Rusty Blackbird Habitat Selection and Survivorship during Nesting and Post-Fledging

Abstract
1. Introduction
2. Materials and Methods
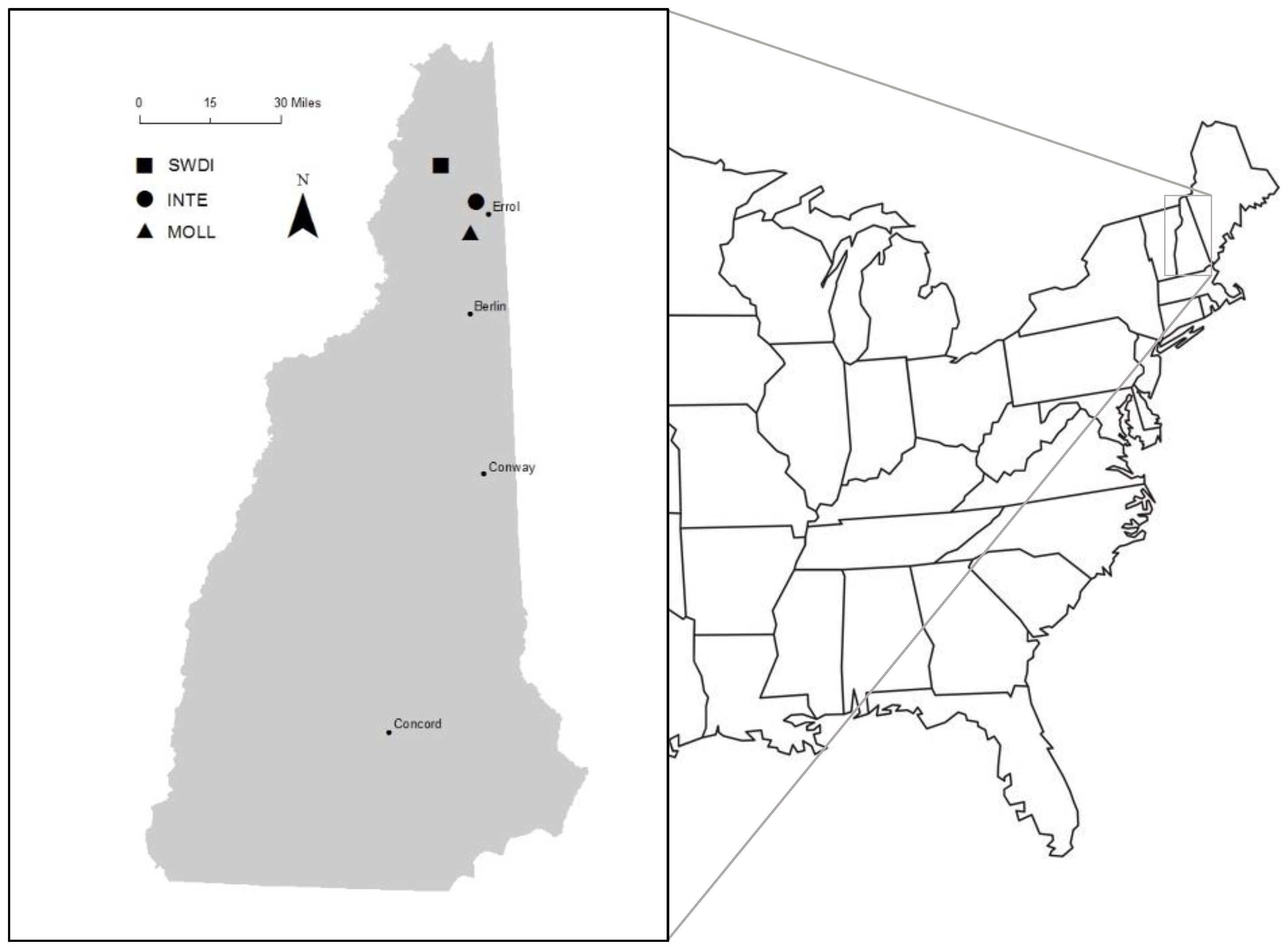
2.1. Study Area
2.2. Field Procedures
2.3. Data Analyses
2.3.1. Habitat Selection
2.3.2. Nest Success and Adult and Fledgling Survivorship
3. Results
3.1. General
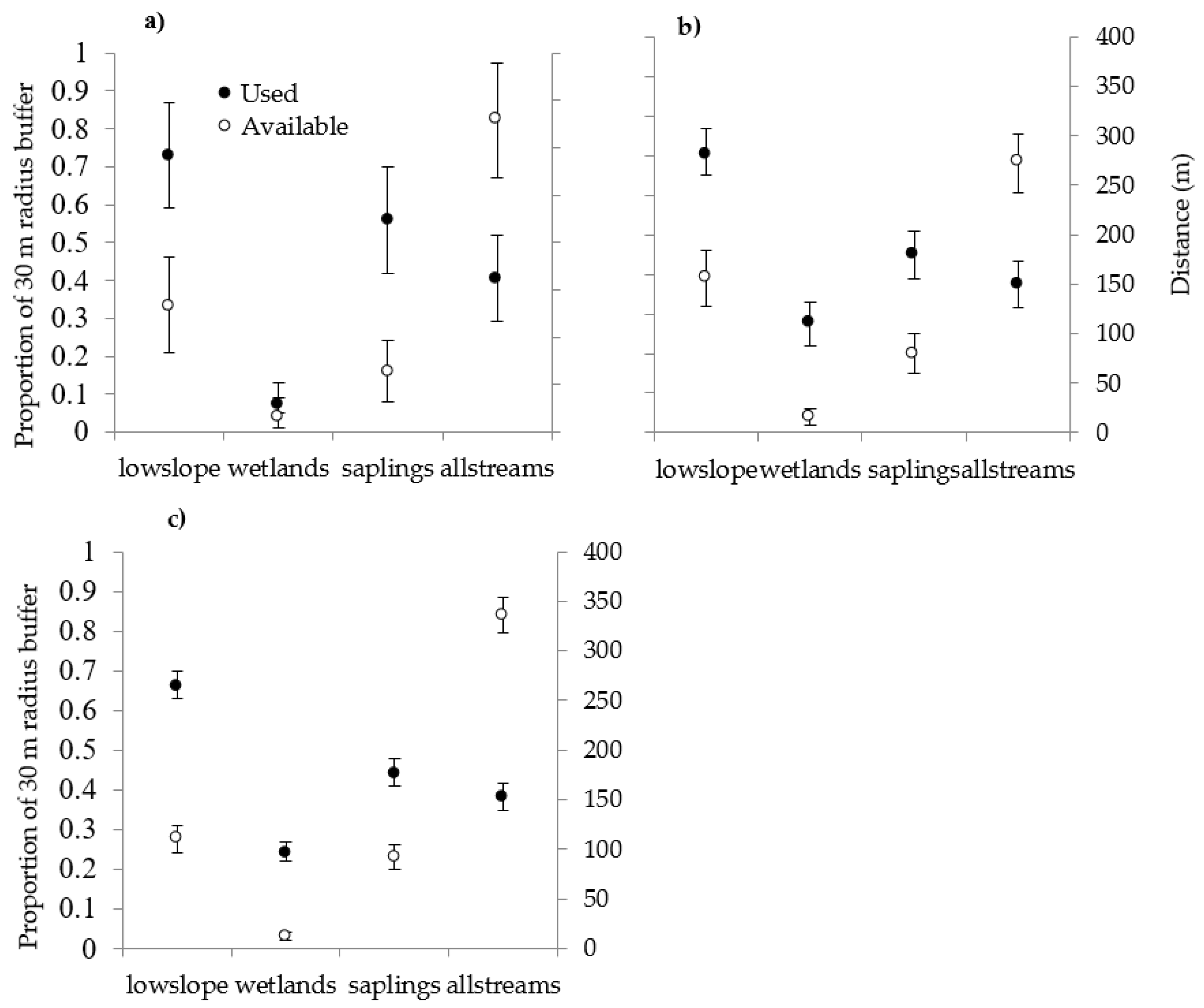
3.2. Habitat Selection
3.3. Nest Success
3.4. Blackbird Survival
4. Discussion
4.1. Survival and Habitat Selection
4.2. Recommendations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
| Year | Drainage Tagged | Total Birds | Total Points | Birds in Analysis | Adults in Analysis | Fledges in Analysis | Points in Analysis |
| 2010 | SWDI | 8 | 167 | 4 | 1M,1F | 2 | 64 |
| 2010 | MOLL | 8 | 130 | 6 | 3M,2F | 1 | 73 |
| 2010 | 2010 Total | 16 | 297 | 10 | 7 | 3 | 137 |
| 2011 | SWDI | 14 | 687 | 10 | 5M,2F | 3 | 230 |
| 2011 | MOLL | 6 | 120 | 4 | 2M | 2 | 60 |
| 2011 | INTE | 4 | 186 | 4 | 2M,1F | 1 | 84 |
| 2011 | 2011 Total | 24 | 993 | 18 | 12 | 6 | 374 |
| 2012 | SWDI | 12 | 486 | 10 | 5M,4F | 1 | 211 |
| 2012 | MOLL | 0 | 0 | 0 | 0 | 0 | 0 |
| 2012 | INTE | 4 | 89 | 2 | 2F | 0 | 49 |
| 2012 | 2012 Total | 16 | 575 | 12 | 11 | 1 | 300 |
| Total | 56 | 1865 | 40 | 331 | 10 | 771 |
References
- Vitz, A.C.; Rodewald, A.D. Can regenerating clearcuts benefit mature-forest songbirds? An examination of post-breeding ecology. Biol. Conserv. 2006, 127, 477–486. [Google Scholar] [CrossRef]
- King, D.I.; Degraaf, R.M.; Smith, M.L.; Buonaccorsi, J.P. Habitat selection and habitat-specific survival of fledgling ovenbirds (Seiurus aurocapilla). J. Zool. 2006, 269, 414–421. [Google Scholar] [CrossRef]
- Rush, S.A.; Stutchbury, B.J.M. Survival of fledgling hooded warblers (Wilsonia citriana) in small and large forest fragments. Auk 2008, 125, 183–191. [Google Scholar] [CrossRef]
- Temple, S.; Cary, J. Modeling the dynamics of forest-interior bird populations in a fragmented landscape. Conserv. Biol. 1988, 2, 305–312. [Google Scholar] [CrossRef]
- Faaborg, J.; Holmes, R.T.; Anders, A.D.; Bildstein, K.L.; Dugger, K.M.; Gauthreaux, S.A.; Heglund, P.J.; Hobson, K.A.; Jahn, A.E.; Johnson, D.H.; et al. Recent advances in understanding migration systems of New World land birds. Ecol. Monogr. 2010, 80, 3–48. [Google Scholar] [CrossRef]
- Mitchell, G.W.; Taylor, P.D.; Warkentin, I.G. Assessing the function of broad-scale movements made by juvenile songbirds prior to migration. Condor 2010, 112, 644–654. [Google Scholar] [CrossRef]
- Moore, L.C.; Stutchbury, B.J.M.; Burke, D.M.; Elliott, K. Effects of forest management on post-fledging survival of rose-breasted grosbeaks (Pheucticus ludovicianus). Auk 2010, 127, 185–194. [Google Scholar] [CrossRef]
- Vitz, A.C.; Rodewald, A.D. Influence of condition and habitat use on survival of post-fledging songbirds. Condor 2011, 113, 400–411. [Google Scholar] [CrossRef]
- Evans, D.; Hobson, K.A.; Kusack, J.W.; Cadman, M.D.; Falconer, C.M.; Mitchell, G.W. Individual condition, but not fledging phenology, carries over to affect post-fledging survival in a Neotropical migratory songbird. IBIS 2019, 162, 331–344. [Google Scholar] [CrossRef]
- Anders, A.D.; Dearborn, D.C.; Faaborg, J.; Thompson, F.R., III. Juvenile survival in a population of neotropical migrant birds. Conserv. Biol. 1997, 11, 698–707. [Google Scholar] [CrossRef]
- Burke, A.D.; Thompson, F.R., III; Faaborg, J. Variation in early-successional habitat use among independent juvenile forest breeding birds. Wilson J. Ornithol. 2017, 129, 235–246. [Google Scholar] [CrossRef]
- Marshall, M.R.; DeCecco, J.A.; Williams, A.B.; Gale, G.A.; Cooper, R.J. Use of regenerating clearcuts by late-successional bird species and their young during the post-fledging period. For. Ecol. Manag. 2003, 183, 127–135. [Google Scholar] [CrossRef]
- White, J.D.; Gardali, T.; Thompson, F.R., III; Faaborg, J. Resource selection by juvenile Swainson’s thrushes during the post-fledging period. Condor 2005, 107, 388–401. [Google Scholar] [CrossRef]
- Jenkins, J.M.A.; Thompson, F.R., III.; Faaborg, J. Species-specific variation in nesting and post-fledging resource selection for two forest breeding migrant songbirds. PLoS ONE 2017, 12, e0179524. [Google Scholar] [CrossRef] [PubMed]
- Delancey, C.D.; Islam, K. Post-fledging habitat use in a declining songbird. PeerJ 2019, 7, e7358. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.C.; Knadle, G.E.; Brubaker, D.L.; Brubaker, K.S. Nest success is not an adequate comparative estimate of avian reproduction. J. Field Ornithol. 2001, 72, 527–536. [Google Scholar] [CrossRef]
- Avery, M.L. Rusty Blackbird (Euphagus carolinus). In The Birds of North America, version 2.0.; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2013. [Google Scholar] [CrossRef]
- Glennon, M.J. Dynamics of boreal birds at the edge of their range in the Adirondack Park, NY. Northeast. Nat. 2014, 21. [Google Scholar] [CrossRef]
- McNulty, S.; Glennon, M.; McCormack, M. Rusty blackbirds in New York State: Ecology, current status, and future. Adirond. J. Environ. Stud. 2015, 20, 87–99. [Google Scholar]
- Greenberg, R.; Droege, S. On the decline of the rusty blackbird and the use of ornithological literature to document long-term population trends. Conserv. Biol. 1999, 13, 553–559. [Google Scholar] [CrossRef]
- Niven, D.K.; Sauer, J.R.; Butcher, G.S.; Link, W.A. Christmas Bird Count provides insights into population change in land birds that breed in the boreal forest. Am. Birds 2004, 58, 10–20. [Google Scholar]
- Sauer, J.R.; Hines, J.E.; Fallon, J. The North American Breeding Bird Survey, Results and Analysis 1966–2005; Version 6.2.; USGS Patuxent Wildlife Research Center: Laurel, ML, USA, 2006.
- Greenberg, R.; Matsuoka, S.M. Rusty blackbird: Mysteries of a species in decline. Condor 2010, 112, 770–777. [Google Scholar] [CrossRef]
- Greenberg, R.; Demarest, D.W.; Matsuoka, S.M.; Mettke-Hofmann, C.; Evers, D.; Hamel, P.B.; Luscier, J.; Powell, L.L.; Shaw, D.; Avery, M.L.; et al. Understanding declines in rusty blackbirds. In Boreal Birds of North America: A Hemispheric View of Their Conservation Links and Significance; Studies in Avian Biology: 41; Wells, J.V., Ed.; University of California Press: Berkeley, CA, USA, 2011; pp. 107–126. [Google Scholar]
- U.S. Fish and Wildlife Service. Birds of Conservation Concern; U.S. Fish and Wildlife Service, Division of Migratory Bird Management: Arlington, VA, USA, 2008.
- International Union for Conservation of Nature and Natural Resources [IUCN]. Euphagus Carolinus, IUCN Red List of Threatened Species, version 2010. 2; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2010. [Google Scholar]
- Matsuoka, S.; Shaw, D.; Sinclair, P.H.; Johnson, J.A.; Corcoran, R.M.; Dau, N.C.; Meyers, P.M.; Rojek, N.A. Nesting ecology of the rusty blackbird in Alaska and Canada. Condor 2010, 112, 810–824. [Google Scholar] [CrossRef]
- Powell, L.L.; Hodgman, T.P.; Glanz, W.E.; Osenton, J.D.; Fisher, C.M. Nest-site selection and nest survival of the rusty blackbird: Does timber management adjacent to wetlands create ecological traps? Condor 2010, 112, 800–809. [Google Scholar] [CrossRef]
- Buckley Luepold, S.H.; Hodgman, T.P.; McNulty, S.A.; Cohen, J.; Foss, C.R. Habitat selection, nest survival, and nest predators of rusty blackbirds in northern New England, USA. Condor 2015, 117, 609–623. [Google Scholar] [CrossRef]
- Martin, T.E. Avian life history evolution in relation to nest sites, nest predation, and food. Ecol. Monogr. 1995, 65, 101–127. [Google Scholar] [CrossRef]
- Pachomski, A.L.; McNulty, S.; Foss, C.R.; Cohen, J.; Farrell, S. Rusty Blackbird (Euphagus carolinus) foraging habitat and prey availability in New England: Implications for conservation of a declining boreal bird species. Diversity 2020, 12. [Google Scholar]
- Schmidt, K.A.; Rush, S.A.; Ostfeld, R.S. Wood thrush nest success and post-fledging survival across a temporal pulse of small mammal abundance in an oak forest. J. Anim. Ecol. 2008, 77, 830–837. [Google Scholar] [CrossRef]
- Thomas, D.W.; Blondel, J.; Perret, P.; Lambrechts, M.M.; Speakman, J.R. Energetic and fitness costs of mismatching resource supply and demand in seasonally breeding birds. Science 2001, 291, 2598–2600. [Google Scholar] [CrossRef]
- Bailey, R.G. Description of the Ecoregions of the United States; 2nd edition revised and expanded. Misc. Publ. No. 1391 (rev.); USDA Forest Service: Washington, DC, USA, 1995.
- Appalachian Mountain Club. Ecological Atlas of the Upper Androscoggin River Watershed; Appalachian Mountain Club: Boston, MA, USA, 2003. [Google Scholar]
- U.S. Climate Data. Available online: https://www.usclimatedata.com/climate/berlin/new-hampshire/united-states/usnh0020 (accessed on 14 May 2020).
- Rappole, J.H.; Tipton, A.R. New harness design for attachment of radio transmitters to small passerines. J. Field Ornithol. 1991, 62, 335–337. [Google Scholar]
- Lele, S.R.; Merrill, E.H.; Keim, J.; Boyce, M.S. Selection, use, choice and occupancy: Clarifying concepts in resource selection studies. J. Anim. Ecol. 2013, 82, 1183–1191. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Fieberg, J.; Matthiopoulos, J.; Hebblewhite, M.; Boyce, M.S.; Frair, J.L. Correlation and studies of habitat selection: Problem, red herring or opportunity? Philos. Trans. R. Soc. B 2010, 365, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Neter, J.; Wasserman, W.; Kutner, M.H. Applied Linear Statistical Models R.D.; Irwin: Homewood, IL, USA, 1990. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Inference: An Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Royall, R.M. Statistical Evidence: A Likelihood Paradigm; Chapman and Hall: London, UK, 1997. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 1st ed.; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- Ott, R.L.; Longnecker, M. An Introduction to Statistical Methods and Data Analysis, 1st ed.; Wadsworth Group: Pacific Groove, CA, USA, 2001. [Google Scholar]
- Laake, J.L. RMark: An R Interface for Analysis of Capture-Recapture Data with MARK; U.S. Department of Commerce: Seattle, WA, USA, 2017.
- Cooch, E.; White, G. Program. MARK: A Gentle Introduction, 20th ed. 2018. Available online: http://www.phidot.org/software/mark/docs/book/ (accessed on 1 June 2020).
- White, G.C.; Burnham, K.P. Program. MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46, S120–S139. [Google Scholar]
- Batzer, D.P.; Rader, R.B.; Wissinger, S.A. Invertebrates in Freshwater Wetlands of North America: Ecology and Management; John Wiley and Sons: New York, NY, USA, 1999. [Google Scholar]
- Arndt, A.; Domdei, J. Influence of beaver ponds on the macroinvertebrate benthic community in lowland brooks. Pol. J. Ecol. 2011, 59, 799–811. [Google Scholar]
- McClure, C.J.W.; Rolek, B.W.; McDonald, K.; Hill, G.E. Climate change and the decline of a once common bird. Ecol. Evol. 2012, 2, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Bush, B.M.; Wissinger, S.A. Invertebrates in beaver-created wetlands and ponds. In Invertebrates in Freshwater Wetlands; Batzer, D., Boix, D., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 411–449. [Google Scholar]
- Westwood, A.R. Conservation of Three Forest Landbird Species at Risk: Characterizing and Modelling Habitat at Multiple Scales to Guide Management Planning. Ph.D. Thesis, Dalhousie University, Halifax, NS, Canada, 2016. [Google Scholar]
- Powell, L.L.; Hodgman, T.P.; Glanz, W.E. Home ranges of Rusty Blackbirds breeding in wetlands: How much would buffers from timber harvest protect habitat? Condor 2010, 112, 834–840. [Google Scholar] [CrossRef]
- Bohnett, E.; Hulse, D.; Ahmad, B.; Hoctor, T. Multi-level, multi-scale modeling and predictive mapping for jaguars in the Brazilian Pantanal. Open J. Ecol. 2020, 10, 243–263. [Google Scholar] [CrossRef]
- Schaefer, J.A.; Mayor, S.J. Geostatistics reveal the scale of habitat selection. Ecol. Model. 2007, 209, 401–406. [Google Scholar] [CrossRef]
- Duquette, C.A.; Hovick, T.J.; Limb, R.F.; McGranahan, D.A.; Sedivec, K.K. Restored fire and grazing regimes influence nest selection and survival in Brewer’s Blackbirds Euphagus Cyanocephalus. Acta Ornithol. 2019, 54, 171–180. [Google Scholar] [CrossRef]
- Hepp, M.; Ware, L.; van Oort, H.; Beauchesne, S.M.; Cooper, J.M.; Green, D.J. Postfledging survival and local recruitment of a riparian songbird in habitat influenced by reservoir operations. Avian Conserv. Ecol. 2018, 13, 12. [Google Scholar] [CrossRef]
- Gruebler, M.E.; Korner-Nievergelt, F.; Naef-Daenzer, B. Equal nonbreeding period survival in adults and juveniles of a long-distant migrant bird. Ecol. Evol. 2014, 4, 756–765. [Google Scholar] [CrossRef]
- Noel, B.L.; Bednarz, J.C.; Ruder, M.G.; Keel, M.K. Effects of radio-transmitter methods on pileated woodpeckers: An improved technique for large woodpeckers. Southeast. Nat. 2013, 12, 399–412. [Google Scholar] [CrossRef]
- Bale, S.; Beazley, K.F.; Westwood, A.; Bush, P. The benefits of using topographic features to predict climate-resilient habitat for migratory forest landbirds: An example for the Rusty Blackbird, Olive-sided Flycatcher, and Canada Warbler. Condor 2020, 122. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Silvics of North America: Volume 1. Conifers. In Agriculture Handbook 654; U.S. Forest Service, Department of Agriculture: Washington, DC, USA, 1990. [Google Scholar]
- Powell, L.L. Long Term Monitoring Plan for Rusty Blackbirds in the Atlantic Northern Forest; Version 1.8; University of Maine: Orono, ME, USA, 2008. [Google Scholar]
- Powell, L.L.; Hodgman, T.P.; Fiske, I.J.; Glanz, W.E. Habitat occupancy of rusty blackbirds (Euphagus carolinus) breeding in northern New England, USA. Condor 2014, 116, 122–133. [Google Scholar] [CrossRef]
- Sechrist, J.; Ahlers, D.D.; Zehfuss, K.P.; Doster, R.H.; Paxton, E.H.; Ryan, V.M. Home range and use of habitat of western yellow-billed cuckoos on the middle Rio Grande, New Mexico. Southwest. Nat. 2013, 58, 411–419. [Google Scholar] [CrossRef]
- Halterman, M.D. Sexual Dimorphism, Detection Probability, Home Range, and Parental Care in the Yellow-Billed Cuckoo. Ph.D. Thesis, University of Nevada, Reno, NV, USA, 2009. [Google Scholar]
- Laymon, S.A.; Halterman, M.D. A Proposed Habitat Management Plan for Yellow-Billed Cuckoos in California. In Proceedings of the California Riparian Systems Conference: Protection, Management, and Restoration for the 1990s, Davis, CA, USA, 22–24 September 1988; Gen. Tech. Rep. PSW-GTR-110; Abell, D.L., Ed.; Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Berkeley, CA, USA, 1989; pp. 272–277. [Google Scholar]
- Halterman, M.D.; Johnson, M.J.; Holmes, J.A.; Laymon, S.A. A Natural History Summary and Survey Protocol for the Western Distinct Population Segment of the Yellow-Billed Cuckoo; U.S. Fish and Wildlife Service: Washington, DC, USA, 2015; p. 45.



{kind=link}
{kind=link}
| Variable | Description |
|---|---|
| lowdrainage | % of buffer in class poorly and very poorly drained |
| lowslope | % of buffer that is 1−8% slope |
| stream123 | mean distance of buffer to 1st−3rd order streams |
| stream3456 | mean distance of buffer to 3rd−6th order streams |
| allstreams | mean distance of buffer to 1st−6th order streams |
| softwood | % softwood stands in buffer |
| swsaplings 1 | % softwood or mixed softwood (>75% SW) |
| mixage | % mixed-age stands in buffer |
| wetlands | % any vegetated palustrine wetland in buffer |
| alder | % alder wetland in buffer |
| beaver | % beaver pond wetland buffer |
| forestwl | % forested wetland in buffer |
| seasonfl | % seasonally flooded wetland in buffer |
| Candidate Models | k | AICc | ΔAICc | ωi | cum. ωi | LL |
|---|---|---|---|---|---|---|
| Nesting | ||||||
| allstreams+swsaplings+lowslope | 5 | 93.28 | 0 | 0.62 | 0.62 | −41.3 |
| allstreams+swsaplings+lowslope+wetlands | 6 | 95.24 | 1.95 | 0.23 | 0.85 | −41.14 |
| stream123+swsaplings+lowslope+wetlands | 6 | 96.81 | 3.53 | 0.11 | 0.96 | −41.92 |
| allstreams+swsaplings+wetlands | 5 | 99.54 | 6.25 | 0.03 | 0.99 | −44.43 |
| Null | 2 | 130.16 | 36.87 | 0 | 1 | −63.01 |
| Fledgling | ||||||
| allstreams+swsaplings+lowslope+wetlands | 6 | 404.91 | 0 | 0.73 | 0.73 | −196.35 |
| allstreams+swsaplings+lowslope+wetlands+softwood | 7 | 406.97 | 2.06 | 0.26 | 0.99 | −196.34 |
| Null | 3 | 543.62 | 138.71 | 0 | 1 | −268.78 |
| Adult | ||||||
| allstreams+swsaplings+lowslope+wetlands | 6 | 1164.69 | 0 | 0.73 | 0.73 | −576.31 |
| allstreams+swsaplings+lowslope+wetlands+softwood | 7 | 1166.68 | 1.99 | 0.27 | 0.99 | −576.29 |
| allstreams+mixage+lowslope+wetlands+softwood | 7 | 1174.46 | 9.77 | 0.01 | 1 | −580.18 |
| Null | 3 | 1568.82 | 404.13 | 0 | 1 | −781.40 |
| Variable | Estimate | SE | LCI | UCI | Unit | Scaled | Scaled CL |
|---|---|---|---|---|---|---|---|
| Change | Estimate | ||||||
| Fledglings | |||||||
| Intercept | −0.92 | 0.38 | −1.66 | −0.18 | |||
| distance to streams (1st–6th) | −30.3 | 7.76 | −45.5 | −15 | 50m | 4.55 | 2.1, 9.7 |
| proportion swsaplings | 1.8 | 0.34 | 1.1 | 2.4 | 10% | 1.2 | 1.1, 1.3 |
| proportion low slope (1−8%) | 1.18 | 0.3 | 0.59 | 1.77 | -- | -- | |
| proportion wetland | 2.9 | 0.6 | 1.71 | 4 | 10% | 1.33 | 1.2, 1.5 |
| Adults | |||||||
| Intercept | −0.33 | 0.15 | −0.62 | −0.04 | |||
| distance to streams (1st–6th) | −3.7 | 0.4 | −4.4 | −2.9 | 50m | 1.2 | 1.2, 1.3 |
| proportion swsaplings | 1.2 | 0.2 | 0.83 | 1.56 | -- | -- | |
| proportion low slope (1−8%) | 0.87 | 0.2 | 0.56 | 1.17 | -- | -- | |
| proportion wetland | 2.7 | 0.4 | 1.95 | 3.38 | 10% | 1.31 | 1.2, 1.4 |
| Nests | |||||||
| Intercept | −0.87 | 0.55 | −1.95 | 0.21 | |||
| distance to streams (1st–6th) | −5.16 | 1.65 | −8.39 | −1.93 | 50m | 1.3 | 1.1, 1.5 |
| proportion swsaplings | 0.02 | 0.01 | 0.01 | 0.04 | 10% | 1.2 | 1.1, 1.5 |
| proportion low slope (1−8%) | 0.01 | 0.01 | 0 | 0.03 | 10% | 1.1 | 1, 1.3 |
| proportion wetland | 0.01 | 0.01 | −0.02 | 0.04 | -- | -- |
| Candidate Models | k | AICc | ΔAICc | ωi | Deviance |
|---|---|---|---|---|---|
| Nests | |||||
| S(~wetlands) | 2 | 72.8 | 0 | 0.19 | 68.76 |
| S(~nestage + wetlands) | 3 | 73.5 | 0.69 | 0.13 | 67.42 |
| S(~nestage + wetlands + allstreams) | 4 | 74.3 | 1.55 | 0.09 | 66.24 |
| S(~allstreams) | 2 | 74.5 | 1.72 | 0.08 | 70.48 |
| S(~wetlands + lowslope) | 3 | 74.8 | 2.00 | 0.07 | 68.73 |
| S(~nestage + wetlands + lowslope) | 4 | 75.1 | 2.26 | 0.06 | 66.95 |
| S(~1) | 1 | 75.2 | 2.38 | 0.06 | 73.16 |
| S(~nestage + allstreams) | 3 | 75.9 | 3.07 | 0.04 | 69.8 |
| S(~nestage) | 2 | 76.0 | 3.16 | 0.04 | 71.92 |
| S(~allstreams + swsaplings) | 3 | 76.3 | 3.55 | 0.03 | 70.28 |
| Adult males and females and fledglings (hy) | |||||
| S(~swsaplings + hy + female + male) | 5 | 100.2 | 0 | 0.32 | 90.19 |
| S(~hy + female + male) | 4 | 102.0 | 1.8 | 0.13 | 94.02 |
| S(~lowslope + allstreams) | 3 | 102.9 | 2.6 | 0.09 | 96.84 |
| S(~time + hy + female + male) | 5 | 103.3 | 3.1 | 0.07 | 93.29 |
| S(~lowslope + hy + female + male) | 5 | 103.9 | 3.7 | 0.057 | 93.88 |
| S(~lowslope) | 2 | 104.0 | 3.8 | 0.05 | 99.98 |
| S(~1) | 1 | 104.1 | 3.9 | 0.05 | 102.09 |
| S(~SWDI) | 2 | 104.1 | 3.9 | 0.05 | 100.13 |
| S(~wetlands + lowslope) | 3 | 105.0 | 4.7 | 0.035 | 98.95 |
| Survivorship Parameters | Survival Estimate | SE | LCI | UCI | Overall |
|---|---|---|---|---|---|
| Nest survival daily | 0.975 | 0.26 | 0.95 | 0.99 | 0.48 |
| Fledgling survivorship | 0.988 | 0.336 | 0.97 | 0.99 | 0.49 |
| Female survivorship | 0.998 | 0.63 | 0.99 | 1 | 0.89 1 |
| Male survivorship | 0.996 | 0.44 | 0.99 | 1 | 0.79 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wohner, P.J.; Foss, C.R.; Cooper, R.J. Rusty Blackbird Habitat Selection and Survivorship during Nesting and Post-Fledging. Diversity 2020, 12, 221. https://doi.org/10.3390/d12060221
Wohner PJ, Foss CR, Cooper RJ. Rusty Blackbird Habitat Selection and Survivorship during Nesting and Post-Fledging. Diversity. 2020; 12(6):221. https://doi.org/10.3390/d12060221
Chicago/Turabian StyleWohner, Patricia J., Carol R. Foss, and Robert J. Cooper. 2020. "Rusty Blackbird Habitat Selection and Survivorship during Nesting and Post-Fledging" Diversity 12, no. 6: 221. https://doi.org/10.3390/d12060221
APA StyleWohner, P. J., Foss, C. R., & Cooper, R. J. (2020). Rusty Blackbird Habitat Selection and Survivorship during Nesting and Post-Fledging. Diversity, 12(6), 221. https://doi.org/10.3390/d12060221