
The Therapeutic and Diagnostic Potential of Phospholipase C Zeta, Oocyte Activation, and Calcium in Treating Human Infertility

 , and
, and 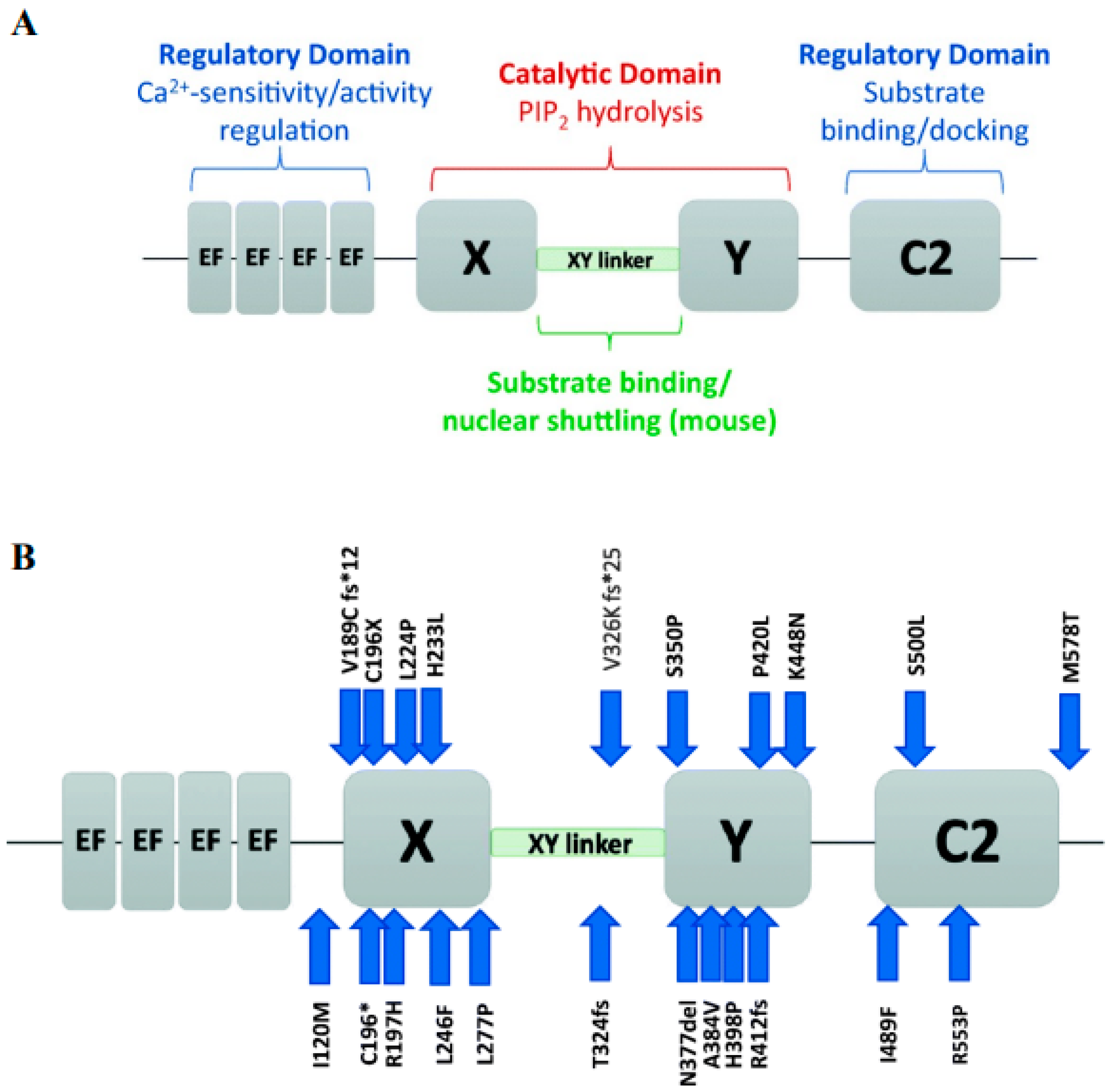{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Endoplasmic Reticulum (ER)
3. Oocyte Mitochondria
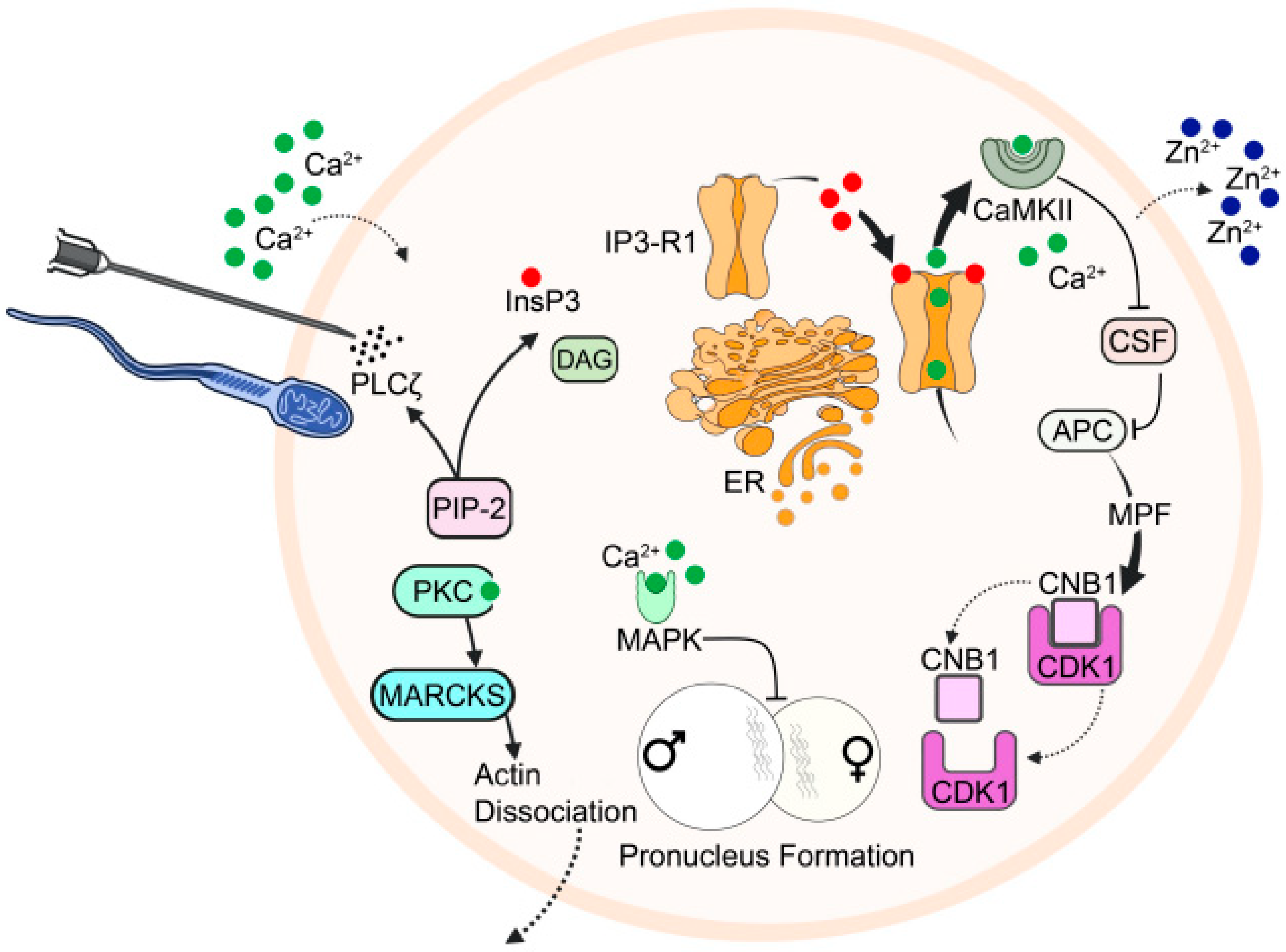
4. Ca2+ Oscillation Models
5. The Ca2+ Conduit Model
6. The Membrane Receptor Model
7. The Soluble Sperm Factor
8. The Mammalian Sperm Factor: Phospholipase C Zeta
9. PLCζ Structure and Function
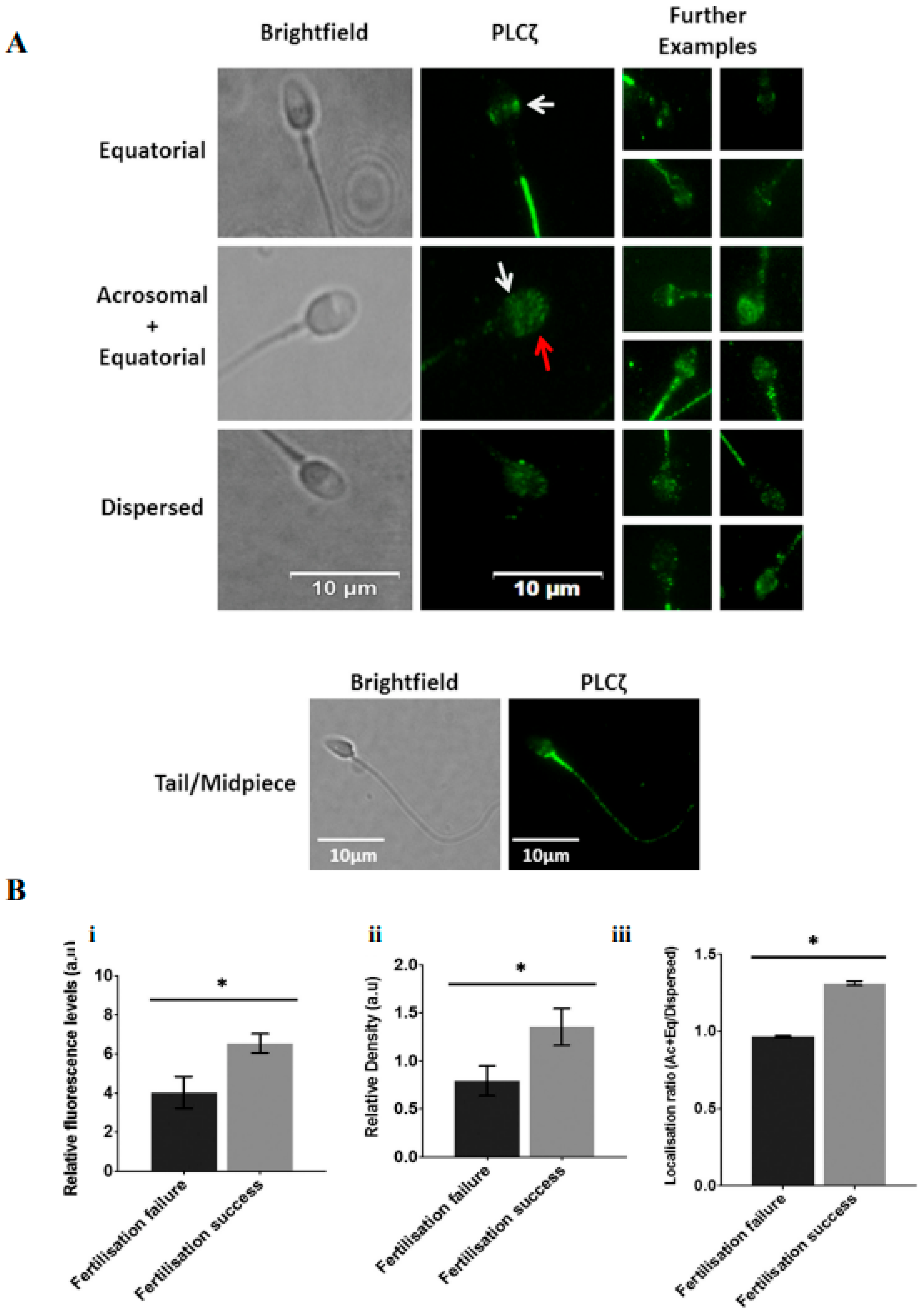
10. Abnormal Expression and Localization in Sperm
11. PLCzeta in Human Male Infertility
12. Assisted Oocyte Activation (AOA)
13. Electrical Activation
14. Mechanical Activation
15. Chemical Activation
16. Therapeutic and Diagnostic Options for PLCζ

17. Alternative Diagnostic and Therapeutic Targets of OA
18. Actin-Mediated Cytoskeletal Movements
19. Modulators of Ca2+ Homeostasis
20. The Role of Zinc (Zn2+)
21. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, C.J. Signalling mechanisms of mammalian oocyte activation. Hum. Reprod. Update 2002, 8, 313–321. [Google Scholar] [CrossRef]
- Robker, R.L.; Hennebold, J.D.; Russell, D.L. Coordination of Ovulation and Oocyte Maturation: A Good Egg at the Right Time. Endocrinology 2018, 159, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Caiazza, F. Oocyte maturation: A story of arrest and release. FBS 2013, 5, 451–477. [Google Scholar] [CrossRef] [PubMed]
- Thanassoulas, A.; Swann, K.; Lai, F.A.; Nomikos, M. SPERM FACTORS AND EGG ACTIVATION: The structure and function relationship of sperm PLCZ1. Reproduction 2022, 164, F1–F8. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S. Thirty years of calcium signals at fertilization. Semin. Cell Dev. Biol. 2006, 17, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Savy, V.; Stein, P.; Shi, M.; Williams, C.J. PMCA1 depletion in mouse eggs amplifies calcium signaling and impacts offspring growth†. Biol. Reprod. 2022, ioac180. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Ganesh, D.; Jones, C.; Coward, K. Oocyte activation deficiency and assisted oocyte activation: Mechanisms, obstacles and prospects for clinical application. Hum. Reprod. Open 2022, 2022, hoac003. [Google Scholar] [CrossRef]
- Swann, K.; Yu, Y. The dynamics of calcium oscillations that activate mammalian eggs. Int. J. Dev. Biol. 2008, 52, 585–594. [Google Scholar] [CrossRef]
- Fulton, B.P.; Whittingham, D.G. Activation of mammalian oocytes by intracellular injection of calcium. Nature 1978, 273, 149–151. [Google Scholar] [CrossRef]
- Miyazaki, S.; Shirakawa, H.; Nakada, K.; Honda, Y. Essential role of the inositol 1,4,5-trisphosphate receptor/Ca2+ release channel in Ca2+ waves and Ca2+ oscillations at fertilization of mammalian eggs. Dev. Biol. 1993, 158, 62–78. [Google Scholar] [CrossRef]
- Brind, S.; Swann, K.; Carroll, J. Inositol 1,4,5-Trisphosphate Receptors Are Downregulated in Mouse Oocytes in Response to Sperm or Adenophostin A but Not to Increases in Intracellular Ca2+ or Egg Activation. Dev. Biol. 2000, 223, 251–265. [Google Scholar] [CrossRef]
- Jellerette, T.; He, C.L.; Wu, H.; Parys, J.B.; Fissore, R.A. Down-regulation of the Inositol 1,4,5-Trisphosphate Receptor in Mouse Eggs Following Fertilization or Parthenogenetic Activation. Dev. Biol. 2000, 223, 238–250. [Google Scholar] [CrossRef]
- Xu, Z.; Williams, C.J.; Kopf, G.S.; Schultz, R.M. Maturation-associated increase in IP3 receptor type 1: Role in conferring increased IP3 sensitivity and Ca2+ oscillatory behavior in mouse eggs. Dev. Biol. 2003, 254, 163–171. [Google Scholar] [CrossRef]
- Boulware, M.J.; Marchant, J.S. Timing in cellular Ca2+ signaling. Curr. Biol. 2008, 18, R769–R776. [Google Scholar] [CrossRef]
- Alhajeri, M.M.; Alkhanjari, R.R.; Hodeify, R.; Khraibi, A.; Hamdan, H. Neurotransmitters, neuropeptides and calcium in oocyte maturation and early development. Front. Cell Dev. Biol. 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Yang, D.; Lan, X.; Li, K.; Li, X.; Chen, J.; Zhang, Y.; Xiao, R.-P.; Han, Q.; Cheng, H. Nuclear Ca2+ sparks and waves mediated by inositol 1,4,5-trisphosphate receptors in neonatal rat cardiomyocytes. Cell Calcium 2008, 43, 165–174. [Google Scholar] [CrossRef]
- Gordienko, D.V.; Bolton, T.B. Crosstalk between ryanodine receptors and IP3 receptors as a factor shaping spontaneous Ca2+-release events in rabbit portal vein myocytes. J. Physiol. 2002, 542, 743–762. [Google Scholar] [CrossRef]
- Sun, B.; Yeh, J. Calcium Oscillatory Patterns and Oocyte Activation During Fertilization: A Possible Mechanism for Total Fertilization Failure (TFF) in Human In Vitro Fertilization? Reprod. Sci. 2021, 28, 639–648. [Google Scholar] [CrossRef]
- Wakai, T.; Vanderheyden, V.; Fissore, R.A. Ca2+ signaling during mammalian fertilization: Requirements, players, and adaptations. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Reber, S.; Over, S.; Kronja, I.; Gruss, O.J. CaM kinase II initiates meiotic spindle depolymerization independently of APC/C activation. J. Cell Biol. 2008, 183, 1007–1017. [Google Scholar] [CrossRef]
- Castro, A.; Bernis, C.; Vigneron, S.; Labbé, J.C.; Lorca, T. The anaphase-promoting complex: A key factor in the regulation of cell cycle. Oncogene 2005, 24, 314–325. [Google Scholar] [CrossRef]
- Jones, K.T.; Lane, S.I.R. Molecular causes of aneuploidy in mammalian eggs. Development 2013, 140, 3719–3730. [Google Scholar] [CrossRef]
- Ducibella, T.; Schultz, R.M.; Ozil, J.P. Role of calcium signals in early development. Semin. Cell Dev. Biol. 2006, 17, 324–332. [Google Scholar] [CrossRef]
- Igarashi, H.; Knott, J.G.; Schultz, R.M.; Williams, C.J. Alterations of PLCβ1 in mouse eggs change calcium oscillatory behavior following fertilization. Dev. Biol. 2007, 312, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Buitrago, M.; Bonte, D.; Dhaenens, L.; Vermorgen, S.; Lu, Y.; De Sutter, P.; Heindryckx, B. Assessment of the calcium releasing machinery in oocytes that failed to fertilize after conventional ICSI and assisted oocyte activation. Reprod. BioMed. Online 2019, 38, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Stein, P.; Savy, V.; Williams, A.M.; Williams, C.J. Modulators of calcium signalling at fertilization. Open Biol. 2020, 10, 200118. [Google Scholar] [CrossRef]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef]
- Kashir, J.; Jones, C.; Coward, K. Calcium oscillations, oocyte activation, and phospholipase C zeta. Adv. Exp. Med. Biol. 2012, 740, 1095–1121. [Google Scholar] [CrossRef]
- Nakano, Y. Spatiotemporal dynamics of intracellular calcium in the mouse egg injected with a spermatozoon. Mol. Hum. Reprod. 1997, 3, 1087–1093. [Google Scholar] [CrossRef]
- Tesarik, J.; Sousa, M. Comparison of Ca2+ responses in human oocytes fertilized by subzonal insemination and by intracytoplasmic sperm injection. Fertil. Steril. 1994, 62, 1197–1204. [Google Scholar] [CrossRef]
- Kurokawa, M.; Fissore, R.A. ICSI-generated mouse zygotes exhibit altered calcium oscillations, inositol 1, 4, 5-trisphosphate receptor-1 down-regulation, and embryo development. Mol. Hum. Reprod. 2003, 9, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Nomikos, M.; Lai, F.A.; Swann, K. Sperm-induced Ca2+ release during egg activation in mammals. Biochem. Biophys. Res. Commun. 2014, 450, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Ducibella, T.; Fissore, R. The roles of Ca2+, downstream protein kinases, and oscillatory signaling in regulating fertilization and the activation of development. Dev. Biol. 2008, 315, 257–279. [Google Scholar] [CrossRef]
- Lee, B.; Yoon, S.-Y.; Fissore, R.A. Regulation of fertilization-initiated [Ca2+]i oscillations in mammalian eggs: A multi-pronged approach. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2006; pp. 274–284. [Google Scholar]
- Swann, K. A cytosolic sperm factor stimulates repetitive calcium increases and mimics fertilization in hamster eggs. Development 1990, 110, 1295–1302. [Google Scholar] [CrossRef]
- Swann, K.; Igusa, Y.; Miyazaki, S. Evidence for an inhibitory effect of protein kinase C on G-protein-mediated repetitive calcium transients in hamster eggs. EMBO J. 1989, 8, 3711–3718. [Google Scholar] [CrossRef]
- Aarabi, M.; Qin, Z.; Xu, W.; Mewburn, J.; Oko, R. Sperm-borne protein, PAWP, initiates zygotic development in Xenopus laevis by eliciting intracellular calcium release. Mol. Reprod. Dev. 2010, 77, 249–256. [Google Scholar] [CrossRef]
- Aarabi, M.; Balakier, H.; Bashar, S.; Moskovtsev, S.I.; Sutovsky, P.; Librach, C.L.; Oko, R. Sperm-derived WW domain-binding protein, PAWP, elicits calcium oscillations and oocyte activation in humans and mice. FASEB J. 2014, 28, 4434–4440. [Google Scholar] [CrossRef]
- Wu, A.T.; Sutovsky, P.; Manandhar, G.; Xu, W.; Katayama, M.; Day, B.N.; Park, K.W.; Yi, Y.J.; Xi, Y.W.; Prather, R.S.; et al. PAWP, a sperm-specific WW domain-binding protein, promotes meiotic resumption and pronuclear development during fertilization. J. Biol. Chem. 2007, 282, 12164–12175. [Google Scholar] [CrossRef]
- Nomikos, M.; Sanders, J.R.; Theodoridou, M.; Kashir, J.; Matthews, E.; Nounesis, G.; Lai, F.A.; Swann, K. Sperm-specific post-acrosomal WW-domain binding protein (PAWP) does not cause Ca2+ release in mouse oocytes. Mol. Hum. Reprod. 2014, 20, 938–947. [Google Scholar] [CrossRef]
- Sette, C.; Bevilacqua, A.; Bianchini, A.; Mangia, F.; Geremia, R.; Rossi, P. Parthenogenetic activation of mouse eggs by microinjection of a truncated c-kit tyrosine kinase present in spermatozoa. Development 1997, 124, 2267–2274. [Google Scholar] [CrossRef]
- Sette, C.; Paronetto, M.P.; Barchi, M.; Bevilacqua, A.; Geremia, R.; Rossi, P. Tr-kit-induced resumption of the cell cycle in mouse eggs requires activation of a Src-like kinase. EMBO J. 2002, 21, 5386–5395. [Google Scholar] [CrossRef]
- Nomikos, M.; Swann, K.; Lai, F.A. Starting a new life: Sperm PLC-zeta mobilizes the Ca2+ signal that induces egg activation and embryo development: An essential phospholipase C with implications for male infertility. Bioessays 2012, 34, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Nomikos, M.; Kashir, J.; Swann, K.; Lai, F.A. Sperm PLCζ: From structure to Ca2+ oscillations, egg activation and therapeutic potential. FEBS Lett. 2013, 587, 3609–3616. [Google Scholar] [CrossRef] [PubMed]
- Nomikos, M.; Kashir, J.; Lai, F.A. The role and mechanism of action of sperm PLC-zeta in mammalian fertilisation. Biochem. J. 2017, 474, 3659–3673. [Google Scholar] [CrossRef]
- Berridge, M.J.; Irvine, R.F. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature 1984, 312, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Inanobe, S.; Kanemaru, K.; Nakamura, Y. Phospholipase C is a key enzyme regulating intracellular calcium and modulating the phosphoinositide balance. Prog. Lipid Res. 2010, 49, 429–437. [Google Scholar] [CrossRef]
- Nishizuka, Y. The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature 1988, 334, 661–665. [Google Scholar] [CrossRef]
- Hwang, J.I.; Oh, Y.S.; Shin, K.J.; Kim, H.; Ryu, S.H.; Suh, P.G. Molecular cloning and characterization of a novel phospholipase C, PLC-eta. Biochem. J. 2005, 389, 181–186. [Google Scholar] [CrossRef]
- Nakahara, M.; Shimozawa, M.; Nakamura, Y.; Irino, Y.; Morita, M.; Kudo, Y.; Fukami, K. A Novel Phospholipase C, PLCη2, Is a Neuron-specific Isozyme*. J. Biol. Chem. 2005, 280, 29128–29134. [Google Scholar] [CrossRef]
- Rhee, S.G. Regulation of phosphoinositide-specific phospholipase C. Annu. Rev. Biochem. 2001, 70, 281–312. [Google Scholar] [CrossRef]
- Zhou, Y.; Wing, M.R.; Sondek, J.; Harden, T.K. Molecular cloning and characterization of PLC-eta2. Biochem. J. 2005, 391, 667–676. [Google Scholar] [CrossRef]
- Nomikos, M.; Blayney, L.M.; Larman, M.G.; Campbell, K.; Rossbach, A.; Saunders, C.M.; Swann, K.; Lai, F.A. Role of phospholipase C-zeta domains in Ca2+-dependent phosphatidylinositol 4,5-bisphosphate hydrolysis and cytoplasmic Ca2+ oscillations. J. Biol. Chem. 2005, 280, 31011–31018. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Kashir, J.; Thanassoulas, A.; Safieh-Garabedian, B.; Lai, F.A.; Nomikos, M. Essential Role of Sperm-Specific PLC-Zeta in Egg Activation and Male Factor Infertility: An Update. Front. Cell Dev. Biol. 2020, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Suh, P.G.; Park, J.I.; Manzoli, L.; Cocco, L.; Peak, J.C.; Katan, M.; Fukami, K.; Kataoka, T.; Yun, S.; Ryu, S.H. Multiple roles of phosphoinositide-specific phospholipase C isozymes. BMB Rep. 2008, 41, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J. Increasing associations between defects in phospholipase C zeta and conditions of male infertility: Not just ICSI failure? J. Assist. Reprod. Genet. 2020, 37, 1273–1293. [Google Scholar] [CrossRef]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLC zeta: A sperm-specific trigger of Ca2+ oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar] [CrossRef]
- Kouchi, Z.; Fukami, K.; Shikano, T.; Oda, S.; Nakamura, Y.; Takenawa, T.; Miyazaki, S. Recombinant Phospholipase Cζ Has High Ca2+ Sensitivity and Induces Ca2+ Oscillations in Mouse Eggs. J. Biol. Chem. 2004, 279, 10408–10412. [Google Scholar] [CrossRef] [PubMed]
- Cox, L. Sperm phospholipase Czeta from humans and cynomolgus monkeys triggers Ca2+ oscillations, activation and development of mouse oocytes. Reproduction 2002, 124, 611–623. [Google Scholar] [CrossRef]
- Kouchi, Z.; Shikano, T.; Nakamura, Y.; Shirakawa, H.; Fukami, K.; Miyazaki, S. The Role of EF-hand Domains and C2 Domain in Regulation of Enzymatic Activity of Phospholipase Cζ. J. Biol. Chem. 2005, 280, 21015–21021. [Google Scholar] [CrossRef]
- Escoffier, J.; Yassine, S.; Lee, H.C.; Martinez, G.; Delaroche, J.; Coutton, C.; Karaouzène, T.; Zouari, R.; Metzler-Guillemain, C.; Pernet-Gallay, K.; et al. Subcellular localization of phospholipase Cζ in human sperm and its absence in DPY19L2-deficient sperm are consistent with its role in oocyte activation. Mol. Hum. Reprod. 2015, 21, 157–168. [Google Scholar] [CrossRef]
- Fujimoto, S.; Yoshida, N.; Fukui, T.; Amanai, M.; Isobe, T.; Itagaki, C.; Izumi, T.; Perry, A.C. Mammalian phospholipase Czeta induces oocyte activation from the sperm perinuclear matrix. Dev. Biol. 2004, 274, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, V.; Kastelic, J.P.; Thundathil, J.C. Ouabain-induced activation of phospholipase C zeta and its contributions to bovine sperm capacitation. Cell Tissue Res. 2021, 385, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Hachem, A.; Godwin, J.; Ruas, M.; Lee, H.C.; Ferrer Buitrago, M.; Ardestani, G.; Bassett, A.; Fox, S.; Navarrete, F.; de Sutter, P.; et al. PLCzeta is the physiological trigger of the Ca2+ oscillations that induce embryogenesis in mammals but conception can occur in its absence. Development 2017, 144, 2914–2924. [Google Scholar] [CrossRef]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-borne phospholipase C zeta-1 ensures monospermic fertilization in mice. Sci. Rep. 2018, 8, 1315. [Google Scholar] [CrossRef] [PubMed]
- Swann, K. The role of Ca2+ in oocyte activation during In Vitro fertilization: Insights into potential therapies for rescuing failed fertilization. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1830–1837. [Google Scholar] [CrossRef]
- Nomikos, M. Novel signalling mechanism and clinical applications of sperm-specific PLCζ. Biochem. Soc. Trans. 2015, 43, 371–376. [Google Scholar] [CrossRef]
- Heytens, E.; Parrington, J.; Coward, K.; Young, C.; Lambrecht, S.; Yoon, S.Y.; Fissore, R.A.; Hamer, R.; Deane, C.M.; Ruas, M.; et al. Reduced amounts and abnormal forms of phospholipase C zeta (PLCzeta) in spermatozoa from infertile men. Hum. Reprod. 2009, 24, 2417–2428. [Google Scholar] [CrossRef]
- Kashir, J.; Jones, C.; Lee, H.C.; Rietdorf, K.; Nikiforaki, D.; Durrans, C.; Ruas, M.; Tee, S.T.; Heindryckx, B.; Galione, A.; et al. Loss of activity mutations in phospholipase C zeta (PLCζ) abolishes calcium oscillatory ability of human recombinant protein in mouse oocytes. Hum. Reprod. 2011, 26, 3372–3387. [Google Scholar] [CrossRef]
- Kashir, J.; Konstantinidis, M.; Jones, C.; Lemmon, B.; Lee, H.C.; Hamer, R.; Heindryckx, B.; Deane, C.M.; De Sutter, P.; Fissore, R.A.; et al. A maternally inherited autosomal point mutation in human phospholipase C zeta (PLCζ) leads to male infertility. Hum. Reprod. 2012, 27, 222–231. [Google Scholar] [CrossRef]
- Kashir, J.; Konstantinidis, M.; Jones, C.; Heindryckx, B.; De Sutter, P.; Parrington, J.; Wells, D.; Coward, K. Characterization of two heterozygous mutations of the oocyte activation factor phospholipase C zeta (PLCζ) from an infertile man by use of minisequencing of individual sperm and expression in somatic cells. Fertil. Steril. 2012, 98, 423–431. [Google Scholar] [CrossRef]
- Theodoridou, M.; Nomikos, M.; Parthimos, D.; Gonzalez-Garcia, J.R.; Elgmati, K.; Calver, B.L.; Sideratou, Z.; Nounesis, G.; Swann, K.; Lai, F.A. Chimeras of sperm PLCζ reveal disparate protein domain functions in the generation of intracellular Ca2+ oscillations in mammalian eggs at fertilization. Mol. Hum. Reprod. 2013, 19, 852–864. [Google Scholar] [CrossRef]
- Escoffier, J.; Lee, H.C.; Yassine, S.; Zouari, R.; Martinez, G.; Karaouzène, T.; Coutton, C.; Kherraf, Z.E.; Halouani, L.; Triki, C.; et al. Homozygous mutation of PLCZ1 leads to defective human oocyte activation and infertility that is not rescued by the WW-binding protein PAWP. Hum. Mol. Genet. 2015, 25, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J. The initiation and regulation of Ca2+ signalling at fertilization in mammals. Semin. Cell Dev. Biol. 2001, 12, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Swann, K.; Saunders, C.M.; Rogers, N.T.; Lai, F.A. PLCzeta(zeta): A sperm protein that triggers Ca2+ oscillations and egg activation in mammals. Semin. Cell Dev. Biol. 2006, 17, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Shikano, T.; Kuroda, K.; Miyazaki, S. Relationship between nuclear sequestration of PLCzeta and termination of PLCzeta-induced Ca2+ oscillations in mouse eggs. Cell Calcium 2008, 44, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Marangos, P.; FitzHarris, G.; Carroll, J. Ca2+ oscillations at fertilization in mammals are regulated by the formation of pronuclei. Development 2003, 130, 1461–1472. [Google Scholar] [CrossRef]
- Larman, M.G.; Saunders, C.M.; Carroll, J.; Lai, F.A.; Swann, K. Cell cycle-dependent Ca2+ oscillations in mouse embryos are regulated by nuclear targeting of PLCzeta. J. Cell Sci. 2004, 117, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Ito, M.; Shikano, T.; Awaji, T.; Yoda, A.; Takeuchi, H.; Kinoshita, K.; Miyazaki, S. The Role of X/Y Linker Region and N-terminal EF-hand Domain in Nuclear Translocation and Ca2+ Oscillation-inducing Activities of Phospholipase Cζ, a Mammalian Egg-activating Factor*. J. Biol. Chem. 2006, 281, 27794–27805. [Google Scholar] [CrossRef]
- Singal, T.; Dhalla, N.S.; Tappia, P.S. Phospholipase C may be involved in norepinephrine-induced cardiac hypertrophy. Biochem. Biophys. Res. Commun. 2004, 320, 1015–1019. [Google Scholar] [CrossRef]
- Yoda, A.; Oda, S.; Shikano, T.; Kouchi, Z.; Awaji, T.; Shirakawa, H.; Kinoshita, K.; Miyazaki, S. Ca2+ oscillation-inducing phospholipase C zeta expressed in mouse eggs is accumulated to the pronucleus during egg activation. Dev. Biol. 2004, 268, 245–257. [Google Scholar] [CrossRef]
- Fissore, R.A.; Dobrinsky, J.R.; Balise, J.J.; Duby, R.T.; Robl, J.M. Patterns of intracellular Ca2+ concentrations in fertilized bovine eggs. Biol. Reprod. 1992, 47, 960–969. [Google Scholar] [CrossRef][Green Version]
- Kline, D.; Kline, J.T. Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse egg. Dev. Biol. 1992, 149, 80–89. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Ito, M.; Kuroda, K.; Takeda, S.; Tanaka, A. The establishment of appropriate methods for egg-activation by human PLCZ1 RNA injection into human oocyte. Cell Calcium 2017, 65, 22–30. [Google Scholar] [CrossRef]
- Ducibella, T.; Huneau, D.; Angelichio, E.; Xu, Z.; Schultz, R.M.; Kopf, G.S.; Fissore, R.; Madoux, S.; Ozil, J.P. Egg-to-embryo transition is driven by differential responses to Ca2+ oscillation number. Dev. Biol. 2002, 250, 280–291. [Google Scholar] [CrossRef]
- Lawrence, Y.; Whitaker, M.; Swann, K. Sperm-egg fusion is the prelude to the initial Ca2+ increase at fertilization in the mouse. Development 1997, 124, 233–241. [Google Scholar] [CrossRef]
- Sone, Y.; Ito, M.; Shirakawa, H.; Shikano, T.; Takeuchi, H.; Kinoshita, K.; Miyazaki, S. Nuclear translocation of phospholipase C-zeta, an egg-activating factor, during early embryonic development. Biochem. Biophys. Res. Commun. 2005, 330, 690–694. [Google Scholar] [CrossRef]
- Yu, Y.; Halet, G.; Lai, F.A.; Swann, K. Regulation of diacylglycerol production and protein kinase C stimulation during sperm- and PLCζ-mediated mouse egg activation. Biol. Cell 2008, 100, 633–643. [Google Scholar] [CrossRef][Green Version]
- Swann, K.; Lai, F.A. PLCζ and the initiation of Ca2+ oscillations in fertilizing mammalian eggs. Cell Calcium 2013, 53, 55–62. [Google Scholar] [CrossRef]
- Yu, Y.; Nomikos, M.; Theodoridou, M.; Nounesis, G.; Lai, F.A.; Swann, K. PLCζ causes Ca2+ oscillations in mouse eggs by targeting intracellular and not plasma membrane PI(4,5)P2. Mol. Biol. Cell 2011, 23, 371–380. [Google Scholar] [CrossRef]
- Satouh, Y.; Ikawa, M. New Insights into the Molecular Events of Mammalian Fertilization. Trends Biochem. Sci. 2018, 43, 818–828. [Google Scholar] [CrossRef]
- Young, C.; Grasa, P.; Coward, K.; Davis, L.C.; Parrington, J. Phospholipase C zeta undergoes dynamic changes in its pattern of localization in sperm during capacitation and the acrosome reaction. Fertil. Steril. 2009, 91, 2230–2242. [Google Scholar] [CrossRef]
- Yoon, S.-Y.; Jellerette, T.; Salicioni, A.M.; Lee, H.C.; Yoo, M.-s.; Coward, K.; Parrington, J.; Grow, D.; Cibelli, J.B.; Visconti, P.E.; et al. Human sperm devoid of PLC, zeta 1 fail to induce Ca2+ release and are unable to initiate the first step of embryo development. J. Clin. Investig. 2008, 118, 3671–3681. [Google Scholar] [CrossRef]
- Grasa, P.; Coward, K.; Young, C.; Parrington, J. The pattern of localization of the putative oocyte activation factor, phospholipase Cζ, in uncapacitated, capacitated, and ionophore-treated human spermatozoa. Hum. Reprod. 2008, 23, 2513–2522. [Google Scholar] [CrossRef]
- Nakai, M.; Ito, J.; Sato, K.-i.; Noguchi, J.; Kaneko, H.; Kashiwazaki, N.; Kikuchi, K. Pre-treatment of sperm reduces success of ICSI in the pig. Reproduction 2011, 142, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Kaewmala, K.; Uddin, M.J.; Cinar, M.U.; Große-Brinkhaus, C.; Jonas, E.; Tesfaye, D.; Phatsara, C.; Tholen, E.; Looft, C.; Schellander, K. Investigation into Association and Expression of PLCz and COX-2 as Candidate Genes for Boar Sperm Quality and Fertility. Reprod. Domest. Anim. 2012, 47, 213–223. [Google Scholar] [CrossRef]
- Bedford-Guaus, S.J.; McPartlin, L.A.; Xie, J.; Westmiller, S.L.; Buffone, M.G.; Roberson, M.S. Molecular Cloning and Characterization of Phospholipase C Zeta in Equine Sperm and Testis Reveals Species-Specific Differences in Expression of Catalytically Active Protein1. Biol. Reprod. 2011, 85, 78–88. [Google Scholar] [CrossRef]
- Kashir, J.; Mistry, B.V.; BuSaleh, L.; Abu-Dawas, R.; Nomikos, M.; Ajlan, A.; Abu-Dawud, R.; AlYacoub, N.; AlHassan, S.; Lai, F.A.; et al. Phospholipase C zeta profiles are indicative of optimal sperm parameters and fertilisation success in patients undergoing fertility treatment. Andrology 2020, 8, 1143–1159. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Meng, X.; Coward, K. SPERM FACTORS AND EGG ACTIVATION: Phospholipase C zeta (PLCZ1) and the clinical diagnosis of oocyte activation deficiency. Reproduction 2022, 164, F53–F66. [Google Scholar] [CrossRef]
- Aarabi, M.; Yu, Y.; Xu, W.; Tse, M.Y.; Pang, S.C.; Yi, Y.-J.; Sutovsky, P.; Oko, R. The testicular and epididymal expression profile of PLCζ in mouse and human does not support its role as a sperm-borne oocyte activating factor. PLoS ONE 2012, 7, e33496. [Google Scholar] [CrossRef]
- Kashir, J.; Jones, C.; Child, T.; Williams, S.A.; Coward, K. Viability Assessment for Artificial Gametes: The Need for Biomarkers of Functional Competency. Biol. Reprod. 2012, 87. [Google Scholar] [CrossRef] [PubMed]
- Nomikos, M.; Thanassoulas, A.; Beck, K.; Theodoridou, M.; Kew, J.; Kashir, J.; Calver, B.L.; Matthews, E.; Rizkallah, P.; Sideratou, Z.; et al. Mutations in PLCδ1 associated with hereditary leukonychia display divergent PIP2 hydrolytic function. FEBS J. 2016, 283, 4502–4514. [Google Scholar] [CrossRef] [PubMed]
- Yaqinuddin, A.; Kvietys, P.; Kashir, J. COVID-19: Role of neutrophil extracellular traps in acute lung injury. Respir. Investig. 2020, 58, 419–420. [Google Scholar] [CrossRef]
- Kashir, J.; Mistry, B.; Gumssani, M.; Rajab, M.; Abu-Dawas, R.; AlMohanna, F.; Nomikos, M.; Jones, C.; Abu-Dawud, R.; Al-Yacoub, N.; et al. Advancing male age differentially alters levels and localization patterns of PLCzeta in sperm and testes from different mouse strains. Asian J. Androl. 2021, 23, 178–187. [Google Scholar] [CrossRef]
- Yelumalai, S.; Kashir, J.; Jones, C.; Bagheri, H.; Oo, S.L.; McLaren, L.; Coward, K. Clinician-induced (iatrogenic) damage incurred during human infertility treatment: Detrimental effects of sperm selection methods and cryopreservation upon the viability, DNA integrity, and function of human sperm. Asian Pac. J. Reprod. 2012, 1, 69–75. [Google Scholar] [CrossRef]
- Kashir, J.; Yelumalai, S.; Jones, C.; Coward, K. Clinician-Induced (Iatrogenic) Damage Incurred during Human Fertility Treatment: Detrimental Effects upon Gamete and Embryo Viability and the Potential for Epigenetic Risk. Hum. Genet. Embryol. 2012. [Google Scholar] [CrossRef]
- Sermondade, N.; Hafhouf, E.; Dupont, C.; Bechoua, S.; Palacios, C.; Eustache, F.; Poncelet, C.; Benzacken, B.; Lévy, R.; Sifer, C. Successful childbirth after intracytoplasmic morphologically selected sperm injection without assisted oocyte activation in a patient with globozoospermia. Hum. Reprod. 2011, 26, 2944–2949. [Google Scholar] [CrossRef][Green Version]
- Meng, X.; Melo, P.; Jones, C.; Ross, C.; Mounce, G.; Turner, K.; Child, T.; Coward, K. Use of phospholipase C zeta analysis to identify candidates for artificial oocyte activation: A case series of clinical pregnancies and a proposed algorithm for patient management. Fertil. Steril. 2020, 114, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Uh, K.; Hay, A.; Chen, P.; Reese, E.; Lee, K. Design of novel oocyte activation methods: The role of zinc. Biol. Reprod. 2022, 106, 264–273. [Google Scholar] [CrossRef]
- Cardona Barberán, A.; Boel, A.; Vanden Meerschaut, F.; Stoop, D.; Heindryckx, B. SPERM FACTORS AND EGG ACTIVATION: Fertilization failure after human ICSI and the clinical potential of PLCZ1. Reproduction 2022, 164, F39–F51. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, K.; Fujikura, Y.; Katayose, H. The present status of artificial oocyte activation in assisted reproductive technology. Reprod. Med. Biol. 2008, 7, 133–142. [Google Scholar] [CrossRef]
- Batista Napotnik, T.; Reberšek, M.; Vernier, P.T.; Mali, B.; Miklavčič, D. Effects of high voltage nanosecond electric pulses on eukaryotic cells (in vitro): A systematic review. Bioelectrochemistry 2016, 110, 1–12. [Google Scholar] [CrossRef]
- Yanagida, K.; Katayose, H.; Yazawa, H.; Kimura, Y.; Sato, A.; Yanagimachi, H.; Yanagimachi, R. Successful fertilization and pregnancy following ICSI and electrical oocyte activation. Hum. Reprod. 1999, 14, 1307–1311. [Google Scholar] [CrossRef]
- Egashira, A.; Murakami, M.; Haigo, K.; Horiuchi, T.; Kuramoto, T. A successful pregnancy and live birth after intracytoplasmic sperm injection with globozoospermic sperm and electrical oocyte activation. Fertil. Steril. 2009, 92, 2037.e2035–2037.e2039. [Google Scholar] [CrossRef] [PubMed]
- Vanden Meerschaut, F.; Nikiforaki, D.; Heindryckx, B.; De Sutter, P. Assisted oocyte activation following ICSI fertilization failure. Reprod. Biomed. Online 2014, 28, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.J.; Jang, G.; Kwon, D.K.; Kang, J.T.; Kwon, O.S.; Park, H.J.; Kang, S.K.; Lee, B.C. Electrical activation induces reactive oxygen species in porcine embryos. Theriogenology 2008, 70, 1111–1118. [Google Scholar] [CrossRef]
- Tesarik, J.; Rienzi, L.; Ubaldi, F.; Mendoza, C.; Greco, E. Use of a modified intracytoplasmic sperm injection technique to overcome sperm-borne and oocyte-borne oocyte activation failures. Fertil. Steril. 2002, 78, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Ebner, T.; Moser, M.; Sommergruber, M.; Jesacher, K.; Tews, G. Complete oocyte activation failure after ICSI can be overcome by a modified injection technique. Hum. Reprod. 2004, 19, 1837–1841. [Google Scholar] [CrossRef]
- Sanusi, R.; Yu, Y.; Nomikos, M.; Lai, F.A.; Swann, K. Rescue of failed oocyte activation after ICSI in a mouse model of male factor infertility by recombinant phospholipase Cζ. Mol. Hum. Reprod. 2015, 21, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tappia, P.S.; Dhalla, N.S. Phospholipases in Health and Disease, 9th ed.; Springer: Berlin/Heidelberg, Germany, 2014; p. 410. [Google Scholar]
- Ebner, T.; Köster, M.; Shebl, O.; Moser, M.; Van der Ven, H.; Tews, G.; Montag, M. Application of a ready-to-use calcium ionophore increases rates of fertilization and pregnancy in severe male factor infertility. Fertil. Steril. 2012, 98, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Ebner, T.; Montag, M.; Montag, M.; Van der Ven, K.; Van der Ven, H.; Ebner, T.; Shebl, O.; Oppelt, P.; Hirchenhain, J.; Krüssel, J.; et al. Live birth after artificial oocyte activation using a ready-to-use ionophore: A prospective multicentre study. Reprod. Biomed. Online 2015, 30, 359–365. [Google Scholar] [CrossRef]
- Ebner, T.; Montag, M. Artificial oocyte activation: Evidence for clinical readiness. Reprod. Biomed. Online 2016, 32, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, S.; Aksünger, Ö.; Ata, C.; Sağıroglu, Y.; Keskin, İ. Artificial oocyte activation with calcium ionophore for frozen sperm cycles. Syst. Biol. Reprod. Med. 2018, 64, 381–388. [Google Scholar] [CrossRef]
- Martínez, M.; Durban, M.; Santaló, J.; Rodríguez, A.; Vassena, R. Assisted oocyte activation effects on the morphokinetic pattern of derived embryos. J. Assist. Reprod. Genet. 2021, 38, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Bae, I.H.; Kim, H.J.; Jang, J.M.; Hur, Y.S.; Kim, H.K.; Yoon, S.H.; Lee, W.D.; Lim, J.H. Analysis of clinical outcomes with respect to spermatozoan origin after artificial oocyte activation with a calcium ionophore. J. Assist. Reprod. Genet. 2013, 30, 1569–1575. [Google Scholar] [CrossRef][Green Version]
- Deemeh, M.R.; Tavalaee, M.; Nasr-Esfahani, M.H. Health of Children Born Through Artificial Oocyte Activation. Reprod. Sci. 2015, 22, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.; Biron-Shental, T.; Sukenik-Halevy, R.; Klement, A.H.; Sharony, R.; Berkovitz, A. Oocyte activation by calcium ionophore and congenital birth defects: A retrospective cohort study. Fertil. Steril. 2016, 106, 590–596.e592. [Google Scholar] [CrossRef]
- Economou, K.A.; Christopikou, D.; Tsorva, E.; Davies, S.; Mastrominas, M.; Cazlaris, H.; Koutsilieris, M.; Angelogianni, P.; Loutradis, D. The combination of calcium ionophore A23187 and GM-CSF can safely salvage aged human unfertilized oocytes after ICSI. J. Assist. Reprod. Genet. 2017, 34, 33–41. [Google Scholar] [CrossRef]
- Cardona Barberán, A.; Boel, A.; Vanden Meerschaut, F.; Stoop, D.; Heindryckx, B. Diagnosis and Treatment of Male Infertility-Related Fertilization Failure. J. Clin. Med. 2020, 9, 3899. [Google Scholar] [CrossRef]
- Jones, K.T. Mammalian egg activation: From Ca2+ spiking to cell cycle progression. Reproduction 2005, 130, 813–823. [Google Scholar] [CrossRef]
- Collas, P.; Chang, T.; Long, C.; Robl, J.M. Inactivation of histone H1 kinase by Ca2+ in rabbit oocytes. Mol. Reprod Dev. 1995, 40, 253–258. [Google Scholar] [CrossRef]
- Bos-Mikich, A.; Swann, K.; Whittingham, D.G. Calcium oscillations and protein synthesis inhibition synergistically activate mouse oocytes. Mol. Reprod Dev. 1995, 41, 84–90. [Google Scholar] [CrossRef]
- Swann, K.; Ozil, J.-P. Dynamics of the Calcium Signal That Triggers Mammalian Egg Activation. Int. Rev. Cytol. 1994, 183–222. [Google Scholar] [CrossRef]
- Tsai, T.-E.; Lin, P.-H.; Lian, P.-F.; Li, C.-J.; Vitale, S.G.; Mikuš, M.; Su, W.-P.; Tsai, H.-W.; Tsui, K.-H.; Lin, L.-T. Artificial oocyte activation may improve embryo quality in older patients with diminished ovarian reserve undergoing IVF-ICSI cycles. J. Ovarian Res. 2022, 15, 102. [Google Scholar] [CrossRef]
- Swann, K.; Windsor, S.; Campbell, K.; Elgmati, K.; Nomikos, M.; Zernicka-Goetz, M.; Amso, N.; Lai, F.A.; Thomas, A.; Graham, C. Phospholipase C-ζ-induced Ca2+ oscillations cause coincident cytoplasmic movements in human oocytes that failed to fertilize after intracytoplasmic sperm injection. Fertil. Steril. 2012, 97, 742–747. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, S.K.; Kim, J.; Kim, J.H.; Chang, J.H.; Jee, B.C.; Kim, S.H. Relationship between phospholipase C zeta immunoreactivity and DNA fragmentation and oxidation in human sperm. Obstet. Gynecol. Sci. 2015, 58, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Tavalaee, M.; Kiani-Esfahani, A.; Nasr-Esfahani, M.H. Relationship between phospholipase C-zeta, semen parameters, and chromatin status. Syst. Biol. Reprod. Med. 2017, 63, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Tavalaee, M.; Kiani-Esfahani, A.; Nasr-Esfahani, M.H. Relationship between Potential Sperm Factors Involved in Oocyte Activation and Sperm DNA Fragmentation with Intra-Cytoplasmic Sperm Injection Clinical Outcomes. Cell J. 2017, 18, 588–596. [Google Scholar] [CrossRef]
- Kashir, J.; Heindryckx, B.; Jones, C.; De Sutter, P.; Parrington, J.; Coward, K. Oocyte activation, phospholipase C zeta and human infertility. Hum. Reprod. Update 2010, 16, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Vaquer, A.; Barragan, M.; Freour, T.; Vernaeve, V.; Vassena, R. PLCζ sequence, protein levels, and distribution in human sperm do not correlate with semen characteristics and fertilization rates after ICSI. J. Assist. Reprod. Genet. 2016, 33, 747–756. [Google Scholar] [CrossRef]
- Torra-Massana, M.; Cornet-Bartolomé, D.; Barragán, M.; Durban, M.; Ferrer-Vaquer, A.; Zambelli, F.; Rodriguez, A.; Oliva, R.; Vassena, R. Novel phospholipase C zeta 1 mutations associated with fertilization failures after ICSI. Hum. Reprod. 2019, 34, 1494–1504. [Google Scholar] [CrossRef]
- Azad, N.; Nazarian, H.; Ghaffari Novin, M.; Masteri Farahani, R.; Piryaei, A.; Heidari, M.H.; Abdollahpour Alitappeh, M. Oligoasthenoteratozoospermic (OAT) men display altered phospholipase C ζ (PLCζ) localization and a lower percentage of sperm cells expressing PLCζ and post-acrosomal sheath WW domain-binding protein (PAWP). Bosn. J. Basic Med. Sci. 2018, 18, 178–184. [Google Scholar] [CrossRef]
- Azad, N.; Nazarian, H.; Ghaffari Novin, M.; Masteri Farahani, R.; Piryaei, A.; Heidari, M.H. Phospholipase C zeta parameters in sperm from polymorphic teratozoospermic men. Ann. Anat.-Anat. Anz. 2018, 215, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Rahimizadeh, P.; Topraggaleh, T.R.; Nasr-Esfahani, M.H.; Ziarati, N.; Mirshahvaladi, S.; Esmaeili, V.; Seifi, S.; Eftekhari-Yazdi, P.; Shahverdi, A. The alteration of PLCζ protein expression in unexplained infertile and asthenoteratozoospermic patients: A potential effect on sperm fertilization ability. Mol. Reprod. Dev. 2019, 87, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Janghorban-Laricheh, E.; Ghazavi-Khorasgani, N.; Tavalaee, M.; Zohrabi, D.; Abbasi, H.; Nasr- Esfahani, M.H. An association between sperm PLCζ levels and varicocele? J. Assist. Reprod. Genet. 2016, 33, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.L.; Yoon, S.Y.; Morshedi, M.S.; Lacey, D.R.; Jellerette, T.; Fissore, R.A.; Oehninger, S. Complete globozoospermia associated with PLCζ deficiency treated with calcium ionophore and ICSI results in pregnancy. Reprod. Biomed. Online 2010, 20, 559–564. [Google Scholar] [CrossRef]
- Kamali-Dolat Abadi, M.; Tavalaee, M.; Shahverdi, A.; Nasr-Esfahani, M.H. Evaluation of PLCζ and PAWP Expression in Globozoospermic Individuals. Cell J. 2016, 18, 438–445. [Google Scholar] [CrossRef]
- Tejera, A.; Mollá, M.; Muriel, L.; Remohí, J.; Pellicer, A.; De Pablo, J.L. Successful pregnancy and childbirth after intracytoplasmic sperm injection with calcium ionophore oocyte activation in a globozoospermic patient. Fertil. Steril. 2008, 90, 1202.e1201–1202.e1205. [Google Scholar] [CrossRef]
- Aghajanpour, S.; Ghaedi, K.; Salamian, A.; Deemeh, M.R.; Tavalaee, M.; Moshtaghian, J.; Parrington, J.; Nasr-Esfahani, M.H. Quantitative expression of phospholipase C zeta, as an index to assess fertilization potential of a semen sample. Hum. Reprod. 2011, 26, 2950–2956. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Sermondade, N.; Sifer, C.; Oo, S.L.; Jones, C.; Mounce, G.; Turner, K.; Child, T.; McVeigh, E.; Coward, K. Motile sperm organelle morphology evaluation-selected globozoospermic human sperm with an acrosomal bud exhibits novel patterns and higher levels of phospholipase C zeta. Hum. Reprod. 2012, 27, 3150–3160. [Google Scholar] [CrossRef]
- Yeste, M.; Jones, C.; Amdani, S.N.; Patel, S.; Coward, K. Oocyte activation deficiency: A role for an oocyte contribution? Hum. Reprod. Update 2016, 22, 23–47. [Google Scholar] [CrossRef]
- Kashir, J.; Mistry, B.V.; BuSaleh, L.; Nomikos, M.; Almuqayyil, S.; Abu-Dawud, R.; AlYacoub, N.; Hamdan, H.; AlHassan, S.; Lai, F.A.; et al. Antigen Unmasking Is Required to Clinically Assess Levels and Localisation Patterns of Phospholipase C Zeta in Human Sperm. Pharmaceuticals 2023, 16, 198. [Google Scholar] [CrossRef]
- Nikiforaki, D.; Vanden Meerschaut, F.; De Gheselle, S.; Qian, C.; Van den Abbeel, E.; De Vos, W.H.; Deroo, T.; De Sutter, P.; Heindryckx, B. Sperm involved in recurrent partial hydatidiform moles cannot induce the normal pattern of calcium oscillations. Fertil. Steril. 2014, 102, 581–588.e581. [Google Scholar] [CrossRef]
- Uh, K.; Ryu, J.; Zhang, L.; Errington, J.; Machaty, Z.; Lee, K. Development of novel oocyte activation approaches using Zn2+ chelators in pigs. Theriogenology 2019, 125, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Yelumalai, S.; Yeste, M.; Jones, C.; Amdani, S.N.; Kashir, J.; Mounce, G.; Da Silva, S.J.M.; Barratt, C.L.; McVeigh, E.; Coward, K. Total levels, localization patterns, and proportions of sperm exhibiting phospholipase C zeta are significantly correlated with fertilization rates after intracytoplasmic sperm injection. Fertil. Steril. 2015, 104, 561–568.e564. [Google Scholar] [CrossRef]
- Kashir, J.; Jones, C.; Mounce, G.; Ramadan, W.M.; Lemmon, B.; Heindryckx, B.; de Sutter, P.; Parrington, J.; Turner, K.; Child, T.; et al. Variance in total levels of phospholipase C zeta (PLC-ζ) in human sperm may limit the applicability of quantitative immunofluorescent analysis as a diagnostic indicator of oocyte activation capability. Fertil. Steril. 2013, 99, 107–117.e103. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.; Xie, P.; Parrella, A.; Keating, D.; Rosenwaks, Z.; Palermo, G.D. Identification and treatment of men with phospholipase Cζ–defective spermatozoa. Fertil. Steril. 2020, 114, 535–544. [Google Scholar] [CrossRef]
- Jones, K.T. Mammalian sperm contain two factors for calcium release and egg activation: Phospholipase C zeta and a cryptic activating factor. Mol. Hum. Reprod. 2018, 24, 465–468. [Google Scholar] [CrossRef]
- Xie, X.; Percipalle, P. An actin-based nucleoskeleton involved in gene regulation and genome organization. Biochem. Biophys. Res. Commun. 2018, 506, 378–386. [Google Scholar] [CrossRef]
- Lange, K. Microvillar Ca++ signaling: A new view of an old problem. J. Cell. Physiol. 1999, 180, 19–34. [Google Scholar] [CrossRef]
- Lange, K. Fundamental role of microvilli in the main functions of differentiated cells: Outline of an universal regulating and signaling system at the cell periphery. J. Cell. Physiol. 2011, 226, 896–927. [Google Scholar] [CrossRef]
- Janmey, P.A.; Bucki, R.; Radhakrishnan, R. Regulation of actin assembly by PI(4,5)P2 and other inositol phospholipids: An update on possible mechanisms. Biochem. Biophys. Res. Commun. 2018, 506, 307–314. [Google Scholar] [CrossRef]
- Lim, D.; Lange, K.; Santella, L. Activation of oocytes by latrunculin A. FASEB J. 2002, 16, 1050–1056. [Google Scholar] [CrossRef]
- Moccia, F. Latrunculin A depolarizes starfish oocytes. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 148, 845–852. [Google Scholar] [CrossRef]
- Vasilev, F.; Limatola, N.; Park, D.R.; Kim, U.H.; Santella, L.; Chun, J.T. Disassembly of Subplasmalemmal Actin Filaments Induces Cytosolic Ca2+ Increases in Astropecten aranciacus Eggs. Cell. Physiol. Biochem. 2018, 48, 2011–2034. [Google Scholar] [CrossRef] [PubMed]
- Santella, L.; Limatola, N.; Chun, J.T. Cellular and molecular aspects of oocyte maturation and fertilization: A perspective from the actin cytoskeleton. Zool. Lett. 2020, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- York-Andersen, A.H.; Hu, Q.; Wood, B.W.; Wolfner, M.F.; Weil, T.T. A calcium-mediated actin redistribution at egg activation in Drosophila. Mol. Reprod. Dev. 2020, 87, 293–304. [Google Scholar] [CrossRef]
- Chun, J.T.; Limatola, N.; Vasilev, F.; Santella, L. Early events of fertilization in sea urchin eggs are sensitive to actin-binding organic molecules. Biochem. Biophys. Res. Commun. 2014, 450, 1166–1174. [Google Scholar] [CrossRef]
- Yonemura, S.; Mabuchi, I. Wave of cortical actin polymerization in the sea urchin egg. Cell Motil. Cytoskelet. 1987, 7, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, Y.; Mabuchi, I. Effects of phalloidin microinjection and localization of fluorescein-labeled phalloidin in living sand dollar eggs. Cell Motil. 1982, 2, 103–113. [Google Scholar] [CrossRef]
- Terasaki, M. Actin filament translocations in sea urchin eggs. Cell Motil. Cytoskelet. 1996, 34, 48–56. [Google Scholar] [CrossRef]
- Puppo, A.; Chun, J.T.; Gragnaniello, G.; Garante, E.; Santella, L. Alteration of the cortical actin cytoskeleton deregulates Ca2+ signaling, monospermic fertilization, and sperm entry. PLoS ONE 2008, 3, e3588. [Google Scholar] [CrossRef]
- Santella, L.; Limatola, N.; Chun, J.T. Calcium and actin in the saga of awakening oocytes. Biochem. Biophys. Res. Commun. 2015, 460, 104–113. [Google Scholar] [CrossRef]
- Limatola, N.; Vasilev, F.; Chun, J.T.; Santella, L. Altered actin cytoskeleton in ageing eggs of starfish affects fertilization process. Exp. Cell Res. 2019, 381, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K. The role of calcium in cell division. Cell Calcium 1994, 16, 322–330. [Google Scholar] [CrossRef]
- Ajduk, A.; Ilozue, T.; Windsor, S.; Yu, Y.; Seres, K.B.; Bomphrey, R.J.; Tom, B.D.; Swann, K.; Thomas, A.; Graham, C.; et al. Rhythmic actomyosin-driven contractions induced by sperm entry predict mammalian embryo viability. Nat. Commun. 2011, 2, 417. [Google Scholar] [CrossRef] [PubMed]
- Coskun, S.; Maddirevula, S.; Awartani, K.; Aldeery, M.; Qubbaj, W.; Kashir, J.; Alkuraya, F.S. Recurrent spontaneous oocyte activation causes female infertility. J. Assist. Reprod. Genet. 2022, 39, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.S.; Buck, W.R. Sources of calcium in sea urchin eggs during the fertilization response. Dev. Biol. 1993, 157, 157–169. [Google Scholar] [CrossRef]
- Miyazaki, S.I.; Ohmori, H.; Sasaki, S. Action potential and non-linear current-voltage relation in starfish oocytes. J. Physiol. 1975, 246, 37–54. [Google Scholar] [CrossRef]
- Vasilev, F.; Limatola, N.; Chun, J.T.; Santella, L. Contributions of suboolemmal acidic vesicles and microvilli to the intracellular Ca2+ increase in the sea urchin eggs at fertilization. Int. J. Biol. Sci. 2019, 15, 757–775. [Google Scholar] [CrossRef]
- Galione, A.; McDougall, A.; Busa, W.B.; Willmott, N.; Gillot, I.; Whitaker, M. Redundant mechanisms of calcium-induced calcium release underlying calcium waves during fertilization of sea urchin eggs. Science 1993, 261, 348–352. [Google Scholar] [CrossRef]
- Lee, H.C.; Aarhus, R.; Walseth, T.F. Calcium mobilization by dual receptors during fertilization of sea urchin eggs. Science 1993, 261, 352–355. [Google Scholar] [CrossRef]
- Chini, E.N.; Beers, K.W.; Dousa, T.P. Nicotinate adenine dinucleotide phosphate (NAADP) triggers a specific calcium release system in sea urchin eggs. J. Biol. Chem. 1995, 270, 3216–3223. [Google Scholar] [CrossRef]
- Lim, D.; Kyozuka, K.; Gragnaniello, G.; Carafoli, E.; Santella, L. NAADP+ initiates the Ca2+ response during fertilization of starfish oocytes. FASEB J. 2001, 15, 2257–2267. [Google Scholar] [CrossRef]
- Moccia, F.; Lim, D.; Kyozuka, K.; Santella, L. NAADP triggers the fertilization potential in starfish oocytes. Cell Calcium 2004, 36, 515–524. [Google Scholar] [CrossRef]
- Kaneuchi, T.; Sartain, C.V.; Takeo, S.; Horner, V.L.; Buehner, N.A.; Aigaki, T.; Wolfner, M.F. Calcium waves occur as Drosophila oocytes activate. Proc. Natl. Acad. Sci. USA 2015, 112, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Wolfner, M.F. The Drosophila Trpm channel mediates calcium influx during egg activation. Proc. Natl. Acad. Sci. USA 2019, 116, 18994–19000. [Google Scholar] [CrossRef]
- Lee, K.W.; Webb, S.E.; Miller, A.L. A wave of free cytosolic calcium traverses zebrafish eggs on activation. Dev. Biol. 1999, 214, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.L.; Hertzler, P.L.; Clark, W.H., Jr. Extracellular Mg2+ induces an intracellular Ca2+ wave during oocyte activation in the marine shrimp Sicyonia ingentis. Dev. Biol. 1992, 152, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Limatola, N.; Chun, J.T.; Santella, L. Effects of Salinity and pH of Seawater on the Reproduction of the Sea Urchin Paracentrotus lividus. Biol. Bull. 2020, 239, 13–23. [Google Scholar] [CrossRef]
- York-Andersen, A.H.; Wood, B.W.; Wilby, E.L.; Berry, A.S.; Weil, T.T. Osmolarity-regulated swelling initiates egg activation in Drosophila. Open Biol. 2021, 11, 210067. [Google Scholar] [CrossRef]
- Duncan, F.E.; Que, E.L.; Zhang, N.; Feinberg, E.C.; O’Halloran, T.V.; Woodruff, T.K. The zinc spark is an inorganic signature of human egg activation. Sci. Rep. 2016, 6, 24737. [Google Scholar] [CrossRef]
- Madgwick, S.; Hansen, D.V.; Levasseur, M.; Jackson, P.K.; Jones, K.T. Mouse Emi2 is required to enter meiosis II by reestablishing cyclin B1 during interkinesis. J. Cell Biol. 2006, 174, 791–801. [Google Scholar] [CrossRef]
- Kim, A.M.; Bernhardt, M.L.; Kong, B.Y.; Ahn, R.W.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc sparks are triggered by fertilization and facilitate cell cycle resumption in mammalian eggs. ACS Chem. Biol. 2011, 6, 716–723. [Google Scholar] [CrossRef]
- Que, E.L.; Bleher, R.; Duncan, F.E.; Kong, B.Y.; Gleber, S.C.; Vogt, S.; Chen, S.; Garwin, S.A.; Bayer, A.R.; Dravid, V.P.; et al. Quantitative mapping of zinc fluxes in the mammalian egg reveals the origin of fertilization-induced zinc sparks. Nat. Chem. 2015, 7, 130–139. [Google Scholar] [CrossRef]
- Que, E.L.; Duncan, F.E.; Lee, H.C.; Hornick, J.E.; Vogt, S.; Fissore, R.A.; O’Halloran, T.V.; Woodruff, T.K. Bovine eggs release zinc in response to parthenogenetic and sperm-induced egg activation. Theriogenology 2019, 127, 41–48. [Google Scholar] [CrossRef]
- Zhao, M.H.; Kim, N.H.; Cui, X.S. Zinc depletion activates porcine metaphase II oocytes independently of the protein kinase C pathway. Vitr. Cell Dev. Biol. Anim. 2014, 50, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.Y.; Bernhardt, M.L.; Kim, A.M.; O’Halloran, T.V.; Woodruff, T.K. Zinc maintains prophase I arrest in mouse oocytes through regulation of the MOS-MAPK pathway. Biol. Reprod. 2012, 87, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, M.L.; Kim, A.M.; O’Halloran, T.V.; Woodruff, T.K. Zinc requirement during meiosis I-meiosis II transition in mouse oocytes is independent of the MOS-MAPK pathway. Biol. Reprod. 2011, 84, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Duncan, F.E.; Nowakowski, A.B.; Antipova, O.A.; Woodruff, T.K.; O’Halloran, T.V.; Wolfner, M.F. Zinc Dynamics during Drosophila Oocyte Maturation and Egg Activation. iScience 2020, 23, 101275. [Google Scholar] [CrossRef] [PubMed]
- Converse, A.; Thomas, P. The zinc transporter ZIP9 (Slc39a9) regulates zinc dynamics essential to egg activation in zebrafish. Sci. Rep. 2020, 10, 15673. [Google Scholar] [CrossRef]
- Kim, A.M.; Vogt, S.; O’Halloran, T.V.; Woodruff, T.K. Zinc availability regulates exit from meiosis in maturing mammalian oocytes. Nat. Chem. Biol. 2010, 6, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Lisle, R.S.; Anthony, K.; Randall, M.A.; Diaz, F.J. Oocyte-cumulus cell interactions regulate free intracellular zinc in mouse oocytes. Reproduction 2013, 145, 381–390. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, N.; Duncan, F.E.; Que, E.L.; O’Halloran, T.V.; Woodruff, T.K. The fertilization-induced zinc spark is a novel biomarker of mouse embryo quality and early development. Sci. Rep. 2016, 6, 22772. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, A.; Terakawa, J.; Ito, J.; Kashiwazaki, N. Roles of zinc signaling in mammalian reproduction. Met. Res. 2022, 2, rev-64–rev-73. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdulsamad, H.M.R.; Murtaza, Z.F.; AlMuhairi, H.M.; Bafleh, W.S.; AlMansoori, S.A.; AlQubaisi, S.A.; Hamdan, H.; Kashir, J. The Therapeutic and Diagnostic Potential of Phospholipase C Zeta, Oocyte Activation, and Calcium in Treating Human Infertility. Pharmaceuticals 2023, 16, 441. https://doi.org/10.3390/ph16030441
Abdulsamad HMR, Murtaza ZF, AlMuhairi HM, Bafleh WS, AlMansoori SA, AlQubaisi SA, Hamdan H, Kashir J. The Therapeutic and Diagnostic Potential of Phospholipase C Zeta, Oocyte Activation, and Calcium in Treating Human Infertility. Pharmaceuticals. 2023; 16(3):441. https://doi.org/10.3390/ph16030441
Chicago/Turabian StyleAbdulsamad, Haia M. R., Zoha F. Murtaza, Hessa M. AlMuhairi, Wjdan S. Bafleh, Salma A. AlMansoori, Shaikha A. AlQubaisi, Hamdan Hamdan, and Junaid Kashir. 2023. "The Therapeutic and Diagnostic Potential of Phospholipase C Zeta, Oocyte Activation, and Calcium in Treating Human Infertility" Pharmaceuticals 16, no. 3: 441. https://doi.org/10.3390/ph16030441
APA StyleAbdulsamad, H. M. R., Murtaza, Z. F., AlMuhairi, H. M., Bafleh, W. S., AlMansoori, S. A., AlQubaisi, S. A., Hamdan, H., & Kashir, J. (2023). The Therapeutic and Diagnostic Potential of Phospholipase C Zeta, Oocyte Activation, and Calcium in Treating Human Infertility. Pharmaceuticals, 16(3), 441. https://doi.org/10.3390/ph16030441