
Morphostructural Characterization of the Heterogeneous Rhodolith Bed at the Marine Protected Area “Capo Carbonara” (Italy) and Hydrodynamics

 ,
, 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
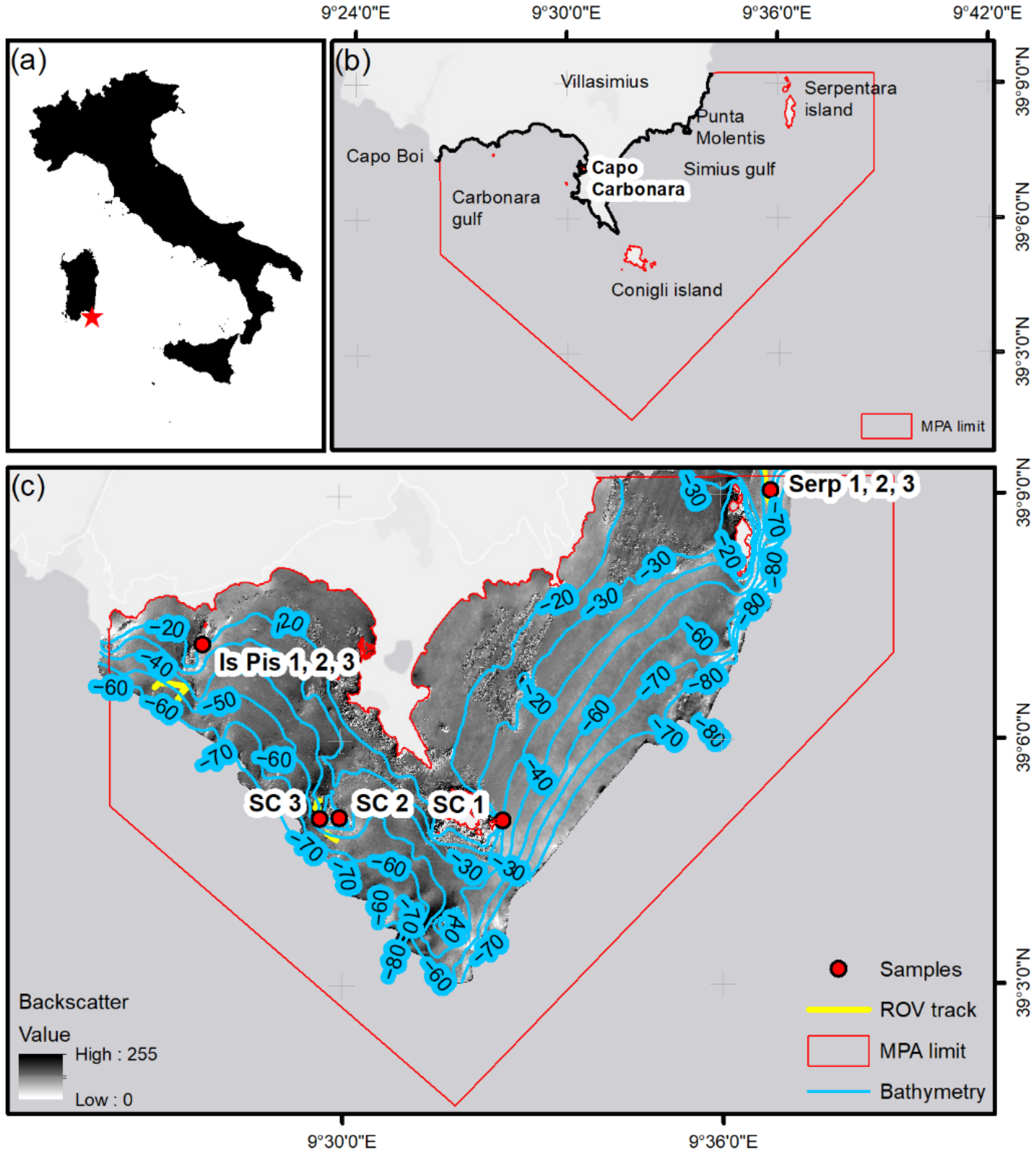
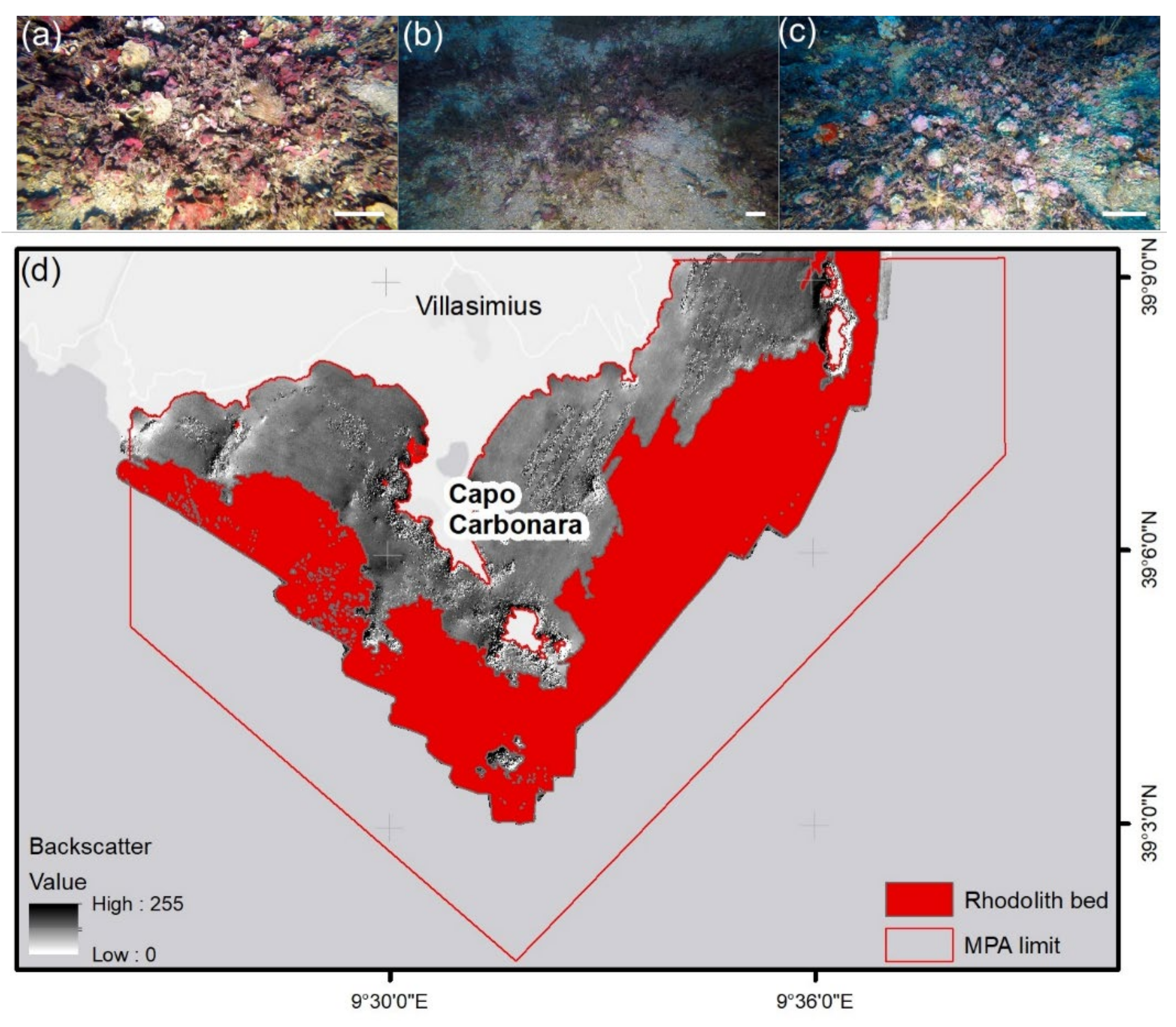
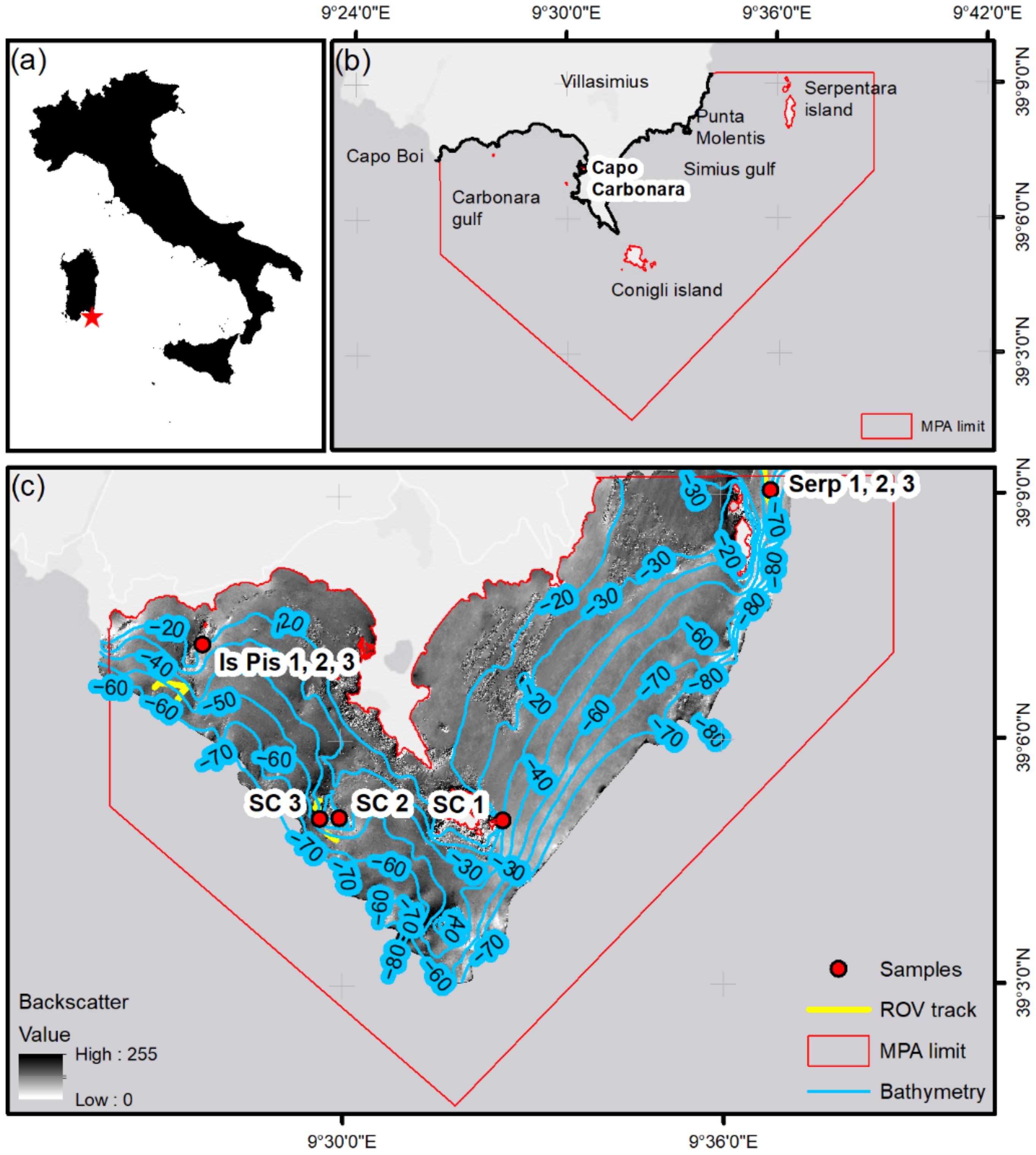
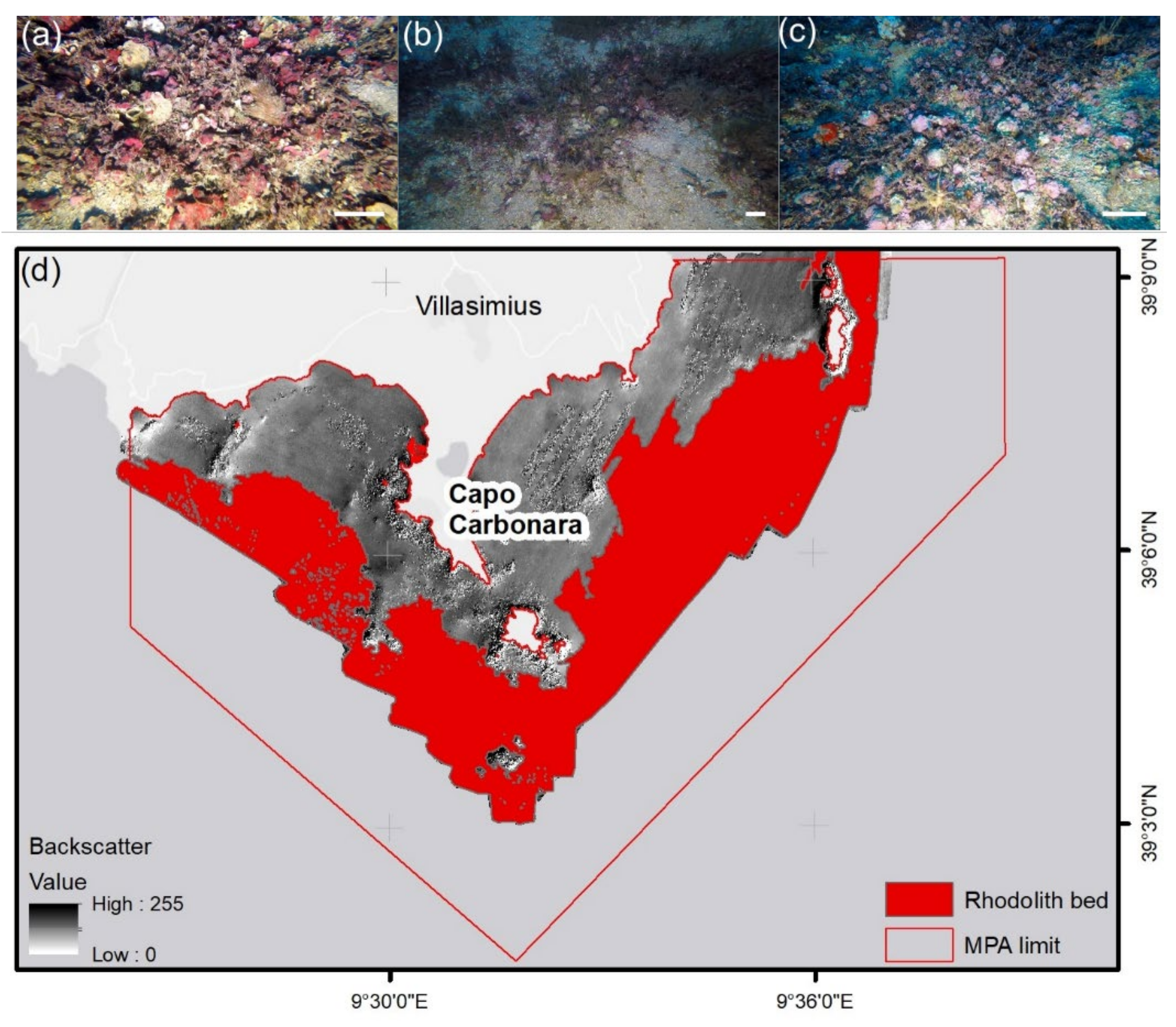
2. Study Site
3. Materials and Methods
3.1. Sampling
3.2. Statistical Analysis
3.3. Environmental Data
4. Results
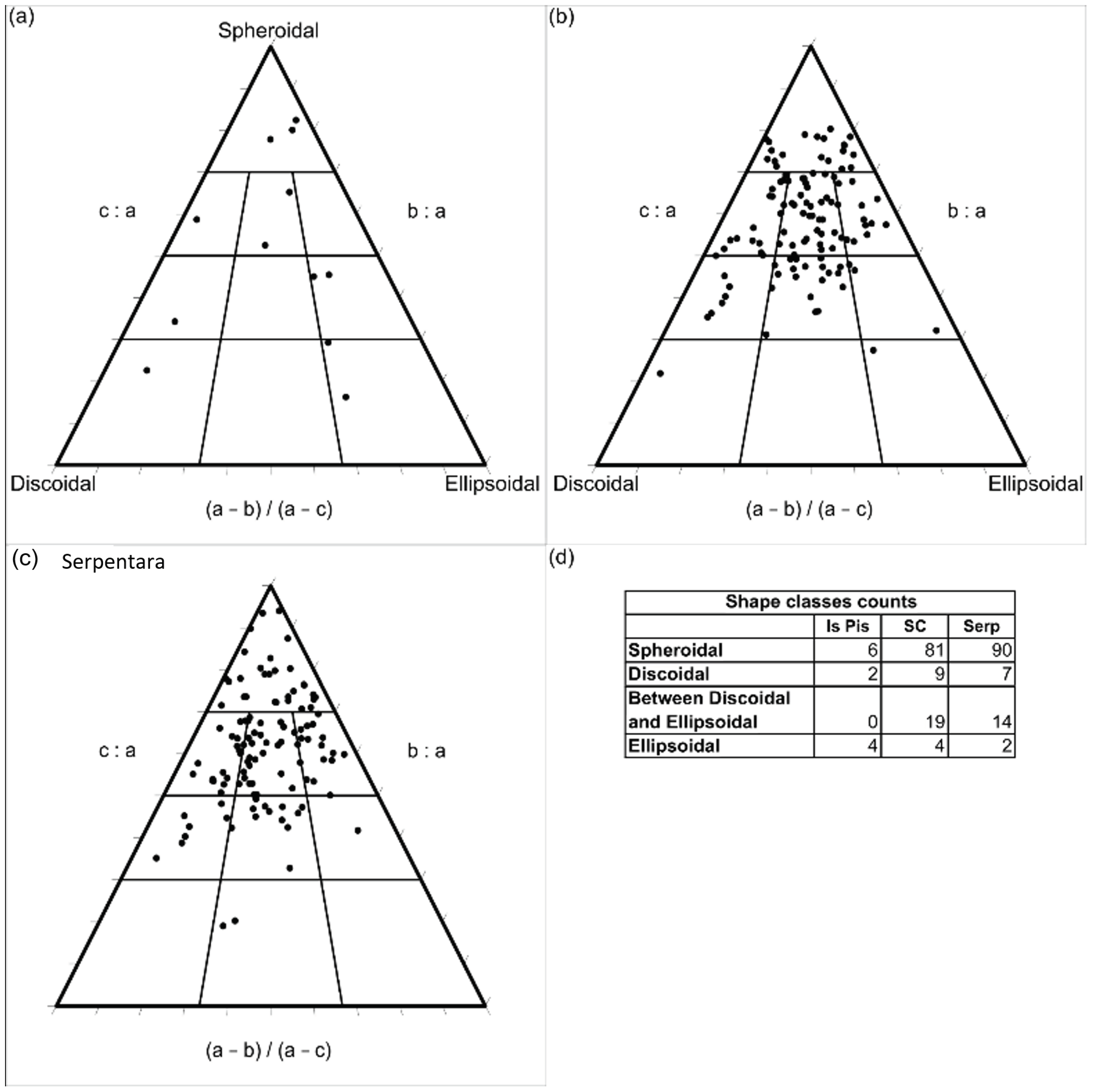
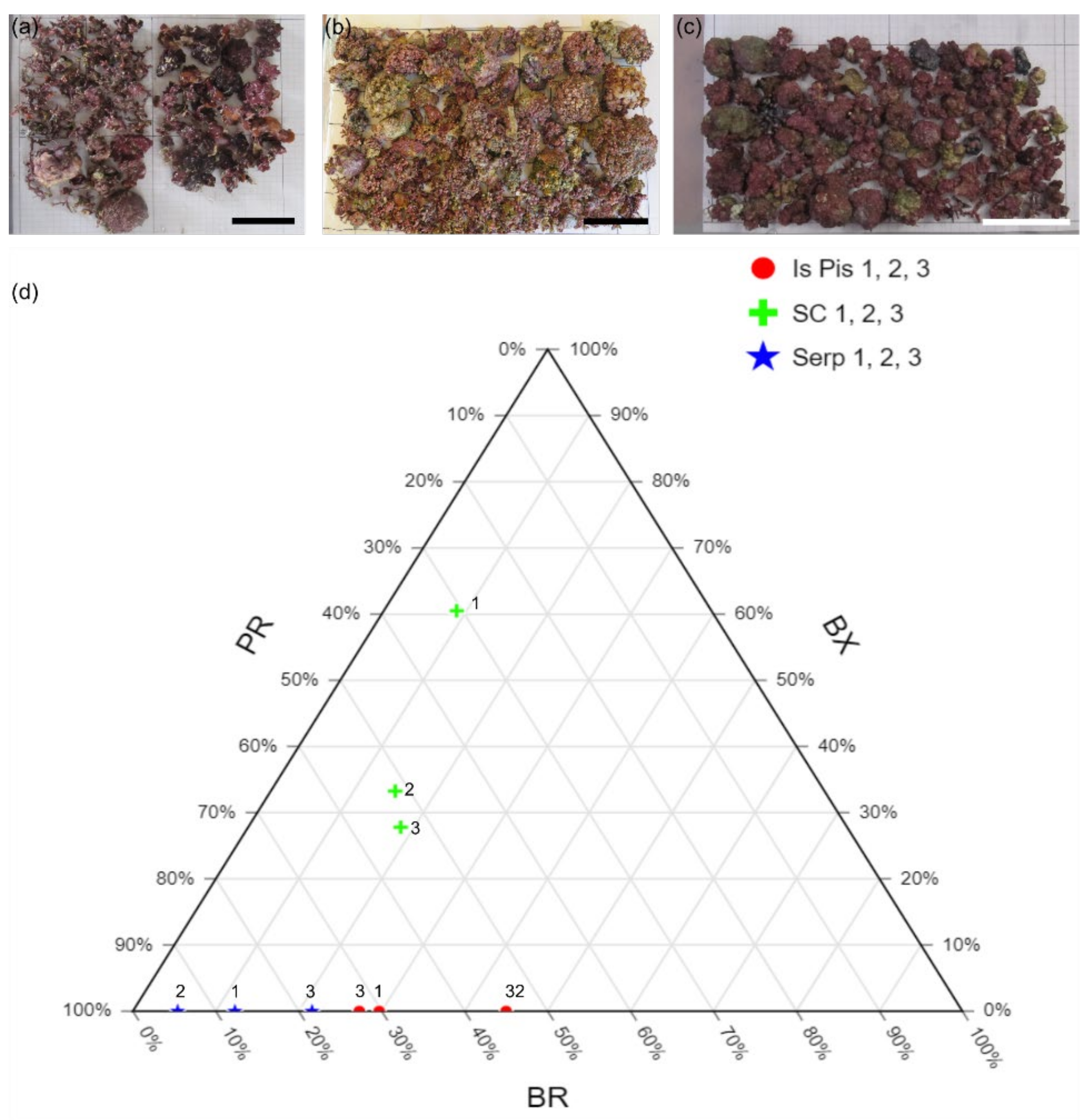
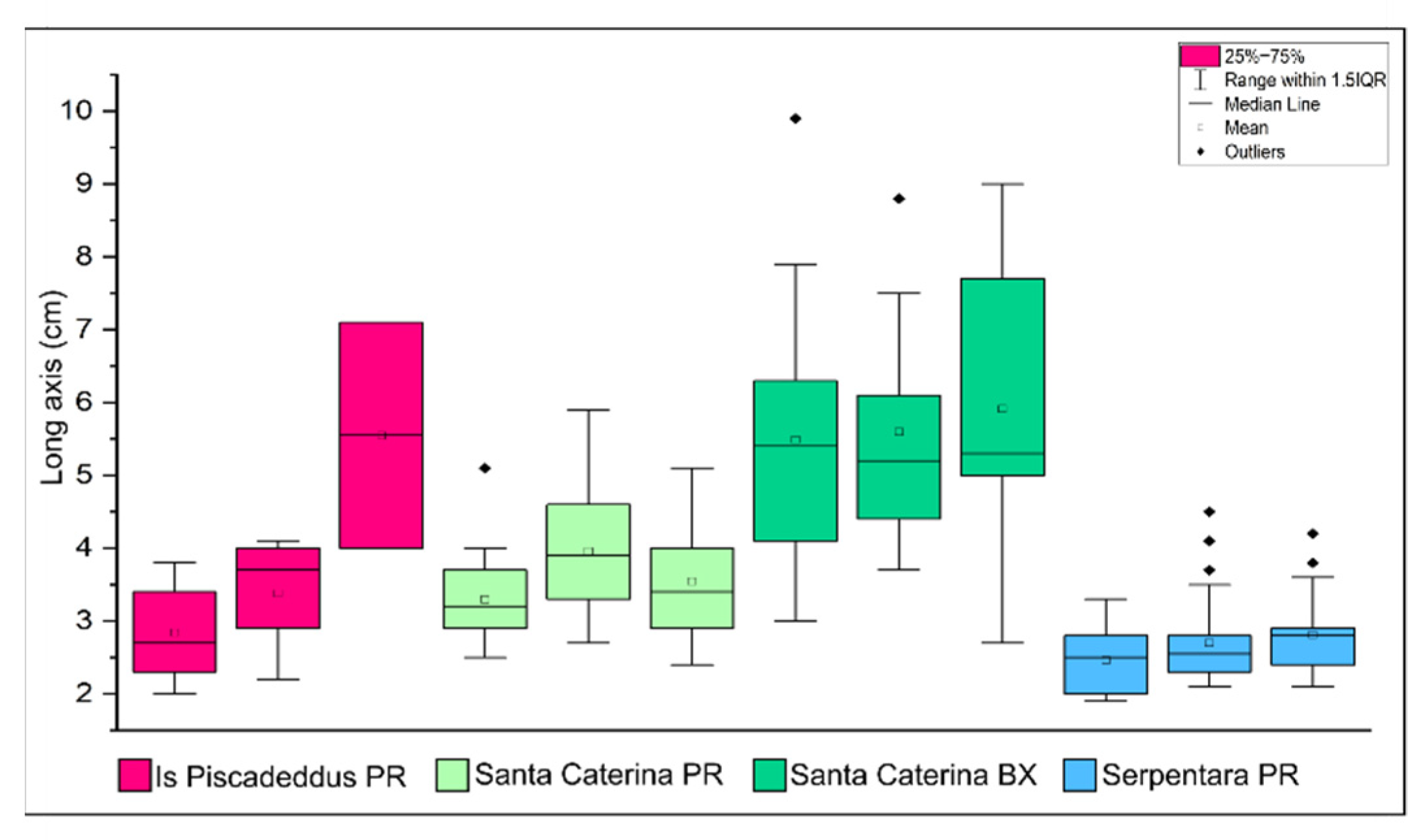
4.1. Sampling
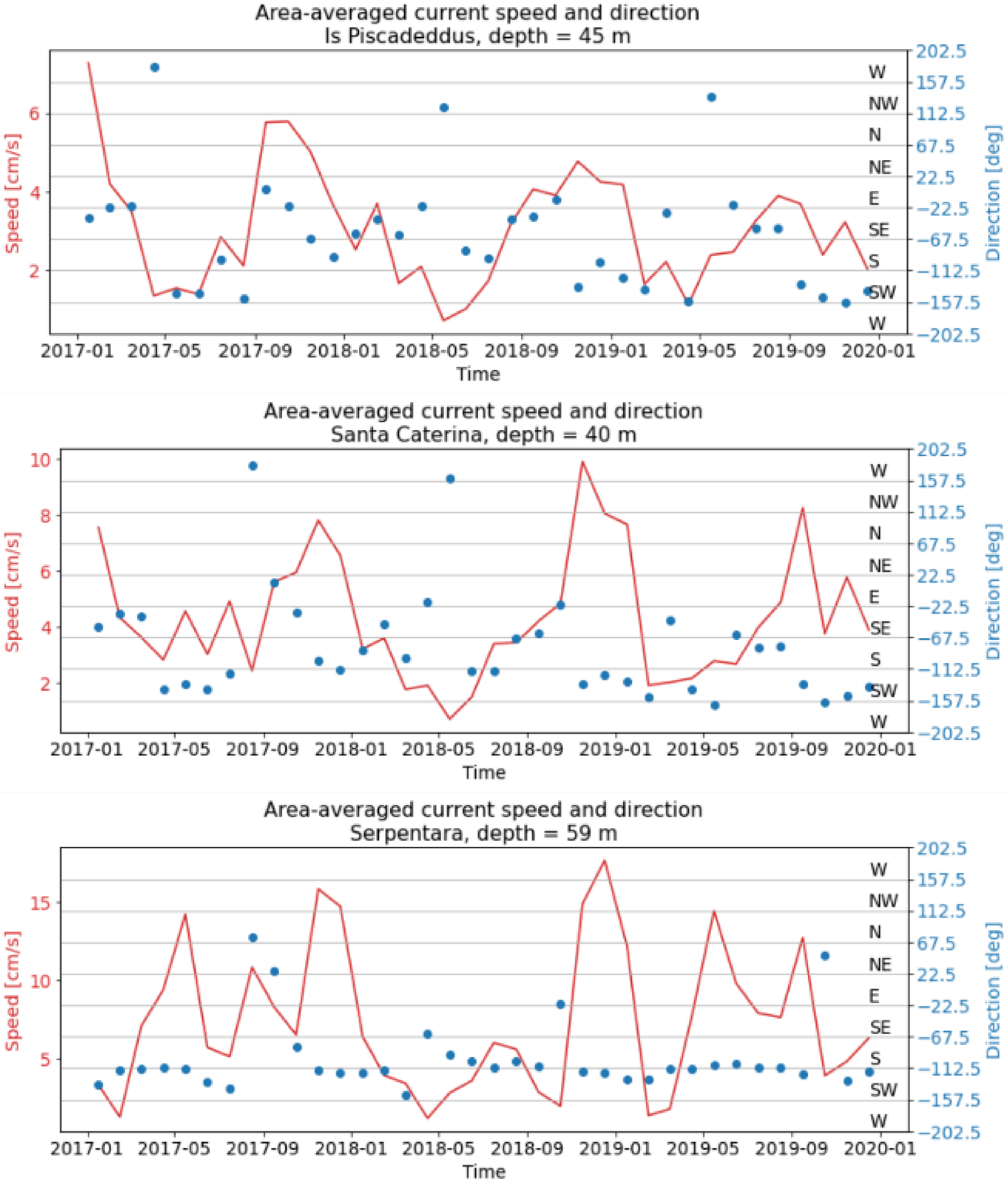
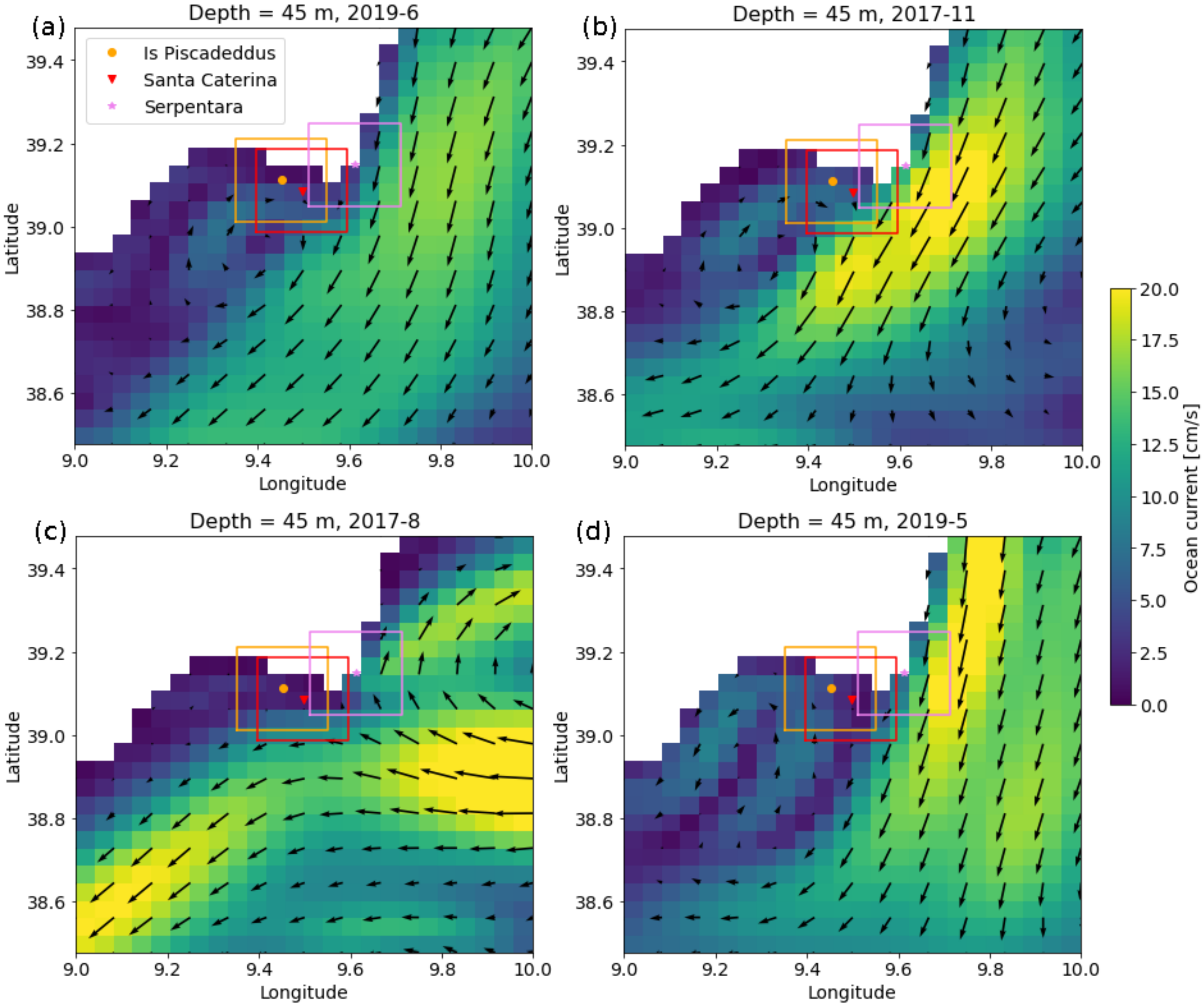
4.2. Environmental Data
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosence, D.W.J. Description and classification of rhodoliths (rhodoids, rhodolites). In Coated Grains; Peryt, T.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 217–224. [Google Scholar]
- Steneck, R.S. The ecology of coralline algae crusts: Convergent patterns and adaptive strategies. Ann. Rev. Ecol. Syst. 1986, 17, 273–303. [Google Scholar] [CrossRef]
- Bosence, D.W.J. The occurrence and ecology of recent rhodoliths—A review. In Coated Grains; Peryt, T.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 225–242. [Google Scholar]
- Woelkerling, J.; Irvine, L.M.; Harvey, A.S. Growth forms in non-geniculate coralline red algae (Corallinales, Rhodophyta). Aust. Syst. Bot. 1993, 6, 277–293. [Google Scholar] [CrossRef]
- Freiwald, A.; Henrich, R. Reefal coralline algal build-ups within the Arctic Circle: Morphology and sedimentary dynamics under extreme environmental seasonality. Sedimentology 1994, 41, 963–984. [Google Scholar] [CrossRef]
- Bosence, D.W.J. Ecological studies on two unattached coralline algae from Western Ireland. Palaeontology 1976, 19, 365–395. [Google Scholar]
- Basso, D. Deep rhodolith distribution in the Pontian Islands, Italy: A model for the paleoecology of a temperate sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 137, 173–187. [Google Scholar] [CrossRef]
- Braga, J.C.; Martín, J.M. Neogene coralline-algal growth-forms and their palaeoenvironments in the Almanzora River Valley (Almeria, S.E. Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 67, 285–303. [Google Scholar] [CrossRef]
- Aguirre, J.; Braga, J.C.; Martín, J.M. Algal nodules in the upper Pliocene deposits at the coast of Cadiz (S Spain). Boll. Soc. Paleontol. Ital. 1993, 1, 1–7. [Google Scholar]
- Steller, D.L.; Foster, M.S. Environmental factors influencing distribution and morphology of rhodoliths on Bahía Concepción, B.CS, México. J. Exp. Mar. Biol. Ecol. 1995, 194, 201–212. [Google Scholar] [CrossRef]
- BIOMAERL Team. Conservation and management of northeast Atlantic and Mediterranean maërl beds. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, S65–S76. [Google Scholar] [CrossRef]
- Bordehore, C.; Borg, J.A.; Lanfranco, E.; Ramos-Esplá, A.A.; Rizzo, F.; Schembri, P.J. Trawling as a major threat to Mediterranean maërl beds. Proceedings of the First Mediterranean symposium on Marine Vegetation, Ajaccio. Mednature 2002, 1, 105–109. [Google Scholar]
- Bordehore, C.; Ramos-Esplá, A.A.; Riosmena-Rodríguez, R. Comparative study of two maërl beds with different otter trawling history, southeast Iberian Peninsula. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, S43–S54. [Google Scholar] [CrossRef]
- Bahia, R.G.; Abrantes, D.P.; Brasileiro, P.S.; Pereira-Filho, G.H.; Amado-Filho, G.M. Rhodolith bed structure along a depth gradient on the northern coast of Bahia State, Brazil. Braz. J. Oceanog. 2010, 58, 323–337. [Google Scholar] [CrossRef]
- Riul, P.; Targino, C.H.; Farias, J.D.N.; Visscher, P.T.; Horta, P.A. Decrease of Lithothamnion sp. (Rhodophyta) primary production due to the deposition of a thin sediment layer. J. Mar. Biol. Ass. UK 2008, 88, 17–19. [Google Scholar]
- Bosellini, A.; Ginsburg, R.N. Form and internal structure of recent algal nodules (rhodolites), Bermuda. J. Geol. 1971, 79, 669–682. [Google Scholar] [CrossRef]
- Foster, M.S.; Riosmena-Rodríguez, R.; Steller, D.L.; Woelkerling, W.J. Living rhodolith beds in the Gulf of California and their implications for paleoenvironmental interpretation. Geol. Soc. Am. Bull. 1997, 318, 127–139. [Google Scholar]
- Steller, D.L.; Riosmena-Rodríguez, R.; Foster, M.S.; Roberts, C.A. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of disturbance. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, S5–S20. [Google Scholar] [CrossRef]
- Barberá, C.; Moranta, J.; Ordines, F.; Ramón, M.; de Mesa, A.; Díaz-Valdés, M.; Grau, A.M.; Massutí, E. Biodiversity and habitat mapping of Menorca Channel (western Mediterranean): Implications for conservation. Biodivers. Conserv. 2012, 21, 701–728. [Google Scholar] [CrossRef]
- Micallef, A.; Le Bas, T.; Huvenne, V.A.I.; Blondel, P.; Huhnerbach, V.; Deidun, A. A multi-method approach for benthic habitat mapping of shallow coastal areas with high-resolution multibeam data. Cont. Shelf Res. 2012, 39, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Sañé, E.; Chiocci, F.L.; Basso, D.; Martorelli, E. Environmental factors controlling the distribution of rhodoliths: An integrated study based on seafloor sampling, ROV and side scan sonar data, offshore the W-Pontine Archipelago. Cont. Shelf Res. 2016, 129, 10–22. [Google Scholar] [CrossRef]
- Bracchi, V.A.; Angeletti, L.; Marchese, F.; Taviani, M.; Cardone, F.; Hajdas, I.; Grande, V.; Prampolini, M.; Caragnano, A.; Corselli, C.; et al. A resilient deep-water rhodolith bed off the Egadi Archipelago (Mediterranean Sea) and its actual paleontological significance. Alp. Mediterr. Quat. 2019, 32, 1–20. [Google Scholar]
- Millar, K.R.; Gagnon, P. Mechanisms of stability of rhodolith beds: Sedimentological aspects. Mar. Ecol. Prog. Ser. 2018, 594, 65–83. [Google Scholar] [CrossRef] [Green Version]
- O’connell, L.G.; James, N.P.; Harvey, A.S.; Luick, J.; Bone, Y.; Shepherd, S.A. Reevaluation of the inferred relationship between living rhodolith morphologies, their movement, and water energy: Implications for interpreting palaeoceanographic conditions. Palaios 2021, 35, 543–556. [Google Scholar] [CrossRef]
- Marrack, E.C. The relationship between water motion and living rhodolith beds in the southwestern Gulf of California, Mexico. Palaios 1999, 14, 159–171. [Google Scholar] [CrossRef]
- Basso, D.; Tomaselli, V. Paleoecological potentiality of rhodoliths: A Mediterranean case history. Bollettino Soc. Paleontol. Ital. 1994, 2, 17–27. [Google Scholar]
- Harris, P.T.; Tsuji, Y.; Marshall, J.F.; Davies, P.J.; Honda, N.; Matsuda, H. Sand and rhodolith-gravel entrainment on the mid- to outer-shelf under a western boundary current: Fraser Island continental shelf, eastern Australia. Mar. Geol. 1996, 129, 313–330. [Google Scholar] [CrossRef]
- Basso, D.; Nalin, R.; Campbell, S.N. Shallow-water Sporolithon rhodoliths from north island (New Zealand). Palaios 2009, 24, 92–103. [Google Scholar] [CrossRef]
- Basso, D.; Babbini, L.; Kaleb, S.; Bracchi, V.; Falace, A. Monitoring deep Mediterranean rhodolith beds. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 26, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, R.; Pastor, X.; de la Torriente, A.; García, S. Deep-sea coralligenous beds observed with ROV on four seamounts in the Western Mediterranean. In UNEP—MAP—RAC/SPA, Proceedings of the 1st Mediterranean Symposium on the Conservation of the Coralligenous and Other Calcareous Bio-Concretions, Tabarka, Tunisia, 15–16 January 2009; Pergent-Martini, C., Brichet, M., Eds.; CAR/ASP Publishing: Tabarka, Tunisia, 2009; pp. 148–150. [Google Scholar]
- Riosmena-Rodríguez, R. Natural history of Rhodolith/Maërl beds: Their role in near-shore biodiversity and management. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Coastal Research Library; Springer International Publishing: Cham, Switzerland, 2017; Volume 15, pp. 3–26. [Google Scholar]
- Foster, M.S. Rhodoliths: Between rocks and soft places. J. Phycol. 2001, 37, 659–667. [Google Scholar] [CrossRef]
- Foster, M.S.; Amado Filho, G.M.; Kamenos, K.A.; Riosmena-Rodríguez, R.; Steller, D.L. Rhodoliths and rhodolith beds. In Research and Discoveries: The Revolution of Science Through SCUBA; American Academy of Underwater Sciences: Mobile, AL, USA, 2013; pp. 143–155. [Google Scholar]
- Basso, D.; Babbini, L.; Ramos-Esplá, A.A.; Salomidi, M. Mediterranean rhodolith beds. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Coastal Research Library; Springer: Cham, Switzerland, 2017; Volume 15, pp. 281–298. [Google Scholar]
- UNEP-MAP-RAC/SPA. Action Plan for the Conservation of the Coralligenous and Other Calcareous Bio-Concretions in the Mediterranean Sea; Pergent-Martini, C., Brichet, M., Eds.; RAC/SPA: Tunis, Tunisia, 2008; 21p. [Google Scholar]
- Orrù, P.; Cocco, A.; Panizza, V. Subaqueous geomorphological investigations from Capo Boi to Is Cappuccinus (south-eastern Sardinia). Mem. Descr. Carta Geol. d’Ital. 1994, LII, 163–176. [Google Scholar]
- Graham, D.J.; Midgley, N.G. Graphical representation of particle shape using triangular diagrams: An Excel spreadsheet method. Earth Surf. Proc. Landf. 2000, 25, 1473–1477. [Google Scholar] [CrossRef]
- Sneed, E.D.; Folk, R.L. Pebbles in the lower Colorado river, Texas. A study in particle morphogenesis. J. Geol. 1958, 66, 114–150. [Google Scholar] [CrossRef]
- Underwood, A.J.; Chapman, M.G.; Richards, S.A. GMAV-5 for Windows. An Analysis of Variance Programme; University of Sydney: Sidney, Australia, 2002. [Google Scholar]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Express: Cambridge, UK, 1997; Volume 117, p. 524. [Google Scholar]
- Escudier, R.; Clementi, E.; Omar, M.; Cipollone, A.; Pistoia, J.; Aydogdu, A.; Drudi, M.; Grandi, A.; Lyubartsev, V.; Lecci, R.; et al. Mediterranean Sea Physical Reanalysis (CMEMS MED-Currents) (Version 1) Set; Copernicus Monitoring Environment Marine Service (CMEMS): Toulouse, France, 2020. [Google Scholar]
- Chimienti, G.; Rizzo, L.; Kaleb, S.; Falace, A.; Fraschetti, S.; Giosa, F.D.; Tursi, A.; Barbone, E.; Ungaro, N.; Mastrototaro, F. Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea). J. Mar. Sci. Eng. 2020, 8, 813. [Google Scholar] [CrossRef]
- Rendina, F.; Kaleb, S.; Caragnano, A.; Ferrigno, F.; Appolloni, L.; Donnarumma, L.; Russo, G.F.; Sandulli, R.; Roviello, V.; Falace, A. Distribution and Characterization of Deep Rhodolith Beds off the Campania coast (SW Italy, Mediterranean Sea). Plants 2020, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Adey, W.H.; Macintyre, I.G. Crustose coralline algae: A re-evaluation in the geological sciences. Geol. Soc. Am. Bull. 1973, 84, 883–904. [Google Scholar] [CrossRef]
- Littler, M.M.; Littler, D.S.; Hanisak, M.D. Deep-water rhodolith distribution, productivity and growth history at sites of formation and subsequent degradation. J. Mar. Biol. Ecol. 1991, 150, 163–182. [Google Scholar] [CrossRef]
- Riul, P.; Lacouth, P.; Pagliosa, P.R.; Christoffersen, M.L.; Horta, P. Rhodolith beds at the Easter- most extreme of South America: Community structure on an endangered environment. Aquat. Bot. 2009, 90, 315–320. [Google Scholar] [CrossRef]
- Amado-Filho, G.M.; Maneveldt, G.; Manso, R.C.C.; Marins-Rosa, B.V.; Pacheco, M.R.; Guimarães, S.M.P.B. Structure of rhodolith beds from 4 to 55 m deep along the southern coast of Espírito Santo State, Brazil. Cienc. Mar. 2007, 33, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, P.; Matheson, K.; Stapleton, M. Variation in rhodolith morphology and biogenic potential of newly discovered rhodolith beds in Newfoundland and Labrador (Canada). Bot. Mar. 2012, 55, 85–99. [Google Scholar] [CrossRef]
- Minnery, G.A.; Rezak, R.; Bright, T.J. Depth zonation and growth form of crustose coralline algae: Flower Garden Banks, northwestern Gulf of Mexico. In Paleoalgology: Contemporary Research and Applications; Toomey, D.F., Nitecki, M.H., Eds.; Springer: Berlin, Germany, 1985; pp. 237–247. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Year | Coordinates | Depth (m) | Type |
|---|---|---|---|---|
| Is Piscadeddus 1 | 2018 | 39.112; 9.4518 | 45 | Van Veen grab |
| Is Piscadeddus 2 | 2018 | 39.112; 9.4518 | ||
| Is Piscadeddus 3 | 2018 | 39.1074, 9.4558 | ||
| Santa Caterina 1 | 2017 | 39.0865, 9.4966 | 40 | Scuba diver |
| Santa Caterina 2 | 2017 | 39.0863, 9.4964 | ||
| Santa Caterina 3 | 2017 | 39.0861, 9.4963 | ||
| Serpentara 1 | 2019 | 39.1493, 9.6124 | 59 | Scuba diver |
| Serpentara 2 | 2019 | 39.1493, 9.6124 | ||
| Serpentara 3 | 2019 | 39.1493, 9.6124 |
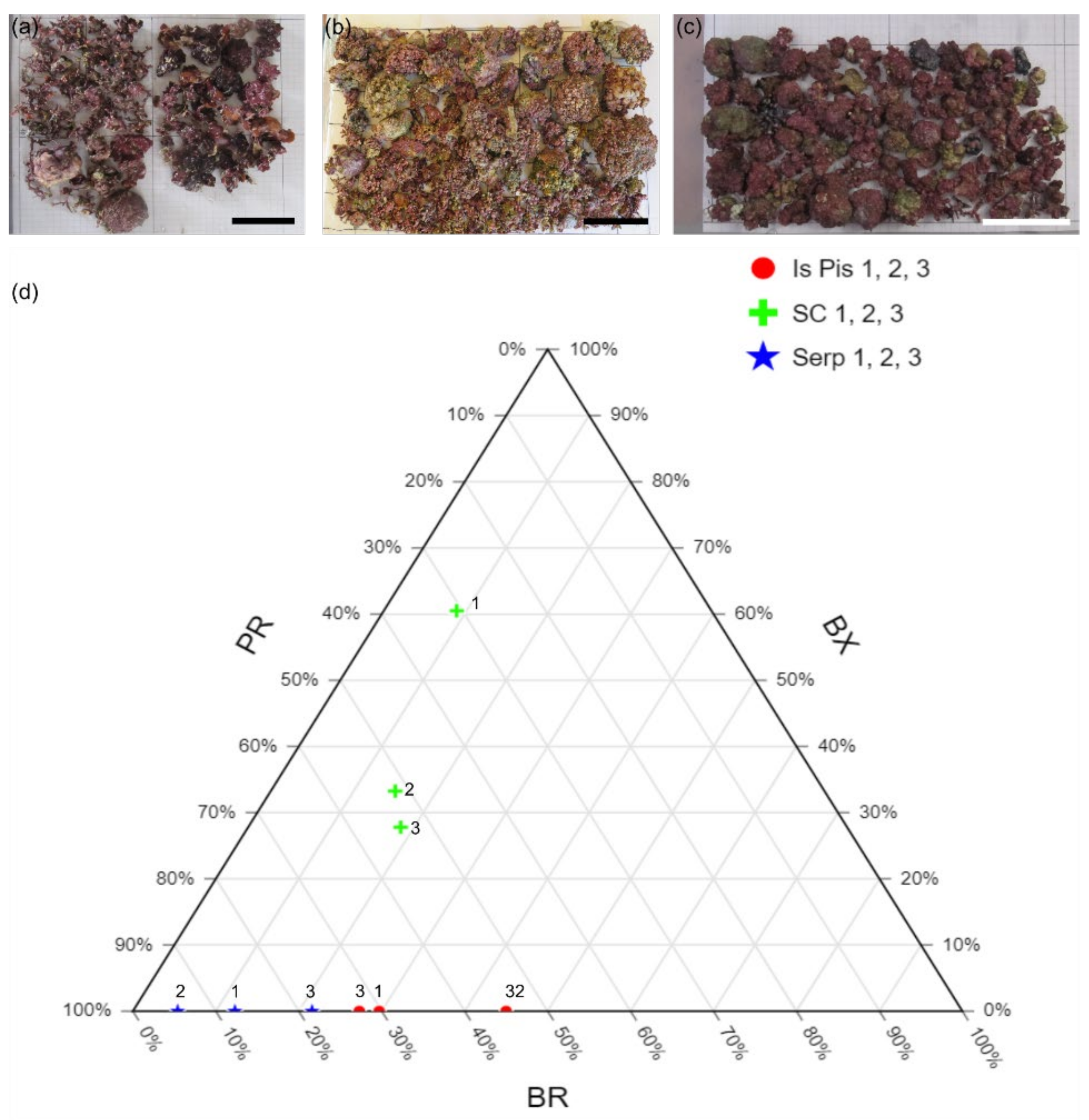
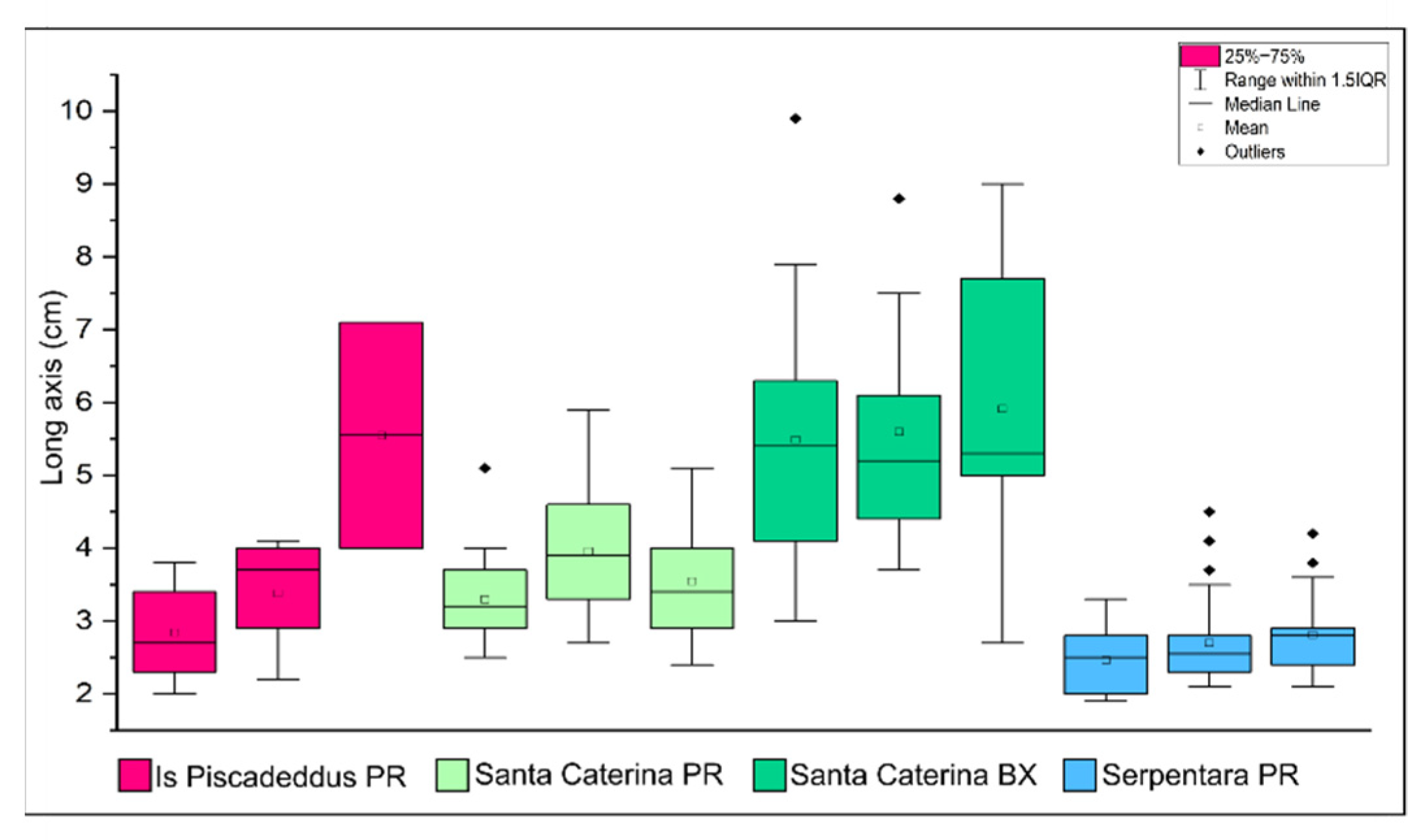
| Sample | BX (cm2) | PR (cm2) | Morphotype (%) | Rhodoliths Coverage | Grain Size (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| <2 cm | >2 cm | <2 cm | >2 cm | BX | PR | BR | cm2 | % | G | S | M | |
| Is Piscadeddus 1 | 0 | 0 | 19.7 | 34 | 0 | 70.3 | 29.7 | 76.4 | 6.95 | 11.7 | 72.9 | 15.4 |
| Is Piscadeddus 2 | 0 | 0 | 24.7 | 26.7 | 0 | 55 | 45 | 93.5 | 8.50 | 12.3 | 73.5 | 14.2 |
| Is Piscadeddus 3 | 0 | 0 | 27.4 | 24.6 | 0 | 72.7 | 27.3 | 71.5 | 6.50 | 7.98 | 76.1 | 16 |
| Santa Caterina 1 | 0 | 535 | 135 | 137 | 60.5 | 30.7 | 8.8 | 884 | 55.25 | 26.5 | 72.4 | 1.12 |
| Santa Caterina 2 | 0 | 214 | 45.4 | 347 | 33.2 | 51.7 | 15 | 642.7 | 40.17 | 29.3 | 70.5 | 0.2 |
| Santa Caterina 3 | 0 | 201 | 149 | 240 | 27.8 | 53.8 | 18.4 | 723.48 | 45.22 | 32 | 67.5 | 0.46 |
| Serpentara 1 | 0 | 0 | 109 | 84.8 | 0 | 87.7 | 12.3 | 220.6 | 13.79 | 35.3 | 59.1 | 5.56 |
| Serpentara 2 | 0 | 0 | 297 | 184 | 0 | 94.6 | 5.4 | 507.9 | 31.74 | 50.7 | 43.7 | 5.57 |
| Serpentara 3 | 0 | 0 | 234 | 113 | 0 | 78.4 | 21.6 | 442.4 | 27.65 | 36.5 | 57 | 6.52 |
| Sites | T (°C) | Salinity (psu) | Current Velocity (cm/s) | ||||
|---|---|---|---|---|---|---|---|
| Max | Min | Max | Min | Max | Min | Mean | |
| Is Piscadeddus | 19.9 | 13.9 | 38.5 | 37.9 | 7.3 | 0.7 | 3.1 |
| Santa Caterina | 21.1 | 13.9 | 38.5 | 37.9 | 9.9 | 0.7 | 4.3 |
| Serpentara | 17.5 | 13.9 | 38.5 | 37.9 | 17.7 | 1.2 | 7.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracchi, V.A.; Caronni, S.; Meroni, A.N.; Burguett, E.G.; Atzori, F.; Cadoni, N.; Marchese, F.; Basso, D. Morphostructural Characterization of the Heterogeneous Rhodolith Bed at the Marine Protected Area “Capo Carbonara” (Italy) and Hydrodynamics. Diversity 2022, 14, 51. https://doi.org/10.3390/d14010051
Bracchi VA, Caronni S, Meroni AN, Burguett EG, Atzori F, Cadoni N, Marchese F, Basso D. Morphostructural Characterization of the Heterogeneous Rhodolith Bed at the Marine Protected Area “Capo Carbonara” (Italy) and Hydrodynamics. Diversity. 2022; 14(1):51. https://doi.org/10.3390/d14010051
Chicago/Turabian StyleBracchi, Valentina A., Sarah Caronni, Agostino N. Meroni, Esteban Gottfried Burguett, Fabrizio Atzori, Nicoletta Cadoni, Fabio Marchese, and Daniela Basso. 2022. "Morphostructural Characterization of the Heterogeneous Rhodolith Bed at the Marine Protected Area “Capo Carbonara” (Italy) and Hydrodynamics" Diversity 14, no. 1: 51. https://doi.org/10.3390/d14010051
APA StyleBracchi, V. A., Caronni, S., Meroni, A. N., Burguett, E. G., Atzori, F., Cadoni, N., Marchese, F., & Basso, D. (2022). Morphostructural Characterization of the Heterogeneous Rhodolith Bed at the Marine Protected Area “Capo Carbonara” (Italy) and Hydrodynamics. Diversity, 14(1), 51. https://doi.org/10.3390/d14010051