
The Neuroprotective Effect of Therapeutic Hypothermia in Cognitive Impairment of an Ischemia/Reperfusion Injury Mouse Model

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
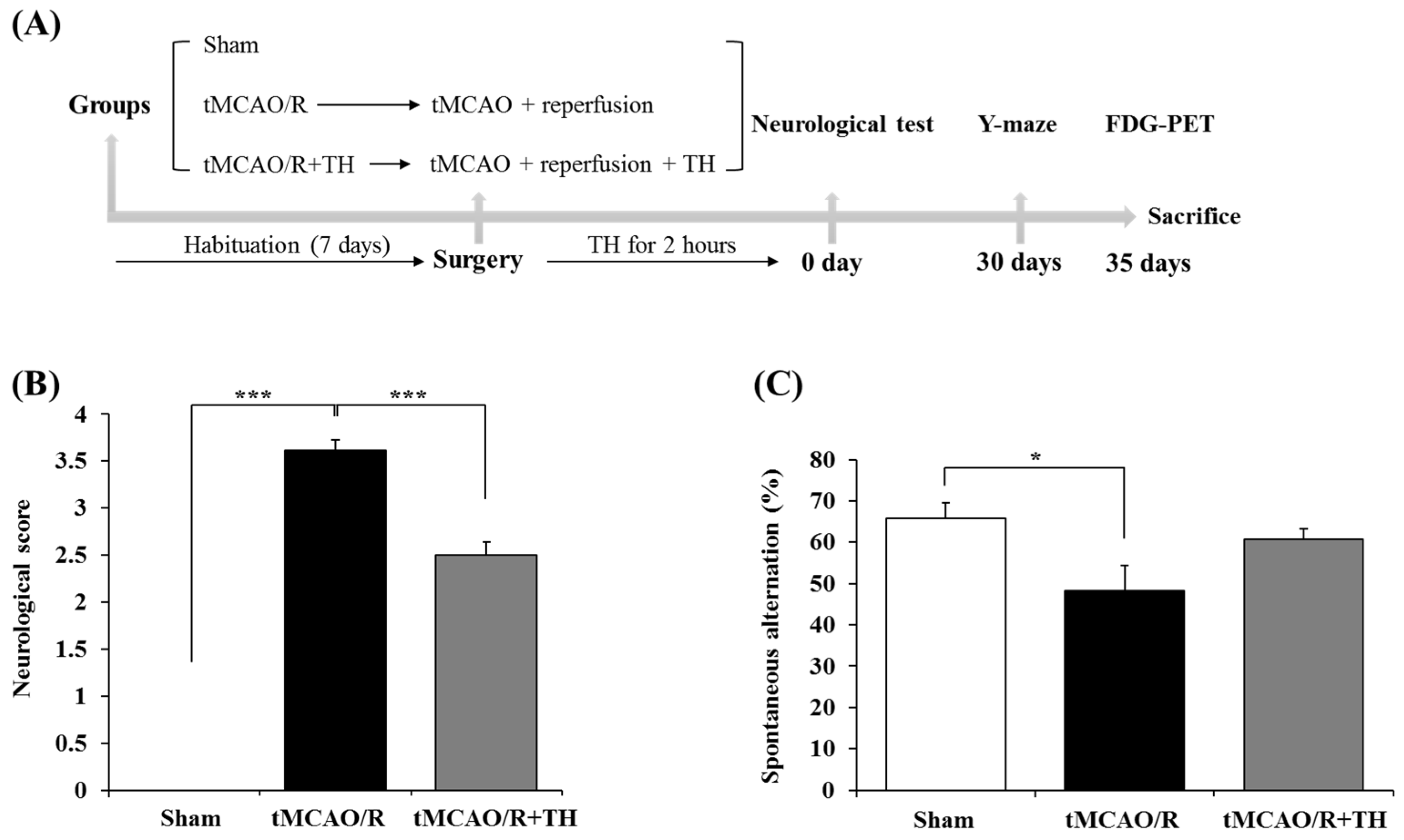
3.1. Hypothermia Improved Functional Outcome and Cognitive Function
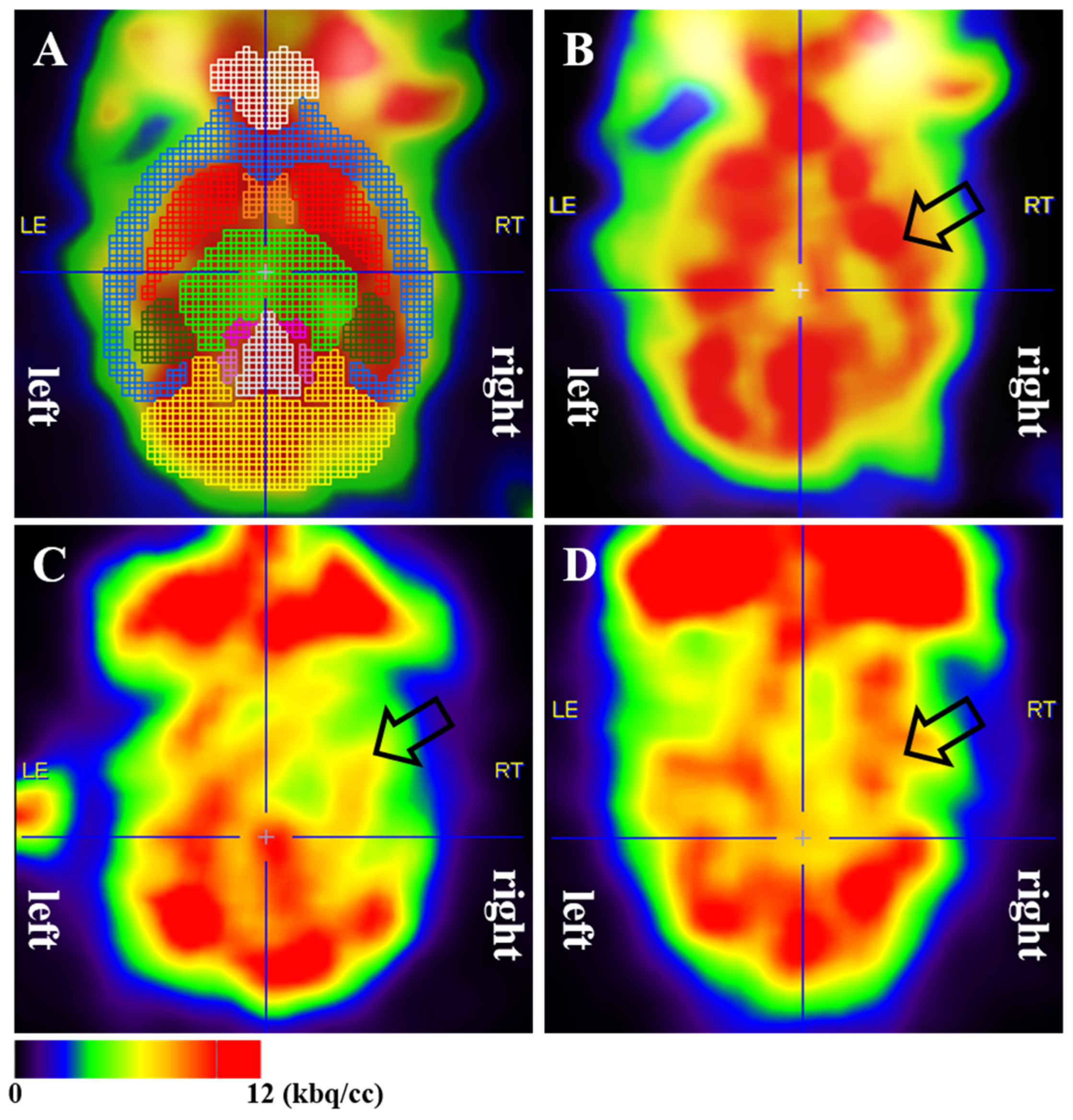
3.2. Cerebral Glucose Metabolism
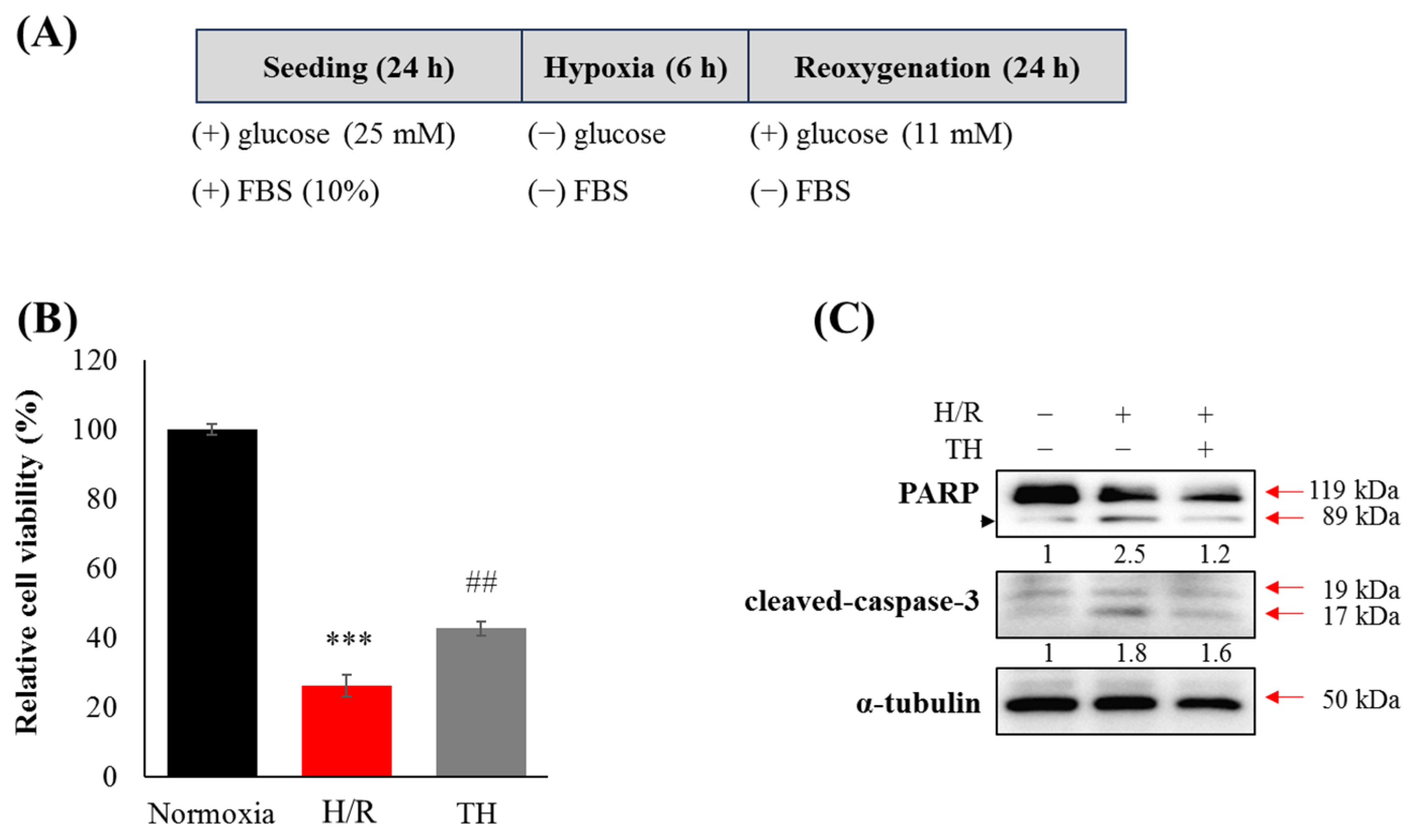
3.3. Therapeutic Hypothermia Attenuated Hypoxia/Reoxygenation(H/R)-Induced Neuronal Apoptosis
4. Discussion
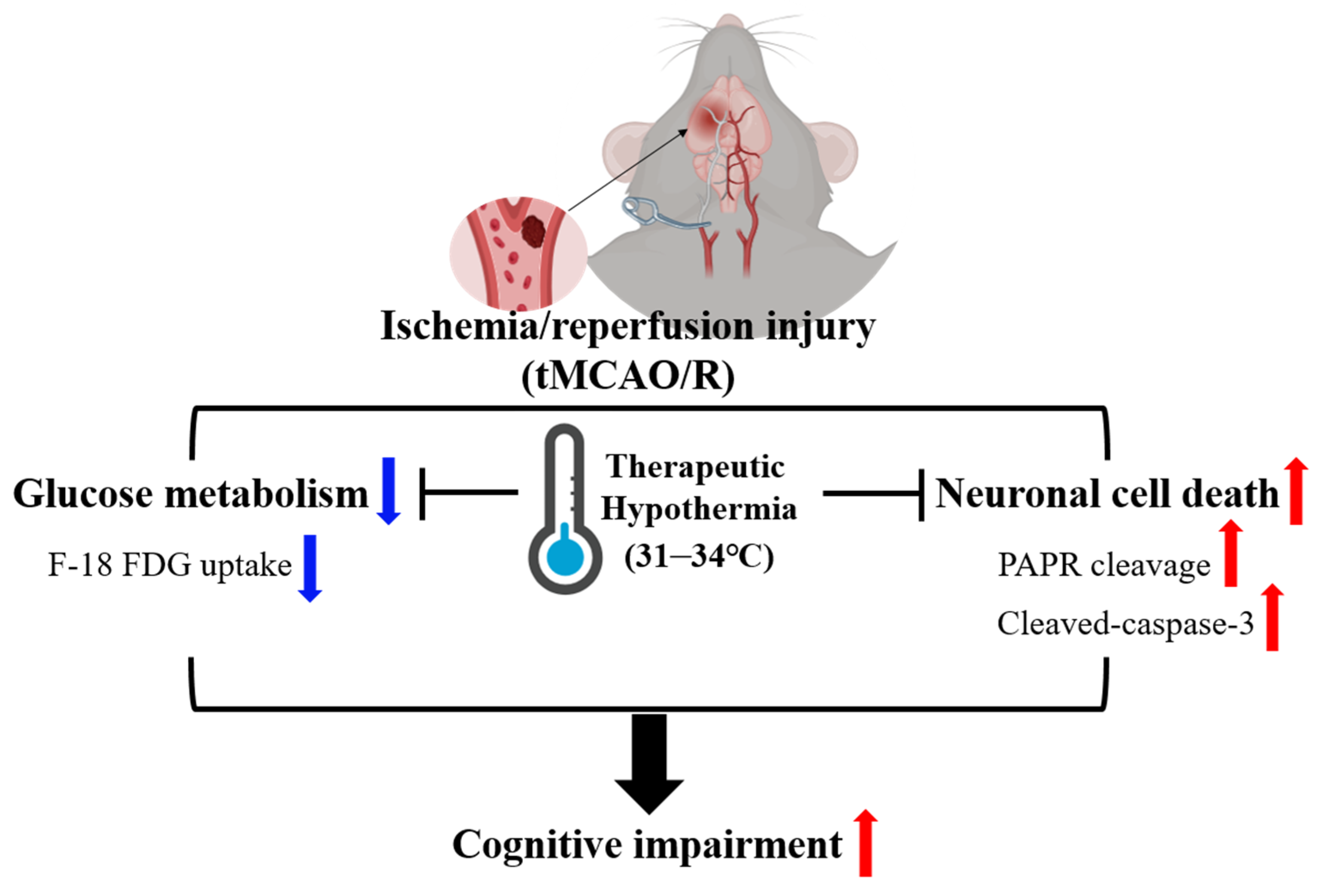
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blanc, R.; Escalard, S.; Baharvadhat, H.; Desilles, J.P.; Boisseau, W.; Fahed, R.; Redjem, H.; Ciccio, G.; Smajda, S.; Maier, B.; et al. Recent advances in devices for mechanical thrombectomy. Expert Rev. Med. Devices 2020, 17, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Mizuma, A.; You, J.S.; Yenari, M.A. Targeting reperfusion injury in the age of mechanical thrombectomy. Stroke 2018, 49, 1796–1802. [Google Scholar] [CrossRef]
- Higashi, Y.; Aratake, T.; Shimizu, T.; Shimizu, S.; Saito, M. Protective role of glutathione in the hippocampus after brain ischemia. Int. J. Mol. Sci. 2021, 22, 7765. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Du, Y.S.; Xu, W.S.; Li, C.J.; Sun, H.; Hu, K.R.; Hu, Y.Z.; Yu, T.J.; Guo, H.M.; Xie, L.; et al. Exogenous glutathione exerts a therapeutic effect in ischemic stroke rats by interacting with intrastriatal dopamine. Acta Pharmacol. Sin. 2022, 43, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Pathak, D.; Sriram, K. Neuron-astrocyte omnidirectional signaling in neurological health and disease. Front. Mol. Neurosci. 2023, 16, 1169320. [Google Scholar] [CrossRef] [PubMed]
- Iskusnykh, I.Y.; Zakharova, A.A.; Pathak, D. Glutathione in brain disorders and aging. Molecules 2022, 27, 324. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Ettelt, P.; Maier, I.L.; Schnieder, M.; Bahr, M.; Behme, D.; Psychogios, M.N.; Liman, J.; Collaborators, G.-E. Bridging therapy is associated with improved cognitive function after large vessel occlusion stroke—An analysis of the German Stroke Registry. Neurol. Res. Pract. 2020, 2, 29. [Google Scholar] [CrossRef]
- Sun, J.H.; Tan, L.; Yu, J.T. Post-stroke cognitive impairment: Epidemiology, mechanisms and management. Ann. Transl. Med. 2014, 2, 80. [Google Scholar] [CrossRef]
- Boysen, G.; Christensen, H. Stroke severity determines body temperature in acute stroke. Stroke 2001, 32, 413–417. [Google Scholar] [CrossRef]
- Walter, E.J.; Carraretto, M. The neurological and cognitive consequences of hyperthermia. Crit. Care 2016, 20, 199. [Google Scholar] [CrossRef] [PubMed]
- Diprose, W.K.; Liem, B.; Wang, M.T.M.; Sutcliffe, J.A.; Brew, S.; Caldwell, J.R.; McGuinness, B.; Campbell, D.; Barber, P.A. Impact of body temperature before and after endovascular thrombectomy for large vessel occlusion stroke. Stroke 2020, 51, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Sawada, M.; Alkayed, N.J.; Goto, S.; Crain, B.J.; Traystman, R.J.; Shaivitz, A.; Nelson, R.J.; Hurn, P.D. Estrogen receptor antagonist ICI182, 780 exacerbates ischemic injury in female mouse. J. Cereb. Blood Flow Metab. 2000, 20, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Park, M.-N.; Lim, J.S.; Baek, H.S.; Kim, S.; Kim, H.W.; Hong, J.-H. the effect of hypothermia on cognitive impairment and anxiety-like behavior in a cardiac arrest mouse model. Quant. BioSci 2023, 42, 9–16. [Google Scholar]
- Maier, C.M.; Ahern, K.v.; Cheng, M.L.; Lee, J.E.; Yenari, M.A.; Steinberg, G.K. Optimal depth and duration of mild hypothermia in a focal model of transient cerebral ischemia: Effects on neurologic outcome, infarct size, apoptosis, and inflammation. Stroke 1998, 29, 2171–2180. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, X.; Zhao, J.; Tan, X.; Liu, B.; Zhang, G.; Sun, L.; Han, D.; Chen, H.; Wang, M. Hypothermia-induced ischemic tolerance is associated with Drp1 inhibition in cerebral ischemia-reperfusion injury of mice. Brain Res. 2016, 1646, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Hattori, K.; Lee, H.; Hurn, P.D.; Crain, B.J.; Traystman, R.J.; DeVries, A.C. Cognitive deficits after focal cerebral ischemia in mice. Stroke 2000, 31, 1939–1944. [Google Scholar] [CrossRef]
- Bieber, M.; Gronewold, J.; Scharf, A.C.; Schuhmann, M.K.; Langhauser, F.; Hopp, S.; Mencl, S.; Geuss, E.; Leinweber, J.; Guthmann, J.; et al. Validity and reliability of neurological scores in mice exposed to middle cerebral artery occlusion. Stroke 2019, 50, 2875–2882. [Google Scholar] [CrossRef]
- Choi, M.; Lee, Y.; Cho, S.-H. Angelica tenuissima Nakai ameliorates cognitive impairment and promotes neurogenesis in mouse model of Alzheimer’s disease. Chin. J. Integr. Med. 2018, 24, 378–384. [Google Scholar] [CrossRef]
- Park, J.-H.; Hong, J.-H.; Lee, S.-W.; Ji, H.D.; Jung, J.-A.; Yoon, K.-W.; Lee, J.-I.; Won, K.S.; Song, B.-I.; Kim, H.W. The effect of chronic cerebral hypoperfusion on the pathology of Alzheimer’s disease: A positron emission tomography study in rats. Sci. Rep. 2019, 9, 14102. [Google Scholar] [CrossRef]
- Casteels, C.; Vunckx, K.; Aelvoet, S.-A.; Baekelandt, V.; Bormans, G.; Van Laere, K.; Koole, M. Construction and evaluation of quantitative small-animal PET probabilistic atlases for [18F] FDG and [18F] FECT functional mapping of the mouse brain. PLoS ONE 2013, 8, e65286. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Bang, Y.; Kim, K.M.; Choi, H.J. Differentiated HT22 cells as a novel model for screening of serotonin reuptake inhibitors. Front. Pharmacol. 2023, 13, 1062650. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Zhang, Y.T.; Jiang, S.N.; Xie, N.; Zhang, Y.; Meng, X.L.; Wang, X.B. Salidroside intensifies mitochondrial function of CoCl-damaged HT22 cells by stimulating PI3K-AKT-MAPK signaling pathway. Phytomedicine 2023, 109, 154568. [Google Scholar] [CrossRef] [PubMed]
- Thiel, A.; Cechetto, D.F.; Heiss, W.-D.; Hachinski, V.; Whitehead, S.N. Amyloid burden, neuroinflammation, and links to cognitive decline after ischemic stroke. Stroke 2014, 45, 2825–2829. [Google Scholar] [CrossRef] [PubMed]
- Rost, N.S.; Brodtmann, A.; Pase, M.P.; van Veluw, S.J.; Biffi, A.; Duering, M.; Hinman, J.D.; Dichgans, M. Post-stroke cognitive impairment and dementia. Circ. Res. 2022, 130, 1252–1271. [Google Scholar] [CrossRef]
- Dichgans, M.; Leys, D. Vascular cognitive impairment. Circ. Res. 2017, 120, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting oxidative stress and inflammation to prevent ischemia-reperfusion injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef]
- Sarmah, D.; Kaur, H.; Saraf, J.; Vats, K.; Pravalika, K.; Wanve, M.; Kalia, K.; Borah, A.; Kumar, A.; Wang, X.; et al. Mitochondrial dysfunction in stroke: Implications of stem cell therapy. Transl. Stroke Res. 2018, 10, 121–136. [Google Scholar] [CrossRef]
- Sekerdag, E.; Solaroglu, I.; Gursoy-Ozdemir, Y. Cell death mechanisms in stroke and novel molecular and cellular treatment options. Curr. Neuropharmacol. 2018, 16, 1396–1415. [Google Scholar] [CrossRef]
- Fricker, M.; Tolkovsky, A.M.; Borutaite, V.; Coleman, M.; Brown, G.C. Neuronal cell death. Physiol. Rev. 2018, 98, 813–880. [Google Scholar] [CrossRef]
- Datta, A.; Sarmah, D.; Mounica, L.; Kaur, H.; Kesharwani, R.; Verma, G.; Veeresh, P.; Kotian, V.; Kalia, K.; Borah, A.; et al. Cell death pathways in ischemic stroke and targeted pharmacotherapy. Transl. Stroke Res. 2020, 11, 1185–1202. [Google Scholar] [CrossRef]
- Fisher, M.; Feuerstein, G.; Howells, D.W.; Hurn, P.D.; Kent, T.A.; Savitz, S.I.; Lo, E.H. Update of the stroke therapy academic industry roundtable preclinical recommendations. Stroke 2009, 40, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Kallmünzer, B.; Kollmar, R. Temperature management in stroke–an unsolved, but important topic. Cerebrovasc. Dis. 2011, 31, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Faridar, A.; Bershad, E.M.; Emiru, T.; Iaizzo, P.A.; Suarez, J.I.; Divani, A.A. Therapeutic hypothermia in stroke and traumatic brain injury. Front. Neurol. 2011, 2, 80. [Google Scholar] [CrossRef] [PubMed]
- Tveita, T.; Sieck, G.C. Physiological impact of hypothermia: The good, the bad, and the ugly. Physiology 2022, 37, 69–87. [Google Scholar] [CrossRef]
- Tahir, R.A.; Pabaney, A.H. Therapeutic hypothermia and ischemic stroke: A literature review. Surg. Neurol. Int. 2016, 7, S381. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Huang, Z.-G.; Smith, K.E.; Buchan, A.M. Immediate or delayed mild hypothermia prevents focal cerebral infarction. Brain Res. 1992, 587, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Barber, P.A.; Hoyte, L.; Colbourne, F.; Buchan, A.M. Temperature-regulated model of focal ischemia in the mouse: A study with histopathological and behavioral outcomes. Stroke 2004, 35, 1720–1725. [Google Scholar] [CrossRef]
- Yenari, M.A.; Hemmen, T.M. Therapeutic hypothermia for brain ischemia: Where have we come and where do we go? Stroke 2010, 41, S72–S74. [Google Scholar] [CrossRef]
- Wu, D.; Chen, J.; Zhang, X.; Ilagan, R.; Ding, Y.; Ji, X. Selective therapeutic cooling: To maximize benefits and minimize side effects related to hypothermia. J. Cereb. Blood Flow Metab. 2022, 42, 213–215. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Zhong, H.; Li, X.; Wu, Y.; Ju, J.; Liu, B.; Zhang, Z.; Yan, H.; Wang, Y. Hypothermia evoked by stimulation of medial preoptic nucleus protects the brain in a mouse model of ischaemia. Nat. Commun. 2022, 13, 6890. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Halliwell, B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat. Rev. Neurosci. 2019, 20, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Inui, Y.; Nakamura, A.; Ito, K. Brain fluorodeoxyglucose (FDG) PET in dementia. Ageing Res. Rev. 2016, 30, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kellar, D.; Craft, S. Brain insulin resistance in Alzheimer’s disease and related disorders: Mechanisms and therapeutic approaches. Lancet Neurol. 2020, 19, 758–766. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Varma, V.R.; Varma, S.; Casanova, R.; Dammer, E.; Pletnikova, O.; Chia, C.W.; Egan, J.M.; Ferrucci, L.; Troncoso, J. Evidence for brain glucose dysregulation in Alzheimer’s disease. Alzheimers Dement. 2018, 14, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Weaver, N.A.; Kuijf, H.J.; Aben, H.P.; Abrigo, J.; Bae, H.-J.; Barbay, M.; Best, J.G.; Bordet, R.; Chappell, F.M.; Chen, C.P. Strategic infarct locations for post-stroke cognitive impairment: A pooled analysis of individual patient data from 12 acute ischaemic stroke cohorts. Lancet Neurol. 2021, 20, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Biesbroek, J.M.; Shi, L.; Liu, W.; Kuijf, H.J.; Chu, W.W.; Abrigo, J.M.; Lee, R.K.; Leung, T.W.; Lau, A.Y. Strategic infarct location for post-stroke cognitive impairment: A multivariate lesion-symptom mapping study. J. Cereb. Blood Flow Metab. 2018, 38, 1299–1311. [Google Scholar] [CrossRef]
- Karussis, D.; Leker, R.; Abramsky, O. Cognitive dysfunction following thalamic stroke: A study of 16 cases and review of the literature. J. Neurol. Sci. 2000, 172, 25–29. [Google Scholar] [CrossRef]
- Obayashi, S. Cognitive and linguistic dysfunction after thalamic stroke and recovery process: Possible mechanism. AIMS Neurosci. 2022, 9, 1. [Google Scholar] [CrossRef]
- Borgstedt, L.; Bratke, S.; Blobner, M.; Pötzl, C.; Ulm, B.; Jungwirth, B.; Schmid, S. Isoflurane has no effect on cognitive or behavioral performance in a mouse model of early-stage Alzheimer’s disease. Front. Neurosci. 2022, 16, 1033729. [Google Scholar] [CrossRef]
- Lin, D.; Zuo, Z. Isoflurane induces hippocampal cell injury and cognitive impairments in adult rats. Neuropharmacology 2011, 61, 1354–1359. [Google Scholar] [CrossRef]
- Pathak, D.; Sriram, K. Molecular mechanisms underlying neuroinflammation elicited by occupational injuries and toxicants. Int. J. Mol. Sci. 2023, 24, 2272. [Google Scholar] [CrossRef]
- Rakers, C.; Schleif, M.; Blank, N.; Matusková, H.; Ulas, T.; Händler, K.; Torres, S.V.; Schumacher, T.; Tai, K.; Schultze, J.L.; et al. Stroke target identification guided by astrocyte transcriptome analysis. Glia 2019, 67, 619–633. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | Side | Mean (Standard Deviation) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Sham | tMCAO/R | tMCAO/R + TH | Sham vs. tMCAO/R | Sham vs. tMCAO/R + TH | tMCAO vs. tMCAO/R + TH | ||
| Amygdala | left | 0.828 (0.081) | 0.904 (0.064) | 0.890 (0.066) | 0.115 | 0.213 | 1.000 |
| right | 0.883 (0.107) | 0.856 (0.087) | 0.904 (0.088) | 1.000 | 1.000 | 0.883 | |
| Cortex | left | 0.960 (0.036) | 0.970 (0.043) | 0.943 (0.035) | 1.000 | 1.000 | 0.438 |
| right | 0.998 (0.085) | 0.948 (0.107) | 0.942 (0.094) | 0.867 | 0.624 | 1.000 | |
| Hippocampus | left | 1.141 (0.070) | 1.062 (0.079) | 1.151 (0.079) | 0.134 | 1.000 | 0.065 |
| right | 1.187 (0.109) | 1.062 (0.078) | 1.162 (0.102) | 0.044 * | 1.000 | 0.127 | |
| Midbrain | left | 1.280 (0.131) | 1.175 (0.086) | 1.260 (0.108) | 0.193 | 1.000 | 0.362 |
| right | 1.282 (0.152) | 1.143 (0.081) | 1.243 (0.135) | 0.106 | 1.000 | 0.345 | |
| Striatum | left | 1.165 (0.078) | 1.082 (0.031) | 1.133 (0.069) | 0.040 * | 0.887 | 0.307 |
| right | 1.191 (0.116) | 1.074 (0.052) | 1.145 (0.091) | 0.043 * | 0.845 | 0.345 | |
| Thalamus | left | 1.224 (0.086) | 1.102 (0.049) | 1.213 (0.079) | 0.007 ** | 1.000 | 0.012 * |
| right | 1.280 (0.136) | 1.135 (0.067) | 1.248 (0.113) | 0.038 * | 1.000 | 0.127 | |
| Inferior colliculi | left | 1.336 (0.165) | 1.205 (0.074) | 0.265 (0.084) | 0.083 | 0.570 | 0.850 |
| right | 1.390 (0.209) | 1.222 (0.083) | 1.331 (0.124) | 0.089 | 1.000 | 0.412 | |
| Superior colliculi | 1.318 (0.152) | 1.182 (0.053) | 1.261 (0.099) | 0.056 | 0.827 | 0.438 | |
| Basal forebrain | 1.046 (0.041) | 1.031 (0.037) | 1.006 (0.042) | 1.000 | 0.126 | 0.602 | |
| Central gray | 1.434 (0.177) | 1.237 (0.079) | 0.328 (0.138) | 0.023 * | 0.325 | 0.551 | |
| Hypothalamus | 1.078 (0.086) | 1.043 (0.058) | 1.083 (0.053) | 0.899 | 1.000 | 0.675 | |
| Olfactory bulb | 1.377 (0.135) | 1.321 (0.173) | 1.248 (0.142) | 1.000 | 0.220 | 0.944 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.S.; Kim, S.; Park, M.-N.; Lee, H.; Baek, H.S.; Kim, J.K.; Kim, H.W.; Hong, J.-H. The Neuroprotective Effect of Therapeutic Hypothermia in Cognitive Impairment of an Ischemia/Reperfusion Injury Mouse Model. Medicina 2024, 60, 350. https://doi.org/10.3390/medicina60030350
Lim JS, Kim S, Park M-N, Lee H, Baek HS, Kim JK, Kim HW, Hong J-H. The Neuroprotective Effect of Therapeutic Hypothermia in Cognitive Impairment of an Ischemia/Reperfusion Injury Mouse Model. Medicina. 2024; 60(3):350. https://doi.org/10.3390/medicina60030350
Chicago/Turabian StyleLim, Ji Sun, Shin Kim, Mee-Na Park, Hyunsu Lee, Hye Suk Baek, Jin Kyung Kim, Hae Won Kim, and Jeong-Ho Hong. 2024. "The Neuroprotective Effect of Therapeutic Hypothermia in Cognitive Impairment of an Ischemia/Reperfusion Injury Mouse Model" Medicina 60, no. 3: 350. https://doi.org/10.3390/medicina60030350
APA StyleLim, J. S., Kim, S., Park, M.-N., Lee, H., Baek, H. S., Kim, J. K., Kim, H. W., & Hong, J.-H. (2024). The Neuroprotective Effect of Therapeutic Hypothermia in Cognitive Impairment of an Ischemia/Reperfusion Injury Mouse Model. Medicina, 60(3), 350. https://doi.org/10.3390/medicina60030350