Are Urban Communities in Successional Stasis? A Case Study on Epiphytic Lichen Communities



Abstract
1. Introduction
- The history of change in pollution levels in London would mean that older and therefore larger trees may have spanned both high and low atmospheric pollution and that this could alter the tree DBH-lichen diversity relationship.
- Lichen communities show a shift in functional trait diversity with sexual reproduction and basidophilous substrate pH preference becoming less abundant as tree size increases.
2. Materials and Methods
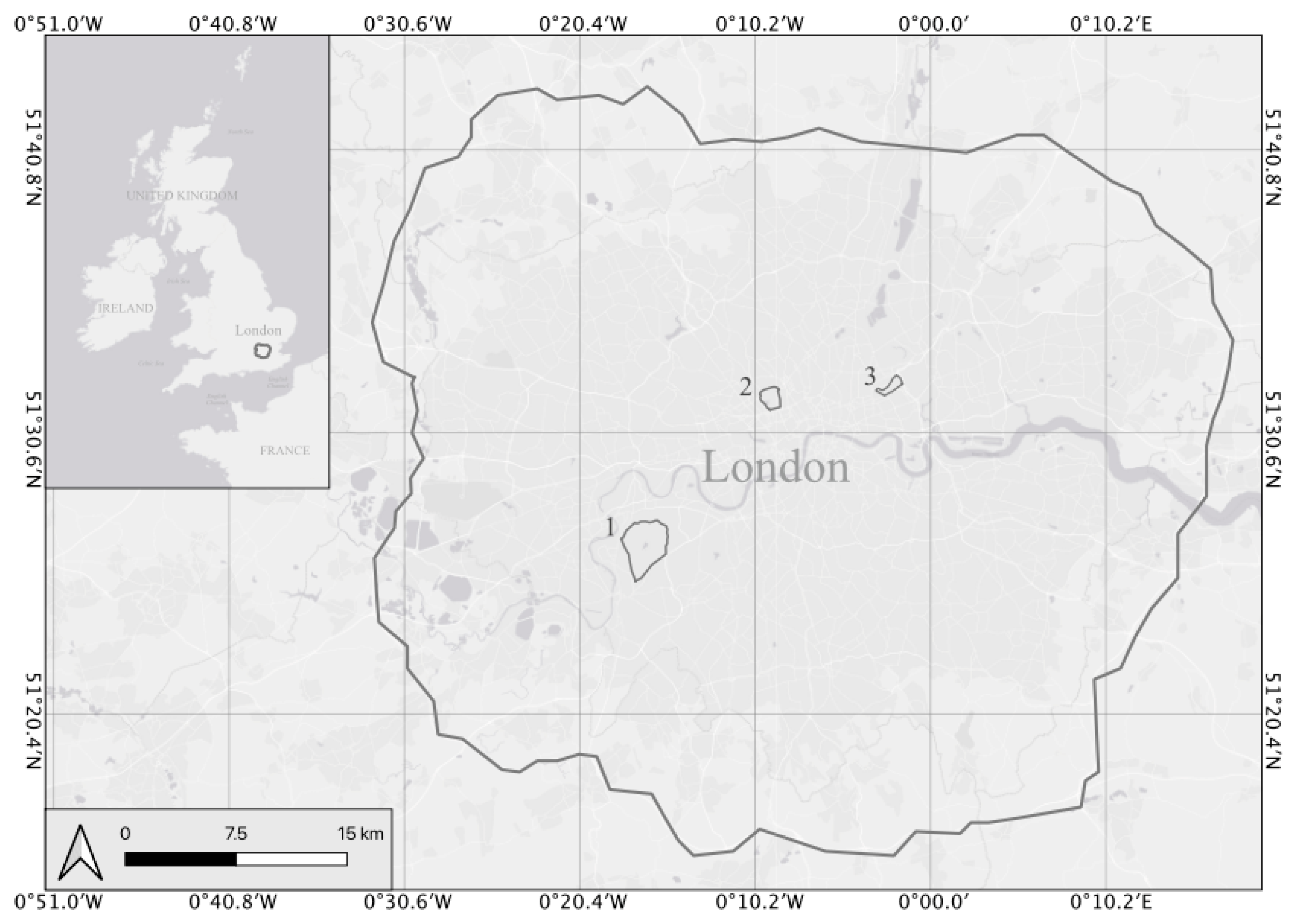
2.1. Survey Sites
2.2. Host Species
2.3. Sampling Method
2.4. Statistical Analysis
2.4.1. Generalized Linear Modeling
2.4.2. Species Co-Occurrence and Community Detection
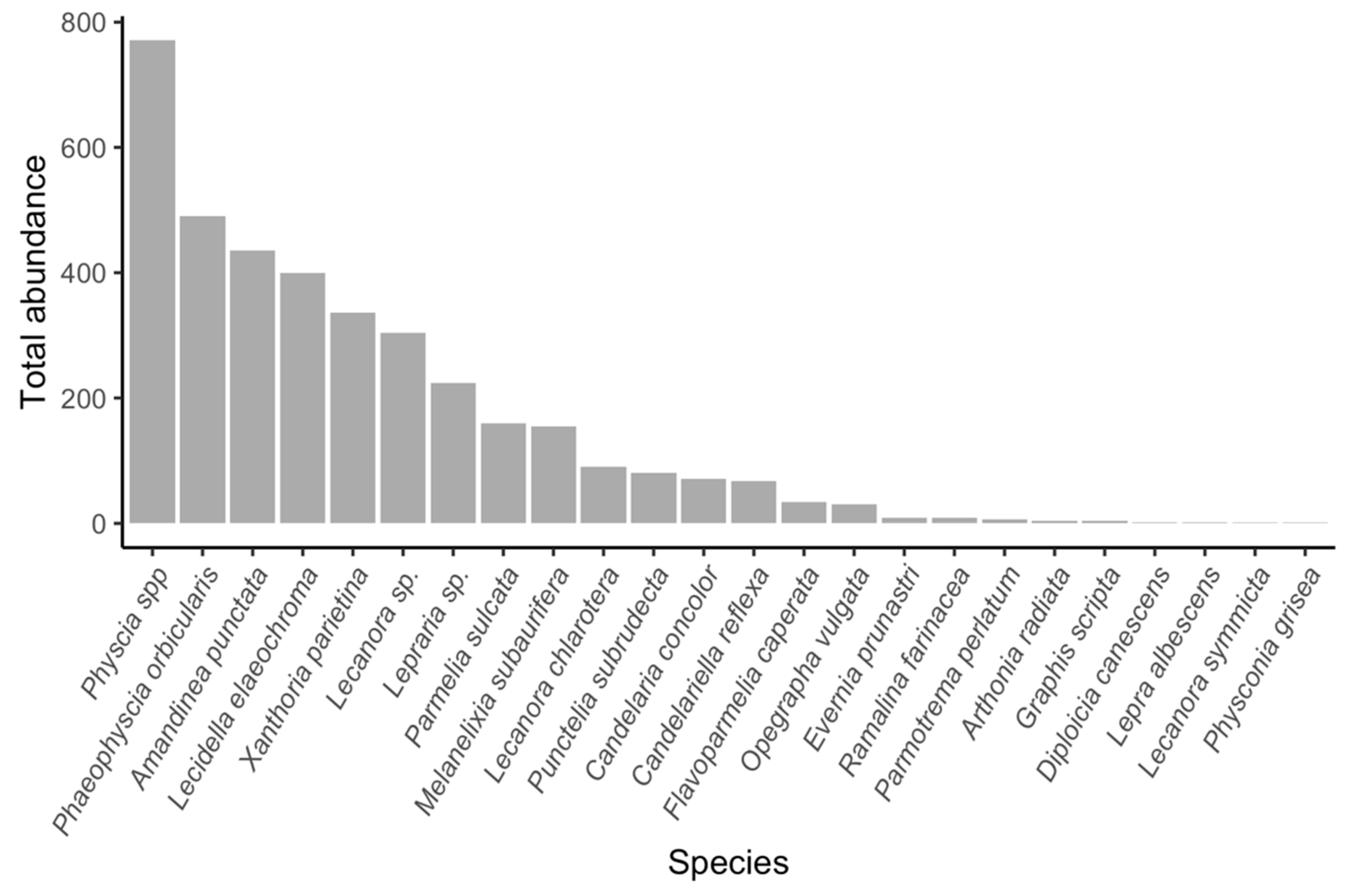
3. Results
3.1. Generalized Linear Modeling
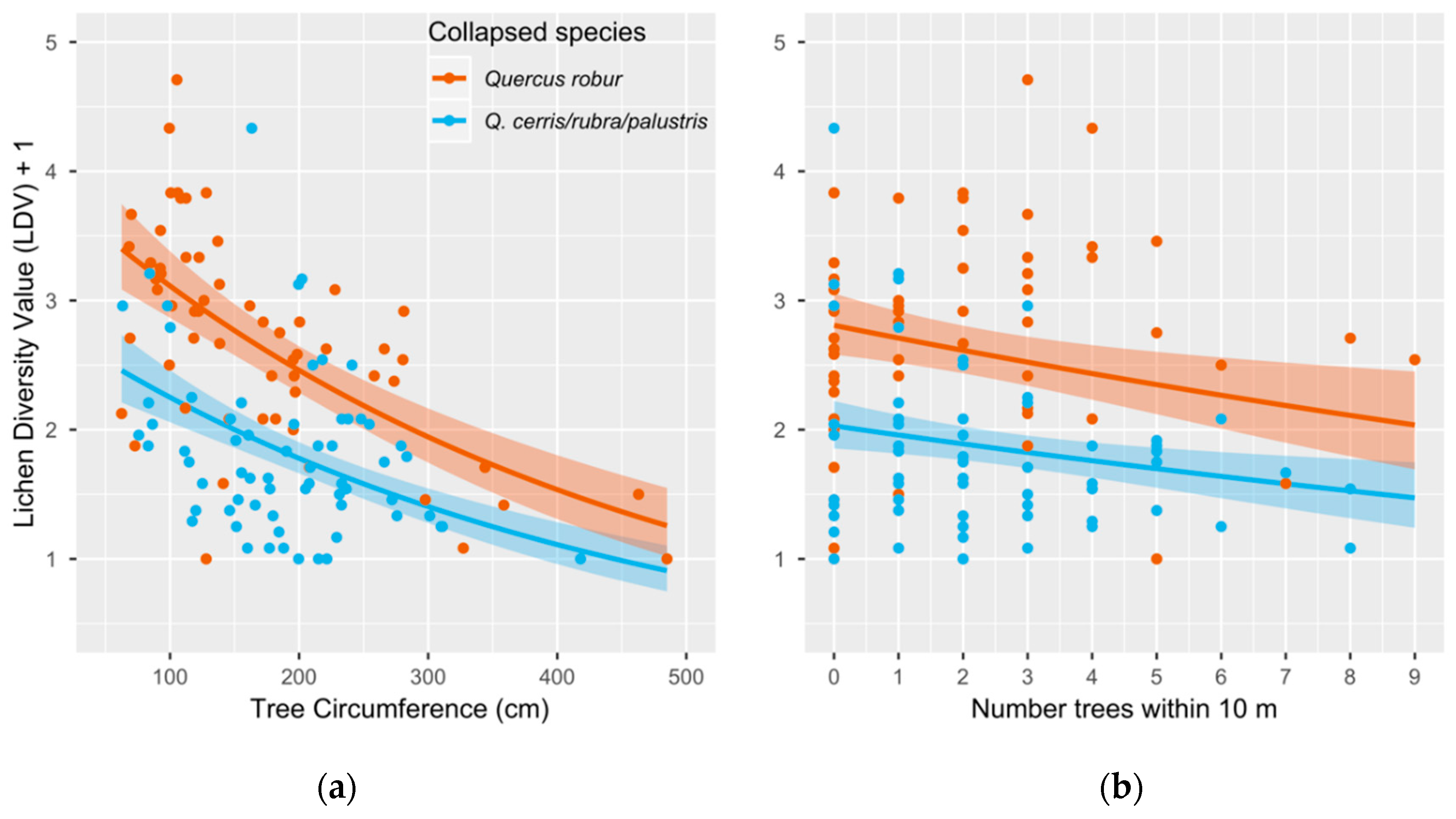
3.1.1. Lichen Diversity Value (LDV)
3.1.2. Functional Trait Analysis
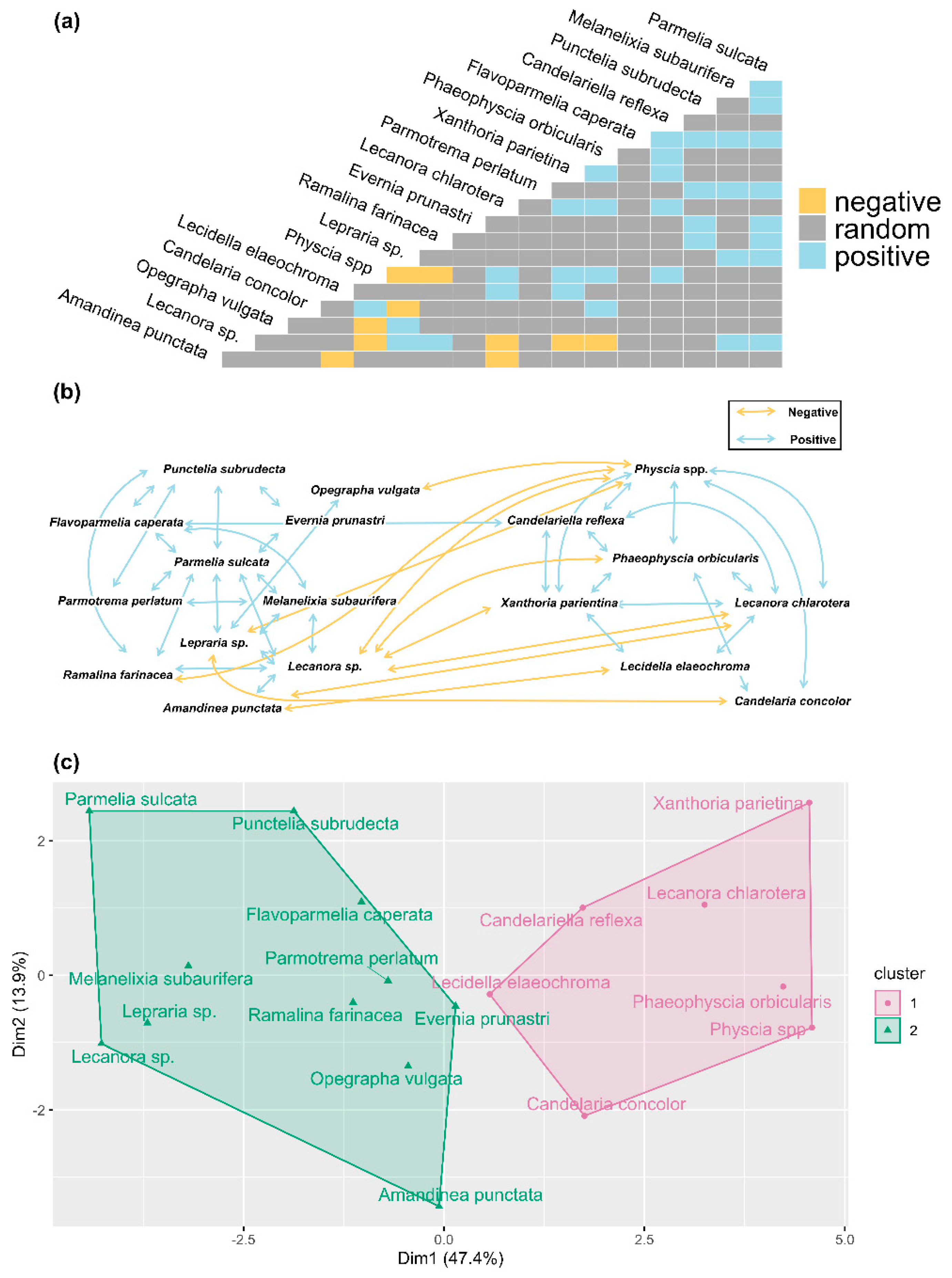
3.2. Species Co-Occurrence and Community Detection
4. Discussion
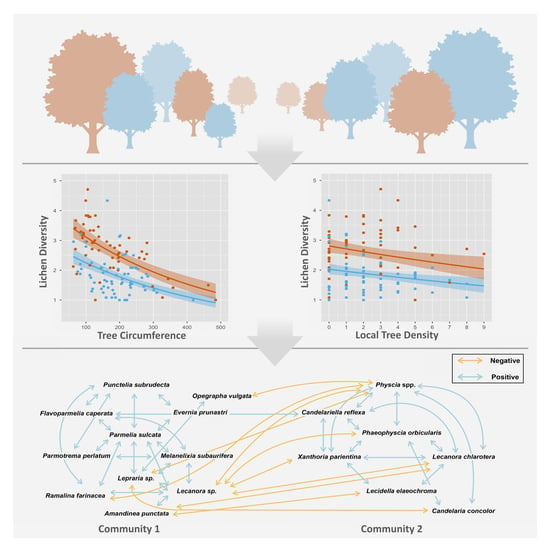
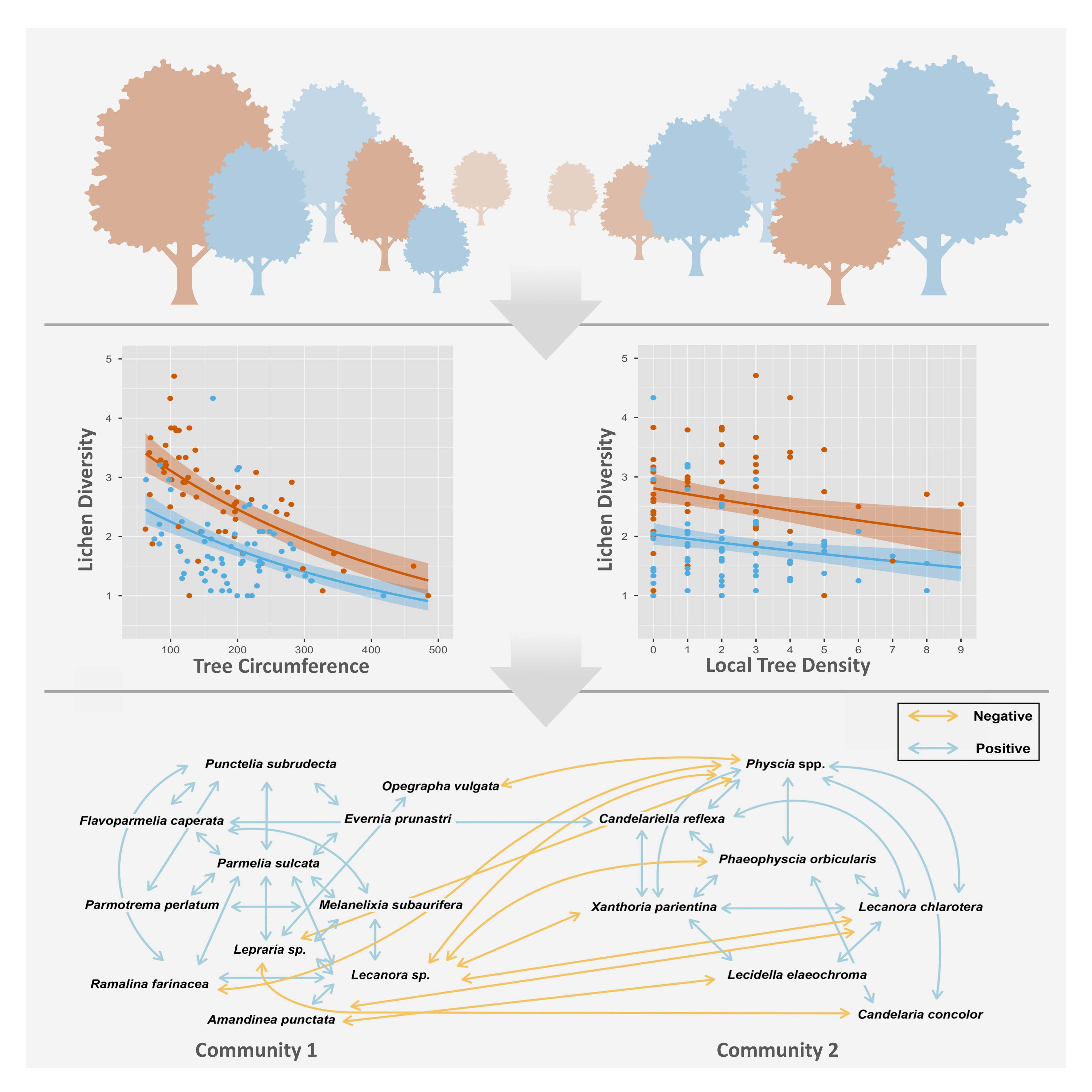
4.1. Lichen Diversity Is Significantly Affected by Host Tree Circumference, Tree Species and Local Crowding
4.2. Species Co-Occurrence Reveals Two Distinct Early Successional Lichen Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Kaye, J.P.; Groffman, P.M.; Grimm, N.B.; Baker, L.A.; Pouyat, R.V. A distinct urban biogeochemistry? Trends Ecol. Evol. 2006, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, J. Is there a need for a theory of urban ecology? Urban Ecosyst. 1999, 3, 57–65. [Google Scholar] [CrossRef]
- Drinnan, I.N. The search for fragmentation thresholds in a Southern Sydney Suburb. Biol. Conserv. 2005, 124, 339–349. [Google Scholar] [CrossRef]
- Gledhill, D.G.; James, P.; Davies, D.H. Pond density as a determinant of aquatic species richness in an urban landscape. Landsc. Ecol. 2008, 23, 1219–1230. [Google Scholar] [CrossRef]
- Turrini, T.; Knop, E. A landscape ecology approach identifies important drivers of urban biodiversity. Glob. Chang. Biol. 2015, 21, 1652–1667. [Google Scholar] [CrossRef]
- Ferenc, M.; Sedláček, O.; Fuchs, R.; Dinetti, M.; Fraissinet, M.; Storch, D. Are cities different? Patterns of species richness and beta diversity of urban bird communities and regional species assemblages in Europe. Glob. Ecol. Biogeogr. 2014, 23, 479–489. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K.L.; Mattox, J.D.; Yaege, C. Massively parallel 454-sequencing of fungal communities in Quercus spp. ectomycorrhizas indicates seasonal dynamics in urban and rural sites. Mol. Ecol. 2010, 19, 41–53. [Google Scholar] [CrossRef]
- Niemelä, J.; Kotze, D.J.; Venn, S.; Penev, L.; Stoyanov, I.; Spence, J.; Hartley, D.; Montes de Oca, E. Carabid beetle assemblages (Coleoptera, Carabidae) across urban-rural gradients: An international comparison. Landsc. Ecol. 2002, 17, 387–401. [Google Scholar] [CrossRef]
- Pavao-Zuckerman, M.A.; Coleman, D.C. Urbanization alters the functional composition, but not taxonomic diversity, of the soil nematode community. Appl. Soil Ecol. 2007, 35, 329–339. [Google Scholar] [CrossRef]
- Bates, S.T.; Cropsey, G.W.G.; Caporaso, J.G.; Knight, R.; Fierer, N. Bacterial communities associated with the lichen symbiosis. Appl. Environ. Microbiol. 2011, 77, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Spribille, T.; Tuovinen, V.; Resl, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome-Heller, M.; Thor, G.; et al. Basidiomycete yeasts in the cortex of ascomycete macrolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Matos, P.; Mexia, T.; Silva, P.; Lopes, N.; Freitas, C.; Correia, O.; Santos-Reis, M.; Branquinho, C.; Pinho, P. Green spaces are not all the same for the provision of air purification and climate regulation services: The case of urban parks. Environ. Res. 2018, 160, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Henriksson, E.; Simu, B. Nitrogen fixation by lichens. Oikos 2012, 22, 119–121. [Google Scholar] [CrossRef]
- Gunnarsson, B.; Hake, M.; Hultengren, S. A functional relationship between species richness of spiders and lichens in spruce. Biodivers. Conserv. 2004, 13, 685–693. [Google Scholar] [CrossRef]
- Pettersson, R.B.; Ball, J.P.; Renhorn, K.E.; Esseen, P.A.; Sjöberg, K. Invertebrate communities in boreal forest canopies as influenced by forestry and lichens with implications for passerine birds. Biol. Conserv. 1995, 74, 57–63. [Google Scholar] [CrossRef]
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota—Approaching one thousand genera. Bryologist 2017, 119, 361–416. [Google Scholar] [CrossRef]
- Nash, T. Lichen Biology; Cambridge University Press: Cambridge, UK, 2008; ISBN 9780874216561. [Google Scholar]
- Purvis, O.W.; Chimonides, J.; Din, V.; Erotokritou, L.; Jeffries, T.; Jones, G.C.; Louwhoff, S.; Read, H.; Spiro, B. Which factors are responsible for the changing lichen floras of London? Sci. Total Environ. 2003, 310, 179–189. [Google Scholar] [CrossRef]
- Lisowska, M. Lichen recolonisation in an urban-industrial area of southern Poland as a result of air quality improvement. Environ. Monit. Assess. 2011, 179, 177–190. [Google Scholar] [CrossRef]
- McMullin, R.T.; Bennett, L.L.; Bjorgan, O.J.; Bourque, D.A.; Burke, C.J.; Clarke, M.A.; Gutgesell, M.K.; Krawiec, P.L.; Malyon, R.; Mantione, A.; et al. Relationships between air pollution, population density, and lichen biodiversity in the Niagara Escarpment World Biosphere Reserve. Lichenologist 2016, 48, 593–605. [Google Scholar] [CrossRef]
- Nascimbene, J.; Tretiach, M.; Corana, F.; Lo Schiavo, F.; Kodnik, D.; Dainese, M.; Mannucci, B. Patterns of traffic polycyclic aromatic hydrocarbon pollution in mountain areas can be revealed by lichen biomonitoring: A case study in the Dolomites (Eastern Italian Alps). Sci. Total Environ. 2014, 475, 90–96. [Google Scholar] [CrossRef]
- Cislaghi, C.; Nimis, P.L. Lichens, air pollution and lung cancer. Nature 1997, 387, 463–464. [Google Scholar] [CrossRef] [PubMed]
- Llop, E.; Pinho, P.; Matos, P.; Pereira, M.J.; Branquinho, C. The use of lichen functional groups as indicators of air quality in a Mediterranean urban environment. Ecol. Indic. 2012, 13, 215–221. [Google Scholar] [CrossRef]
- Koch, N.M.; Matos, P.; Branquinho, C.; Pinho, P.; Lucheta, F.; Martins, S.M.d.A.; Vargas, V.M.F. Selecting lichen functional traits as ecological indicators of the effects of urban environment. Sci. Total Environ. 2019, 654, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Gadsdon, S.R.; Dagley, J.R.; Wolseley, P.A.; Power, S.A. Relationships between lichen community composition and concentrations of NO2 and NH3. Environ. Pollut. 2010, 158, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- Giordani, P.; Malaspina, P. Do tree-related factors mediate the response of lichen functional groups to eutrophication? Plant Biosyst. 2016, 151, 1062–1072. [Google Scholar] [CrossRef]
- Laundon, J.R. A study of the lichen flora of London. Lichenologist 1967, 3, 277–327. [Google Scholar] [CrossRef]
- Rose, C.I.; Hawksworth, D.L. Lichen recolonization in London’s cleaner air. Nature 1981, 289, 289–292. [Google Scholar] [CrossRef]
- Larsen, R.S.; Bell, J.N.B.; James, P.W.; Chimonides, P.J.; Rumsey, F.J.; Tremper, A.; Purvis, O.W. Lichen and bryophyte distribution on oak in London in relation to air pollution and bark acidity. Environ. Pollut. 2007, 146, 332–340. [Google Scholar] [CrossRef]
- Sernander, R. Stockholms Natur; Almqvist & Wiksell: Uppsala, Sweden, 1926. [Google Scholar]
- Laundon, J.R. London’s lichens. Lond. Nat. 1970, 49, 20–69. [Google Scholar]
- Davies, L.; James, P.W.; Chimonides, J.H.; Purvis, O.W. Part 1: Corticolous lichens in London. In Effects of NOx and NH3 on Lichen Communities and Urban Ecosystems: A Pilot Study; Technical Report; The Natural History Museum and Imperial College of Science, Technology and Medicine: London, UK, 2002; pp. 8–29. [Google Scholar]
- Jovan, S.; Riddell, J.; Padgett, P.E.; Nash, T.H. Eutrophic lichens respond to multiple forms of N: Implications for critical levels and critical loads research. Ecol. Appl. 2012, 22, 1910–1922. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.; Bates, J.W.; Bell, J.N.B.; James, P.W.; Purvis, O.W. Diversity and sensitivity of epiphytes to oxides of nitrogen in London. Environ. Pollut. 2007, 146, 299–310. [Google Scholar] [CrossRef]
- Welden, N.A.; Wolseley, P.A.; Ashmore, M.R. Citizen science identifies the effects of nitrogen deposition, climate and tree species on epiphytic lichens across the UK. Environ. Pollut. 2017, 232, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Buckley, H.L. Isolation affects tree-scale epiphytic lichen community structure on New Zealand mountain beech trees. J. Veg. Sci. 2011, 22, 1062–1071. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Motta, R.; Nimis, P.L. Influence of tree age, tree size and crown structure on lichen communities in mature Alpine spruce forests. Biodivers. Conserv. 2009, 18, 1509–1522. [Google Scholar] [CrossRef]
- Reynolds, C.L.; Er, O.A.H.; Winder, L.; Blanchon, D.J. Distribution and community composition of lichens on mature mangroves (Avicennia marina subsp. australasica (Walp.) J.Everett) in New Zealand. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Thor, G.; Johansson, P.; Jönsson, M.T. Lichen diversity and red-listed lichen species relationships with tree species and diameter in wooded meadows. Biodivers. Conserv. 2010, 19, 2307–2328. [Google Scholar] [CrossRef]
- Lie, M.H.; Arup, U.; Grytnes, J.A.; Ohlson, M. The importance of host tree age, size and growth rate as determinants of epiphytic lichen diversity in boreal spruce forests. Biodivers. Conserv. 2009, 18, 3579–3596. [Google Scholar] [CrossRef]
- Ranius, T.; Johansson, P.; Berg, N.; Niklasson, M. The influence of tree age and microhabitat quality on the occurrence of crustose lichens associated with old oaks. J. Veg. Sci. 2008, 19, 653–662. [Google Scholar] [CrossRef]
- Johansson, P.; Rydin, H.; Thor, G. Tree age relationships with epiphytic lichen diversity and lichen life history traits on ash in southern Sweden. Ecoscience 2007, 14, 81–91. [Google Scholar] [CrossRef]
- Fritz, Ö.; Brunet, J.; Caldiz, M. Interacting effects of tree characteristics on the occurrence of rare epiphytes in a Swedish beech forest area. Bryologist 2009, 112, 488–505. [Google Scholar] [CrossRef]
- Fritz, Ö.; Niklasson, M.; Churski, M. Tree age is a key factor for the conservation of epiphytic lichens and bryophytes in beech forests. Appl. Veg. Sci. 2009, 12, 93–106. [Google Scholar] [CrossRef]
- Mežaka, A.; Brūmelis, G.; Piterāns, A. Tree and stand-scale factors affecting richness and composition of epiphytic bryophytes and lichens in deciduous woodland key habitats. Biodivers. Conserv. 2012, 21, 3221–3241. [Google Scholar] [CrossRef]
- Adams, D.B.; Risser, P.G. Some factors influencing the frequency of bark lichens in north central Oklahoma. Am. J. Bot. 1971, 58, 752. [Google Scholar] [CrossRef]
- Ellis, C.J.; Ellis, S.C. Signatures of autogenic epiphyte succession for an aspen chronosequence. J. Veg. Sci. 2013, 24, 688–701. [Google Scholar] [CrossRef]
- Sales, K.; Kerr, L.; Gardner, J. Factors influencing epiphytic moss and lichen distribution within killarney national park. Biosci. Horiz. 2016, 9. [Google Scholar] [CrossRef]
- Jüriado, I.; Liira, J.; Paal, J.; Suija, A. Tree and stand level variables influencing diversity of lichens on temperate broad-leaved trees in boreo-nemoral floodplain forests. Biodivers. Conserv. 2009, 18, 105–125. [Google Scholar] [CrossRef]
- Culberson, W.L. The corticolous communities of lichens and bryophytes in the upland forests of northern Wisconsin. Source Ecol. Monogr. 1955, 25, 215–231. [Google Scholar] [CrossRef]
- Gauslaa, Y. The ecology of Lobarion pulmonariae and Parmelion caperatae in Quercus dominated forests in south-west Norway. Lichenologist 1985, 17, 117–140. [Google Scholar] [CrossRef]
- Bates, J.W. Influence of chemical and physical factors on Quercus and Fraxinus epiphytes at Loch Sunart, western Scotland: A multivariate analysis. J. Ecol. 1992, 80, 163–179. [Google Scholar] [CrossRef]
- Löbel, S.; Snäll, T.; Rydin, H. Species richness patterns and metapopulation processes—Evidence from epiphyte communities in boreo-nemoral forests. Ecography 2006, 29, 169–182. [Google Scholar] [CrossRef]
- Degtjarenko, P.; Matos, P.; Marmor, L.; Branquinho, C.; Randlane, T. Functional traits of epiphytic lichens respond to alkaline dust pollution. Fungal Ecol. 2018, 36, 81–88. [Google Scholar] [CrossRef]
- McDonald, L.; Van Woudenberg, M.; Dorin, B.; Adcock, A.M.; McMullin, R.T.; Cottenie, K. The effects of bark quality on corticolous lichen community composition in urban parks of southern Ontario. Botany 2017, 95, 1141–1149. [Google Scholar] [CrossRef]
- Van Herk, C.M. Bark pH and susceptibility to toxic air pollutants as independent causes of changes in epiphytic lichen composition in space and time. Lichenologist 2001, 33, 419–441. [Google Scholar] [CrossRef]
- Van Herk, C.M. Mapping of ammonia pollution with epiphytic lichens in the Netherlands. Lichenologist 1999, 31, 9–20. [Google Scholar] [CrossRef]
- Nimis, P.L.; Martellos, S. ITALIC—The Information System on Italian Lichens Version 5.0. University of Trieste, Dept. of Biology, 2017. Available online: http://dryades.units.it/italic (accessed on 19 December 2018).
- LeBlanc, F.; Barkman, J.J. Phytosociology and ecology of cryptogamic epiphytes. Bryologist 1971, 74, 407. [Google Scholar] [CrossRef]
- Cleavitt, N.L.; Dibble, A.C.; Werier, D.A. Influence of tree composition upon epiphytic macrolichens and bryophytes in old forests of Acadia National Park, Maine. Bryologist 2009, 112, 467–487. [Google Scholar] [CrossRef]
- Király, I.; Ódor, P. The effect of stand structure and tree species composition on epiphytic bryophytes in mixed deciduous-coniferous forests of Western Hungary. Biol. Conserv. 2010, 143, 2063–2069. [Google Scholar] [CrossRef]
- Spier, L.; Van Dobben, H.; Van Dort, K. Is bark pH more important than tree species in determining the composition of nitrophytic or acidophytic lichen floras? Environ. Pollut. 2010, 158, 3607–3611. [Google Scholar] [CrossRef]
- Ellis, C.J.; Coppins, B.J. Contrasting functional traits maintain lichen epiphyte diversity in response to climate and autogenic succession. J. Biogeogr. 2006, 33, 1643–1656. [Google Scholar] [CrossRef]
- Ellis, C.J.; Coppins, B.J. Reproductive strategy and the compositional dynamics of crustose lichen communities on aspen (Populus tremula L.) in Scotland. Lichenologist 2007, 39, 377–391. [Google Scholar] [CrossRef]
- Lewis, J.E.J.; Ellis, C.J. Taxon- compared with trait-based analysis of epiphytes, and the role of tree species and tree age in community composition. Plant Ecol. Divers. 2010, 3, 203–210. [Google Scholar] [CrossRef]
- Dettki, H.; Klintberg, P.; Esseen, P.A. Are epiphytic lichens in young forests limited by local dispersal? Ecoscience 2000, 7, 317–325. [Google Scholar] [CrossRef]
- Öckinger, E.; Niklasson, M.; Nilsson, S.G. Is local distribution of the epiphytic lichen Lobaria pulmonaria limited by dispersal capacity or habitat quality? Biodivers. Conserv. 2005, 14, 759–773. [Google Scholar] [CrossRef]
- Kalwij, J.M.; Wagner, H.H.; Scheidegger, C. Effects of stand-level disturbances on the spatial distribution of a lichen indicator. Ecol. Appl. 2005, 15, 2015–2024. [Google Scholar] [CrossRef]
- Sillett, S.C.; McCune, B.; Peck, J.E.; Rambo, T.R.; Ruchty, A. Dispersal limitations of epiphytic lichens result in species dependent on old-growth forests. Ecol. Appl. 2000, 10, 789–799. [Google Scholar] [CrossRef]
- Greater London Authority London Atmospheric Emissions (LAEI) 2016, London Datastore. Available online: https://data.london.gov.uk/dataset/london-atmospheric-emissions-inventory--laei--2016 (accessed on 8 April 2018).
- Zandbergen, P.A.; Barbeau, S.J. Positional accuracy of assisted GPS data from high-sensitivity GPS-enabled mobile phones. J. Navig. 2011, 64, 381–399. [Google Scholar] [CrossRef]
- Asta, J.; Erhardt, W.; Ferretti, M.; Fornasier, F. Mapping lichen diversity as an indicator of environmental quality. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Springer: Dordrecht, The Netherlands, 2002; Volume 7, pp. 273–279. ISBN 978-94-010-0423-7. [Google Scholar]
- Seed, L.; Wolseley, P.; Gosling, L.; Davies, L.; Power, S.A. Modelling relationships between lichen bioindicators, air quality and climate on a national scale: Results from the UK OPAL air survey. Environ. Pollut. 2013, 182, 437–447. [Google Scholar] [CrossRef]
- Dobson, F.S. Lichens: An Illustrated Guide to the British and Irish Species, 6th ed.; Richmond Publishing: Slough, UK, 2011; ISBN 978-0-85546-315-1. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 16 October 2019).
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field—Methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Veech, J.A. A probabilistic model for analysing species co-occurrence. Glob. Ecol. Biogeogr. 2013, 22, 252–260. [Google Scholar] [CrossRef]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic species co-occurrence analysis in R. J. Stat. Softw. 2016, 69, 1–7. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; Studer, M.; Roudier, P.; Gonzalez, J.; Kozlowski, K. Cluster: Cluster analysis basics and extensions. 2018. R Package Version 2.0.7-1. Available online: https://CRAN.R-project.org/package=cluster (accessed on 30 January 2020).
- Hedenaås, H.; Ericson, L. Epiphytic macrolichens as conservation indicators: Successional sequence in Populus tremula stands. Biol. Conserv. 2000, 93, 43–53. [Google Scholar] [CrossRef]
- Wolseley, P.A.; James, P.W.; Theobald, M.R.; Sutton, M.A. Detecting changes in epiphytic lichen communities at sites affected by atmospheric ammonia from agricultural sources. Lichenologist 2006, 38, 161–176. [Google Scholar] [CrossRef]
- Batty, K.; Bates, J.W.; Bell, J.N.B. A transplant experiment on the factors preventing lichen colonization of oak bark in southeast England under declining SO2 pollution. Can. J. Bot. 2003, 81, 439–451. [Google Scholar] [CrossRef]
- London Air Quality Network (LAQN). London Air Quality Network—Summary Report 2018; Techinal Report Environmental Research Group, King’s College London: London, UK, 2018; Available online: https://www.londonair.org.uk/london/reports/2018_LAQN_report.pdf (accessed on 11 August 2020).
- Hawksworth, D. Lichens: Changes in the lichen flora on trees in Epping Forest through periods of increasing and then ameliorating sulphur dioxide air pollution. In Epping Forest Through the Eye of the Naturalist; Hanson, M.W., Ed.; Essex Naturalist, Essex Field Club: London, UK, 1992; pp. 92–101. ISBN 090563716X. [Google Scholar]
- Guest, J. Further recolonisation of Cheshire by epiphytic lichens. Brit. Lichen Soc. Bull. 1989, 64, 29–31. [Google Scholar]
- Hauck, M.; Jung, R.; Runge, M. Does water-holding capacity of bark have an influence on lichen performance in dieback-affected spuce forests? Lichenologist 2000, 32, 407–409. [Google Scholar] [CrossRef]
- Hyvärinen, M.; Halonen, P.; Kauppi, M. Influence of stand age and structure on the epiphytic lichen vegetation in the middle-boreal forests of Finland. Lichenologist 1992, 24, 165–180. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A. Differences in the susceptibility to light stress between epiphytic lichens of ancient and young boreal forest stands. Funct. Ecol. 1996, 10, 344–354. [Google Scholar] [CrossRef]
- Jönsson, M.T.; Thor, G.; Johansson, P. Environmental and historical effects on lichen diversity in managed and unmanaged wooded meadows. Appl. Veg. Sci. 2011, 14, 120–131. [Google Scholar] [CrossRef]
- James, P.W.; Hawksworth, D.L.; Rose, F. Lichen communities in the British Isles: A preliminary conspectus. In Lichen Ecology; Seaward, M.R.D., Ed.; Academic Press: London, UK, 1977; pp. 295–413. [Google Scholar]
- Nascimbene, J.; Marini, L.; Nimis, P.L. Influence of tree species on epiphytic macrolichens in temperate mixed forests of northern Italy. Can. J. For. Res. 2009, 39, 785–791. [Google Scholar] [CrossRef]
- Ronnås, C.; Werth, S.; Ovaskainen, O.; Várkonyi, G.; Scheidegger, C.; Snäll, T. Discovery of long-distance gamete dispersal in a lichen-forming ascomycete. New Phytol. 2017, 216, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.M. Distributional ecology of New Guinea birds. Science 1973, 179, 759–769. [Google Scholar] [CrossRef]
- Flügge, A.J.; Olhede, S.C.; Murrell, D.J. A method to detect subcommunities from multivariate spatial associations. Methods Ecol. Evol. 2014, 5, 1214–1224. [Google Scholar] [CrossRef]
- Ellis, C.J.; Eaton, S.; Theodoropoulos, M.; Elliot, K. Epiphyte Communities and Indicator Species. An Ecological Guide for Scotland’s Woodlands; Royal Botanic Gardens Edinburgh: Edinburgh, UK, 2015; ISBN 978-1-910877-01-2. [Google Scholar]
- Rose, F. Ancient British woodlands and their epiphytes. Br. Wildl. 1993, 5, 83–93. [Google Scholar]
- Paltto, H.; Nordberg, A.; Nordén, B.; Snäll, T. Development of secondary woodland in oak wood pastures reduces the richness of rare epiphytic lichens. PLoS ONE 2011, 6, e24675. [Google Scholar] [CrossRef]
- Outhwaite, C.L.; Gregory, R.D.; Chandler, R.E.; Collen, B.; Isaac, N.J.B. Complex long-term biodiversity change among invertebrates, bryophytes and lichens. Nat. Ecol. Evol. 2020, 4, 384–392. [Google Scholar] [CrossRef]
- Giordani, P.; Malaspina, P.; Benesperi, R.; Incerti, G.; Nascimbene, J. Functional over-redundancy and vulnerability of lichen communities decouple across spatial scales and environmental severity. Sci. Total Environ. 2019, 666, 22–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | 95% CIs | Std. Error | t Value | p-Value | |
|---|---|---|---|---|---|
| Intercept | 1.118 | 0.959–1.277 | 0.081 | 13.818 | <2 × 10−16 |
| Circumference | −0.002 | −0.003–−0.002 | 0.000 | −7.401 | 2.04 × 10−11 |
| Quercus Robur | 0.324 | 0.225–0.423 | 0.051 | 6.42 | 2.85 × 10−9 |
| No. Trees 10 m | −0.036 | −0.059–−0.012 | 0.012 | −2.903 | 0.004 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llewellyn, T.; Gaya, E.; Murrell, D.J. Are Urban Communities in Successional Stasis? A Case Study on Epiphytic Lichen Communities. Diversity 2020, 12, 330. https://doi.org/10.3390/d12090330
Llewellyn T, Gaya E, Murrell DJ. Are Urban Communities in Successional Stasis? A Case Study on Epiphytic Lichen Communities. Diversity. 2020; 12(9):330. https://doi.org/10.3390/d12090330
Chicago/Turabian StyleLlewellyn, Theo, Ester Gaya, and David J. Murrell. 2020. "Are Urban Communities in Successional Stasis? A Case Study on Epiphytic Lichen Communities" Diversity 12, no. 9: 330. https://doi.org/10.3390/d12090330
APA StyleLlewellyn, T., Gaya, E., & Murrell, D. J. (2020). Are Urban Communities in Successional Stasis? A Case Study on Epiphytic Lichen Communities. Diversity, 12(9), 330. https://doi.org/10.3390/d12090330