
Diatom Red List Species Reveal High Conservation Value and Vulnerability of Mountain Lakes


Abstract
:1. Introduction
2. Materials and Methods
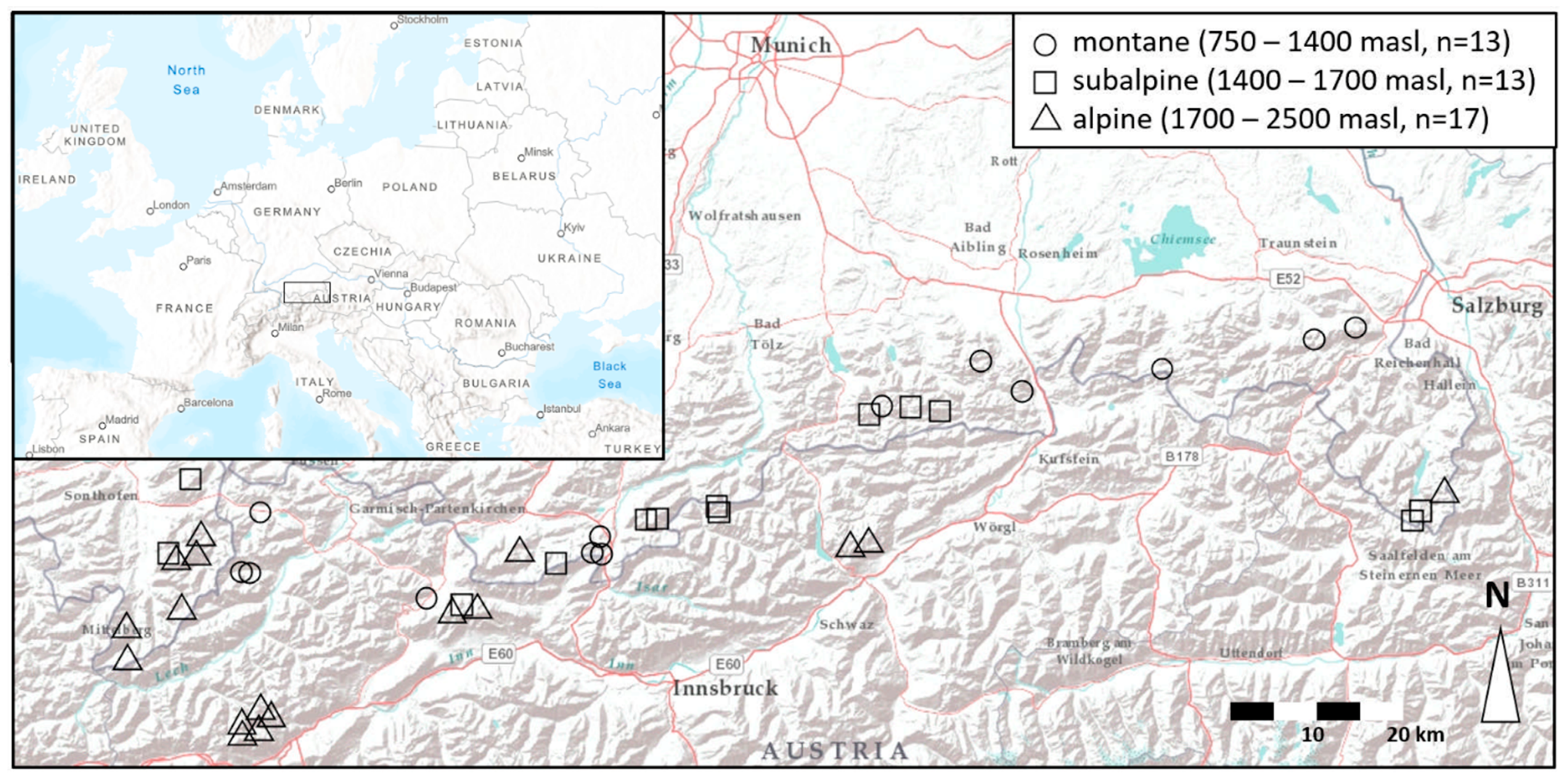
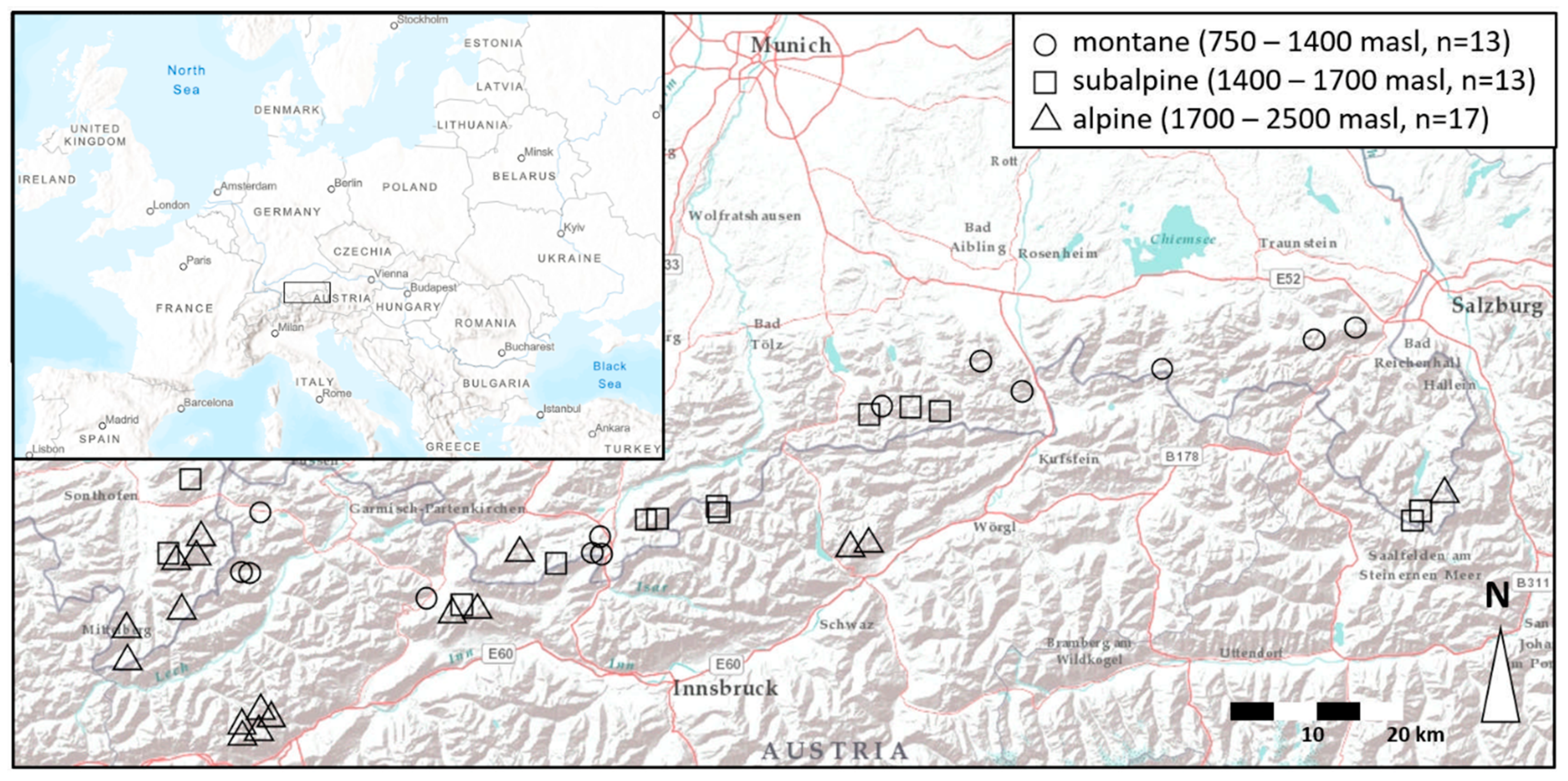
2.1. Study Site
2.2. Sampling and Laboratory Procedures
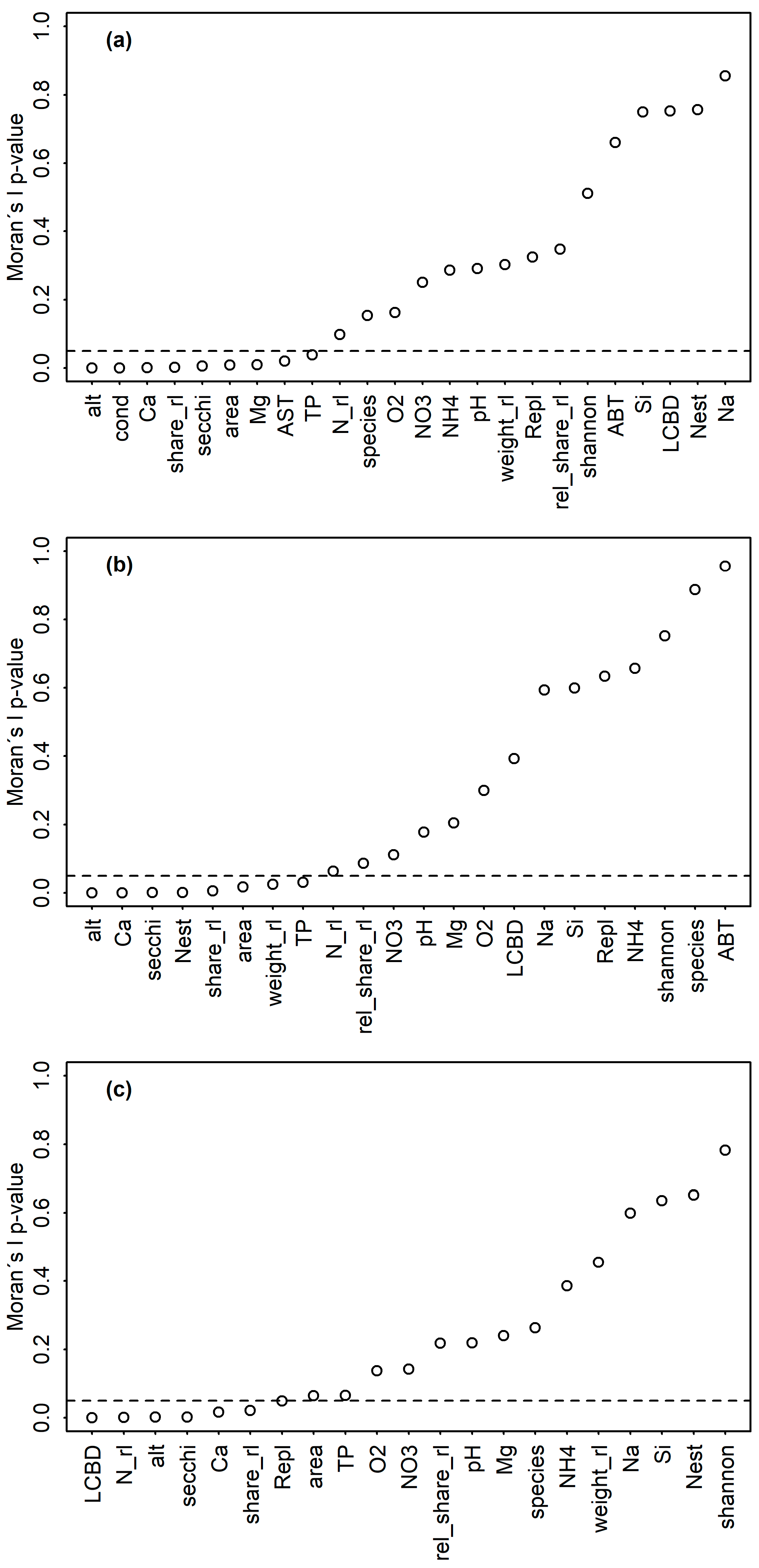
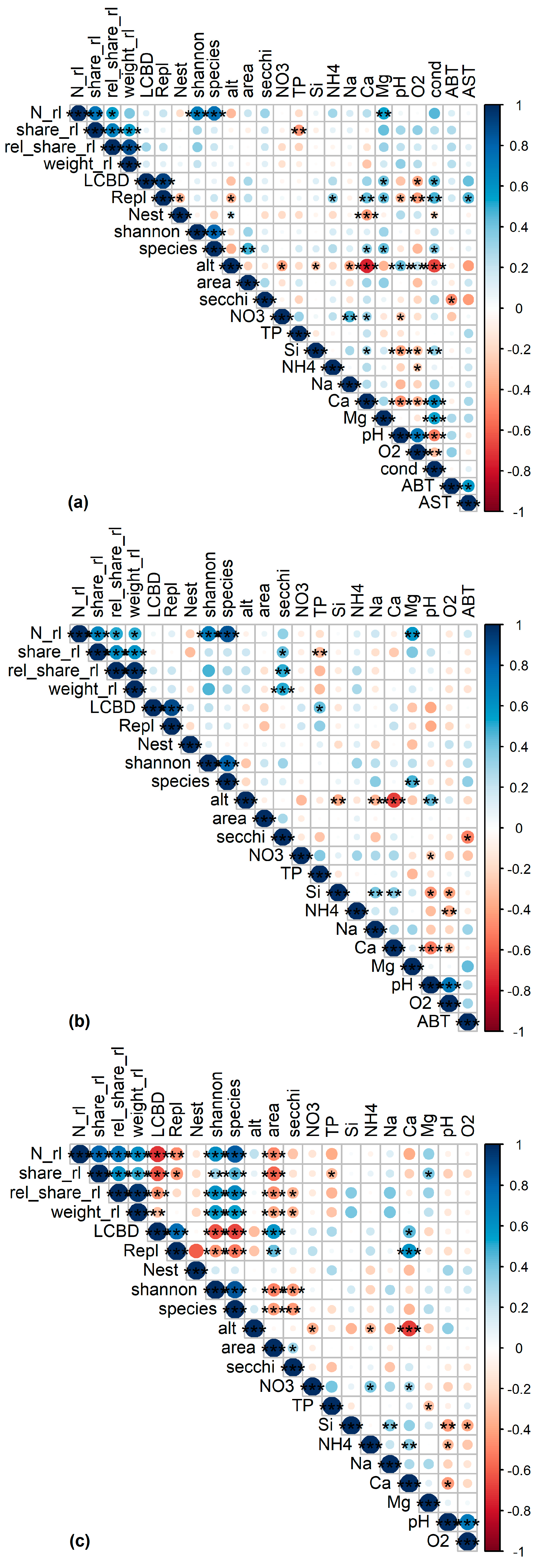
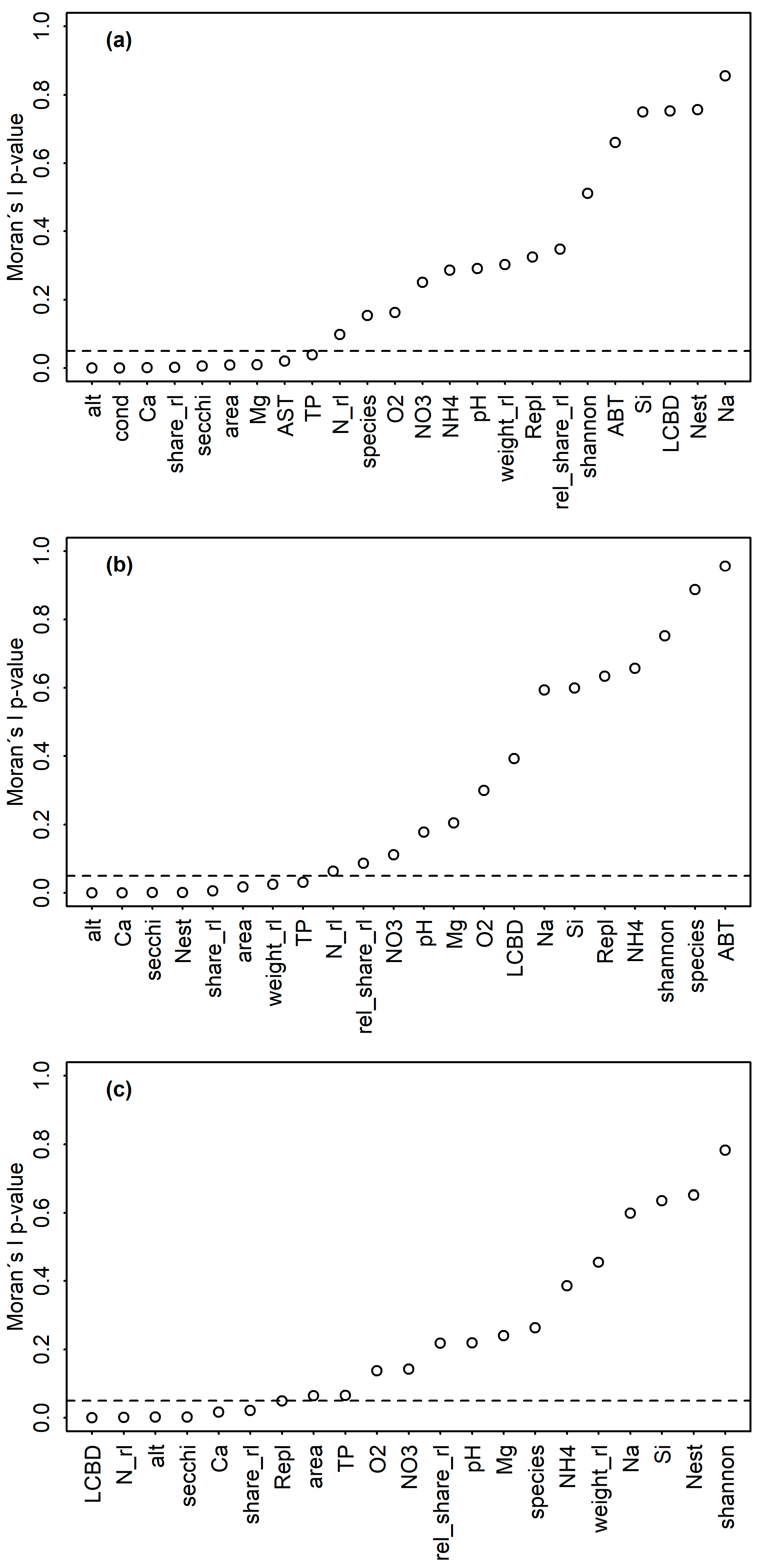
2.3. Data Analysis
- N_rl = sum of RLD
- share_rl = N_rl/N
- rel_share_rl = AbRL/Ab
- weight_rl = rel_share_rl weighted by Red List class
3. Results
4. Discussion
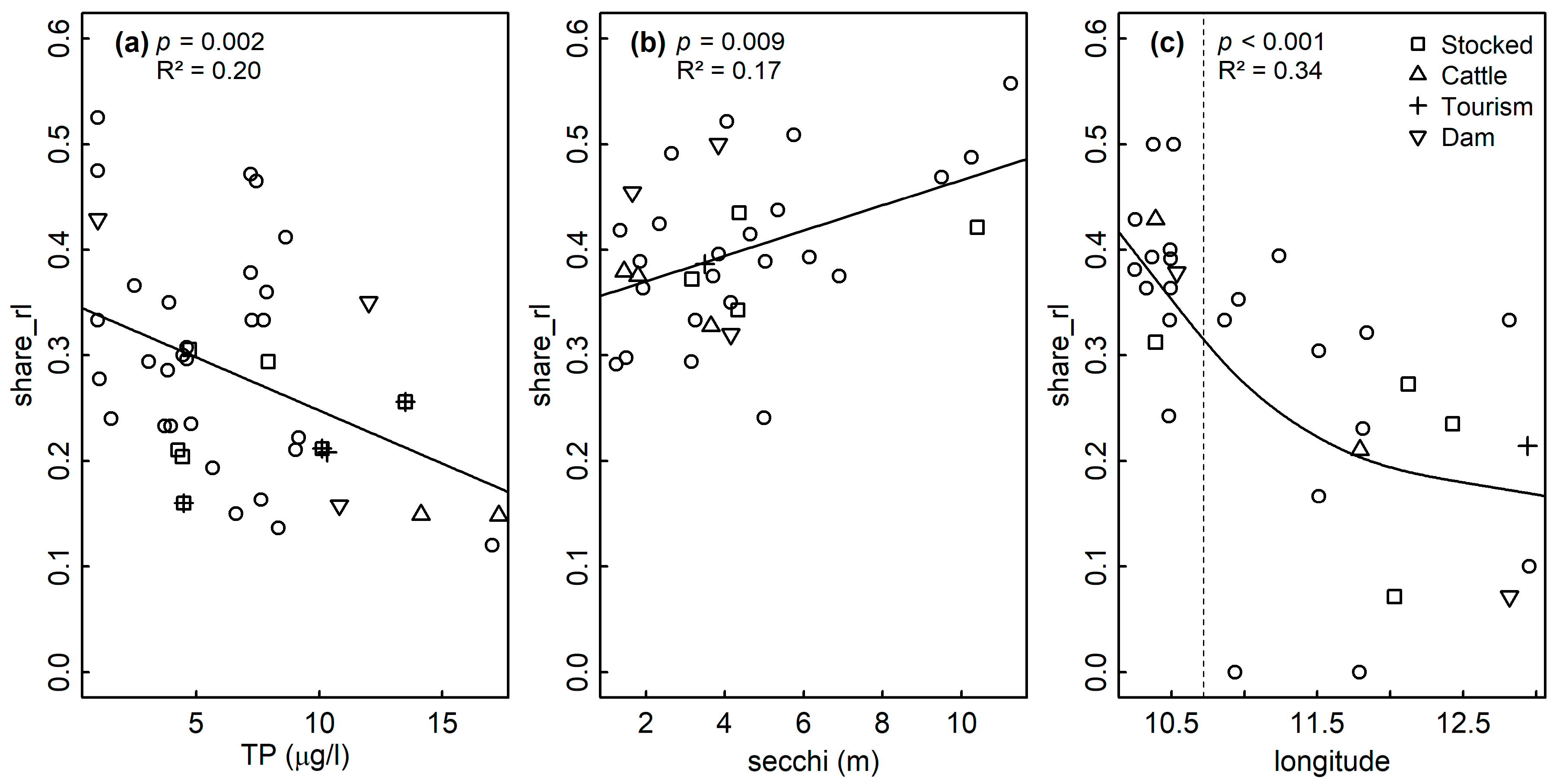
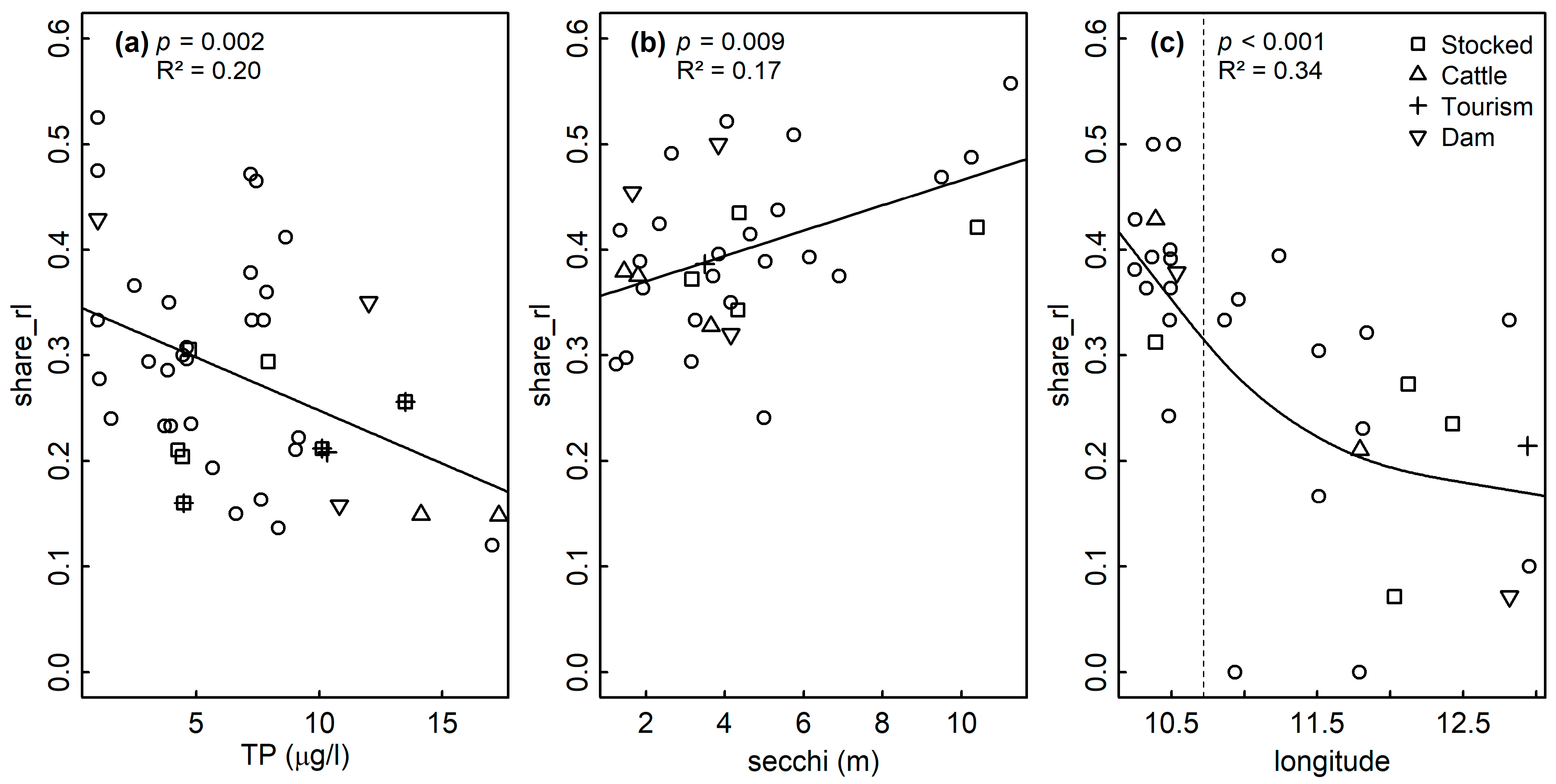
4.1. Nutrients and the Share of Red List Diatoms
4.2. Uniqueness as an Indicator of Fish Stocking
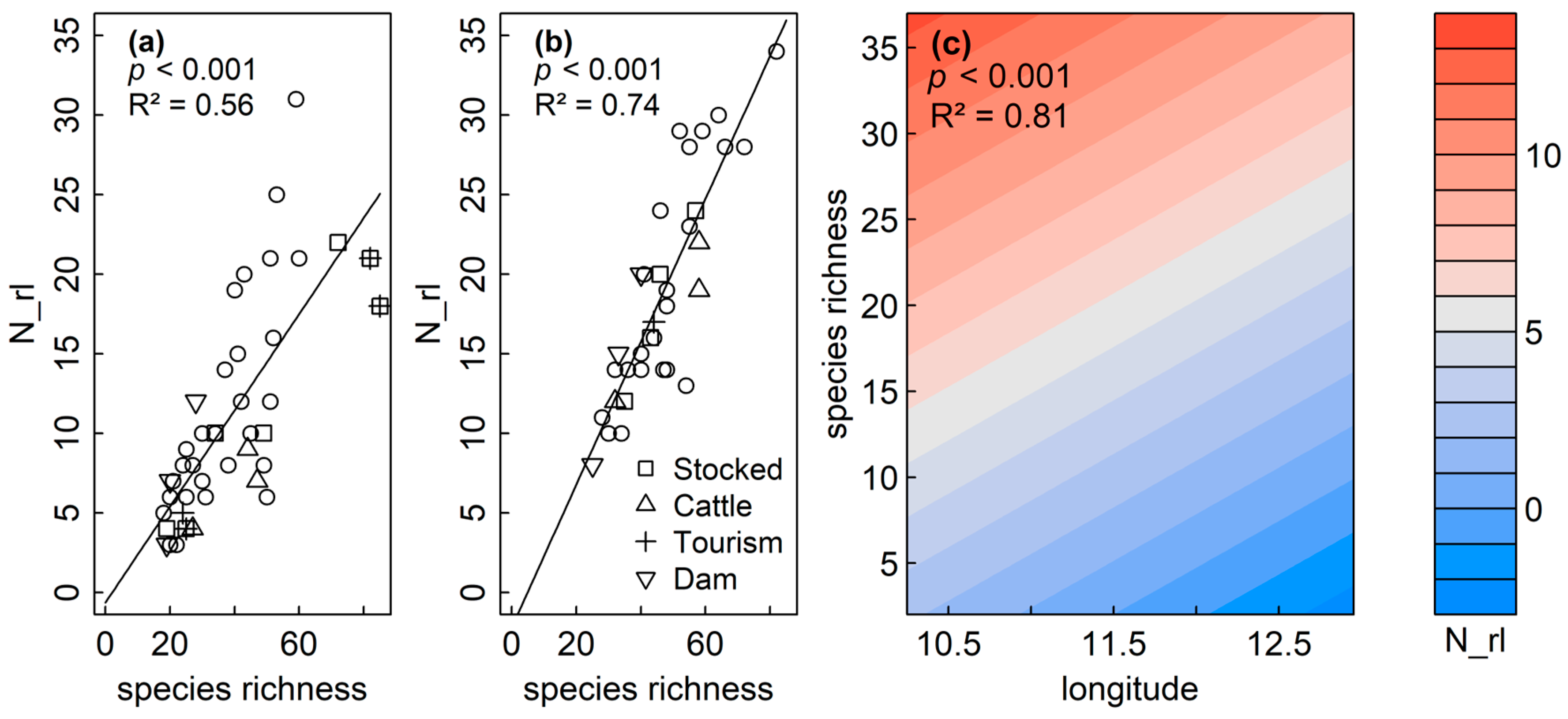
4.3. Diatom Richness and Conservation Value
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mittermeier, R.A.; Myers, N.; Thomsen, J.B.; da Fonseca, G.A.B.; Olivieri, S. Biodiversity hotspots and major tropical wilderness areas: Approaches to setting conservation priorities. Conserv. Biol. 1998, 12, 516–520. [Google Scholar] [CrossRef]
- Darwall, W.R.T.; Vie, J.C. Identifying important sites for conservation of freshwater biodiversity: Extending the species-based approach. Fish. Manag. Ecol. 2005, 12, 287–293. [Google Scholar] [CrossRef]
- Groom, M.; Meffe, G.; Carroll, C. Principles of Conservation Biology; Sinauer Associates: Sunderland, MA, USA, 2012; p. 779. [Google Scholar]
- Knight, A.T.; Cowling, R.M.; Rouget, M.; Balmford, A.; Lombard, A.T.; Campbell, B.M. Knowing but not doing: Selecting priority conservation areas and the research-implementation gap. Conserv. Biol. 2008, 22, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Geist, J. Seven steps towards improving freshwater conservation. Aquat. Conserv.-Mar. Freshw. Ecosyst. 2015, 25, 447–453. [Google Scholar] [CrossRef]
- Geist, J. Integrative freshwater ecology and biodiversity conservation. Ecol. Indic. 2011, 11, 1507–1516. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Leveque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Darwall, W.R.T.; Holland, R.A.; Smith, K.G.; Allen, D.; Brooks, E.G.E.; Katarya, V.; Pollock, C.M.; Shi, Y.; Clausnitzer, V.; Cumberlidge, N.; et al. Implications of bias in conservation research and investment for freshwater species. Conserv. Lett. 2011, 4, 474–482. [Google Scholar] [CrossRef]
- EEA. Introduction to the EU Water Framework Directive. Available online: https://ec.europa.eu/environment/water/water-framework/info/intro_en.htm (accessed on 16 March 2022).
- Kristensen, P.; Globevnik, L. European Small Waterbodies. Biol. Environ.-Proc. R. Ir. Acad. 2014, 114B, 281–287. [Google Scholar] [CrossRef]
- EEA. European Rivers and Lakes—Assessment of Their Environmental State; European Environmental Agency: Copenhagen, Denmark, 1994; p. 124.
- EEA. European Waters—Assessment of Status and Pressures; European Environmental Agency: Copenhagen, Denmark, 2018; p. 90.
- Biggs, J.; von Fumetti, S.; Kelly-Quinn, M. The importance of small waterbodies for biodiversity and ecosystem services: Implications for policy makers. Hydrobiologia 2017, 793, 3–39. [Google Scholar] [CrossRef]
- Martinez-Sanz, C.; Cenzano, C.S.S.; Fernandez-Alaez, M.; Garcia-Criado, F. Relative contribution of small mountain ponds to regional richness of littoral macroinvertebrates and the implications for conservation. Aquat. Conserv.-Mar. Freshw. Ecosyst. 2012, 22, 155–164. [Google Scholar] [CrossRef]
- Sommaruga, R. The role of solar UV radiation in the ecology of alpine lakes. J. Photochem. Photobiol. B Biol. 2001, 62, 35–42. [Google Scholar] [CrossRef]
- Casamayor, E.O. Towards a Microbial Conservation Perspective in High Mountain Lakes. In High Mountain Conservation in a Changing World, 1st ed.; Catalan, J., Ninot, J.M., Aniz, M.M., Eds.; Springer Nature: Cham, Switzerland, 2017; Volume 63, pp. 157–180. [Google Scholar]
- Ortiz-Alvarez, R.; Triado-Margarit, X.; Camarero, L.; Casamayor, E.O.; Catalan, J. High planktonic diversity in mountain lakes contains similar contributions of autotrophic, heterotrophic and parasitic eukaryotic life forms. Sci. Rep. 2018, 8, 4457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, K.A.; Baron, J.S.; Brahney, J.; Oleksy, I.A.; Saros, J.E.; Hundey, E.J.; Sadro, S.; Kopáček, J.; Sommaruga, R.; Kainz, M.J.; et al. Mountain lakes: Eyes on global environmental change. Glob. Planet. Chang. 2019, 178, 77–95. [Google Scholar] [CrossRef] [Green Version]
- Tiberti, R.; Rogora, M.; Tartari, G.; Callieri, C. Ecological impact of transhumance on the trophic state of alpine lakes in Gran Paradiso National Park. Knowl. Manag. Aquat. Ecosyst. 2014, 415, 05. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, A.M.; Kuefner, W.; Mayr, C.; Dubois, N.; Geist, J.; Raeder, U. Unravelling climate change impacts from other anthropogenic influences in a subalpine lake: A multi-proxy sediment study from Oberer Soiernsee (Northern Alps, Germany). Hydrobiologia 2021, 848, 4285–4309. [Google Scholar] [CrossRef]
- Dynowski, P.; Senetra, A.; Zrobek-Sokolnik, A.; Kozlowski, J. The Impact of Recreational Activities on Aquatic Vegetation in Alpine Lakes. Water 2019, 11, 173. [Google Scholar] [CrossRef] [Green Version]
- Tiberti, R.; von Hardenberg, A.; Bogliani, G. Ecological impact of introduced fish in high altitude lakes: A case of study from the European Alps. Hydrobiologia 2014, 724, 1–19. [Google Scholar] [CrossRef]
- Schindler, D.E.; Knapp, R.A.; Leavitt, P.R. Alteration of Nutrient Cycles and Algal Production Resulting from Fish Introductions into Mountain Lakes. Ecosystems 2001, 4, 308–321. [Google Scholar] [CrossRef]
- Detmer, T.; Lewis, W.M. Influences of fish on food web structure and function in mountain lakes. Freshw. Biol. 2019, 64, 1572–1583. [Google Scholar] [CrossRef]
- Ventura, M.; Tiberti, R.; Buchaca, T.; Bunay, D.; Sabas, I.; Miro, A. Why Should We Preserve Fishless High Mountain Lakes? In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J.M., Aniz, M.M., Eds.; Springer Nature: Cham, Switzerland, 2017; Volume 62, pp. 181–205. [Google Scholar]
- Selene, P.; Paolo, P.; Marco, B.; Salvi, G.; Franz, F.; Marino, P.; Pizzul, E. Changes in midge assemblages (Diptera Chironomidae) in an alpine lake from the Italian Western Alps: The role and importance of fish introduction. Hydrobiologia 2020, 847, 2393–2415. [Google Scholar] [CrossRef]
- Tolotti, M.; Thies, H.; Nickus, U.; Psenner, R. Temperature modulated effects of nutrients on phytoplankton changes in a mountain lake. Hydrobiologia 2012, 698, 61–75. [Google Scholar] [CrossRef]
- Kuefner, W.; Hofmann, A.; Ossyssek, S.; Dubois, N.; Geist, J.; Raeder, U. Composition of highly diverse diatom community shifts as response to climate change: A down-core study of 23 central European mountain lakes. Ecol. Indic. 2020, 117, 106590. [Google Scholar] [CrossRef]
- Rogora, M.; Frate, L.; Carranza, M.L.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleksy, I.A.; Baron, J.S.; Leavitt, P.R.; Spaulding, S.A. Nutrients and warming interact to force mountain lakes into unprecedented ecological states. Proc. R. Soc. B-Biol. Sci. 2020, 287, 20200304. [Google Scholar] [CrossRef] [PubMed]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef] [PubMed]
- Rosbakh, S.; Bernhardt-Romermann, M.; Poschlod, P. Elevation matters: Contrasting effects of climate change on the vegetation development at different elevations in the Bavarian Alps. Alp. Bot. 2014, 124, 143–154. [Google Scholar] [CrossRef]
- Schönfelder, I.; Gelbrecht, J.; Schönfelder, J.; Steinberg, C.E.W. Relationships between littoral diatoms and their chemical environment in northeastern German lakes and rivers. J. Phycol. 2002, 38, 66–82. [Google Scholar] [CrossRef]
- Weckstrom, J.; Korhola, A.; Blom, T. Diatoms as quantitative indicators of pH and water temperature in subarctic Fennoscandian lakes. Hydrobiologia 1997, 347, 171–184. [Google Scholar] [CrossRef]
- Kuefner, W.; Hofmann, A.M.; Geist, J.; Raeder, U. Evaluating climate change impacts on mountain lakes by applying the new silicification value to paleolimnological samples. Sci. Total Environ. 2020, 715, 12. [Google Scholar] [CrossRef]
- Bennion, H.; Fluin, J.; Simpson, G.L. Assessing eutrophication and reference conditions for Scottish freshwater lochs using subfossil diatoms. J. Appl. Ecol. 2004, 41, 124–138. [Google Scholar] [CrossRef]
- Battarbee, R.W. Diatom analysis and the acidification of lakes. Philos. Trans. R. Soc. B-Biol. Sci. 1984, 305, 451–477. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The diatoms: Biology and morphology of the genera. J. Mar. Biol. Assoc. UK 1990, 70, 924. [Google Scholar] [CrossRef]
- Guiry, M.D. How many Species of Algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.G.; Droop, S.J.M. Biodiversity, biogeography and conservation of diatoms. Hydrobiologia 1996, 336, 19–32. [Google Scholar] [CrossRef]
- Hofmann, G.; Lange-Bertalot, H.; Werum, M.; Klee, R. Rote Liste und Gesamtartenliste der limnischen Kieselalgen (Bacillariophyta) Deutschlands. In Rote Liste Gefährdeter Tiere, Pflanzen und Pilze Deutschlands, Band 7: Pflanzen; Metzing, D.H.N., Ludwig, G., Matzke-Hajek, G., Eds.; Landwirtschaftsverlag: Münster, Germany, 2018; Naturschutz und Biologische Vielfalt; pp. 601–708. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Drinan, T.J.; Foster, G.N.; Nelson, B.H.; O’Halloran, J.; Harrison, S.S.C. Macroinvertebrate assemblages of peatland lakes: Assessment of conservation value with respect to anthropogenic land-cover change. Biol. Conserv. 2013, 158, 175–187. [Google Scholar] [CrossRef]
- Lukacs, B.A.; Sramko, G.; Molnar, V.A. Plant diversity and conservation value of continental temporary pools. Biol. Conserv. 2013, 158, 393–400. [Google Scholar] [CrossRef]
- Legendre, P.; De Caceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Tonkin, J.D.; Heino, J.; Sundermann, A.; Haase, P.; Jahnig, S.C. Context dependency in biodiversity patterns of central German stream metacommunities. Freshw. Biol. 2016, 61, 607–620. [Google Scholar] [CrossRef]
- Juran, J.; Kastovsky, J. The procedure of compiling the Red List of microscopic algae of the Czech Republic. Biodivers. Conserv. 2019, 28, 2499–2529. [Google Scholar] [CrossRef]
- Cantonati, M.; Hofmann, G.; Spitale, D.; Werum, M.; Lange-Bertalot, H. Diatom Red Lists: Important tools to assess and preserve biodiversity and habitats in the face of direct impacts and environmental change. Biodivers. Conserv. 2022, 31, 453–477. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Decet, F.; Gabrieli, J. Diatoms in very-shallow pools of the site of community importance Danta di Cadore Mires (south-eastern Alps), and the potential contribution of these habitats to diatom biodiversity conservation. Nova Hedwig. 2011, 93, 475–507. [Google Scholar] [CrossRef]
- Mogna, M.; Cantonati, M.; Andreucci, F.; Angeli, N.; Berta, G.; Miserere, L. Diatom communities and vegetation of springs in the south-western Alps. Acta Bot. Croat. 2015, 74, 265–285. [Google Scholar] [CrossRef] [Green Version]
- Falasco, E.; Piano, E.; Bona, F. Diatom flora in Mediterranean streams: Flow intermittency threatens endangered species. Biodivers. Conserv. 2016, 25, 2965–2986. [Google Scholar] [CrossRef]
- Tolotti, M. Phytoplankton and littoral epilithic diatoms in high mountain lakes of the Adamello-Brenta Regional Park (Trentino, Italy) and their relation to trophic status and acidification risk. J. Limnol. 2001, 60, 171–188. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Achnanthidium dolomiticum sp. nov. (Bacillariophyta) from oligotrophic Mountain Springs and Lakes fed by Dolomite Aquifers. J. Phycol. 2006, 42, 1184–1188. [Google Scholar] [CrossRef]
- Cantonati, M.; Segadelli, S.; Spitale, D.; Gabrieli, J.; Gerecke, R.; Angeli, N.; De Nardo, M.T.; Ogata, K.; Wehr, J.D. Geological and hydrochemical prerequisites of unexpectedly high biodiversity in spring ecosystems at the landscape level. Sci. Total Environ. 2020, 740, 140157. [Google Scholar] [CrossRef]
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (south-eastern Alps). Fundam. Appl. Limnol. 2009, 174, 117–133. [Google Scholar] [CrossRef]
- Falasco, E.; Piano, E.; Doretto, A.; Fenoglio, S.; Bona, F. Resilience of benthic diatom communities in Mediterranean streams: Role of endangered species. Mar. Freshw. Res. 2019, 70, 212–224. [Google Scholar] [CrossRef]
- Leira, M.; Chen, G.; Dalton, C.; Irvine, K.; Taylor, D. Patterns in freshwater diatom taxonomic distinctness along an eutrophication gradient. Freshw. Biol. 2009, 54, 1–14. [Google Scholar] [CrossRef]
- Tilman, D.; Kilham, S.S.; Kilham, P. Phytoplankton Community Ecology-The Role of Limiting Nutrients. Annu. Rev. Ecol. Syst. 1982, 13, 349–372. [Google Scholar] [CrossRef]
- Velghe, K.; Vermaire, J.C.; Gregory-Eaves, I. Declines in littoral species richness across both spatial and temporal nutrient gradients: A palaeolimnological study of two taxonomic groups. Freshw. Biol. 2012, 57, 2378–2389. [Google Scholar] [CrossRef]
- CEN. Water Quality—Guidance on Quantitative and Qualitative Sampling of Phytoplankton from Inland Waters; European Committee for Standardization: Brussels, Belgium, 2015; p. 40. [Google Scholar]
- Throndsen, R. Preservation and Storage. In Phytoplankton Manual; Sournia, A., Ed.; UNESCO: Paris, France, 1978; pp. 70–71. [Google Scholar]
- DIN. Water Quality—Guidance for the Routine Sampling and Preparation of Benthic Diatoms from Rivers and Lakes; Beuth Verlag GmbH: Berlin, Germany, 2014; p. 17. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Navone, R. Proposed method for nitrate in potable waters. Am. Water Work. Assoc. 1964, 56, 781–783. [Google Scholar] [CrossRef]
- DIN. Deutsche Einheitsverfahren zur Wasser-, Abwasser-und Schlammuntersuchung; Kationen (Gruppe E); Bestimmung des Ammonium-Stickstoffs (E 5); Beuth Verlag GmbH: Berlin, Germany, 1983. [Google Scholar]
- Nixdorf, B.; Hoehn, E.; Riedmüller, U.; Mischke, U.; Schönfelder, I. Probenahme und Analyse des Phytoplanktons in Seen und Flüssen zur Ökologischen Bewertung gemäß der EU-WRRL. In Handbuch Angewandte Limnologie: Grundlagen-Gewässerbelastung-Restaurierung-Aquatische Ökotoxikologie-Bewertung-Gewässerschutz; Hupfer, M., Fischer, H., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2014. [Google Scholar]
- van der Werff, A.; Macan, T.T. A new method of concentrating and cleaning diatoms and other organisms. Proc. Int. Assoc. Theor. Appl. Limnol. (SIL) 1955, 12, 276–277. [Google Scholar] [CrossRef]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 3. Teil: Centrales, Fragilariaceae, Eunotiaceae; Gustav Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema; Gustav Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae; Gustav Fischer Verlag: Stuttgart, Germany, 1997. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 1. Teil: Naviculaceae; Gustav Fischer Verlag: Stuttgart, Germany, 1997. [Google Scholar]
- Krammer, K. Diatoms of the European Inland Waters and Comparable Habitats: The Genus Pinnularia; A.R.G. Gartner Verlag K.G.: Rugell, Liechtenstein, 2000. [Google Scholar]
- Lange-Bertalot, H. Diatoms of the European Inland Waters and Comparable Habitats: Navicula Sensu Stricto, 10 Genera Separated from Navicula Sensu Stricto, Frustulia; A.R.G. Gartner Verlag K.G.: Rugell, Liechtenstein, 2001. [Google Scholar]
- Krammer, K. Diatoms of the European Inland Waters and Comparable Habitats: Cymbella; A.R.G. Gartner Verlag K.G.: Rugell, Liechtenstein, 2002; p. 584. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe; Over 800 common species used in ecological assessment; Koeltz Botanical Books: Oberreifenberg, Germany, 2017. [Google Scholar]
- Pla-Rabes, S.; Catalan, J. Diatom species variation between lake habitats: Implications for interpretation of paleolimnological records. J. Paleolimnol. 2018, 60, 169–187. [Google Scholar] [CrossRef]
- Rimet, F.; Bouchez, A.; Tapolczai, K. Spatial heterogeneity of littoral benthic diatoms in a large lake: Monitoring implications. Hydrobiologia 2016, 771, 179–193. [Google Scholar] [CrossRef]
- Naimi, B. R Package ‘usdm’; The R Foundation: Vienna, Austria, 2017. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Henry, M.; Stevens, S. The Vegan Package; The R Foundation: Vienna, Austria, 2007. [Google Scholar]
- Baselga, A. Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients. Methods Ecol. Evol. 2013, 4, 552–557. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V.R. R Package “Corrplot”: Visualization of a Correlation Matrix; The R Foundation: Vienna, Austria, 2017. [Google Scholar]
- Ossyssek, S.; Hofmann, A.; Geist, J.; Raeder, U. Sedimentary, planktic and littoral diatom metacommunities in mountain lakes of the northern European Alps show divergent assembly patterns. Hydrobiologia, 2022; submitted. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Bivand, R.S.; Wong, D.W.S. Comparing implementations of global and local indicators of spatial association. TEST 2018, 27, 716–748. [Google Scholar] [CrossRef]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B-Stat. Methodol. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Cantonati, M.; Bilous, O.; Spitale, D.; Angeli, N.; Segadelli, S.; Bernabè, D.; Lichtenwöhrer, K.; Gerecke, R.; Saber, A.A. Diatoms from the Spring Ecosystems Selected for the Long-Term Monitoring of Climate-Change Effects in the Berchtesgaden National Park (Germany). Water 2022, 14, 381. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Diatom biodiversity of springs in the Berchtesgaden National Park (North-Eastern Alps, Germany), with the ecological and morphological characterization of two species new to science. Diatom Res. 2010, 25, 251–280. [Google Scholar] [CrossRef]
- Muri, G.; Cermelj, B.; Jacimovic, R.; Ravnikar, T.; Smuc, A.; Tursic, J.; Vreca, P. Factors that contributed to recent eutrophication of two Slovenian mountain lakes. J. Paleolimnol. 2018, 59, 411–426. [Google Scholar] [CrossRef]
- Sienkiewicz, E.; Gasiorowski, M. Changes in the Trophic Status of Three Mountain Lakes-Natural or Anthropogenic Process? Pol. J. Environ. Stud. 2014, 23, 875–892. [Google Scholar]
- Kuefmann, C. Soil types and eolian dust in high-mountainous karst of the Northern Calcareous Alps (Zugspitzplatt, Wetterstein Mountains, Germany). Catena 2003, 53, 211–227. [Google Scholar] [CrossRef]
- Bergström, A.-K. The use of TN:TP and DIN:TP ratios as indicators for phytoplankton nutrient limitation in oligotrophic lakes affected by N deposition. Aquat. Sci. 2010, 72, 277–281. [Google Scholar] [CrossRef]
- Rogora, M.; Massaferro, J.; Marchetto, A.; Tartari, G.; Mosello, R. The water chemistry of some shallow lakes in Northern Patagonia and their nitrogen status in comparison with remote lakes in different regions of the globe. J. Limnol. 2008, 67, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Muri, G.; Cermelj, B.; Jacimovic, R.; Skaberne, D.; Smuc, A.; Burnik Sturm, M.; Turšič, J.; Vreca, P. Consequences of anthropogenic activity for two remote alpine lakes in NW Slovenia as tracked by sediment geochemistry. J. Paleolimnol. 2013, 50, 457–470. [Google Scholar] [CrossRef]
- Rogora, M.; Arisci, S.; Marchetto, A. The role of nitrogen deposition in the recent nitrate decline in lakes and rivers in Northern Italy. Sci. Total Environ. 2012, 417, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Angeli, N.; Cantonati, M.; Spitale, D.; Bertalot, H.L. A comparison between diatom assemblages in two groups of carbonate, low-altitude springs with different levels of anthropogenic disturbances. Fottea 2010, 10, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Denys, L.; Oosterlynck, P. Diatom assemblages of non-living substrates in petrifying Cratoneurion springs from lower Belgium. Fottea 2015, 15, 123–138. [Google Scholar] [CrossRef]
- Karna, O.M.; Heino, J.; Laamanen, T.; Jyrkankallio-Mikkola, J.; Pajunen, V.; Soininen, J.; Tolonen, K.T.; Tukiainen, H.; Hjort, J. Does catchment geodiversity foster stream biodiversity? Landsc. Ecol. 2019, 34, 2469–2485. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, A.M.; Geist, J.; Nowotny, L.; Raeder, U. Depth-distribution of lake benthic diatom assemblages in relation to light availability and substrate: Implications for paleolimnological studies. J. Paleolimnol. 2020, 64, 315–334. [Google Scholar] [CrossRef]
- Cantonati, M.; Scola, S.; Angeli, N.; Guella, G.; Frassanito, R. Environmental controls of epilithic diatom depth-distribution in an oligotrophic lake characterized by marked water-level fluctuations. Eur. J. Phycol. 2009, 44, 15–29. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Sondergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Jensen, C.; Faafeng, B.; Hessen, D.O.; Sondergaard, M.; Lauridsen, T.; Brettum, P.; Christoffersen, K. The impact of nutrient state and lake depth on top-down control in the pelagic zone of lakes: A study of 466 lakes from the temperate zone to the arctic. Ecosystems 2003, 6, 313–325. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation Body Size and Composition of Plankton. Science 1965, 150, 28. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R.; Cochran, P.A.; Elser, J.J.; Elser, M.M.; Lodge, D.M.; Kretchmer, D.; He, X.; Vonende, C.N. Regulation of Lake Primary Productivity by Food Web Structure. Ecology 1987, 68, 1863–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benndorf, J.; Boing, W.; Koop, J.; Neubauer, I. Top-down control of phytoplankton: The role of time scale, lake depth and trophic state. Freshw. Biol. 2002, 47, 2282–2295. [Google Scholar] [CrossRef] [Green Version]
- Threlkeld, S.T. Planktivory and planktivore Biomass Effects on Zooplankton, Phytoplankton, and the Trophic Cascade. Limnol. Oceanogr. 1988, 33, 1362–1375. [Google Scholar] [CrossRef]
- Pastorino, P.; Prearo, M.; Bertoli, M.; Menconi, V.; Esposito, G.; Righetti, M.; Mugetti, D.; Pederiva, S.; Abete, M.C.; Pizzul, E. Assessment of Biological and Sanitary Condition of Alien Fish from a High-Mountain Lake (Cottian Alps). Water 2020, 12, 559. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.R.; Cottingham, K.L.; Schindler, D.E. Biotic Feedbacks in Lake Phosphorus Cycles. Trends Ecol. Evol. 1992, 7, 332–336. [Google Scholar] [CrossRef]
- Schindler, D.E.; Kitchell, J.F.; He, X.; Carpenter, S.R.; Hodgson, J.R.; Cottingham, K.L. Food-Web Structure and Phosphorus Cycling in Lakes. Trans. Am. Fish. Soc. 1993, 122, 756–772. [Google Scholar] [CrossRef]
- Whitmore, T.J.; Riedinger-Whitmore, M.A.; Lauterman, F.M.; Curtis, J.H. Cyanobacterial influence on diatom community lifeform dynamics in shallow subtropical lakes of Florida USA. J. Paleolimnol. 2018, 60, 223–246. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R. Cascading Trophic Interactions and Lake Productivity. Bioscience 1985, 35, 634–639. [Google Scholar] [CrossRef]
- Guilizzoni, P.; Lami, A.; Manca, M.; Musazzi, S.; Marchetto, A. Palaeoenvironmental changes inferred from biological remains in short lake sediment cores from the Central Alps and Dolomites. Hydrobiologia 2006, 562, 167–191. [Google Scholar] [CrossRef]
- Post, J.R.; McQueen, D.J. The Impact of planktivorous Fish on the Structure of a Plankton Community. Freshw. Biol. 1987, 17, 79–89. [Google Scholar] [CrossRef]
- Gliwicz, Z. Effect of zooplankton grazing on photosynthetic activity and composition of phytoplankton. Int. Ver. Theor. Angew. Limnol. Verh. 1975, 19, 1490–1497. [Google Scholar] [CrossRef]
- Sienkiewicz, E.; Gasiorowski, M. The effect of fish stocking on mountain lake plankton communities identified using palaeobiological analyses of bottom sediment cores. J. Paleolimnol. 2016, 55, 129–150. [Google Scholar] [CrossRef] [Green Version]
- Gliwicz, Z.M. Why do Cladocerans fail to control Algal Blooms? Hydrobiologia 1990, 200, 83–97. [Google Scholar] [CrossRef]
- Kissman, C.E.H.; Williamson, C.E.; Rose, K.C.; Saros, J.E. Response of phytoplankton in an alpine lake to inputs of dissolved organic matter through nutrient enrichment and trophic forcing. Limnol. Oceanogr. 2013, 58, 867–880. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S. The Ecology of Freshwater Phytoplankton; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
- Ossyssek, S.; Geist, J.; Werner, P.; Raeder, U. Identification of the ecological preferences of Cyclotella comensis in mountain lakes of the northern European Alps. Arct. Antarct. Alp. Res. 2020, 52, 512–523. [Google Scholar] [CrossRef]
- Donald, D.B. Assessment of the Outcome of Eight Decades of Trout Stocking in the Mountain National Parks, Canada. N. Am. J. Fish. Manag. 1987, 7, 545–553. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. On the different nature of top-down and bottom-up effects in pelagic food webs. Freshw. Biol. 2002, 47, 2296–2312. [Google Scholar] [CrossRef]
- Reynolds, C.S. Scales of Disturbance and their Role in Plankton Ecology. Hydrobiologia 1993, 249, 157–171. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Wiseman, S.W.; Godfrey, B.M.; Butterwick, C. Some Effects of Artificial Mixing on the Dynamics of Phytoplankton Populations in Large Limnetic Enclosures. J. Plankton Res. 1983, 5, 203–234. [Google Scholar] [CrossRef]
- Benito, X.; Fritz, S.C.; Steinitz-Kannan, M.; Velez, M.I.; McGlue, M.M. Lake regionalization and diatom metacommunity structuring in tropical South America. Ecol. Evol. 2018, 8, 7865–7878. [Google Scholar] [CrossRef] [Green Version]
- Knapp, R.A.; Matthews, K.R. Eradication of nonnative fish by gill netting from a small mountain lake in California. Restor. Ecol. 1998, 6, 207–213. [Google Scholar] [CrossRef]
- Tiberti, R.; Buchaca, T.; Boiano, D.; Knapp, R.A.; Rovira, Q.P.; Tavecchia, G.; Ventura, M.; Tenan, S. Alien fish eradication from high mountain lakes by multiple removal methods: Estimating residual abundance and eradication probability in open populations. J. Appl. Ecol. 2021, 58, 1055–1068. [Google Scholar] [CrossRef]
- Parker, B.R.; Schindler, D.W. Cascading trophic interactions in an oligotrophic species-poor alpine lake. Ecosystems 2006, 9, 157–166. [Google Scholar] [CrossRef]
- Knapp, R.A. Effects of nonnative fish and habitat characteristics on lentic herpetofauna in Yosemite National Park, USA. Biol. Conserv. 2005, 121, 265–279. [Google Scholar] [CrossRef]
- Knapp, R.A.; Matthews, K.R.; Sarnelle, O. Resistance and resilience of alpine lake fauna to fish introductions. Ecol. Monogr. 2001, 71, 401–421. [Google Scholar] [CrossRef]
- Denoel, M.; Scime, P.; Zambelli, N. Newt life after fish introduction: Extirpation of paedomorphosis in a mountain fish lake and newt use of satellite pools. Curr. Zool. 2016, 62, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Bohm, M.; Collen, B.; Baillie, J.E.M.; Bowles, P.; Chanson, J.; Cox, N.; Hammerson, G.; Hoffmann, M.; Livingstone, S.R.; Ram, M.; et al. The conservation status of the world’s reptiles. Biol. Conserv. 2013, 157, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Cushman, S.A. Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar] [CrossRef]
- Gilbert, B.; Levine, J.M. Ecological drift and the distribution of species diversity. Proc. R. Soc. B-Biol. Sci. 2017, 284, 20170507. [Google Scholar] [CrossRef]
- Fodelianakis, S.; Valenzuela-Cuevas, A.; Barozzi, A.; Daffonchio, D. Direct quantification of ecological drift at the population level in synthetic bacterial communities. ISME J. 2021, 15, 55–66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alt. | Area | Depth | Sec. | NO3 | TP | Si | NH4 | Na | Ca | Mg | pH | Cond | O2 | ABT | AST | DIN:TP | Geo. | Reg. | F | C | T | D | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lake | m asl | ha | m | m | mg/L | µg/L | mg/L | µg/L | mg/L | mg/L | mg/L | µS/cm | % | °C | °C | |||||||||
| Fal | 760 | 1.0 | 15.0 | 7.0 | 1.84 | 9.0 | 0.18 | 17.0 | 2.11 | 42.3 | 6.7 | 8.0 | 266.3 | 96 | 5.8 | 12.8 | 205.4 | Ls | C | |||||
| Bic | 955 | 1.4 | 11.0 | 4.4 | 0.76 | 4.3 | 0.77 | 32.3 | 0.30 | 49.2 | 14.7 | 7.9 | 360.2 | 61 | 8.4 | 21.7 | 185.7 | Mi | V | + | ||||
| Fri | 973 | 1.2 | 5.5 | 4.6 | 0.93 | 7.2 | 0.15 | 83.7 | 0.48 | 39.6 | 5.4 | 8.6 | 224.3 | 110 | 13.9 | 18.6 | 140.1 | Ls | C | |||||
| Sut | 995 | 1.4 | 1.5 | 1.5 | 0.67 | 4.8 | 0.76 | 60.8 | 1.62 | 41.2 | 6.7 | 8.0 | 280.7 | 82 | 14.1 | 17.5 | 153.2 | Mi | V | |||||
| Laut | 1013 | 12.0 | 18.0 | 6.0 | 0.21 | 13.5 | 0.25 | 50.8 | 0.10 | 46.1 | 16.0 | 8.6 | 310.0 | 102 | 8.2 | 16.0 | 19.3 | Do | W | + | + | |||
| Fer | 1060 | 10.0 | 19.5 | 6.8 | 0.33 | 10.1 | 0.21 | 43.0 | 0.10 | 47.2 | 18.1 | 8.1 | 304.0 | 102 | 7.4 | 15.6 | 36.9 | Do | W | + | + | |||
| Mit | 1082 | 3.3 | 4.7 | 4.7 | 0.10 | 7.5 | 0.13 | 34.0 | 1.23 | 38.5 | 18.4 | 8.2 | 345.8 | 101 | 18.4 | 22.6 | 17.4 | W | ||||||
| Wild | 1136 | 2.3 | 2.4 | 2.4 | 0.22 | 8.6 | 0.19 | 53.6 | 0.35 | 31.6 | 19.5 | 8.6 | 296.8 | 77 | 20.9 | 21.2 | 31.4 | Do | W | |||||
| Tau | 1138 | 3.6 | 14.6 | 4.3 | 0.66 | 4.4 | 0.20 | 126.9 | 0.49 | 40.4 | 4.1 | 8.0 | 246.8 | 64 | 5.0 | 21.0 | 177.4 | Ls | C | + | ||||
| Hoef | 1192 | 0.6 | 1.9 | 1.9 | 0.47 | 3.1 | 0.14 | 5.3 | 0.14 | 20.7 | 10.1 | 8.8 | 220.6 | 64 | 14.3 | 15.2 | 153.1 | A | ||||||
| SieK | 1205 | 0.2 | 11.3 | 11.3 | 0.56 | 1.0 | 0.23 | 99.4 | 0.36 | 38.6 | 10.8 | 8.2 | 382.4 | 130 | 7.4 | 8.8 | 655.2 | A | ||||||
| SieG | 1207 | 0.8 | 20.2 | 9.5 | 0.55 | 1.0 | 0.24 | 3.8 | 0.51 | 39.1 | 11.8 | 8.1 | 536.7 | 118 | 5.8 | 10.8 | 552.8 | A | ||||||
| GruW | 1393 | 2.3 | 5.8 | 3.7 | 0.18 | 14.1 | 0.18 | 19.9 | 0.29 | 29.7 | 3.2 | 8.1 | 198.6 | 99 | 14.0 | 20.3 | 13.8 | Ma | V | + | ||||
| Roe | 1450 | 1.0 | 7.6 | 1.5 | 2.86 | 20.4 | 0.18 | 78.1 | 0.86 | 28.9 | 3.2 | 8.2 | 248.1 | 120 | 6.0 | 16.4 | 144.2 | Ls | V | + | ||||
| SoiS | 1458 | 4.0 | 8.2 | 3.2 | 0.61 | 7.9 | 0.27 | 13.4 | 0.33 | 29.8 | 3.9 | 8.3 | 222.1 | 112 | 7.3 | 15.0 | 78.7 | Ma/Ls | V | + | ||||
| GruO | 1474 | 3.5 | 6.9 | 5.0 | 0.20 | 8.3 | 0.13 | 60.8 | 0.10 | 26.5 | 1.4 | 8.5 | 194.5 | 112 | 14.1 | 17.5 | 31.3 | Ls | B | |||||
| GaiU | 1508 | 3.5 | 4.1 | 3.8 | 0.25 | 12.0 | 0.29 | 9.6 | 0.18 | 16.4 | 10.1 | 8.9 | 198.3 | 72 | 13.4 | 16.3 | 21.3 | Do | A | + | ||||
| SoiN | 1520 | 0.3 | 4.7 | 4.2 | 0.77 | 10.8 | 0.24 | 13.9 | 0.37 | 36.3 | 6.8 | 8.2 | 251.8 | 115 | 5.4 | 13.4 | 72.5 | Mi | V | + | ||||
| SoE | 1552 | 3.0 | 5.5 | 4.5 | 0.61 | 3.9 | 0.18 | 49.3 | 0.10 | 31.1 | 6.6 | 8.8 | 177.7 | 123 | 7.5 | 15.0 | 168.2 | Ls | K | |||||
| SoW | 1558 | 3.0 | 11.5 | 7.0 | 0.36 | 4.5 | 0.16 | 82.9 | 0.10 | 30.2 | 6.3 | 8.3 | 204.2 | 91 | 14.7 | 14.5 | 97.8 | Ls | K | + | + | |||
| DelN | 1600 | 0.6 | 1.3 | 1.3 | 0.37 | 5.7 | 0.38 | 15.1 | 0.39 | 35.1 | 2.8 | 8.2 | 223.3 | 81 | 11.9 | 12.6 | 67.7 | Ls | K | |||||
| DelS | 1600 | 0.2 | 4.2 | 4.2 | 0.45 | 4.6 | 0.23 | 16.6 | 0.22 | 40.5 | 5.0 | 8.8 | 177.2 | 67 | 9.3 | 14.1 | 101.9 | Ls | K | |||||
| Hoer | 1601 | 0.5 | 1.8 | 1.8 | 0.34 | 17.3 | 0.21 | 20.4 | 0.37 | 24.7 | 1.0 | 7.9 | 225.2 | 90 | 11.8 | 15.3 | 21.1 | Mi | A | + | ||||
| Fun | 1601 | 2.5 | 4.5 | 3.5 | 0.03 | 10.3 | 0.13 | 44.3 | 0.10 | 31.3 | 4.3 | 8.4 | 274.4 | 91 | 10.3 | 14.9 | 7.0 | Ls | B | + | ||||
| Seeb | 1657 | 6.3 | 18.4 | 6.9 | 0.52 | 4.5 | 0.14 | 23.3 | 0.14 | 27.0 | 4.4 | 8.8 | 165.9 | 82 | 6.8 | 13.8 | 121.6 | Ls | M | |||||
| Scha | 1680 | 3.0 | 4.4 | 3.8 | 0.49 | 7.2 | 0.17 | 19.8 | 0.10 | 27.1 | 4.9 | 8.7 | 167.1 | 114 | 12.6 | 16.8 | 71.2 | Ls | W | |||||
| Gug | 1725 | 0.1 | 1.9 | 1.9 | 0.29 | 4.6 | 0.23 | 10.6 | 0.86 | 15.0 | 10.3 | 9.0 | 207.3 | 100 | 11.8 | 13.2 | 64.7 | Do | A | |||||
| GaiO | 1769 | 0.8 | 2.9 | 2.7 | 0.15 | 3.7 | 0.24 | 25.3 | 0.18 | 14.6 | 7.0 | 8.5 | 201.0 | 114 | 9.5 | 16.3 | 46.8 | Do | A | |||||
| Zie | 1799 | 3.0 | 15.1 | 5.0 | 0.14 | 7.6 | 0.15 | 26.7 | 0.10 | 22.5 | 1.6 | 8.2 | 219.8 | 107 | 6.7 | 13.7 | 21.5 | Ls | R | |||||
| Seel | 1809 | 0.4 | 5.4 | 5.4 | 0.59 | 7.9 | 0.12 | 31.5 | 0.10 | 23.8 | 1.0 | 8.7 | 134.2 | 93 | 12.4 | 14.9 | 79.3 | Ls | B | |||||
| Eis | 1827 | 0.7 | 3.9 | 3.9 | 0.24 | 1.1 | 0.21 | 23.3 | 0.36 | 16.0 | 3.6 | 8.2 | 192.7 | 96 | 6.4 | 10.4 | 247.6 | Ma | A | |||||
| Dra | 1874 | 5.3 | 20.7 | 10.3 | 0.30 | 4.0 | 0.19 | 19.6 | 0.10 | 26.8 | 3.1 | 8.6 | 157.7 | 97 | 4.9 | 11.9 | 79.9 | Ls | M | |||||
| Eng | 1876 | 3.0 | 17.3 | 10.4 | 0.04 | 4.7 | 0.59 | 27.0 | 0.31 | 19.8 | 7.2 | 8.2 | 235.6 | 89 | 4.9 | 11.1 | 14.4 | Do | A | + | ||||
| Bre | 1903 | 1.5 | 6.2 | 6.2 | 0.37 | 6.6 | 0.12 | 14.0 | 0.10 | 26.7 | 2.8 | 8.6 | 150.6 | 90 | 4.6 | 11.4 | 58.5 | Ls | M | |||||
| Stu | 1921 | 3.0 | 5.1 | 5.1 | 0.25 | 7.7 | 0.12 | 19.6 | 0.10 | 23.3 | 1.6 | 8.8 | 126.0 | 112 | 12.4 | 16.7 | 35.4 | Ls | W | |||||
| Lauf | 2012 | 0.8 | 5.6 | 3.7 | 0.13 | 3.8 | 0.22 | 24.9 | 0.19 | 12.4 | 6.3 | 8.3 | 168.2 | 99 | 9.4 | 14.7 | 39.1 | Do | A | |||||
| Rap | 2047 | 2.3 | 7.8 | 5.0 | 0.04 | 9.2 | 0.17 | 15.9 | 0.23 | 21.2 | 9.3 | 8.6 | 205.1 | 101 | 11.0 | 15.6 | 6.5 | Do/Ma | A | |||||
| Grub | 2060 | 0.5 | 3.5 | 3.2 | 0.10 | 17.0 | 0.15 | 17.8 | 0.18 | 37.9 | 2.6 | 8.6 | 222.4 | 73 | 9.4 | 13.3 | 6.9 | Ls | R | |||||
| SeeU | 2224 | 2.4 | 1.7 | 1.4 | 0.07 | 2.5 | 0.25 | 21.4 | 0.99 | 19.8 | 9.1 | 8.8 | 208.6 | 113 | 16.1 | 17.0 | 35.7 | Do/Ma | L | |||||
| Adl | 2294 | 1.9 | 1.7 | 1.7 | 0.15 | 1.0 | 0.13 | 30.3 | 0.32 | 19.4 | 9.2 | 8.5 | 159.6 | 94 | 14.0 | 14.7 | 183.3 | Do | L | + | ||||
| Schi | 2300 | 2.0 | 5.5 | 3.3 | 0.34 | 1.0 | 0.14 | 45.7 | 0.29 | 19.8 | 10.1 | 8.3 | 164.7 | 47 | 7.2 | 13.1 | 390.4 | Do | L | |||||
| SeeM | 2424 | 0.5 | 4.1 | 4.1 | 0.20 | 1.5 | 0.13 | 67.8 | 0.14 | 17.6 | 9.9 | 8.7 | 137.8 | 122 | 10.6 | 11.5 | 173.2 | Do | L | |||||
| SeeO | 2469 | 1.6 | 13.1 | 5.8 | 0.11 | 7.3 | 0.18 | 67.9 | 0.23 | 15.4 | 5.5 | 8.6 | 121.4 | 71 | 5.5 | 12.9 | 24.8 | Do | L | |||||
| Response Share_rl | pShapiro | Model Type | Predictor | pmodel | R2adj | Res. Df | plm.moran | AIC |
|---|---|---|---|---|---|---|---|---|
| Sediment | 0.098 | LM | TP | 0.002 | 0.20 | 41 | 0.175 | |
| Littoral | 0.931 | LM | TP | 0.056 | 0.08 | 32 | 0.908 | |
| LM | TP + secchi | TP: 0.182 secchi: 0.029 | 0.19 | 31 | 0.923 | |||
| LM | secchi | 0.009 | 0.17 | 32 | 0.607 | |||
| Plankton | 0.02 | GAM | s(lon) + s(lat) + s(TP) | lon: <0.001 lat: 0.117 TP: 0.327 | 0.38 | 19 | −38.4 | |
| GAM | s(lon) | <0.001 | 0.34 | 27 | −41.3 |
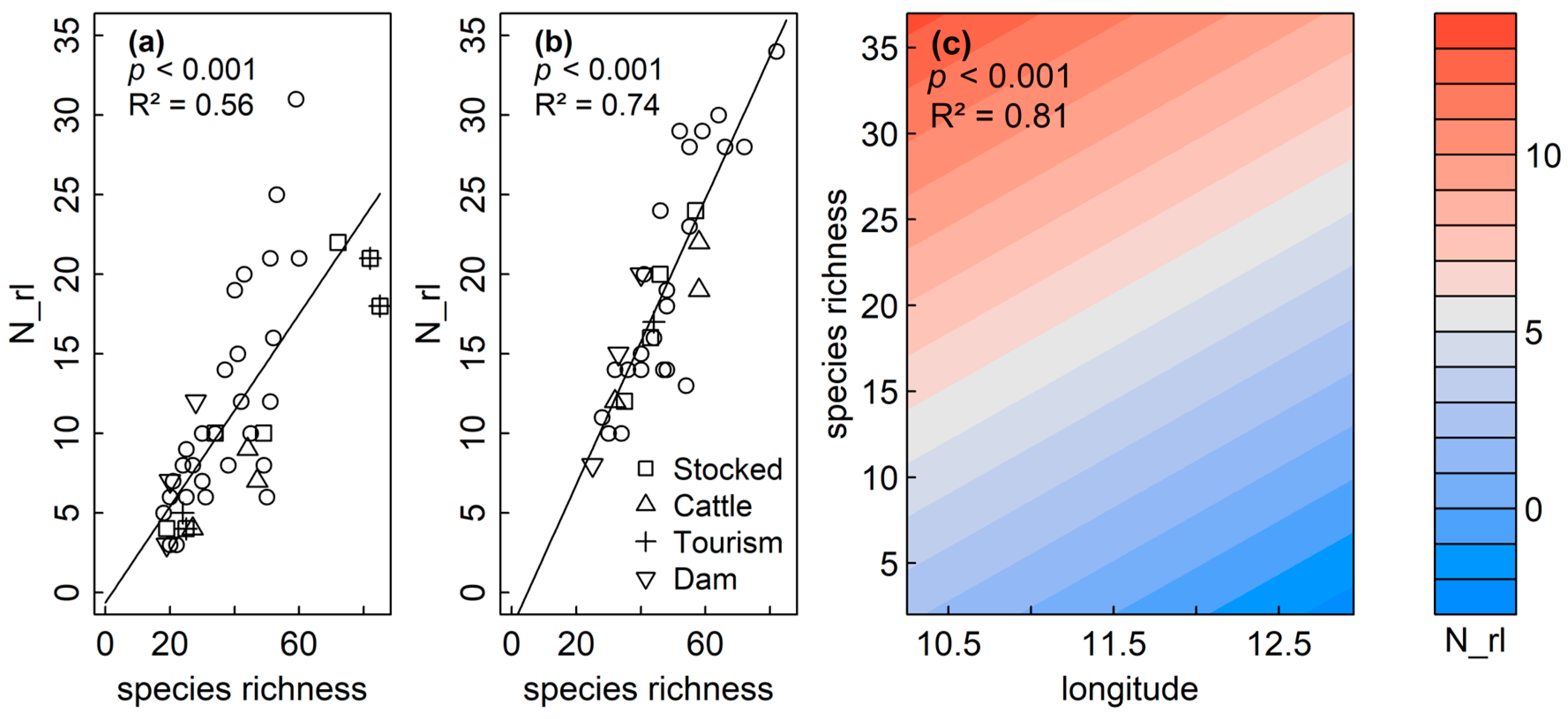
| Response N_rl | pShapiro | Model Type | Predictor | pmodel | R2adj | Res. Df | plm.moran | AIC |
|---|---|---|---|---|---|---|---|---|
| Sediment | <0.001 | GLM | species | <0.001 | 0.56 | 41 | ||
| Littoral | 0.083 | LM | species | <0.001 | 0.74 | 32 | 0.219 | |
| Plankton | 0.092 | GAM | s(lon) + s(lat) + s(species) | lon: 0.008 lat: 0.826 species: <0.001 | 0.80 | 28 | −63.00 | |
| GAM | s(lon) + s(species) | lon: <0.001 species: <0.001 | 0.81 | 29 | −63.02 |
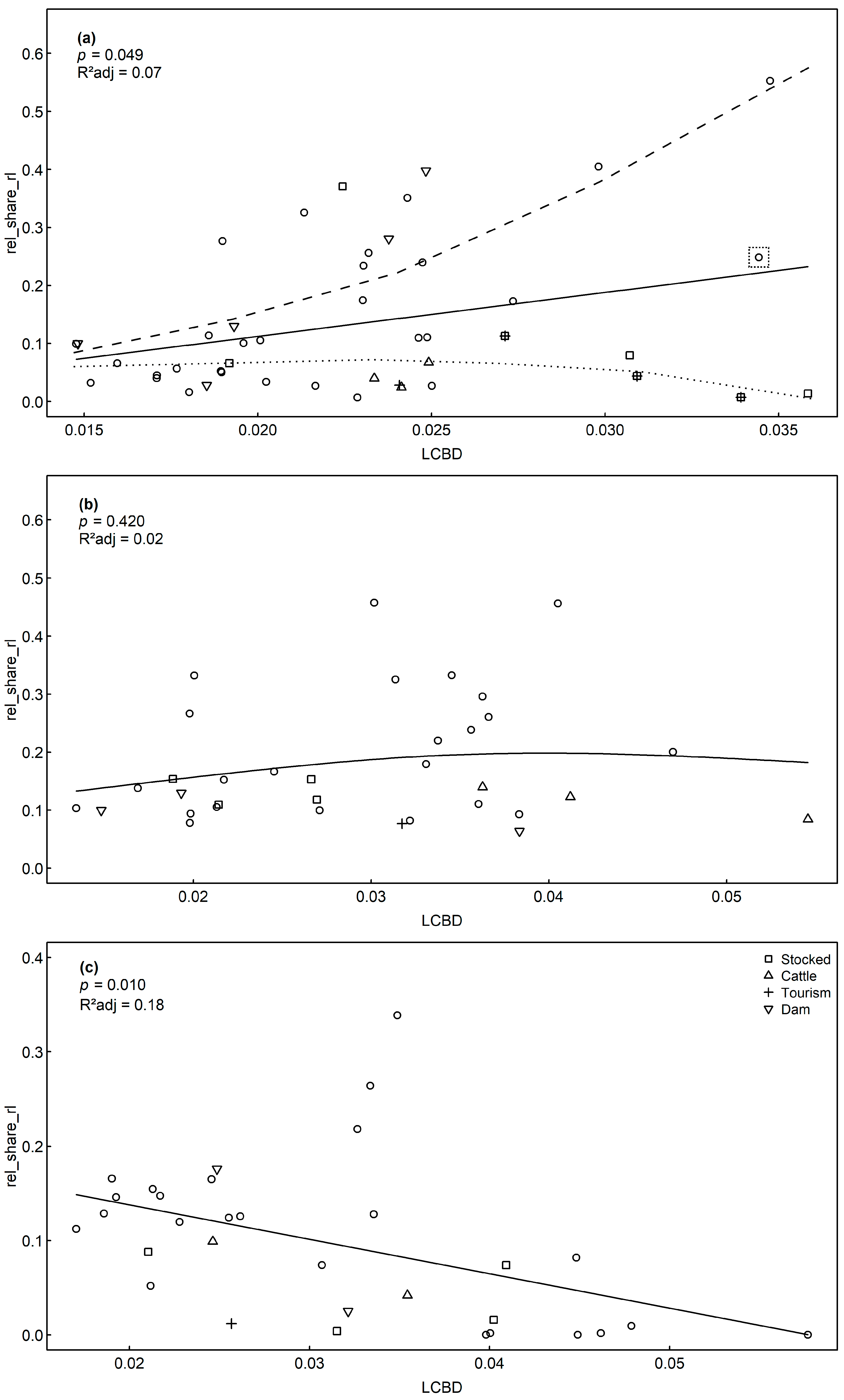
| Response Rel_Share_rl | Model Type | Predictor | pmodel | R2adj | Res. Df |
|---|---|---|---|---|---|
| Sediment | GAM | LCBD | 0.049 | 0.07 | 41 |
| Littoral | GAM | LCBD | 0.420 | 0.02 | 32 |
| Plankton | GAM | LCBD | 0.010 | 0.18 | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ossyssek, S.; Hofmann, A.M.; Geist, J.; Raeder, U. Diatom Red List Species Reveal High Conservation Value and Vulnerability of Mountain Lakes. Diversity 2022, 14, 389. https://doi.org/10.3390/d14050389
Ossyssek S, Hofmann AM, Geist J, Raeder U. Diatom Red List Species Reveal High Conservation Value and Vulnerability of Mountain Lakes. Diversity. 2022; 14(5):389. https://doi.org/10.3390/d14050389
Chicago/Turabian StyleOssyssek, Stefan, Andrea Maria Hofmann, Juergen Geist, and Uta Raeder. 2022. "Diatom Red List Species Reveal High Conservation Value and Vulnerability of Mountain Lakes" Diversity 14, no. 5: 389. https://doi.org/10.3390/d14050389
APA StyleOssyssek, S., Hofmann, A. M., Geist, J., & Raeder, U. (2022). Diatom Red List Species Reveal High Conservation Value and Vulnerability of Mountain Lakes. Diversity, 14(5), 389. https://doi.org/10.3390/d14050389