features of Palmaria sp. (Japan) Plastid Genomes
2.2. Comparison of Amino Acid (AA) Composition between Palmaria sp. (Japan) Plastid Proteins and Proximate AA in P. palmata
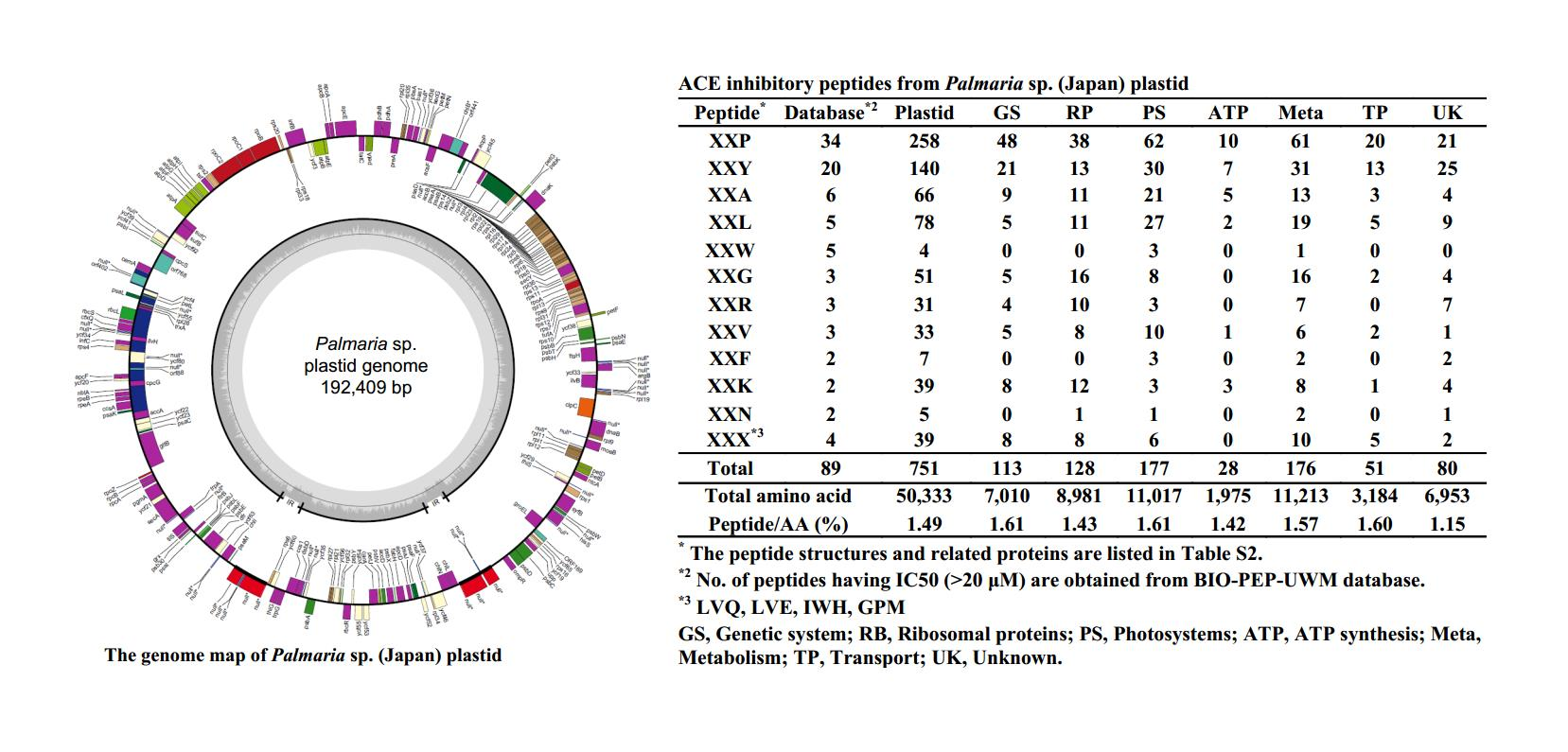
2.3. ACE Inhibitory Peptides in Plastid
2.4. ACE Inhibitory Peptides in Photosystems
2.5. Comparison of ACE Inhibitory Peptides in Palmaria sp. (Japan) and P. palmata
3. Materials and Methods
3.1. Plastid Genome Construction
3.2. Collection of ACE Inhibitory Peptides and Comparison with Plastid Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Renn, D. Biotechnology and the red seaweed polysaccharide industry: Status, needs and prospects. Trends Biotechnol. 1997, 15, 9–14. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Biological activities and potential industrial applications of fucose rich sulfated polysaccharides and fucoidans isolated from brown seaweeds: A review. Carbohydr. Polym. 2012, 88, 13–20. [Google Scholar] [CrossRef]
- Maehre, H.K.; Edvinsen, G.K.; Eilertsen, K.E.; Elvevoll, E.O. Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria Palmata), but not in the brown seaweed winged kelp (Alaria Esculenta). J. Appl. Phycol. 2016, 28, 581–590. [Google Scholar] [CrossRef]
- Mai, K.; Mercer, J.P.; Donlon, J. Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L. and Haliotis discus hannai Ino: II. Amino acid composition of abalone and six species of macroalgae with an assessment of their nutritional value. Aquaculture 1994, 128, 115–130. [Google Scholar] [CrossRef]
- Grote, B. Recent Developments in aquaculture of Palmaria palmata (Linnaeus) (Weber & Mohr 1805): Cultivation and uses. Rev. Aquac. 2019, 11, 25–41. [Google Scholar] [CrossRef]
- Gantt, E. Phycobilisomes. Annu. Rev. Plant Physiol. 1981, 32, 327–347. [Google Scholar] [CrossRef]
- Sato, N.; Furuta, T.; Takeda, T.; Miyabe, Y.; Ura, K.; Takagi, Y.; Yasui, H.; Kumagai, Y.; Kishimura, H. Antioxidant activity of proteins extracted from red alga dulse harvested in Japan. J. Food Biochem. 2018, 43, e12709. [Google Scholar] [CrossRef]
- Sonani, R.R.; Singh, N.K.; Kumar, J.; Thakar, D.; Madamwar, D. Concurrent purification and antioxidant activity of phycobiliproteins from Lyngbya Sp. A09DM: An antioxidant and anti-aging potential of phycoerythrin in Caenorhabditis Elegans. Process Biochem. 2014, 49, 1757–1766. [Google Scholar] [CrossRef]
- Beaulieu, L.; Sirois, M.; Tamigneaux, É. Evaluation of the in vitro biological activity of protein hydrolysates of the edible red alga, Palmaria palmata (dulse) harvested from the Gaspe coast and cultivated in tanks. J. Appl. Phycol. 2016, 28, 3101–3115. [Google Scholar] [CrossRef]
- Cao, D.; Lv, X.; Xu, X.; Yu, H.; Sun, X.; Xu, N. Purification and identification of a novel ACE inhibitory peptide from marine alga Gracilariopsis lemaneiformis protein hydrolysate. Eur. Food Res. Technol. 2017, 243, 1829–1837. [Google Scholar] [CrossRef]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. In vitro assessment of the cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Soler-Vila, A.; Edwards, M.D.; FitzGerald, R.J. The effect of time and origin of harvest on the in vitro biological activity of Palmaria palmata protein hydrolysates. Food Res. Int. 2014, 62, 746–752. [Google Scholar] [CrossRef]
- He, H.L.; Chen, X.L.; Wu, H.; Sun, C.Y.; Zhang, Y.Z.; Zhou, B.C. High throughput and rapid screening of marine protein hydrolysates enriched in peptides with angiotensin-I-converting enzyme inhibitory activity by capillary electrophoresis. Bioresour. Technol. 2007, 98, 3499–3505. [Google Scholar] [CrossRef] [PubMed]
- Kitade, Y.; Miyabe, Y.; Yamamoto, Y.; Takeda, H.; Shimizu, T.; Yasui, H.; Kishimura, H. Structural characteristics of phycobiliproteins from red alga Mazzaella japonica. J. Food Biochem. 2018, 42, 1–10. [Google Scholar] [CrossRef]
- Miyabe, Y.; Furuta, T.; Takeda, T.; Kanno, G.; Shimizu, T.; Tanaka, Y.; Gai, Z.; Yasui, H.; Kishimura, H. Structural properties of phycoerythrin from dulse Palmaria palmata. J. Food Biochem. 2017, 41, e12301. [Google Scholar] [CrossRef]
- Cian, R.E.; Alaiz, M.; Vioque, J.; Drago, S.R. Enzyme proteolysis enhanced extraction of ACE inhibitory and antioxidant compounds (peptides and polyphenols) from Porphyra columbina residual cake. J. Appl. Phycol. 2013, 25, 1197–1206. [Google Scholar] [CrossRef]
- Cha, S.H.; Lee, K.W.; Jeon, Y.J. Screening of extracts from red algae in Jeju for potentials marine angiotensin-I converting enzyme (ACE) inhibitory activity. Algae 2006, 21, 343–348. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A.; Nakano, T. Angiotensin I-converting enzyme inhibitory peptides derived from wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef]
- Cho, C.H.; Choi, J.W.; Lam, D.W.; Kim, K.M.; Yoon, H.S. Plastid genome analysis of three nemaliophycidae red algal species suggests environmental adaptation for iron limited habitats. PLoS ONE 2018, 13, e0196995. [Google Scholar] [CrossRef] [PubMed]
- Janouskovec, J.; Liu, S.L.; Martone, P.T.; Carre, W.; Leblanc, C.; Collen, J.; Keeling, P.J. Evolution of red algal plastid genomes: Ancient architectures, introns, horizontal gene transfer, and taxonomic utility of plastid markers. PLoS ONE 2013, 8, e59001. [Google Scholar] [CrossRef] [PubMed]
- Salomaki, E.D.; Nickles, K.R.; Lane, C.E. The Ghost Plastid of Choreocolax Polysiphoniae. J. Phycol. 2015, 51, 217–221. [Google Scholar] [CrossRef]
- Lee, J.M.; Cho, C.H.; Park, S.I.; Choi, J.W.; Song, H.S.; West, J.A.; Bhattacharya, D.; Yoon, H.S. Parallel evolution of highly conserved plastid genome architecture in red seaweeds and seed plants. BMC Biol. 2016, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, K.M.; Yang, E.C.; Miller, K.A.; Boo, S.M.; Bhattacharya, D.; Yoon, H.S. Reconstructing the complex evolutionary history of mobile plasmids in red algal genomes. Sci. Rep. 2016, 6, 23744. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Nakozoe, J. Production and use of marine algae in Japan. Jpn. Agric. Res. Q. 2001, 35, 281–290. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.V.; Fleurence, J.; Lamghari, R.; Luçon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.P.; Villaume, C.; Guéant, J.L. Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Sfriso, A.A.; Gallo, M.; Baldi, F. Phycoerythrin productivity and diversity from five red macroalgae. J. Appl. Phycol. 2018, 30, 2523–2531. [Google Scholar] [CrossRef]
- Fleurence, J.; Massiani, L.; Guyader, O.; Mabeau, S. Use of enzymatic cell wall degradation for improvement of protein extraction from Chondrus crispus, Gracilaria verrucosa and Palmaria palmata. J. Appl. Phycol. 1995, 7, 393–397. [Google Scholar] [CrossRef]
- Cota-Sánchez, J.H.; Remarchuk, K.; Ubayasena, K. Ready-to-use DNA extracted with a CTAB method adapted for herbarium specimens and mucilaginous plant tissue. Plant Mol. Biol. Rep. 2006, 24, 161–167. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—A suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, 575–581. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Classification | No. | Gene | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genetic System | Maintenance | 2 | dnaB | rne | ||||||
| RNA polymerase | 5 | rpoA | rpoB | rpoC1 | rpoC2 | rpoZ | ||||
| Transcription factors | 4 | ntcA | ompR | rbcR | ycf29 | |||||
| Translation | 4 | infB | infC | tsf | tufA | |||||
| Ribosomal Proteins | Large subunit | 28 | rpl1 | rpl2 | rpl3 | rpl4 | rpl5 | rpl6 | rpl9 | rpl11 |
| rpl12 | rpl13 | rpl14 | rpl16 | rpl18 | rpl19 | rpl20 | rpl21 | |||
| rpl22 | rpl23 | rpl24 | rpl27 | rpl28 | rpl29 | rpl31 | rpl32 | |||
| rpl33 | rpl34 | rpl35 | rpl36 | |||||||
| Small subunit | 19 | rps1 | rps2 | rps3 | rps4 | rps5 | rps6 | rps7 | rps8 | |
| rps9 | rps10 | rps11 | rps12 | rps13 | rps14 | rps16 | rps17 | |||
| rps18 | rps19 | rps20 | ||||||||
| tRNA processing | 1 | tilS | ||||||||
| Protein quality control | 4 | clpC | dnaK | ftsH | groEL | |||||
| Photosystems | Phycobilisomes | 12 | apcA | apcB | apcD | apcE | apcF | rpcA | rpcB | cpcG |
| cpcS | rpeA | rpeB | nblA | |||||||
| Photosystem I | 13 | psaA | psaB | psaC | psaD | psaE | psaF | psaI | psaJ | |
| psaK | psaL | psaM | ycf3 | ycf4 | ||||||
| Photosystem II | 19 | psbA | psbB | psbC | psbD | psbE | psbF | psbH | psbI | |
| psbJ | psbK | psbL | psbN | psbT | psbV | |||||
| psbW | psbX | psbY | psbZ | psb30 | ||||||
| Cytochrome complex | 11 | ccs1 | ccsA | petA | petB | petD | petF | petG | petJ | |
| petL | petM | petN | ||||||||
| Redox system | 7 | acsF | bas1 | dsbD | ftrB | grx | pbsA | trxA | ||
| ATP Synthesis | ATP synthase | 8 | atpA | atpB | atpD | atpE | atpF | atpG | atpH | atpI |
| Metabolism | Carbohydrates | 6 | cfxQ | pdhA | pdhB | pgmA | rbcL | rbcS | ||
| Lipids | 5 | accA | accB | accD | acpP | fabH | ||||
| Nucleotides | 2 | carA | upp | |||||||
| Amino acids (AAs) | 8 | argB | gltB | ilvB | ilvH | hisS | syfB | trpA | trpG | |
| Cofactors | 8 | chlB | chlI | chlL | chlN | moeB | preA | thiG | thiS | |
| Secondary metabolites | 1 | dfr | ||||||||
| Transport | Transport | 9 | cemA | secA | secG | secY | sufB | sufC | tatC | ycf38 |
| ycf63 | ||||||||||
| Unknown | Conserved open reding frame (ORF)s | 23 | ycf19 | ycf20 | ycf21 | ycf22 | ycf23 | ycf33 | ycf34 | ycf35 |
| ycf36 | ycf37 | ycf39 | ycf41 | ycf45 | ycf46 | ycf52 | ycf53 | |||
| ycf54 | ycf55 | ycf56 | ycf60 | ycf65 | ycf80 | ycf92 | ||||
| Unique ORFs | 6 | orf55 | orf88 | orf169 | orf402 | orf441 | orf768 | |||
| Total genes | 205 | |||||||||
| Subclass | Species | General Characteristics | RNAs | GenBank Accession | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|
| Total nt | GC% * | Introns | PCG *2 | tRNA | rRNA | ||||
| Nemaliophycidae | Palmaria sp. (Japan) | 192,410 | 34.6 | 2 | 205 | 33 | 6 | AB807662 | This study |
| Palmaria palmata | 192,960 | 33.9 | 2 | 205 | 33 | 6 | NC_031147 | [25] | |
| Kumanoa americana hys120 | 184,025 | 29.3 | 2 | 201 | 32 | 3 | NC_031178 | [25] | |
| Thorea hispida hsy077 | 175,193 | 28.3 | 2 | 194 | 31 | 3 | NC_031171 | [25] | |
| Corallinophycidae | Calliarthron tuberculosum | 178,981 | 29.2 | 2 | 202 | 33 | 3 | NC_021075 | [26] |
| Sporolithon durum | 191,464 | 29.3 | 2 | 207 | 30 | 3 | NC_029857 | [27] | |
| Ahnfeltiophycidae | Ahnfeltia plicata | 190,451 | 32.5 | 1 | 207 | 31 | 6 | NC_031145 | [28] |
| Rhodymeniophycidae | Asparagopsis taxiformis | 177,091 | 29.4 | 2 | 205 | 32 | 3 | NC_031148 | [28] |
| Ceramium japonicum | 171,634 | 27.8 | 1 | 202 | 29 | 3 | NC_031174 | [28] | |
| Rhodymenia pseudopalmata | 194,153 | 32.0 | 1 | 202 | 32 | 6 | NC_031144 | [28] | |
| Vertebrata lanosa | 167,158 | 30.0 | 0 | 193 | 28 | 3 | KP308097 | [29] | |
| AA | Plastid | GS | RP | PS | ATP | Meta | TP | UK | P. palmata | |
|---|---|---|---|---|---|---|---|---|---|---|
| % of AA | % of Total AA a or Protein b | |||||||||
| Alanine | 6.4 | 5.6 | 6.8 | 7.6 | 8.7 | 6.4 | 5.8 | 4.5 | 7.5 a | 6.7 b |
| Arginine | 4.6 | 5.1 | 6.8 | 4.1 | 3.8 | 4.0 | 3.6 | 4.1 | 6.2 a | 5.1 b |
| Aspartic acid | 4.5 | 5.6 | 4.1 | 4.0 | 4.6 | 5.4 | 3.9 | 3.8 | 9.3 a | 18.5 b |
| Asparagine | 5.5 | 5.9 | 5.2 | 4.8 | 4.2 | 5.6 | 5.6 | 6.4 | ||
| Cystine | 1.1 | 0.8 | 0.7 | 1.0 | 0.2 | 1.4 | 1.2 | 1.5 | 1.3 a | 0 b |
| Glutamic acid | 5.7 | 6.4 | 6.2 | 4.6 | 7.2 | 6.0 | 6.2 | 4.7 | 13 a | 9.9 b |
| Glutamine | 4.2 | 4.5 | 4.1 | 3.8 | 5.5 | 4.3 | 3.9 | 4.3 | ||
| Glycine | 6.3 | 5.7 | 7.0 | 7.6 | 7.1 | 6.6 | 5.4 | 3.7 | 7.2 a | 13.3 b |
| Histidine | 1.9 | 1.9 | 1.9 | 1.9 | 0.7 | 2.3 | 1.4 | 2.1 | 2.1 a | 0.5 b |
| Isoleucine | 9.0 | 10.0 | 8.7 | 7.5 | 9.3 | 9.0 | 11.1 | 10.0 | 5.3 a | 3.7 b |
| Leucine | 10.6 | 10.1 | 8.7 | 10.4 | 12.0 | 10.4 | 12.2 | 12.9 | 7.8 a | 7.1 b |
| Lysine | 6.5 | 7.3 | 9.3 | 4.4 | 5.5 | 5.8 | 5.3 | 7.0 | 8.2 a | 3.3 b |
| Methionine | 2.2 | 1.8 | 2.1 | 2.6 | 2.0 | 2.4 | 1.9 | 1.8 | 1.9 a | 2.7 b |
| Phenylalanine | 4.1 | 3.2 | 2.7 | 5.8 | 3.4 | 3.5 | 4.8 | 5.1 | 5.2 a | 5.1 b |
| Proline | 3.7 | 3.6 | 3.6 | 4.1 | 3.7 | 3.9 | 3.1 | 3.4 | 4.4 a | |
| Serine | 7.4 | 7.0 | 6.3 | 7.8 | 6.8 | 7.2 | 7.8 | 8.6 | 4.6 a | 6.3 b |
| Threonine | 5.6 | 5.2 | 5.8 | 5.6 | 5.9 | 5.6 | 6.0 | 5.3 | 4.5 a | 3.6 b |
| Tryptophan | 1.0 | 0.5 | 0.5 | 1.9 | 0.4 | 0.8 | 0.9 | 1.3 | ||
| Tyrosine | 3.6 | 3.5 | 2.6 | 3.9 | 2.4 | 3.4 | 4.2 | 4.7 | 4.5 a | 3.4 b |
| Valine | 6.3 | 6.6 | 7.0 | 6.6 | 6.9 | 6.2 | 5.9 | 4.9 | 7.3 a | 6.9 b |
| Total AA | 50,333 | 7010 | 8981 | 11,017 | 1975 | 11,213 | 3184 | 6953 | ||
| Peptide * | Database *2 | Plastid | GS | RP | PS | ATP | Meta | TP | UK |
|---|---|---|---|---|---|---|---|---|---|
| XXP | 34 | 260 | 48 | 38 | 62 | 10 | 61 | 20 | 21 |
| XXY | 20 | 140 | 21 | 13 | 30 | 7 | 31 | 13 | 25 |
| XXA | 6 | 66 | 9 | 11 | 21 | 5 | 13 | 3 | 4 |
| XXL | 5 | 78 | 5 | 11 | 27 | 2 | 19 | 5 | 9 |
| XXW | 5 | 4 | 0 | 0 | 3 | 0 | 1 | 0 | 0 |
| XXG | 3 | 51 | 5 | 16 | 8 | 0 | 16 | 2 | 4 |
| XXR | 3 | 31 | 4 | 10 | 3 | 0 | 7 | 0 | 7 |
| XXV | 3 | 33 | 5 | 8 | 10 | 1 | 6 | 2 | 1 |
| XXF | 2 | 7 | 0 | 0 | 3 | 0 | 2 | 0 | 2 |
| XXK | 2 | 39 | 8 | 12 | 3 | 3 | 8 | 1 | 4 |
| XXN | 2 | 5 | 0 | 1 | 1 | 0 | 2 | 0 | 1 |
| XXX *3 | 4 | 39 | 8 | 8 | 6 | 0 | 10 | 5 | 2 |
| Total | 89 | 751 | 113 | 128 | 177 | 28 | 176 | 51 | 80 |
| Total AA | 50,333 | 7010 | 8981 | 11,017 | 1975 | 11,213 | 3184 | 6953 | |
| Peptide/AA (%) | 1.49 | 1.61 | 1.43 | 1.61 | 1.42 | 1.57 | 1.60 | 1.15 | |
| Peptide * | PBS | PSI | PSII | Cc | Red |
|---|---|---|---|---|---|
| XXP | 11 | 21 | 17 | 7 | 6 |
| XXY | 9 | 8 | 7 | 3 | 3 |
| XXA | 6 | 4 | 8 | 1 | 2 |
| XXL | 6 | 12 | 4 | 5 | 0 |
| XXW | 0 | 3 | 0 | 0 | 0 |
| XXG | 3 | 1 | 4 | 0 | 0 |
| XXR | 1 | 1 | 1 | 0 | 0 |
| XXV | 3 | 2 | 3 | 2 | 0 |
| XXF | 0 | 0 | 2 | 1 | 0 |
| XXK | 0 | 1 | 2 | 0 | 0 |
| XXN | 0 | 0 | 0 | 0 | 1 |
| XXX ** | 3 | 0 | 3 | 0 | 0 |
| Total | 42 | 53 | 51 | 19 | 12 |
| Total AA | 2644 | 2654 | 2582 | 1784 | 1353 |
| Peptide/AA (%) | 1.59 | 2.00 | 1.98 | 1.07 | 0.89 |
| PBS | No. * | PSI | No. * | PSII | No. * | PSII | No. * | Cc | No. * | Red | No. * |
|---|---|---|---|---|---|---|---|---|---|---|---|
| apcA | 2 | psaA | 18 | psbA | 9 | psbV | 2 | ccs1 | 5 | acsF | 2 |
| apcB | 2 | psaB | 13 | psbB | 11 | psbW | 1 | ccsA | 0 | bas1 | 1 |
| apcD | 3 | psaC | 1 | psbC | 10 | psbX | 0 | petA | 5 | dsbD | 4 |
| apcE | 14 | psaD | 1 | psbD | 6 | psbY | 1 | petB | 3 | ftrB | 1 |
| apcF | 3 | psaE | 2 | psbE | 1 | psbZ | 2 | petD | 1 | grx | 0 |
| cpcG | 0 | psaF | 5 | psbF | 0 | psb30 | 0 | petF | 3 | pbsA | 2 |
| cpcS | 2 | psaI | 1 | psbH | 3 | petG | 1 | trxA | 2 | ||
| rpcA | 2 | psaJ | 0 | psbI | 0 | petJ | 0 | ||||
| rpcB | 3 | psaL | 3 | psbJ | 1 | petL | 0 | ||||
| rpeA | 6 | psaM | 1 | psbK | 1 | petM | 1 | ||||
| rpeB | 4 | psbH | 1 | psbL | 1 | petN | 0 | ||||
| nblA | 1 | ycf3 | 3 | psbN | 0 | ||||||
| ycf4 | 4 | psbT | 2 | ||||||||
| Total | 42 | 53 | 51 | 19 | 12 |
| Peptide * | Database *2 | Plastid | GS | RP | PS | ATP | Meta | TP | UK |
|---|---|---|---|---|---|---|---|---|---|
| XXP | 34 | 263 | 47 | 39 | 63 | 10 | 63 | 20 | 21 |
| XXY | 20 | 133 | 23 | 13 | 29 | 7 | 28 | 12 | 21 |
| XXA | 6 | 68 | 8 | 13 | 20 | 6 | 13 | 3 | 5 |
| XXL | 5 | 79 | 6 | 10 | 25 | 2 | 20 | 5 | 11 |
| XXW | 5 | 4 | 0 | 0 | 3 | 0 | 1 | 0 | 0 |
| XXG | 3 | 51 | 6 | 16 | 8 | 0 | 16 | 2 | 3 |
| XXR | 3 | 27 | 2 | 10 | 3 | 0 | 7 | 0 | 5 |
| XXV | 3 | 33 | 5 | 8 | 9 | 1 | 7 | 2 | 1 |
| XXF | 2 | 6 | 0 | 0 | 3 | 0 | 2 | 0 | 1 |
| XXK | 2 | 37 | 9 | 11 | 3 | 4 | 7 | 1 | 2 |
| XXN | 2 | 5 | 0 | 1 | 1 | 0 | 2 | 0 | 1 |
| XXX *3 | 4 | 36 | 7 | 8 | 6 | 0 | 9 | 5 | 1 |
| Total | 89 | 742 | 113 | 129 | 173 | 30 | 175 | 50 | 72 |
| Total AA | 50,229 | 7009 | 8981 | 11,013 | 1970 | 11,237 | 3188 | 6831 | |
| Peptide/AA (%) | 1.48 | 1.61 | 1.44 | 1.57 | 1.52 | 1.56 | 1.57 | 1.05 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumagai, Y.; Miyabe, Y.; Takeda, T.; Adachi, K.; Yasui, H.; Kishimura, H. In Silico Analysis of Relationship between Proteins from Plastid Genome of Red Alga Palmaria sp. (Japan) and Angiotensin I Converting Enzyme Inhibitory Peptides. Mar. Drugs 2019, 17, 190. https://doi.org/10.3390/md17030190
Kumagai Y, Miyabe Y, Takeda T, Adachi K, Yasui H, Kishimura H. In Silico Analysis of Relationship between Proteins from Plastid Genome of Red Alga Palmaria sp. (Japan) and Angiotensin I Converting Enzyme Inhibitory Peptides. Marine Drugs. 2019; 17(3):190. https://doi.org/10.3390/md17030190
Chicago/Turabian StyleKumagai, Yuya, Yoshikatsu Miyabe, Tomoyuki Takeda, Kohsuke Adachi, Hajime Yasui, and Hideki Kishimura. 2019. "In Silico Analysis of Relationship between Proteins from Plastid Genome of Red Alga Palmaria sp. (Japan) and Angiotensin I Converting Enzyme Inhibitory Peptides" Marine Drugs 17, no. 3: 190. https://doi.org/10.3390/md17030190
APA StyleKumagai, Y., Miyabe, Y., Takeda, T., Adachi, K., Yasui, H., & Kishimura, H. (2019). In Silico Analysis of Relationship between Proteins from Plastid Genome of Red Alga Palmaria sp. (Japan) and Angiotensin I Converting Enzyme Inhibitory Peptides. Marine Drugs, 17(3), 190. https://doi.org/10.3390/md17030190