


Therapeutic Potential of Cannabinoids in Glaucoma
 , ,
, , 
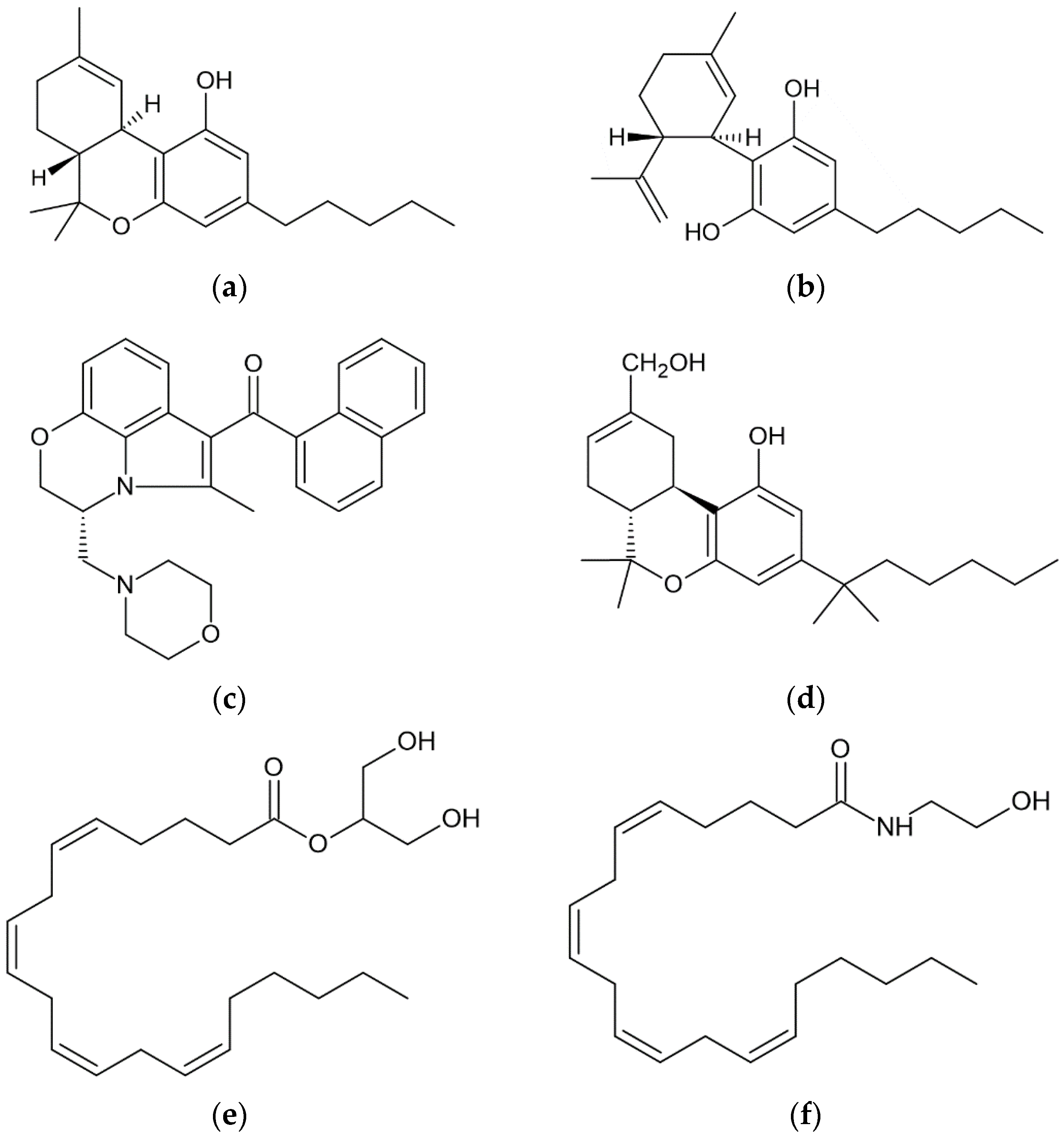{kind=link}
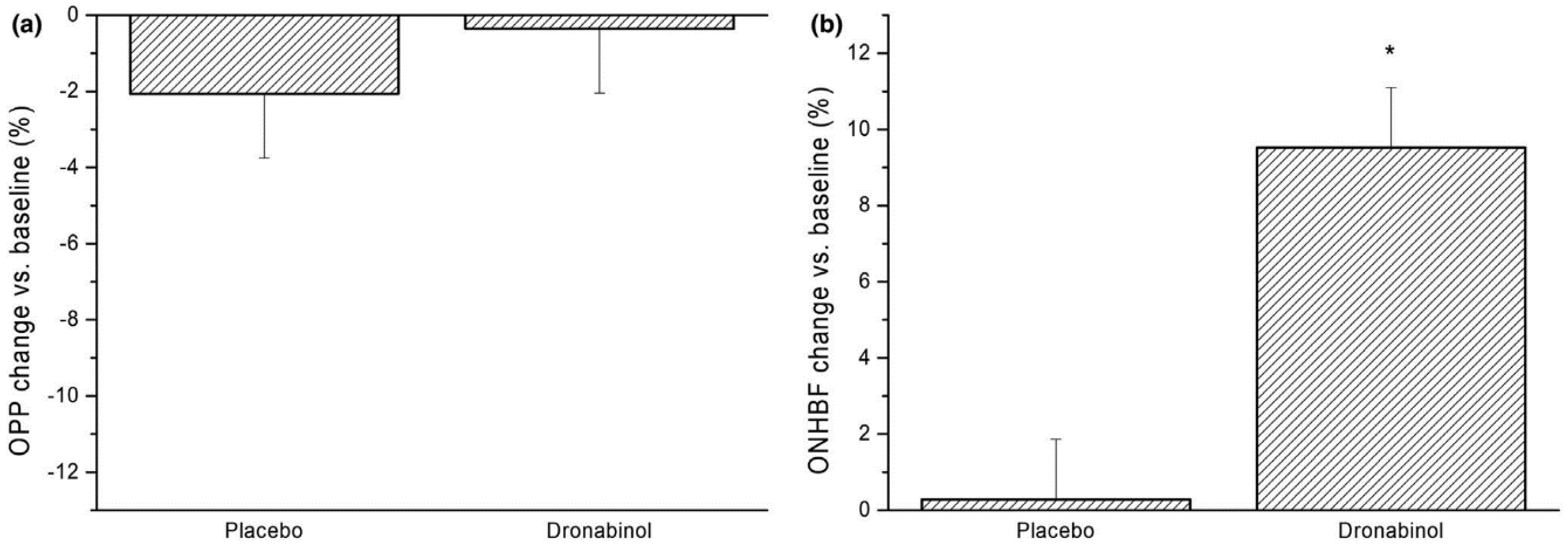{kind=link}
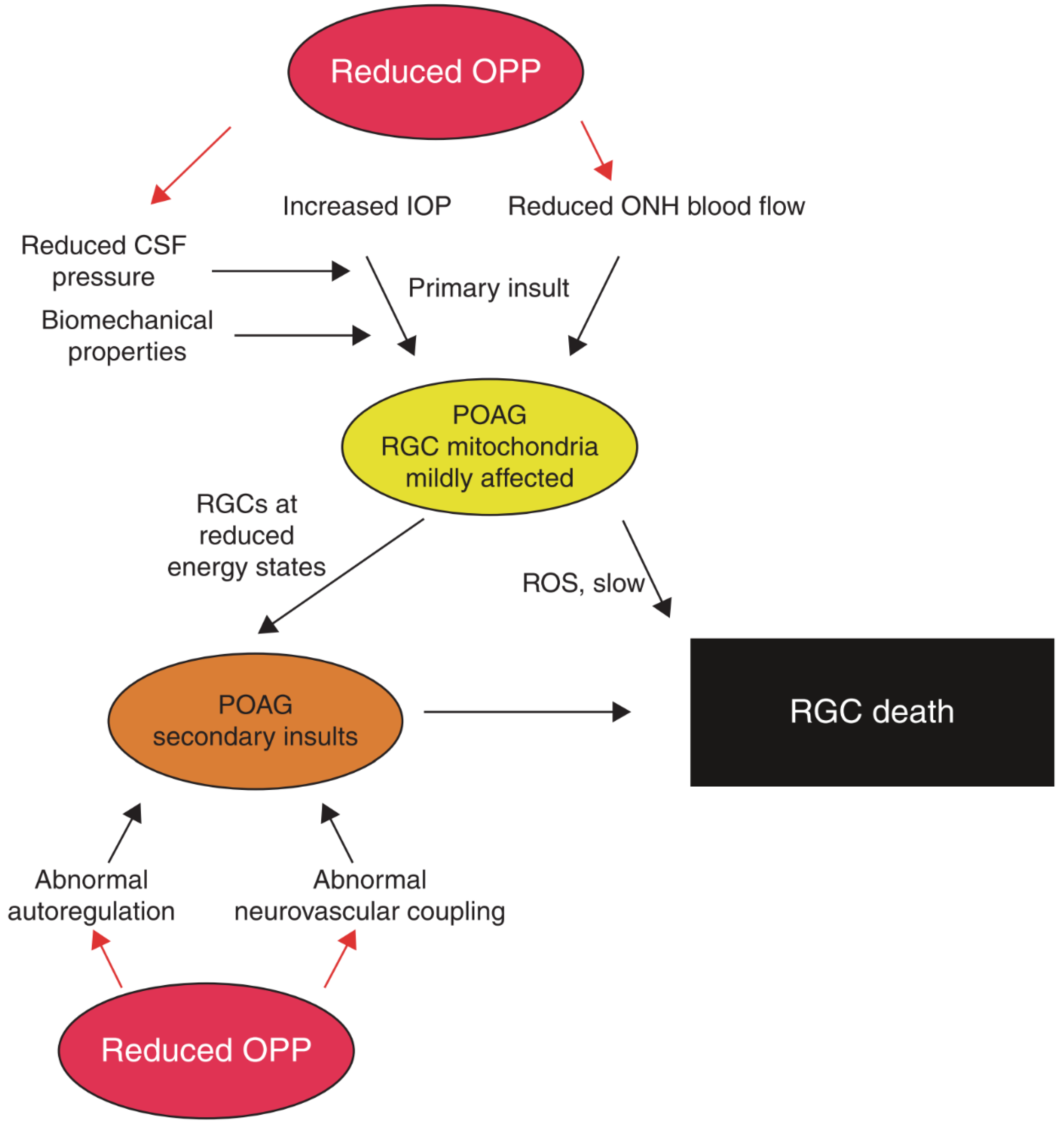{kind=link}
Abstract
1. Introduction
2. Glaucoma
3. Marijuana and Endocannabinoids
3.1. Phytocannabinoids
3.2. Synthetic Cannabinoids
3.3. Endocannabinoids
4. Molecular Targets and Mechanisms for Cannabinoid-Induced Ocular Effects
4.1. CB1 and CB2 Receptors
4.2. Non-Cannabinoid Receptors
5. Therapeutic Approaches of Cannabinoids on IOP
5.1. Clinical Studies on the IOP-Lowering Effect of Cannabinoids
5.2. Side Effects
5.3. Novel Topical Formulations
6. Neuroprotective Actions of Cannabinoids
6.1. Targeting Mechanisms in Neuroprotection
6.2. Evidence for Neuroprotection
7. Vascular Targets of Cannabinoids and the Effect on Ocular Hemodynamics
7.1. Vascular Targets
7.2. Evidence for Hemodynamic Effects
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schilling, S.; Melzer, R.; McCabe, P.F. Cannabis sativa. Curr. Biol. 2020, 30, R8–R9. [Google Scholar] [CrossRef]
- Crocq, M.A. History of cannabis and the endocannabinoid system. Dialogues Clin. Neurosci. 2020, 22, 223–228. [Google Scholar] [CrossRef]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef]
- MacDonald, E.; Adams, A. CADTH Rapid Response Reports. In The Use of Medical Cannabis with Other Medications: A Review of Safety and Guidelines—An Update; Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2019. [Google Scholar]
- Stasiłowicz, A.; Tomala, A.; Podolak, I.; Cielecka-Piontek, J. Cannabis sativa L. as a Natural Drug Meeting the Criteria of a Multitarget Approach to Treatment. Int. J. Mol. Sci. 2021, 22, 778. [Google Scholar] [CrossRef] [PubMed]
- Hepler, R.S.; Frank, I.R. Marihuana smoking and intraocular pressure. JAMA 1971, 217, 1392. [Google Scholar] [CrossRef]
- Tomida, I.; Pertwee, R.G.; Azuara-Blanco, A. Cannabinoids and glaucoma. Br. J. Ophthalmol. 2004, 88, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Passani, A.; Posarelli, C.; Sframeli, A.T.; Perciballi, L.; Pellegrini, M.; Guidi, G.; Figus, M. Cannabinoids in Glaucoma Patients: The Never-Ending Story. J. Clin. Med. 2020, 9, 3978. [Google Scholar] [CrossRef] [PubMed]
- Rapino, C.; Tortolani, D.; Scipioni, L.; Maccarrone, M. Neuroprotection by (endo)Cannabinoids in Glaucoma and Retinal Neurodegenerative Diseases. Curr. Neuropharmacol. 2018, 16, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.T.M.; Danesh-Meyer, H.V. Cannabinoids and the eye. Surv. Ophthalmol. 2021, 66, 327–345. [Google Scholar] [CrossRef]
- Nucci, C.; Bari, M.; Spanò, A.; Corasaniti, M.; Bagetta, G.; Maccarrone, M.; Morrone, L.A. Potential roles of (endo)cannabinoids in the treatment of glaucoma: From intraocular pressure control to neuroprotection. Prog. Brain Res. 2008, 173, 451–464. [Google Scholar] [CrossRef]
- Cairns, E.A.; Baldridge, W.H.; Kelly, M.E. The Endocannabinoid System as a Therapeutic Target in Glaucoma. Neural Plast. 2016, 2016, 9364091. [Google Scholar] [CrossRef] [PubMed]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Cherecheanu, A.P.; Garhofer, G.; Schmidl, D.; Werkmeister, R.; Schmetterer, L. Ocular perfusion pressure and ocular blood flow in glaucoma. Curr. Opin. Pharmacol. 2013, 13, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Pezzullo, L.; Streatfeild, J.; Simkiss, P.; Shickle, D. The economic impact of sight loss and blindness in the UK adult population. BMC Health Serv. Res. 2018, 18, 63. [Google Scholar] [CrossRef]
- Coleman, A.L.; Miglior, S. Risk factors for glaucoma onset and progression. Surv. Ophthalmol. 2008, 53 (Suppl. S1), S3–S10. [Google Scholar] [CrossRef]
- Schmidl, D.; Schmetterer, L.; Garhöfer, G.; Popa-Cherecheanu, A. Pharmacotherapy of glaucoma. J. Ocul. Pharmacol. Ther. 2015, 31, 63–77. [Google Scholar] [CrossRef]
- Lu, L.J.; Tsai, J.C.; Liu, J. Novel Pharmacologic Candidates for Treatment of Primary Open-Angle Glaucoma. Yale J. Biol. Med. 2017, 90, 111–118. [Google Scholar]
- Shalaby, W.S.; Shankar, V.; Razeghinejad, R.; Katz, L.J. Current and new pharmacotherapeutic approaches for glaucoma. Expert Opin. Pharmacother. 2020, 21, 2027–2040. [Google Scholar] [CrossRef]
- Wang, S.Y.; Singh, K. Management of the glaucoma patient progressing at low normal intraocular pressure. Curr. Opin. Ophthalmol. 2020, 31, 107–113. [Google Scholar] [CrossRef]
- Bengtsson, B. The prevalence of glaucoma. Br. J. Ophthalmol. 1981, 65, 46–49. [Google Scholar] [CrossRef]
- Yorio, T.; Krishnamoorthy, R.; Prasanna, G. Endothelin: Is it a contributor to glaucoma pathophysiology? J. Glaucoma 2002, 11, 259–270. [Google Scholar] [CrossRef]
- Guo, Y.; Shrestha, M.; Marshak-Rothstein, A.; Gregory-Ksander, M.S. Caspase-8-mediated inflammation but not apoptosis drives the development of glaucoma. Investig. Ophthalmol. Vis. Sci. 2021, 62, 2350. [Google Scholar]
- Evangelho, K.; Mogilevskaya, M.; Losada-Barragan, M.; Vargas-Sanchez, J.K. Pathophysiology of primary open-angle glaucoma from a neuroinflammatory and neurotoxicity perspective: A review of the literature. Int. Ophthalmol. 2019, 39, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Adornetto, A.; Rombolà, L.; Morrone, L.A.; Nucci, C.; Corasaniti, M.T.; Bagetta, G.; Russo, R. Natural Products: Evidence for Neuroprotection to Be Exploited in Glaucoma. Nutrients 2020, 12, 3158. [Google Scholar] [CrossRef] [PubMed]
- Sim, R.H.; Sirasanagandla, S.R.; Das, S.; Teoh, S.L. Treatment of Glaucoma with Natural Products and Their Mechanism of Action: An Update. Nutrients 2022, 14, 534. [Google Scholar] [CrossRef]
- Alves, V.L.; Gonçalves, J.L.; Aguiar, J.; Teixeira, H.M.; Câmara, J.S. The synthetic cannabinoids phenomenon: From structure to toxicological properties. A review. Crit. Rev. Toxicol. 2020, 50, 359–382. [Google Scholar] [CrossRef]
- Turner, S.E.; Williams, C.M.; Iversen, L.; Whalley, B.J. Molecular Pharmacology of Phytocannabinoids. Prog. Chem. Org. Nat. Prod. 2017, 103, 61–101. [Google Scholar] [CrossRef]
- An, D.; Peigneur, S.; Hendrickx, L.A.; Tytgat, J. Targeting Cannabinoid Receptors: Current Status and Prospects of Natural Products. Int. J. Mol. Sci. 2020, 21, 5064. [Google Scholar] [CrossRef]
- Alves, P.; Amaral, C.; Teixeira, N.; Correia-da-Silva, G. Cannabis sativa: Much more beyond Δ9-tetrahydrocannabinol. Pharmacol. Res. 2020, 157, 104822. [Google Scholar] [CrossRef]
- Metna-Laurent, M.; Mondésir, M.; Grel, A.; Vallée, M.; Piazza, P.V. Cannabinoid-Induced Tetrad in Mice. Curr. Protoc. Neurosci. 2017, 80, 9–59. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef]
- Martínez-Pinilla, E.; Varani, K.; Reyes-Resina, I.; Angelats, E.; Vincenzi, F.; Ferreiro-Vera, C.; Oyarzabal, J.; Canela, E.I.; Lanciego, J.L.; Nadal, X.; et al. Binding and Signaling Studies Disclose a Potential Allosteric Site for Cannabidiol in Cannabinoid CB2 Receptors. Front. Pharmacol. 2017, 8, 744. [Google Scholar] [CrossRef]
- Roque-Bravo, R.; Silva, R.S.; Malheiro, R.F.; Carmo, H.; Carvalho, F.; da Silva, D.D.; Silva, J.P. Synthetic Cannabinoids: A Pharmacological and Toxicological Overview. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 187–209. [Google Scholar] [CrossRef]
- U.S. Food & Drug Administration. FDA and Cannabis: Research and Drug Approval Process. Available online: https://www.fda.gov/news-events/public-health-focus/fda-and-cannabis-research-and-drug-approval-process (accessed on 29 June 2023).
- Accelerated Cure Project for Multiple Sclerosis. Sativex—The Road to FDA Approval. Available online: https://www.acceleratedcure.org/sites/default/files/images/Sativex-TheRoadtoFDAApproval.pdf (accessed on 29 June 2023).
- Yazulla, S. Endocannabinoids in the retina: From marijuana to neuroprotection. Prog. Retin. Eye. Res. 2008, 27, 501–526. [Google Scholar] [CrossRef] [PubMed]
- Aiello, F.; Gallo Afflitto, G.; Li, J.O.; Martucci, A.; Cesareo, M.; Nucci, C. CannabinEYEds: The Endocannabinoid System as a Regulator of the Ocular Surface Nociception, Inflammatory Response, Neovascularization and Wound Healing. J. Clin. Med. 2020, 9, 4036. [Google Scholar] [CrossRef] [PubMed]
- Schwitzer, T.; Schwan, R.; Angioi-Duprez, K.; Giersch, A.; Laprevote, V. The Endocannabinoid System in the Retina: From Physiology to Practical and Therapeutic Applications. Neural Plast. 2016, 2016, 2916732. [Google Scholar] [CrossRef]
- Cairns, E.A.; Toguri, J.T.; Porter, R.F.; Szczesniak, A.M.; Kelly, M.E. Seeing over the horizon-targeting the endocannabinoid system for the treatment of ocular disease. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Mackie, K. Review of the Endocannabinoid System. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 607–615. [Google Scholar] [CrossRef]
- Di Marzo, V.; Piscitelli, F. The Endocannabinoid System and its Modulation by Phytocannabinoids. Neurotherapeutics 2015, 12, 692–698. [Google Scholar] [CrossRef]
- Chen, J.; Matias, I.; Dinh, T.; Lu, T.; Venezia, S.; Nieves, A.; Woodward, D.F.; Di Marzo, V. Finding of endocannabinoids in human eye tissues: Implications for glaucoma. Biochem. Biophys. Res. Commun. 2005, 330, 1062–1067. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Cannabinoid Receptors: An Update on Cell Signaling, Pathophysiological Roles and Therapeutic Opportunities in Neurological, Cardiovascular, and Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 7693. [Google Scholar] [CrossRef] [PubMed]
- Alexander, S.P.; Kendall, D.A. The complications of promiscuity: Endocannabinoid action and metabolism. Br. J. Pharmacol. 2007, 152, 602–623. [Google Scholar] [CrossRef] [PubMed]
- Rösch, S.; Ramer, R.; Brune, K.; Hinz, B. R(+)-methanandamide and other cannabinoids induce the expression of cyclooxygenase-2 and matrix metalloproteinases in human nonpigmented ciliary epithelial cells. J. Pharmacol. Exp. Ther. 2006, 316, 1219–1228. [Google Scholar] [CrossRef]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Pertwee, R.G. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol. Ther. 1997, 74, 129–180. [Google Scholar] [CrossRef]
- Howlett, A.C. Cannabinoid receptor signaling. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 53–79. [Google Scholar] [CrossRef]
- Szabo, B.; Schlicker, E. Effects of cannabinoids on neurotransmission. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 327–365. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Straiker, A.J.; Maguire, G.; Mackie, K.; Lindsey, J. Localization of cannabinoid CB1 receptors in the human anterior eye and retina. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2442–2448. [Google Scholar]
- Somvanshi, R.K.; Zou, S.; Kadhim, S.; Padania, S.; Hsu, E.; Kumar, U. Cannabinol modulates neuroprotection and intraocular pressure: A potential multi-target therapeutic intervention for glaucoma. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166325. [Google Scholar] [CrossRef]
- Järvinen, T.; Pate, D.W.; Laine, K. Cannabinoids in the treatment of glaucoma. Pharmacol. Ther. 2002, 95, 203–220. [Google Scholar] [CrossRef]
- Murataeva, N.; Miller, S.; Dhopeshwarkar, A.; Leishman, E.; Daily, L.; Taylor, X.; Morton, B.; Lashmet, M.; Bradshaw, H.; Hillard, C.J.; et al. Cannabinoid CB2R receptors are upregulated with corneal injury and regulate the course of corneal wound healing. Exp. Eye Res. 2019, 182, 74–84. [Google Scholar] [CrossRef]
- Zhong, L.; Geng, L.; Njie, Y.; Feng, W.; Song, Z.H. CB2 cannabinoid receptors in trabecular meshwork cells mediate JWH015-induced enhancement of aqueous humor outflow facility. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1988–1992. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, X.; Wang, L. Presence and regulation of cannabinoid receptors in human retinal pigment epithelial cells. Mol. Vis. 2009, 15, 1243–1251. [Google Scholar]
- Gallo Afflitto, G.; Aiello, F.; Scuteri, D.; Bagetta, G.; Nucci, C. CB1R, CB2R and TRPV1 expression and modulation in in vivo, animal glaucoma models: A systematic review. Biomed. Pharmacother. 2022, 150, 112981. [Google Scholar] [CrossRef]
- Morales, P.; Jagerovic, N. Advances Towards The Discovery of GPR55 Ligands. Curr. Med. Chem. 2016, 23, 2087–2100. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, J.; Lehmann, C. GPR55—A putative “type 3” cannabinoid receptor in inflammation. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 297–302. [Google Scholar] [CrossRef]
- Kumar, A.; Qiao, Z.; Kumar, P.; Song, Z.H. Effects of palmitoylethanolamide on aqueous humor outflow. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4416–4425. [Google Scholar] [CrossRef]
- Bouskila, J.; Javadi, P.; Casanova, C.; Ptito, M.; Bouchard, J.F. Rod photoreceptors express GPR55 in the adult vervet monkey retina. PLoS ONE 2013, 8, e81080. [Google Scholar] [CrossRef]
- Balenga, N.A.; Aflaki, E.; Kargl, J.; Platzer, W.; Schröder, R.; Blättermann, S.; Kostenis, E.; Brown, A.J.; Heinemann, A.; Waldhoer, M. GPR55 regulates cannabinoid 2 receptor-mediated responses in human neutrophils. Cell Res. 2011, 21, 1452–1469. [Google Scholar] [CrossRef]
- Kohno, M.; Hasegawa, H.; Inoue, A.; Muraoka, M.; Miyazaki, T.; Oka, K.; Yasukawa, M. Identification of N-arachidonylglycine as the endogenous ligand for orphan G-protein-coupled receptor GPR18. Biochem. Biophys. Res. Commun. 2006, 347, 827–832. [Google Scholar] [CrossRef]
- Miller, S.; Daily, L.; Leishman, E.; Bradshaw, H.; Straiker, A. Δ9-Tetrahydrocannabinol and Cannabidiol Differentially Regulate Intraocular Pressure. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5904–5911. [Google Scholar] [CrossRef]
- Caldwell, M.D.; Hu, S.S.; Viswanathan, S.; Bradshaw, H.; Kelly, M.E.; Straiker, A. A GPR18-based signalling system regulates IOP in murine eye. Br. J. Pharmacol. 2013, 169, 834–843. [Google Scholar] [CrossRef]
- MacIntyre, J.; Dong, A.; Straiker, A.; Zhu, J.; Howlett, S.E.; Bagher, A.; Denovan-Wright, E.; Yu, D.Y.; Kelly, M.E. Cannabinoid and lipid-mediated vasorelaxation in retinal microvasculature. Eur. J. Pharmacol. 2014, 735, 105–114. [Google Scholar] [CrossRef]
- Miller, S.; Hu, S.S.; Leishman, E.; Morgan, D.; Wager-Miller, J.; Mackie, K.; Bradshaw, H.B.; Straiker, A. A GPR119 Signaling System in the Murine Eye Regulates Intraocular Pressure in a Sex-Dependent Manner. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2930–2938. [Google Scholar] [CrossRef]
- Montell, C. The TRP superfamily of cation channels. Sci. STKE 2005, 2005, re3. [Google Scholar] [CrossRef]
- Yang, T.J.; Yu, Y.; Yang, J.Y.; Li, J.J.; Zhu, J.Y.; Vieira, J.A.C.; Jiang, Q. Involvement of transient receptor potential channels in ocular diseases: A narrative review. Ann. Transl. Med. 2022, 10, 839. [Google Scholar] [CrossRef]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 2018, 11, 487. [Google Scholar] [CrossRef]
- Sappington, R.M.; Sidorova, T.; Long, D.J.; Calkins, D.J. TRPV1: Contribution to retinal ganglion cell apoptosis and increased intracellular Ca2+ with exposure to hydrostatic pressure. Investig. Ophthalmol. Vis. Sci. 2009, 50, 717–728. [Google Scholar] [CrossRef]
- Luo, N.; Conwell, M.D.; Chen, X.; Kettenhofen, C.I.; Westlake, C.J.; Cantor, L.B.; Wells, C.D.; Weinreb, R.N.; Corson, T.W.; Spandau, D.F.; et al. Primary cilia signaling mediates intraocular pressure sensation. Proc. Natl. Acad. Sci. USA 2014, 111, 12871–12876. [Google Scholar] [CrossRef]
- Patel, P.D.; Chen, Y.L.; Kasetti, R.B.; Maddineni, P.; Mayhew, W.; Millar, J.C.; Ellis, D.Z.; Sonkusare, S.K.; Zode, G.S. Impaired TRPV4-eNOS signaling in trabecular meshwork elevates intraocular pressure in glaucoma. Proc. Natl. Acad. Sci. USA 2021, 118, e2022461118. [Google Scholar] [CrossRef]
- Li, Q.; Cheng, Y.; Zhang, S.; Sun, X.; Wu, J. TRPV4-induced Müller cell gliosis and TNF-α elevation-mediated retinal ganglion cell apoptosis in glaucomatous rats via JAK2/STAT3/NF-κB pathway. J. Neuroinflamm. 2021, 18, 271. [Google Scholar] [CrossRef]
- Taylor, L.; Arnér, K.; Ghosh, F. Specific inhibition of TRPV4 enhances retinal ganglion cell survival in adult porcine retinal explants. Exp. Eye Res. 2017, 154, 10–21. [Google Scholar] [CrossRef]
- Choi, H.J.; Sun, D.; Jakobs, T.C. Astrocytes in the optic nerve head express putative mechanosensitive channels. Mol. Vis. 2015, 21, 749–766. [Google Scholar]
- Ryskamp, D.A.; Witkovsky, P.; Barabas, P.; Huang, W.; Koehler, C.; Akimov, N.P.; Lee, S.H.; Chauhan, S.; Xing, W.; Rentería, R.C.; et al. The polymodal ion channel transient receptor potential vanilloid 4 modulates calcium flux, spiking rate, and apoptosis of mouse retinal ganglion cells. J. Neurosci. 2011, 31, 7089–7101. [Google Scholar] [CrossRef]
- Burstein, S. PPAR-gamma: A nuclear receptor with affinity for cannabinoids. Life Sci. 2005, 77, 1674–1684. [Google Scholar] [CrossRef]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef]
- Aebersold, A.S.; Song, Z.H. The Effects of Cannabidiol on Aqueous Humor Outflow and Trabecular Meshwork Cell Signaling. Cells 2022, 11, 3006. [Google Scholar] [CrossRef]
- Thapa, D.; Cairns, E.A.; Szczesniak, A.M.; Toguri, J.T.; Caldwell, M.D.; Kelly, M.E.M. The Cannabinoids Δ8THC, CBD, and HU-308 Act via Distinct Receptors to Reduce Corneal Pain and Inflammation. Cannabis Cannabinoid Res. 2018, 3, 11–20. [Google Scholar] [CrossRef]
- Liou, G.I.; Auchampach, J.A.; Hillard, C.J.; Zhu, G.; Yousufzai, B.; Mian, S.; Khan, S.; Khalifa, Y. Mediation of cannabidiol anti-inflammation in the retina by equilibrative nucleoside transporter and A2A adenosine receptor. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5526–5531. [Google Scholar] [CrossRef]
- Rocha-Sousa, A.; Rodrigues-Araújo, J.; Gouveia, P.; Barbosa-Breda, J.; Azevedo-Pinto, S.; Pereira-Silva, P.; Leite-Moreira, A. New therapeutic targets for intraocular pressure lowering. ISRN Ophthalmol. 2013, 2013, 261386. [Google Scholar] [CrossRef]
- Laine, K.; Järvinen, K.; Järvinen, T. Topically administered CB2-receptor agonist, JWH-133, does not decrease intraocular pressure (IOP) in normotensive rabbits. Life Sci. 2003, 72, 837–842. [Google Scholar] [CrossRef]
- Woodward, D.F.; Carling, R.W.; Cornell, C.L.; Fliri, H.G.; Martos, J.L.; Pettit, S.N.; Liang, Y.; Wang, J.W. The pharmacology and therapeutic relevance of endocannabinoid derived cyclo-oxygenase (COX)-2 products. Pharmacol. Ther. 2008, 120, 71–80. [Google Scholar] [CrossRef]
- Smid, S.D. Role of prostaglandins and specific place in therapy of bimatoprost in the treatment of elevated intraocular pressure and ocular hypertension: A closer look at the agonist properties of bimatoprost and the prostamides. Clin. Ophthalmol. 2009, 3, 663–670. [Google Scholar] [CrossRef][Green Version]
- Maihöfner, C.; Schlötzer-Schrehardt, U.; Gühring, H.; Zeilhofer, H.U.; Naumann, G.O.; Pahl, A.; Mardin, C.; Tamm, E.R.; Brune, K. Expression of cyclooxygenase-1 and -2 in normal and glaucomatous human eyes. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2616–2624. [Google Scholar]
- Porcella, A.; Maxia, C.; Gessa, G.L.; Pani, L. The synthetic cannabinoid WIN55212-2 decreases the intraocular pressure in human glaucoma resistant to conventional therapies. Eur. J. Neurosci. 2001, 13, 409–412. [Google Scholar] [CrossRef]
- Njie, Y.F.; Kumar, A.; Qiao, Z.; Zhong, L.; Song, Z.H. Noladin ether acts on trabecular meshwork cannabinoid (CB1) receptors to enhance aqueous humor outflow facility. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1999–2005. [Google Scholar] [CrossRef]
- Tiedeman, J.S.; Shields, M.B.; Weber, P.A.; Crow, J.W.; Cocchetto, D.M.; Harris, W.A.; Howes, J.F. Effect of synthetic cannabinoids on elevated intraocular pressure. Ophthalmology 1981, 88, 270–277. [Google Scholar] [CrossRef]
- Flach, A.J. Delta-9-tetrahydrocannabinol (THC) in the treatment of end-stage open-angle glaucoma. Trans. Am. Ophthalmol. Soc. 2002, 100, 215–222; discussion 222–214. [Google Scholar]
- Tomida, I.; Azuara-Blanco, A.; House, H.; Flint, M.; Pertwee, R.G.; Robson, P.J. Effect of sublingual application of cannabinoids on intraocular pressure: A pilot study. J. Glaucoma 2006, 15, 349–353. [Google Scholar] [CrossRef]
- Plange, N.; Arend, K.O.; Kaup, M.; Doehmen, B.; Adams, H.; Hendricks, S.; Cordes, A.; Huth, J.; Sponsel, W.E.; Remky, A. Dronabinol and retinal hemodynamics in humans. Am. J. Ophthalmol. 2007, 143, 173–174. [Google Scholar] [CrossRef]
- Newell, F.W.; Stark, P.; Jay, W.M.; Schanzlin, D.J. Nabilone: A pressure-reducing synthetic benzopyran in open-angle glaucoma. Ophthalmology 1979, 86, 156–160. [Google Scholar] [CrossRef]
- Green, K. Marijuana smoking vs cannabinoids for glaucoma therapy. Arch. Ophthalmol. 1998, 116, 1433–1437. [Google Scholar] [CrossRef]
- Sun, X.; Xu, C.S.; Chadha, N.; Chen, A.; Liu, J. Marijuana for Glaucoma: A Recipe for Disaster or Treatment? Yale J. Biol. Med. 2015, 88, 265–269. [Google Scholar]
- Strobbe, E.; Cellini, M.; Campos, E.C. Effectiveness of palmitoylethanolamide on endothelial dysfunction in ocular hypertensive patients: A randomized, placebo-controlled cross-over study. Investig. Ophthalmol. Vis. Sci. 2013, 54, 968–973. [Google Scholar] [CrossRef]
- Pescosolido, N.; Librando, A.; Puzzono, M.; Nebbioso, M. Palmitoylethanolamide effects on intraocular pressure after Nd:YAG laser iridotomy: An experimental clinical study. J. Ocul. Pharmacol. Ther. 2011, 27, 629–635. [Google Scholar] [CrossRef]
- Gagliano, C.; Ortisi, E.; Pulvirenti, L.; Reibaldi, M.; Scollo, D.; Amato, R.; Avitabile, T.; Longo, A. Ocular hypotensive effect of oral palmitoyl-ethanolamide: A clinical trial. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6096–6100. [Google Scholar] [CrossRef]
- Huestis, M.A. Human cannabinoid pharmacokinetics. Chem. Biodivers. 2007, 4, 1770–1804. [Google Scholar] [CrossRef]
- Cooler, P.; Gregg, J.M. Effect of delta-9-tetrahydrocannabinol on intraocular pressure in humans. South. Med. J. 1977, 70, 951–954. [Google Scholar] [CrossRef]
- Purnell, W.D.; Gregg, J.M. Delta(9)-tetrahydrocannabinol,, euphoria and intraocular pressure in man. Ann. Ophthalmol. 1975, 7, 921–923. [Google Scholar]
- Grotenhermen, F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin. Pharmacokinet. 2003, 42, 327–360. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.; Weizman, A.; Weinstein, A. Positive and Negative Effects of Cannabis and Cannabinoids on Health. Clin. Pharmacol. Ther. 2019, 105, 1139–1147. [Google Scholar] [CrossRef]
- Merritt, J.C.; Perry, D.D.; Russell, D.N.; Jones, B.F. Topical delta 9-tetrahydrocannabinol and aqueous dynamics in glaucoma. J. Clin. Pharmacol. 1981, 21, 467s–471s. [Google Scholar] [CrossRef]
- Merritt, J.C.; Olsen, J.L.; Armstrong, J.R.; McKinnon, S.M. Topical delta 9-tetrahydrocannabinol in hypertensive glaucomas. J. Pharm. Pharmacol. 1981, 33, 40–41. [Google Scholar] [CrossRef] [PubMed]
- Green, K.; Roth, M. Ocular effects of topical administration of delta 9-tetrahydrocannabinol in man. Arch. Ophthalmol. 1982, 100, 265–267. [Google Scholar] [CrossRef]
- Jay, W.M.; Green, K. Multiple-drop study of topically applied 1% delta 9-tetrahydrocannabinol in human eyes. Arch. Ophthalmol. 1983, 101, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.Y.; Ashkar, S.; Jain, S.; Marchand, M.; Tran, S.D. Breaking Barriers in Eye Treatment: Polymeric Nano-Based Drug-Delivery System for Anterior Segment Diseases and Glaucoma. Polymers 2023, 15, 1373. [Google Scholar] [CrossRef]
- Kabiri, M.; Kamal, S.H.; Pawar, S.V.; Roy, P.R.; Derakhshandeh, M.; Kumar, U.; Hatzikiriakos, S.G.; Hossain, S.; Yadav, V.G. A stimulus-responsive, in situ-forming, nanoparticle-laden hydrogel for ocular drug delivery. Drug Deliv. Transl. Res. 2018, 8, 484–495. [Google Scholar] [CrossRef]
- Saraiva, S.M.; Martín-Banderas, L.; Durán-Lobato, M. Cannabinoid-Based Ocular Therapies and Formulations. Pharmaceutics 2023, 15, 1077. [Google Scholar] [CrossRef]
- Taskar, P.S.; Patil, A.; Lakhani, P.; Ashour, E.; Gul, W.; ElSohly, M.A.; Murphy, B.; Majumdar, S. Δ9-Tetrahydrocannabinol Derivative-Loaded Nanoformulation Lowers Intraocular Pressure in Normotensive Rabbits. Transl. Vis. Sci. Technol. 2019, 8, 15. [Google Scholar] [CrossRef]
- Levin, L.A. Direct and indirect approaches to neuroprotective therapy of glaucomatous optic neuropathy. Surv. Ophthalmol. 1999, 43 (Suppl. S1), S98–S101. [Google Scholar] [CrossRef] [PubMed]
- Bagli, E.; Kitsos, G. Neuroprotective agents in glaucoma. In The Mistery of Glaucoma; InTech: London, UK, 2011; pp. 115–144. [Google Scholar]
- Baltmr, A.; Duggan, J.; Nizari, S.; Salt, T.E.; Cordeiro, M.F. Neuroprotection in glaucoma—Is there a future role? Exp. Eye Res. 2010, 91, 554–566. [Google Scholar] [CrossRef]
- Coyle, J.T.; Puttfarcken, P. Oxidative stress, glutamate, and neurodegenerative disorders. Science 1993, 262, 689–695. [Google Scholar] [CrossRef]
- El-Remessy, A.B.; Khalil, I.E.; Matragoon, S.; Abou-Mohamed, G.; Tsai, N.J.; Roon, P.; Caldwell, R.B.; Caldwell, R.W.; Green, K.; Liou, G.I. Neuroprotective effect of (-)Delta9-tetrahydrocannabinol and cannabidiol in N-methyl-D-aspartate-induced retinal neurotoxicity: Involvement of peroxynitrite. Am. J. Pathol. 2003, 163, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- van der Stelt, M.; Veldhuis, W.B.; Bär, P.R.; Veldink, G.A.; Vliegenthart, J.F.; Nicolay, K. Neuroprotection by Delta9-tetrahydrocannabinol, the main active compound in marijuana, against ouabain-induced in vivo excitotoxicity. J. Neurosci. 2001, 21, 6475–6479. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Thayer, S.A. Cannabinoid receptor agonists protect cultured rat hippocampal neurons from excitotoxicity. Mol. Pharmacol. 1998, 54, 459–462. [Google Scholar] [CrossRef]
- Nagayama, T.; Sinor, A.D.; Simon, R.P.; Chen, J.; Graham, S.H.; Jin, K.; Greenberg, D.A. Cannabinoids and neuroprotection in global and focal cerebral ischemia and in neuronal cultures. J. Neurosci. 1999, 19, 2987–2995. [Google Scholar] [CrossRef]
- Tadijan, A.; Vlašić, I.; Vlainić, J.; Đikić, D.; Oršolić, N.; Jazvinšćak Jembrek, M. Intracellular Molecular Targets and Signaling Pathways Involved in Antioxidative and Neuroprotective Effects of Cannabinoids in Neurodegenerative Conditions. Antioxidants 2022, 11, 2049. [Google Scholar] [CrossRef]
- Paloczi, J.; Varga, Z.V.; Hasko, G.; Pacher, P. Neuroprotection in Oxidative Stress-Related Neurodegenerative Diseases: Role of Endocannabinoid System Modulation. Antioxid. Redox Signal. 2018, 29, 75–108. [Google Scholar] [CrossRef]
- Benyó, Z.; Ruisanchez, É.; Leszl-Ishiguro, M.; Sándor, P.; Pacher, P. Endocannabinoids in cerebrovascular regulation. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H785–H801. [Google Scholar] [CrossRef] [PubMed]
- Leonelli, M.; Martins, D.O.; Britto, L.R. Retinal cell death induced by TRPV1 activation involves NMDA signaling and upregulation of nitric oxide synthases. Cell Mol. Neurobiol. 2013, 33, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Leonelli, M.; Martins, D.O.; Britto, L.R. TRPV1 receptors are involved in protein nitration and Müller cell reaction in the acutely axotomized rat retina. Exp. Eye Res. 2010, 91, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Kawamata, T.; Ninomiya, T.; Omote, K.; Namiki, A. Endothelin-1 enhances capsaicin-evoked intracellular Ca2+ response via activation of endothelin a receptor in a protein kinase Cepsilon-dependent manner in dorsal root ganglion neurons. Neuroscience 2006, 137, 949–960. [Google Scholar] [CrossRef]
- Lau, J.; Dang, M.; Hockmann, K.; Ball, A.K. Effects of acute delivery of endothelin-1 on retinal ganglion cell loss in the rat. Exp. Eye Res. 2006, 82, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Weitlauf, C.; Ward, N.J.; Lambert, W.S.; Sidorova, T.N.; Ho, K.W.; Sappington, R.M.; Calkins, D.J. Short-term increases in transient receptor potential vanilloid-1 mediate stress-induced enhancement of neuronal excitation. J. Neurosci. 2014, 34, 15369–15381. [Google Scholar] [CrossRef]
- Nucci, C.; Martucci, A.; Giannini, C.; Morrone, L.A.; Bagetta, G.; Mancino, R. Neuroprotective agents in the management of glaucoma. Eye 2018, 32, 938–945. [Google Scholar] [CrossRef]
- Vasudevan, S.K.; Gupta, V.; Crowston, J.G. Neuroprotection in glaucoma. Indian J. Ophthalmol. 2011, 59 (Suppl. S1), S102–S113. [Google Scholar] [CrossRef]
- Crandall, J.; Matragoon, S.; Khalifa, Y.M.; Borlongan, C.; Tsai, N.T.; Caldwell, R.B.; Liou, G.I. Neuroprotective and intraocular pressure-lowering effects of (-)Delta9-tetrahydrocannabinol in a rat model of glaucoma. Ophthalmic Res. 2007, 39, 69–75. [Google Scholar] [CrossRef]
- Pinar-Sueiro, S.; Zorrilla Hurtado, J.; Veiga-Crespo, P.; Sharma, S.C.; Vecino, E. Neuroprotective effects of topical CB1 agonist WIN 55212-2 on retinal ganglion cells after acute rise in intraocular pressure induced ischemia in rat. Exp. Eye Res. 2013, 110, 55–58. [Google Scholar] [CrossRef]
- Hui-Feng, L.; Yuan, H.; Jun, J.; Ming-Li, J.; Jin-Wei, X. Clinical study on intravitreal injection of cannabinoid HU-211 for optic nerve damage in glaucoma rats. Int. Eye Sci. 2014, 14, 1584–1586. [Google Scholar]
- Nucci, C.; Gasperi, V.; Tartaglione, R.; Cerulli, A.; Terrinoni, A.; Bari, M.; De Simone, C.; Agrò, A.F.; Morrone, L.A.; Corasaniti, M.T.; et al. Involvement of the endocannabinoid system in retinal damage after high intraocular pressure-induced ischemia in rats. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2997–3004. [Google Scholar] [CrossRef]
- Zalish, M.; Lavie, V. Dexanabinol (HU-211) has a beneficial effect on axonal sprouting and survival after rat optic nerve crush injury. Vision Res. 2003, 43, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Yoles, E.; Belkin, M.; Schwartz, M. HU-211, a nonpsychotropic cannabinoid, produces short- and long-term neuroprotection after optic nerve axotomy. J. Neurotrauma 1996, 13, 49–57. [Google Scholar] [CrossRef] [PubMed]
- El-Remessy, A.B.; Al-Shabrawey, M.; Khalifa, Y.; Tsai, N.T.; Caldwell, R.B.; Liou, G.I. Neuroprotective and blood-retinal barrier-preserving effects of cannabidiol in experimental diabetes. Am. J. Pathol. 2006, 168, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Lax, P.; Esquiva, G.; Altavilla, C.; Cuenca, N. Neuroprotective effects of the cannabinoid agonist HU210 on retinal degeneration. Exp. Eye Res. 2014, 120, 175–185. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, X.; Liu, S.; Shen, Y. Neuroprotective effect of cannabinoid receptor 1 antagonist in the MNU-induced retinal degeneration model. Exp. Eye Res. 2018, 167, 145–151. [Google Scholar] [CrossRef]
- Kokona, D.; Spyridakos, D.; Tzatzarakis, M.; Papadogkonaki, S.; Filidou, E.; Arvanitidis, K.I.; Kolios, G.; Lamani, M.; Makriyannis, A.; Malamas, M.S.; et al. The endocannabinoid 2-arachidonoylglycerol and dual ABHD6/MAGL enzyme inhibitors display neuroprotective and anti-inflammatory actions in the in vivo retinal model of AMPA excitotoxicity. Neuropharmacology 2021, 185, 108450. [Google Scholar] [CrossRef]
- Slusar, J.E.; Cairns, E.A.; Szczesniak, A.M.; Bradshaw, H.B.; Di Polo, A.; Kelly, M.E. The fatty acid amide hydrolase inhibitor, URB597, promotes retinal ganglion cell neuroprotection in a rat model of optic nerve axotomy. Neuropharmacology 2013, 72, 116–125. [Google Scholar] [CrossRef]
- Maccarone, R.; Rapino, C.; Zerti, D.; di Tommaso, M.; Battista, N.; Di Marco, S.; Bisti, S.; Maccarrone, M. Modulation of Type-1 and Type-2 Cannabinoid Receptors by Saffron in a Rat Model of Retinal Neurodegeneration. PLoS ONE 2016, 11, e0166827. [Google Scholar] [CrossRef]
- Sappington, R.M.; Sidorova, T.; Ward, N.J.; Chakravarthy, R.; Ho, K.W.; Calkins, D.J. Activation of transient receptor potential vanilloid-1 (TRPV1) influences how retinal ganglion cell neurons respond to pressure-related stress. Channels 2015, 9, 102–113. [Google Scholar] [CrossRef]
- Sakamoto, K.; Kuroki, T.; Okuno, Y.; Sekiya, H.; Watanabe, A.; Sagawa, T.; Ito, H.; Mizuta, A.; Mori, A.; Nakahara, T.; et al. Activation of the TRPV1 channel attenuates N-methyl-D-aspartic acid-induced neuronal injury in the rat retina. Eur. J. Pharmacol. 2014, 733, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, R.; Lu, H.; Zhang, X. Systemic administration with tetrahydrocannabinol causes retinal damage in BALB/c mice. Hum. Exp. Toxicol. 2020, 39, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.C.M.; Scudeller, L.; Lumini, C.; Bettio, F.; Picasso, E.; Ruberto, G.; Briola, A.; Mirabile, A.; Paviglianiti, A.; Pasinetti, G.M.; et al. Effect of palmitoylethanolamide on inner retinal function in glaucoma: A randomized, single blind, crossover, clinical trial by pattern-electroretinogram. Sci. Rep. 2020, 10, 10468. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.R.; Lograno, M.D. Involvement of the peroxisome proliferator-activated receptor (PPAR) alpha in vascular response of endocannabinoids in the bovine ophthalmic artery. Eur. J. Pharmacol. 2012, 683, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Schwitzer, T.; Schwan, R.; Albuisson, E.; Giersch, A.; Lalanne, L.; Angioi-Duprez, K.; Laprevote, V. Association Between Regular Cannabis Use and Ganglion Cell Dysfunction. JAMA Ophthalmol. 2017, 135, 54–60. [Google Scholar] [CrossRef]
- Schwitzer, T.; Schwan, R.; Angioi-Duprez, K.; Giersch, A.; Lalanne, L.; Albuisson, E.; Laprevote, V. Delayed bipolar and ganglion cells neuroretinal processing in regular cannabis users: The retina as a relevant site to investigate brain synaptic transmission dysfunctions. J. Psychiatr. Res. 2018, 103, 75–82. [Google Scholar] [CrossRef]
- Schwitzer, T.; Henrion, M.L.; Sarre, D.; Albuisson, E.; Angioi-Duprez, K.; Giersch, A.; Lalanne, L.; Schwan, R.; Laprevote, V. Spatial localization of retinal anomalies in regular cannabis users: The relevance of the multifocal electroretinogram. Schizophr. Res. 2020, 219, 56–61. [Google Scholar] [CrossRef]
- Lucas, A.; Thirion, A.; Schwan, R.; Krieg, J.; Angioi-Duprez, K.; Laprevote, V.; Schwitzer, T. Association between increased retinal background noise and co-occurrent regular cannabis and alcohol use. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 89, 335–340. [Google Scholar] [CrossRef]
- Zantut, P.R.A.; Veras, M.M.; Yariwake, V.Y.; Takahashi, W.Y.; Saldiva, P.H.; Young, L.H.; Damico, F.M.; Fajersztajn, L. Effects of cannabis and its components on the retina: A systematic review. Cutan. Ocul. Toxicol. 2020, 39, 1–9. [Google Scholar] [CrossRef]
- Arend, O.; Plange, N.; Sponsel, W.E.; Remky, A. Pathogenetic aspects of the glaucomatous optic neuropathy: Fluorescein angiographic findings in patients with primary open angle glaucoma. Brain Res. Bull. 2004, 62, 517–524. [Google Scholar] [CrossRef]
- Flammer, J. The vascular concept of glaucoma. Surv. Ophthalmol. 1994, 38, S3–S6. [Google Scholar] [CrossRef]
- Resch, H.; Garhofer, G.; Fuchsjäger-Mayrl, G.; Hommer, A.; Schmetterer, L. Endothelial dysfunction in glaucoma. Acta Ophthalmol. 2009, 87, 4–12. [Google Scholar] [CrossRef]
- Schmidl, D.; Garhofer, G.; Schmetterer, L. The complex interaction between ocular perfusion pressure and ocular blood flow-relevance for glaucoma. Exp. Eye Res. 2011, 93, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Schmetterer, L.; Kiel, J. Ocular Blood Flow; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Yaoeda, K.; Shirakashi, M.; Fukushima, A.; Funaki, S.; Funaki, H.; Abe, H.; Tanabe, N. Relationship between optic nerve head microcirculation and visual field loss in glaucoma. Acta Ophthalmol. Scand. 2003, 81, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Shiga, Y.; Kunikata, H.; Aizawa, N.; Kiyota, N.; Maiya, Y.; Yokoyama, Y.; Omodaka, K.; Takahashi, H.; Yasui, T.; Kato, K.; et al. Optic Nerve Head Blood Flow, as Measured by Laser Speckle Flowgraphy, Is Significantly Reduced in Preperimetric Glaucoma. Curr. Eye Res. 2016, 41, 1447–1453. [Google Scholar] [CrossRef]
- Harris, A.; Zarfati, D.; Zalish, M.; Biller, J.; Sheets, C.W.; Rechtman, E.; Migliardi, R.; Garzozi, H.J. Reduced cerebrovascular blood flow velocities and vasoreactivity in open-angle glaucoma. Am. J. Ophthalmol. 2003, 135, 144–147. [Google Scholar] [CrossRef]
- Calzetti, G.; Mursch-Edlmayr, A.S.; Bata, A.M.; Ungaro, N.; Mora, P.; Chua, J.; Schmidl, D.; Bolz, M.; Garhöfer, G.; Gandolfi, S.; et al. Measuring optic nerve head perfusion to monitor glaucoma: A study on structure-function relationships using laser speckle flowgraphy. Acta Ophthalmol. 2022, 100, e181–e191. [Google Scholar] [CrossRef] [PubMed]
- Garhöfer, G.; Bata, A.M.; Popa-Cherecheanu, A.; Hommer, A.; Vass, C.; Resch, H.; Schmidl, D.; Werkmeister, R.M.; Schmetterer, L. Retinal Oxygen Extraction in Patients with Primary Open-Angle Glaucoma. Int. J. Mol. Sci. 2022, 23, 10152. [Google Scholar] [CrossRef]
- Mozaffarieh, M.; Flammer, J. Is there more to glaucoma treatment than lowering IOP? Surv. Ophthalmol. 2007, 52 (Suppl. S2), S174–S179. [Google Scholar] [CrossRef] [PubMed]
- Bata, A.M.; Fondi, K.; Witkowska, K.J.; Werkmeister, R.M.; Hommer, A.; Vass, C.; Resch, H.; Schmidl, D.; Popa-Cherecheanu, A.; Chua, J.; et al. Optic nerve head blood flow regulation during changes in arterial blood pressure in patients with primary open-angle glaucoma. Acta Ophthalmol. 2019, 97, e36–e41. [Google Scholar] [CrossRef]
- Fuchsjäger-Mayrl, G.; Wally, B.; Georgopoulos, M.; Rainer, G.; Kircher, K.; Buehl, W.; Amoako-Mensah, T.; Eichler, H.G.; Vass, C.; Schmetterer, L. Ocular blood flow and systemic blood pressure in patients with primary open-angle glaucoma and ocular hypertension. Investig. Ophthalmol. Vis. Sci. 2004, 45, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Fuchsjäger-Mayrl, G.; Georgopoulos, M.; Hommer, A.; Weigert, G.; Pemp, B.; Vass, C.; Garhöfer, G.; Schmetterer, L. Effect of dorzolamide and timolol on ocular pressure: Blood flow relationship in patients with primary open-angle glaucoma and ocular hypertension. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1289–1296. [Google Scholar] [CrossRef]
- Garhöfer, G.; Resch, H.; Weigert, G.; Lung, S.; Simader, C.; Schmetterer, L. Short-term increase of intraocular pressure does not alter the response of retinal and optic nerve head blood flow to flicker stimulation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1721–1725. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.E.; Oh, S.; Baek, S.U.; Ahn, S.J.; Park, K.H.; Jeoung, J.W. Ocular Perfusion Pressure and the Risk of Open-Angle Glaucoma: Systematic Review and Meta-analysis. Sci. Rep. 2020, 10, 10056. [Google Scholar] [CrossRef]
- Stanley, C.; O’Sullivan, S.E. Vascular targets for cannabinoids: Animal and human studies. Br. J. Pharmacol. 2014, 171, 1361–1378. [Google Scholar] [CrossRef] [PubMed]
- Lípez-Miranda, V.; Herradón, E.; Martín, M.I. Vasorelaxation caused by cannabinoids: Mechanisms in different vascular beds. Curr. Vasc. Pharmacol. 2008, 6, 335–346. [Google Scholar] [CrossRef]
- Randall, M.D.; Kendall, D.A.; O’Sullivan, S. The complexities of the cardiovascular actions of cannabinoids. Br. J. Pharmacol. 2004, 142, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Hillard, C.J. Endocannabinoids and vascular function. J. Pharmacol. Exp. Ther. 2000, 294, 27–32. [Google Scholar] [PubMed]
- Zoratti, C.; Kipmen-Korgun, D.; Osibow, K.; Malli, R.; Graier, W.F. Anandamide initiates Ca2+ signaling via CB2 receptor linked to phospholipase C in calf pulmonary endothelial cells. Br. J. Pharmacol. 2003, 140, 1351–1362. [Google Scholar] [CrossRef]
- Alswailmi, F.K. A Cross Talk between the Endocannabinoid System and Different Systems Involved in the Pathogenesis of Hypertensive Retinopathy. Pharmaceuticals 2023, 16, 345. [Google Scholar] [CrossRef]
- Chataigneau, T.; Félétou, M.; Thollon, C.; Villeneuve, N.; Vilaine, J.P.; Duhault, J.; Vanhoutte, P.M. Cannabinoid CB1 receptor and endothelium-dependent hyperpolarization in guinea-pig carotid, rat mesenteric and porcine coronary arteries. Br. J. Pharmacol. 1998, 123, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Ronco, A.M.; Llanos, M.; Tamayo, D.; Hirsch, S. Anandamide inhibits endothelin-1 production by human cultured endothelial cells: A new vascular action of this endocannabinoid. Pharmacology 2007, 79, 12–16. [Google Scholar] [CrossRef]
- Su, E.N.; Kelly, M.E.; Cringle, S.J.; Yu, D.Y. Role of Endothelium in Abnormal Cannabidiol-Induced Vasoactivity in Retinal Arterioles. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4029–4037. [Google Scholar] [CrossRef] [PubMed]
- Ripodas, A.; de Juan, J.A.; Roldán-Pallarés, M.; Bernal, R.; Moya, J.; Chao, M.; López, A.; Fernández-Cruz, A.; Fernández-Durango, R. Localisation of endothelin-1 mRNA expression and immunoreactivity in the retina and optic nerve from human and porcine eye. Evidence for endothelin-1 expression in astrocytes. Brain Res. 2001, 912, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.R.; Lograno, M.D. Cannabinoid agonists induce relaxation in the bovine ophthalmic artery: Evidences for CB1 receptors, nitric oxide and potassium channels. Br. J. Pharmacol. 2006, 147, 917–925. [Google Scholar] [CrossRef]
- Sultan, S.R.; Millar, S.A.; O’Sullivan, S.E.; England, T.J. A Systematic Review and Meta-Analysis of the In Vivo Haemodynamic Effects of Δ9-Tetrahydrocannabinol. Pharmaceuticals 2018, 11, 13. [Google Scholar] [CrossRef]
- Green, K.; Wynn, H.; Padgett, D. Effects of delta9-tetrahydrocannabinol on ocular blood flow and aqueous humor formation. Exp. Eye Res. 1978, 26, 65–69. [Google Scholar] [CrossRef]
- Ohlsson, A.; Lindgren, J.E.; Wahlen, A.; Agurell, S.; Hollister, L.E.; Gillespie, H.K. Plasma delta-9 tetrahydrocannabinol concentrations and clinical effects after oral and intravenous administration and smoking. Clin. Pharmacol. Ther. 1980, 28, 409–416. [Google Scholar] [CrossRef]
- Ogunbiyi, M.O.; Hindocha, C.; Freeman, T.P.; Bloomfield, M.A.P. Acute and chronic effects of Δ9-tetrahydrocannabinol (THC) on cerebral blood flow: A systematic review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 101, 109900. [Google Scholar] [CrossRef]
- Hill, M.; Wong, T.Y.; Davis, M.; Meier, M.H. Associations between cannabis use and retinal vessel diameter in young adults. Schizophr. Res. 2020, 219, 62–68. [Google Scholar] [CrossRef]
- Hommer, N.; Kallab, M.; Szegedi, S.; Puchner, S.; Stjepanek, K.; Bauer, M.; Werkmeister, R.M.; Schmetterer, L.; Abensperg-Traun, M.; Garhöfer, G.; et al. The Effect of Orally Administered Dronabinol on Optic Nerve Head Blood Flow in Healthy Subjects-A Randomized Clinical Trial. Clin. Pharmacol. Ther. 2020, 108, 155–161. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindner, T.; Schmidl, D.; Peschorn, L.; Pai, V.; Popa-Cherecheanu, A.; Chua, J.; Schmetterer, L.; Garhöfer, G. Therapeutic Potential of Cannabinoids in Glaucoma. Pharmaceuticals 2023, 16, 1149. https://doi.org/10.3390/ph16081149
Lindner T, Schmidl D, Peschorn L, Pai V, Popa-Cherecheanu A, Chua J, Schmetterer L, Garhöfer G. Therapeutic Potential of Cannabinoids in Glaucoma. Pharmaceuticals. 2023; 16(8):1149. https://doi.org/10.3390/ph16081149
Chicago/Turabian StyleLindner, Theresa, Doreen Schmidl, Laura Peschorn, Viktoria Pai, Alina Popa-Cherecheanu, Jacqueline Chua, Leopold Schmetterer, and Gerhard Garhöfer. 2023. "Therapeutic Potential of Cannabinoids in Glaucoma" Pharmaceuticals 16, no. 8: 1149. https://doi.org/10.3390/ph16081149
APA StyleLindner, T., Schmidl, D., Peschorn, L., Pai, V., Popa-Cherecheanu, A., Chua, J., Schmetterer, L., & Garhöfer, G. (2023). Therapeutic Potential of Cannabinoids in Glaucoma. Pharmaceuticals, 16(8), 1149. https://doi.org/10.3390/ph16081149