
Differential Physiological Responses of Portuguese Bread Wheat (Triticum aestivum L.) Genotypes under Aluminium Stress

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Material and Growth Conditions
2.2. Root Regrowth Measurement
2.3. Determination of Al Accumulation in Roots
2.4. Assessment of Plasma Membrane Integrity Loss and Cell Lipidic Peroxidation
2.5. Callose Detection
2.6. Extraction and Quantification of OAs from Intact Roots
2.7. Statistical Analysis
3. Results
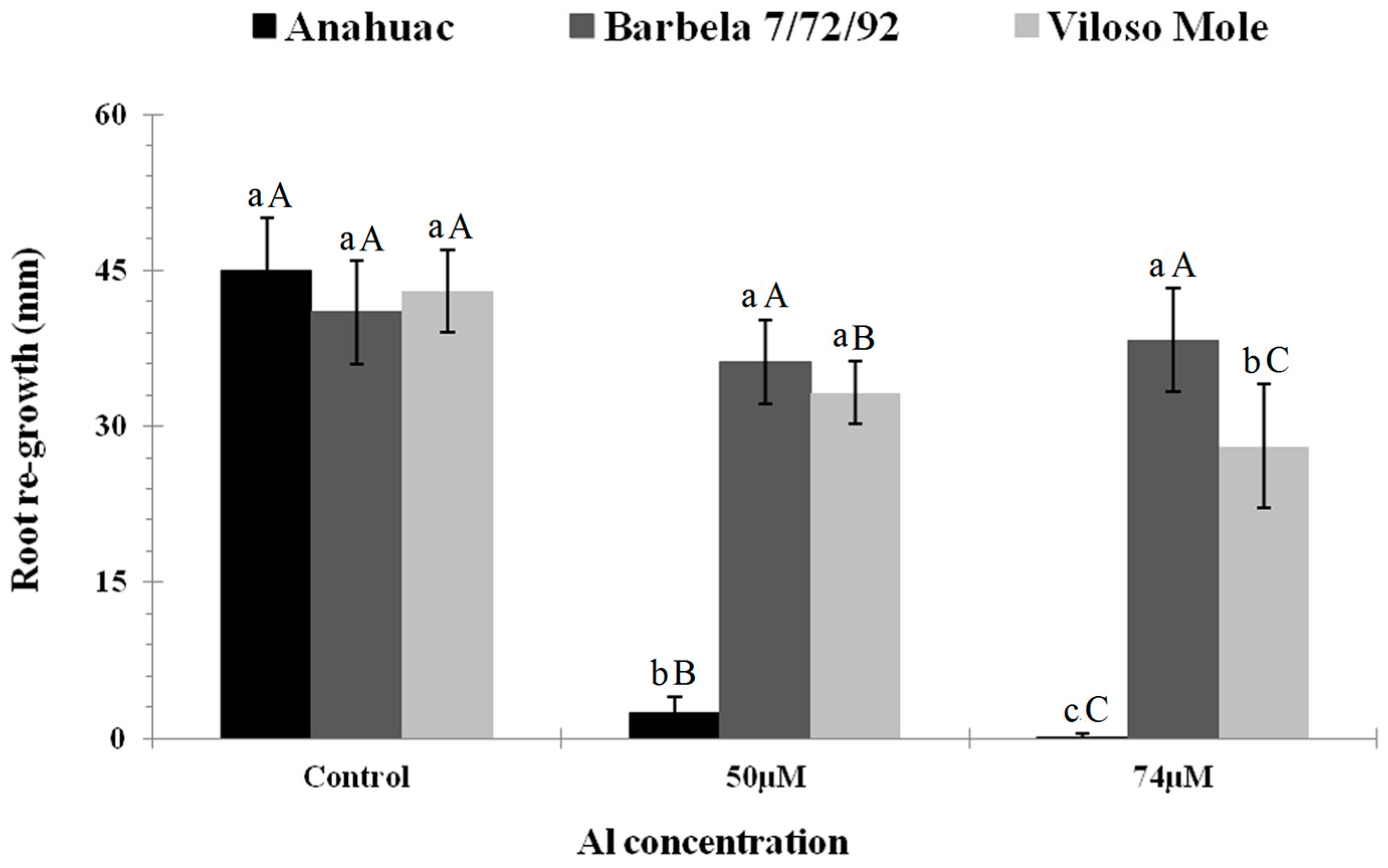
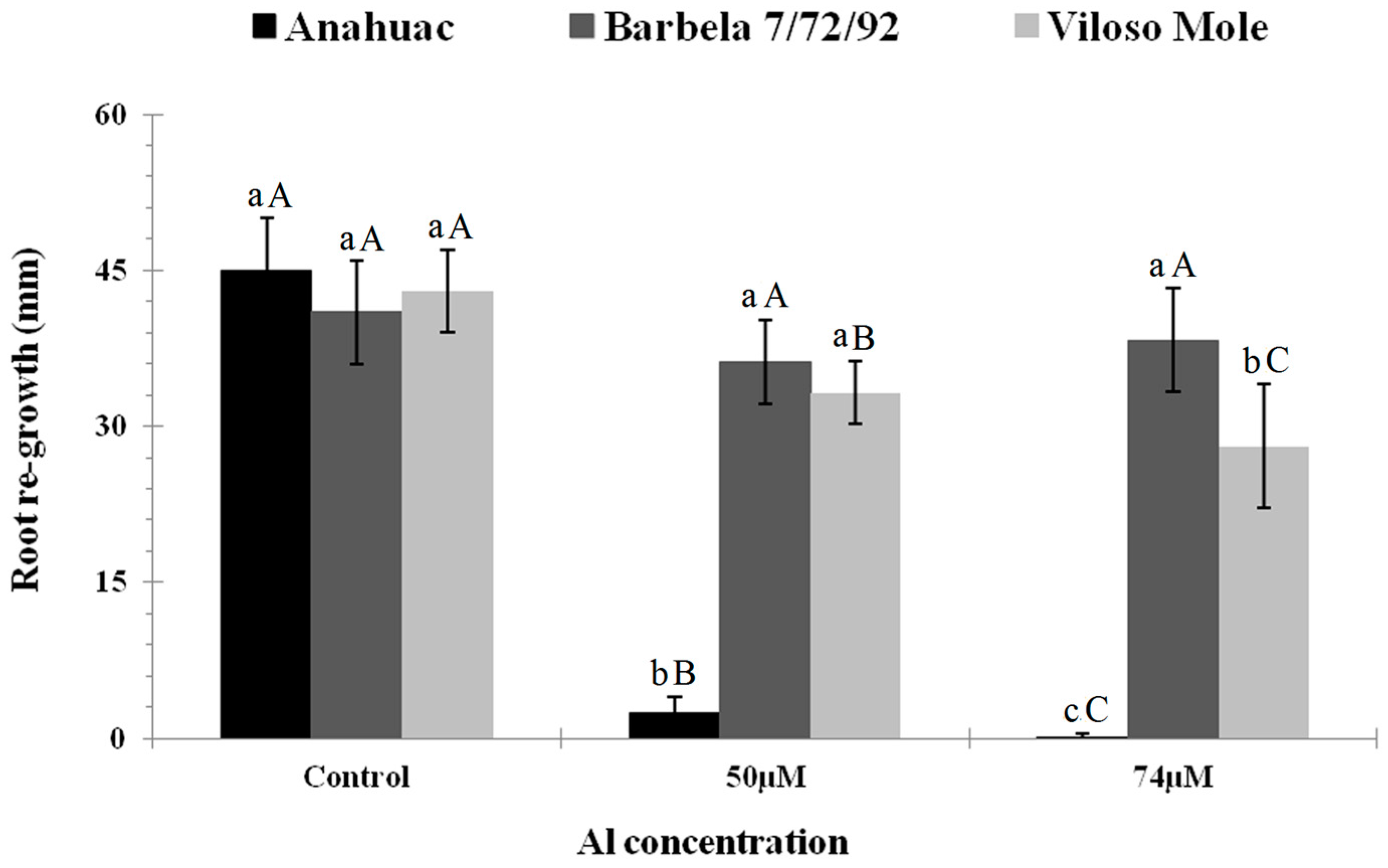
3.1. Root Morphological Response to Al Exposure
3.2. Al Accumulation
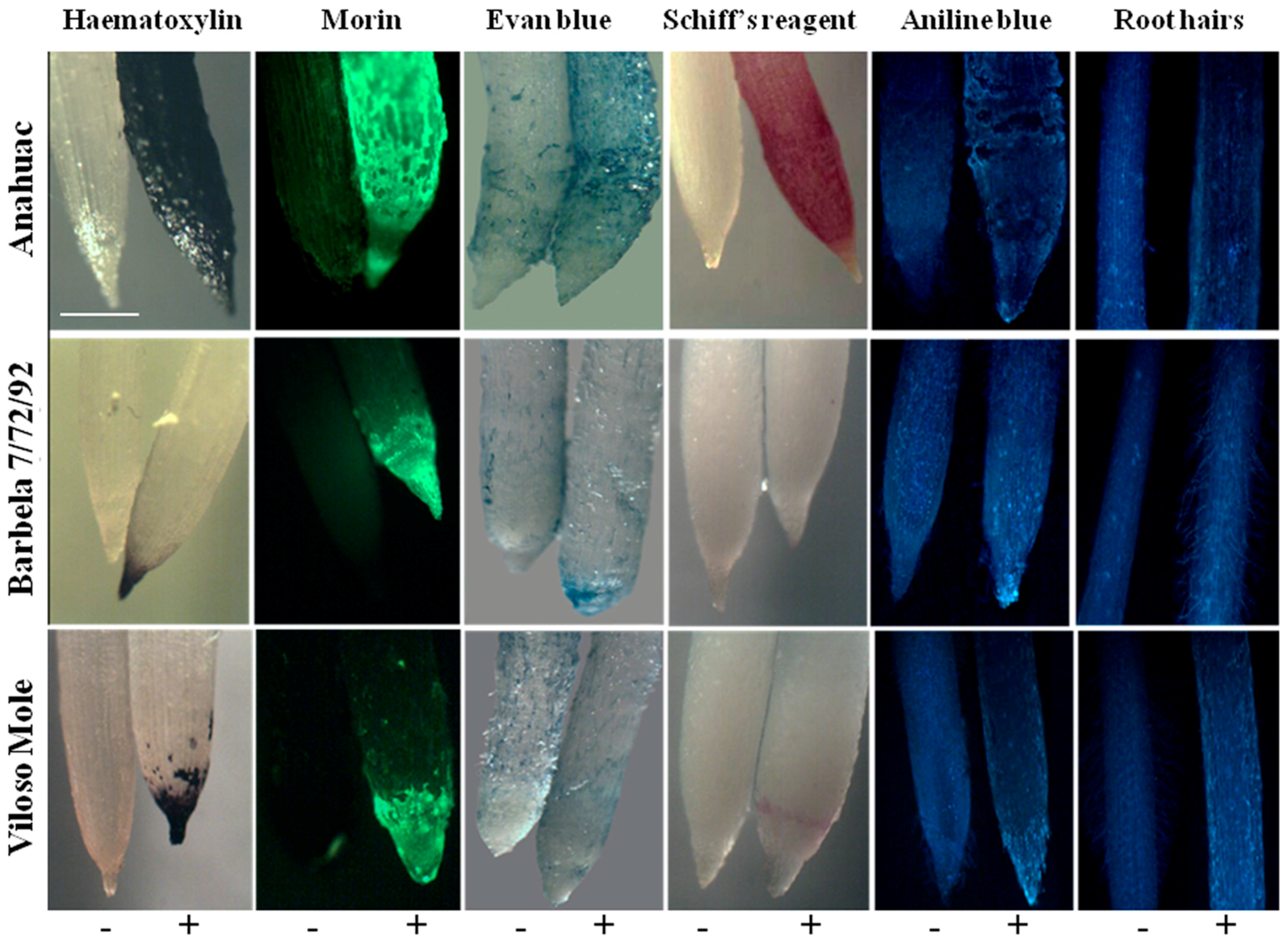
3.3. Plasma Membrane Integrity
3.4. Lipid Peroxidation and Callose Formation under Al Accumulation
3.5. Root Hair Development
3.6. Pattern and Level of OAs in Root Exudates
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eswaran, H.; Reich, P.; Beinroth, F. Global distribution of soils with acidity. In Plant-Soil Interactions at Low pH; Moniz, A.Z., Furlani, A.M.C., Schaffert, R.E., Fageria, N.K., Rosolem, C.A., Cantarella, H., Eds.; Brazilian Soil Science Society: Viçosa, Brazil, 1997; pp. 159–164. [Google Scholar]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Barceló, J.; Poschenrieder, C. Fast root growth responses, root exudates, and internal detoxification as clues to the mechanisms of aluminium toxicity and resistance: A review. Environ. Exp. Bot. 2002, 48, 75–92. [Google Scholar] [CrossRef]
- Ma, J.F.; Furukawa, J. Recent progress in the research of external Al detoxification in higher plants: A minireview. J. Inorg. Biochem. 2003, 97, 46–51. [Google Scholar] [CrossRef]
- Yang, L.T.; Qi, Y.P.; Jiang, H.X.; Chen, L.S. Roles of organic acid anion secretion in aluminium tolerance of higher plants. BioMed. Res. Int. 2013, 2013, 173682. [Google Scholar] [CrossRef] [PubMed]
- Delhaize, E.; James, R.A.; Ryan, P.R. Aluminium tolerance of root hairs underlies genotypic differences in rhizosheath size of wheat (Triticum aestivum) grown on acid soil. New Phytol. 2012, 195, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveira, A.L.; Chander, S.; Barcelo, J.; Poschenrieder, C. Aluminium stress in crop plants. In Recent Advances in Plant Stress Physiology; Yadav, P., Kumar, S., Jain, V., Eds.; Daya Publishing House: New Delhi, India, 2016; pp. 237–263. [Google Scholar]
- Gahoonia, T.S.; Care, D.; Nielsen, N.E. Root hairs and phosphorus acquisition of wheat and barley cultivars. Plant Soil. 1997, 191, 181–188. [Google Scholar] [CrossRef]
- Pinto-Carnide, O.; Guedes-Pinto, H. Aluminum tolerance variability in rye and wheat Portuguese germplasm. Genet. Resour. Crop Evol. 1999, 46, 81–85. [Google Scholar] [CrossRef]
- Martins-Lopes, P.; Maçãs, B.; Guedes-Pinto, H. Portuguese bread wheat germplasm evaluation for aluminium tolerance. Cereal Res. Commun. 2009, 37, 179–188. [Google Scholar] [CrossRef]
- Delhaize, E.; Craig, S.; Beaton, C.D.; Bennet, R.J.; Jagadish, V.C.; Randall, P.J. Aluminum tolerance in wheat (Triticum aestivum L.). I. Uptake and distribution of aluminium in root apices. Plant Physiol. 1993, 103, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.P., Jr.; Carver, B.F.; Baligar, V.C. Expression of aluminium tolerance transferred from Atlas66 to hard winter wheat. Crop Sci. 1997, 37, 103–108. [Google Scholar] [CrossRef]
- Melo, J.O.; Lana, U.G.P.; Piñeros, M.A.; Alves, V.M.C.; Guimarães, C.T.; Liu, J.; Zheng, Y.; Zhong, S.; Fei, Z.; Maron, L.G.; et al. Incomplete transfer of accessory loci influencing SbMATE expression underlies genetic background effects for aluminum tolerance in sorghum. Plant J. 2013, 63, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveira, A.L.; Benito, C.; Prieto, P.; de Andrade Menezes, R.; Rodrigues-Pousada, C.; Guedes-Pinto, H.; Martins-Lopes, P. Molecular characterization of TaSTOP1 homoeologues and their response to aluminium and proton (H+) toxicity in bread wheat (Triticum aestivum L.). BMC Plant Biol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Polle, E.; Konzak, C.F.; Kittrick, J.A. Visual detection of aluminium tolerance levels in wheat by hematoxylin staining of seedling roots. Crop Sci. 1978, 18, 823–827. [Google Scholar] [CrossRef]
- Tice, K.R.; Parker, D.R.; DeMason, D.A. Operationally defined apoplastic and symplastic aluminum fractions in root tips of aluminum-intoxicated wheat. Plant Physiol. 1992, 100, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid peroxidation is an early symptom triggered by aluminum, but not the primary cause of elongation inhibition in pea roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Mano, J.; Wang, S.; Tsuji, W.; Tanaka, K. The involvement of lipid peroxide-derived aldehydes in aluminium toxicity of tobacco roots. Plant Physiol. 2010, 152, 1406–1417. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Blancaflor, E.B.; Kochian, L.V.; Gilroy, S. Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant Cell Environ. 2006, 29, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Delhaize, E.; Ryan, P.R.; Hebb, D.M.; Yamamoto, Y.; Sasaki, T.; Matsumoto, H. Engineering high-level aluminium tolerance in barley with the ALMT1 gene. Proc. Natl. Acad. Sci. USA 2004, 101, 15249–15254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Slaski, J.J.; Archambault, D.J.; Taylor, G.J. Alternation of plasma membrane lipids in aluminum-resistant and aluminum-sensitive wheat genotypes in response to aluminum stress. Physiol. Plant. 1997, 99, 302–308. [Google Scholar] [CrossRef]
- Miyasaka, S.C.; Hawes, M.C. Possible role of root border cells in detection and avoidance of aluminum toxicity. Plant Physiol. 2001, 125, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Boscolo, P.R.S.; Menossi, M.; Jorge, R.A. Aluminum-induced oxidative stress in maize. Phytochemistry 2003, 62, 181–189. [Google Scholar] [CrossRef]
- Doncheva, S.; Amenós, M.; Poschenrieder, C.; Barceló, J. Root cell patterning: A primary target for aluminum toxicity in maize. J. Exp. Bot. 2005, 56, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Narukawa, M.; Kanbara, K.; Tominaga, Y. Chlorogenic acid facilitates root hair formation in lettuce seedlings. Plant Cell Physiol. 2009, 50, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Arroyave, C.; Barceló, J.; Poschenrieder, C.; Tolrà, R. Aluminium-induced changes in root epidermal cell patterning, a distinctive feature of hyperresistance to Al in Brachiaria decumbens. J. Inorg. Biochem. 2011, 105, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Kinraide, T.B. Aluminum enhancement of plant growth in acid rooting media. A case of reciprocal alleviation of toxicity by two toxic cations. Physiol. Plant. 1993, 88, 619–625. [Google Scholar] [CrossRef]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Hue, N.V.; Craddock, G.R.; Adams, F. Effect of organic acids on aluminum toxicity in subsoils. Soil Sci. Soc. Am. J. 1986, 50, 28–34. [Google Scholar] [CrossRef]
- Ginting, S.; Johnson, B.B.; Wilkens, S. Alleviation of aluminium phytotoxicity on soybean growth by organic anions in nutrient solutions. Aust. J. Plant Physiol. 1998, 25, 901–908. [Google Scholar] [CrossRef]
- Ma, J.F. Role of organic acids in detoxification of aluminum in higher plants. Plant Cell Physiol. 2000, 41, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Delhaize, E.; Ryan, P.R.; Randall, P.J. Aluminum tolerance in wheat (Triticum aestivum L.) II. Aluminum-stimulated excretion of malic acid from root apices. Plant Physiol. 1993, 103, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.R.; Raman, H.; Gupta, S.; Horst, W.J.; Delhaize, E. A second mechanism for aluminum resistance in wheat relies on the constitutive efflux of citrate from roots. Plant Physiol. 2009, 149, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveira, A.L.; Poschenrieder, C.; Barcelo, J.; Martins-Lopes, P. Breeding for Al tolerance by unravelling genetic diversity in bread wheat. In Aluminum Stress Adaptation in Plants; Panda, S.K., Baluška, F., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 125–153. [Google Scholar]
- Garcia-Oliveira, A.L.; Martins-Lopes, P.; Tolrá, R.; Poschenrieder, C.; Tarquis, M.; Guedes-Pinto, H.; Benito, C. Molecular characterization of the citrate transporter gene TaMATE1 and expression analysis of upstream genes involved in organic acid transport under Al stress in bread wheat (Triticum aestivum L.). Physiol. Plant. 2014, 152, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Tovkach, A.; Ryan, P.R.; Richardson, A.E.; Lewis, D.C.; Rathjen, T.M.; Ramesh, S.; Tyerman, S.D.; Delhaize, E. Transposon-mediated alteration of TaMATE1B expression in wheat confers constitutive citrate efflux from root apices. Plant Physiol. 2013, 161, 880–892. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, J.V.; Liu, J.; Guimarães, C.T.; Lana, U.G.; Alves, V.M.; Wang, Y.H.; Schaffert, R.E.; Hoekenga, O.A.; Piñeros, M.A.; Shaff, J.E.; et al. A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat. Genet. 2007, 39, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Šukalović, V.H.T.; Vuletić, M.; Marković, K.; Vučinić, Ž. Cell wall-associated malate dehydrogenase activity from maize roots. Plant Sci. 2011, 181, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Yamamoto, Y.; Ezaki, B.; Katsuhara, M.; Ahn, S.J.; Ryan, P.R.; Delhaize, E.; Matsumoto, H. A wheat gene encoding an aluminum-activated malate transporter. Plant J. 2004, 37, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Ryan, P.R.; Delhaize, E.; Hebb, D.M.; Ogihara, Y.; Kawaura, K.; Noda, K.; Kojima, T.; Toyoda, A.; Matsumoto, H.; et al. Sequence upstream of the wheat (Triticum aestivum L.) ALMT1 gene and its relationship to aluminum resistance. Plant Cell Physiol. 2006, 47, 1343–1354. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Oliveira, A.L.; Martins-Lopes, P.; Tolrà, R.; Poschenrieder, C.; Guedes-Pinto, H.; Benito, C. Differential Physiological Responses of Portuguese Bread Wheat (Triticum aestivum L.) Genotypes under Aluminium Stress. Diversity 2016, 8, 26. https://doi.org/10.3390/d8040026
Garcia-Oliveira AL, Martins-Lopes P, Tolrà R, Poschenrieder C, Guedes-Pinto H, Benito C. Differential Physiological Responses of Portuguese Bread Wheat (Triticum aestivum L.) Genotypes under Aluminium Stress. Diversity. 2016; 8(4):26. https://doi.org/10.3390/d8040026
Chicago/Turabian StyleGarcia-Oliveira, Ana Luísa, Paula Martins-Lopes, Roser Tolrà, Charlotte Poschenrieder, Henrique Guedes-Pinto, and César Benito. 2016. "Differential Physiological Responses of Portuguese Bread Wheat (Triticum aestivum L.) Genotypes under Aluminium Stress" Diversity 8, no. 4: 26. https://doi.org/10.3390/d8040026
APA StyleGarcia-Oliveira, A. L., Martins-Lopes, P., Tolrà, R., Poschenrieder, C., Guedes-Pinto, H., & Benito, C. (2016). Differential Physiological Responses of Portuguese Bread Wheat (Triticum aestivum L.) Genotypes under Aluminium Stress. Diversity, 8(4), 26. https://doi.org/10.3390/d8040026