Conservation Lessons from the Study of North American Boreal Birds at Their Southern Periphery

{kind=link}
Abstract
1. Introduction
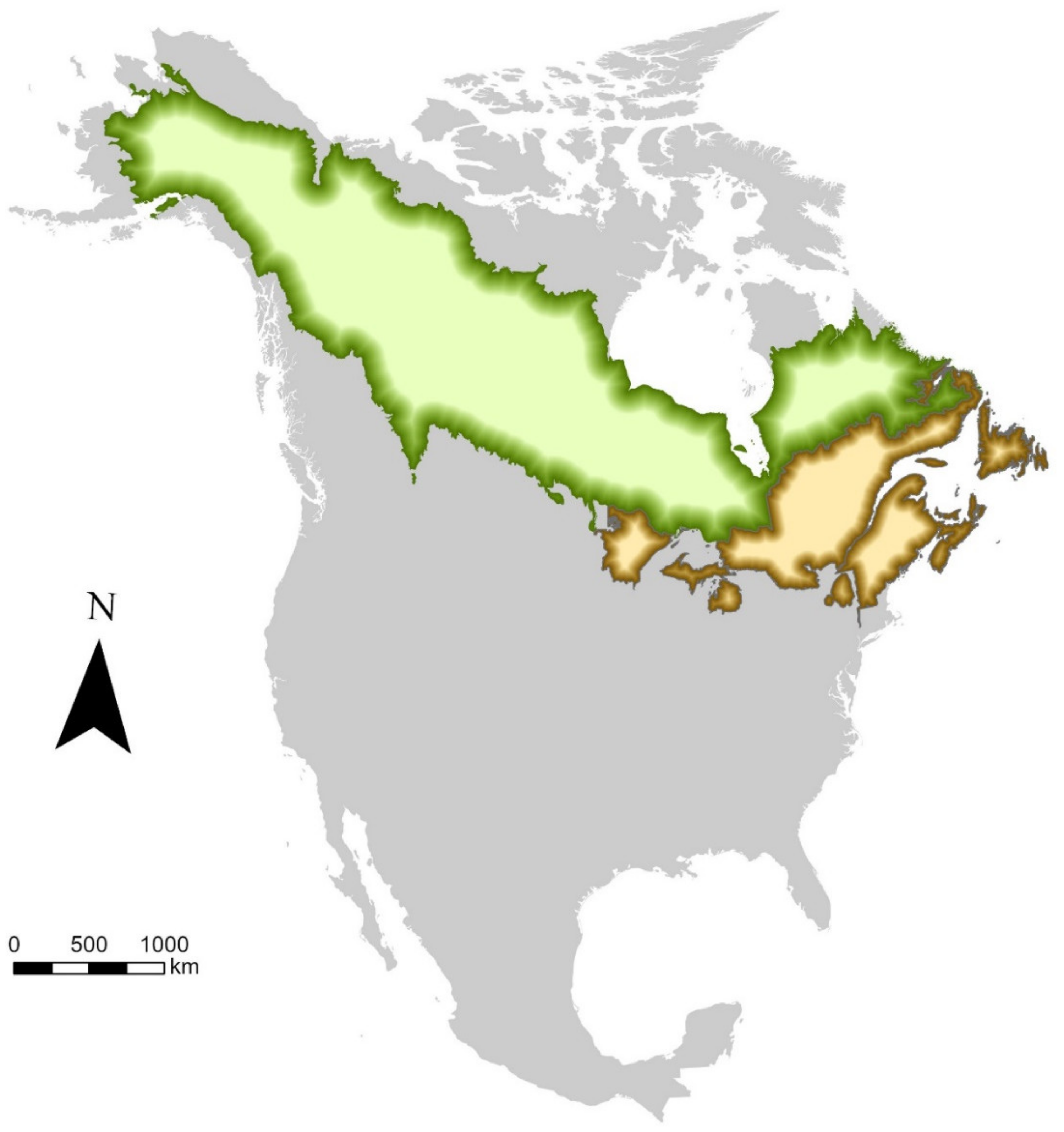
2. Peripheral Populations of Boreal Birds Are Genetically Unique, Threatened by Climate Change, and Declining
3. Species Differ in Their Responses to Changing Environments, Creating a Challenge for Community Level Conservation
4. As Communities Reshuffle, Species Face New Biotic Interactions—Some of Which May Have Unexpected Outcomes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wells, J.; Stralberg, D.; Childs, D. Boreal Forest Refuge: Conserving North America’s Bird Nursery in the Face of Climate Change; Boreal Songbird Initiative: Seattle, WA, USA, 2018. [Google Scholar]
- Blancher, P.; Wells, J. The Boreal Forest Region: North. America’s Bird Nursery; Canadian Boreal Initiative and Boreal Songbird Initiative: Ottawa, ON, Canada, 2005; pp. 1–12. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; 1535p. [Google Scholar]
- Walker, X.J.; Baltzer, J.L.; Cumming, S.G.; Day, N.J.; Ebert, C.; Goetz, S.; Johnstone, J.F.; Potter, S.; Rogers, B.M.; Schuur, E.A.; et al. Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature 2019, 572, 520–525. [Google Scholar] [CrossRef]
- Nelson, T.A.; Coops, N.C.; Wulder, M.A.; Perez, L.; Fitterer, J.; Powers, R.; Fontana, F. Predicting climate change impacts to the Canadian boreal forest. Diversity 2014, 6, 133–157. [Google Scholar] [CrossRef]
- Stralberg, D.; Berteaux, D.; Drever, C.R.; Drever, M.; Naujokaitis-Lewis, I.; Schmiegelow, F.K.A.; Tremblay, J.A. Conservation planning for boreal birds in a changing climate: A framework for action. Avian Conserv. Ecol. 2019, 14, 13. [Google Scholar] [CrossRef]
- Bateman, B.L.; Wilsey, C.; Taylor, L.; Wu, J.; LeBaron, G.S.; Langham, G.M. North American birds require mitigation and adaptation to reduce vulnerability to climate change. Conserv. Sci. Pract. 2019, in press. [Google Scholar]
- Damman, A.W.H. An ecological subdivision of the island of Newfoundland. In Biogeography and Ecology of the Island of Newfoundland; South, G.R., Ed.; Dr. W. Junk Publishers: The Hague, The Netherlands, 1983; pp. 163–206. [Google Scholar]
- Anderson, M.G.; Clark, M.; Ferree, C.E.; Jospe, A.; Olivero Sheldon, A.; Weaver, K.J. Northeast. Habitat Guides: A Companion to the Terrestrial and Aquatic Habitat Maps; The Nature Conservancy, Eastern Conservation Science: Boston, MA, USA, 2013. [Google Scholar]
- Zukerburg, B.; Woods, A.M.; Porter, W.F. Poleward shufts in breeding bird distributions in New York State. Glob. Chang. Biol. 2009, 15, 1866–1883. [Google Scholar] [CrossRef]
- Ralston, J.; Kirchman, J.J. Predicted range shifts in North American boreal forest birds and the effect of climate change on genetic diversity in blackpoll warblers (Setophaga striata). Conserv. Genet. 2013, 14, 543–555. [Google Scholar] [CrossRef]
- DeLuca, W.V.; King, D.I. Montane birds shift downslope despite recent warming in the northern Appalachian Mountains. J. Ornithol. 2017, 158, 493–505. [Google Scholar] [CrossRef]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The spread of invasive species and infectious disease as drivers of ecosystem change. Front. Ecol. Environ. 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Fuller, T.; Bensch, S.; Müller, I.; Novembre, J.; Pérez-Tris, J.; Ricklefs, R.E.; Smith, T.B.; Waldenström, J. The ecology of emerging infectious diseases in migratory birds: An assessment of the role of climate change and priorities for future research. EcoHealth 2012, 9, 80–88. [Google Scholar] [CrossRef]
- Whitaker, D.M. The colonization of Newfoundland by red squirrels (Tamasciurus hudsonicus). Osprey Nat. J. Nfld. Labrador 2015, 46, 23–29. [Google Scholar]
- Imbeau, L.; Mönkkönen, M.; Desrochers, A. Long-term effects of forestry on birds of the eastern Canadian boreal forests: A comparison with Fennoscandia. Conserv. Biol. 2001, 15, 1151–1162. [Google Scholar] [CrossRef]
- Rimmer, C.C.; McFarland, K.P.; Evers, D.C.; Miller, E.K.; Aubry, Y.; Busby, D.; Taylor, R.J. Mercury concentrations in Bicknell’s Thrush and other insectivorous passerines in montane forests of Northeastern North America. Ecotoxicology 2005, 14, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Venier, L.A.; Thompson, I.D.; Fleming, R.; Malcolm, J.; Aubin, I.; Trofymow, J.A.; Langor, D.; Sturrock, R.; Patry, C.; Outerbridge, R.O.; et al. Effects of natural resource development on the terrestrial biodiversity of Canadian boreal forest. Environ. Rev. 2014, 22, 457–490. [Google Scholar] [CrossRef]
- Zlonis, E.J.; Niemi, G.J. Avian communities of managed and wilderness hemiboreal forests. For. Ecol. Manag. 2014, 328, 26–34. [Google Scholar] [CrossRef]
- DeLuca, W.V.; King, D.I. Influence of hiking trails on montane birds. J. Wildlife Manag. 2014, 78, 494–502. [Google Scholar] [CrossRef]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, J.J.; Ralston, J. The Adirondack archipelago. Adirondack J. Environ. Stud. 2015, 20, 5. [Google Scholar]
- Ralston, J.; FitzGerald, A.M.; Burg, T.M.; Starkloff, N.C.; Whitaker, D.M.; Warkentin, I.G.; Norris, D.R.; Kirchman, J.J. Comparative phylogeographic analysis suggests a shared history among eastern North American boreal forest birds. Glob. Ecol. Biogeog. Under review.
- Kirchman, J.J.; Ross, A.M.; Johnson, G. Historical decline of genetic diversity in a range-periphery population of Spruce Grouse (Falcipennis canadensis) inhabiting the Adirondack Mountains. Conserv. Genet. 2020, 21, 373–380. [Google Scholar] [CrossRef]
- Van Els, P.; Cicero, C.; Klicka, J. High latitude and high genetic diversity: Phylogeography of a widespread boreal bird, the gray jay (Perisoreus candensis). Mol. Phylogenet. Evol. 2012, 63, 456–465. [Google Scholar] [CrossRef]
- Dohms, K.M.; Graham, B.A.; Burg, T.M. Multilocus genetic analyses and spatial modeling reveal complex population structure and history in a widespread resident North American passerine (Perisoreus canadensis). Ecol. Evol. 2017, 7, 9869–9889. [Google Scholar] [CrossRef] [PubMed]
- Lait, L.A.; Burg, T.M. When east meets west: Population structure of a high-latitude resident species, the boreal chickadee (Poecile hudsonicus). Heredity 2013, 111, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Ralston, J.; Kirchman, J.J. Continent-scale genetic structure in a boreal forest migrant, the blackpoll warbler (Steophaga striata). Auk 2012, 129, 467–478. [Google Scholar]
- FitzGerald, A.M.; Weir, J.; Ralston, J.; Warkentin, I.G.; Whitaker, D.M.; Kirchman, J.J. Genetic structure and biogeographical history of the Bicknell’s Thrush/Gray-cheeked Thrush species complex. Auk 2020, 137, 1–20. [Google Scholar] [CrossRef]
- Zink, R.M. The geography of mitochondrial DNA variation, population structure, hybridization, and species limits in the fox sparrow (Passerella iliaca). Evolution 1994, 48, 96–111. [Google Scholar]
- Townsend, J.M.; McFarland, K.P.; Rimmer, C.C.; Ellison, W.G.; Goetz, J.E. Bicknell’s Thrush (Catharus bicknelli), version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- FitzGerald, A.M.; Whitaker, D.M.; Ralston, J.; Kirchman, J.J.; Warkentin, I.G. Taxonomy and distribution of the imperiled Newfoundland Gray-cheeked Thrush, Catharus minimus minimus. Avian Conserv. Ecol. 2017, 12, 10. [Google Scholar] [CrossRef]
- FitzGerald, A.M. Division within the North American boreal forest: Ecological niche divergence between the Bicknell’s Thrush (Cathatus bicknelli) and Gray-cheeked Thrush (C. minimus). Ecol. Evol. 2017, 7, 5285–5295. [Google Scholar] [CrossRef] [PubMed]
- Designing Sustainable Landscapes: Modeling Focal Species. Report to the North Atlantic Conservation Cooperative, US Fish and Wildlife Service, Northeast Region. Available online: http://jamba.provost.ads.umass.edu/web/lcc/dsl/technical/DSL_documentation_species.pdf (accessed on 22 May 2020).
- Loman, Z.G.; DeLuca, W.V.; Harrison, D.; Loftin, C.S.; Wood, P.B. Landscape capability models as a tool to predict fine-scale forest bird occupancy and abundance. Landsc. Ecol. 2018, 33, 77–91. [Google Scholar] [CrossRef]
- Able, K.P.; Noon, B.R. Avian community structure along elevational gradients in the Northeastern United States. Oecologia 1979, 26, 275–294. [Google Scholar] [CrossRef]
- Kirchman, J.J.; Van Keuren, A.E. Altitudinal range shifts of birds at the southern periphery of the boreal forest: 40 years of change in the Adirondacks Mountains. Wilson J. Ornithol. 2017, 129, 742–753. [Google Scholar] [CrossRef]
- Beckage, B.; Osborne, B.; Gavin, D.G.; Pucko, C.; Siccama, T.; Perkins, T. A rapid upward shift of the forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc. Natl. Acad. Sci. USA 2008, 105, 4197–4202. [Google Scholar] [CrossRef] [PubMed]
- Elsen, P.R.; Tingley, M.W. Global mountain topography and the fate of montane species under climate change. Nat. Clim. Chang. 2015, 5, 772–776. [Google Scholar] [CrossRef]
- Foster, J.R.; D’Amato, A.W. Montane forest ecotones moved downslope in northeastern USA in spite of warming between 1984 and 2011. Glob. Chang. Biol. 2015, 21, 4497–4507. [Google Scholar] [CrossRef] [PubMed]
- Hanowski, J.A.; Niemi, G.J. A comparison of on- and off-road bird counts: Do you need to go off road to count birds accurately? J. Field Ornithol. 1995, 66, 469–483. [Google Scholar]
- Ralston, J.; King, D.I.; DeLuca, W.V.; Niemi, G.J.; Glennon, M.J.; Scarl, J.C.; Lambert, J.D. Analysis of combined data sets yields trend estimates for vulnerable spruce-fir birds in northern United States. Biol. Conserv. 2015, 187, 270–278. [Google Scholar] [CrossRef]
- Sólymos, P.; Toms, J.D.; Matsuoka, S.M.; Cumming, S.G.; Barker, N.K.; Thogmartin, W.E.; Stralberg, D.; Crosby, A.D.; Dénes, F.V.; Haché, S.; et al. Lessons learned from comparing spatially explicit models and the Partners in Flight approach to estimate population sizes of boreal birds in Alberta, Canada. Condor 2020. [Google Scholar] [CrossRef]
- Partners in Fight Science Committee: Partners in Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States. Available online: https://www.partnersinflight.org/wp-content/uploads/2016/08/pif-continental-plan-final-spread-single.pdf. (accessed on 22 May 2020).
- Howe, R.H.; Roberts, L.J. Sixteen Years of Habitat-Based Bird Monitoring in the Nicolet National Forest; USDA Forest Service General Technical Report PSW-GTR-191; USDA Forest Service: Washington, DC, USA, 2005.
- US Forest Service. White Mountain National Forest, Monitoing and Valuation Report; US Department of Argiculture: Washinginton, DC, USA, 2006.
- Johnson, B. The Ottawa National Forest Breeding Bird Census, an Analysis of Twenty Years of Data, 1992–2011; Report Submitted to Ottawa National Forest; USDA Forest Service: Ironwood, MI, USA, 2012.
- Zlonis, E.J.; Grinde, A.; Bednar, J.; Niemi, G.J. Summary of Breeding Bird Trends in the Chippewa and Superior National Forests of Minnesota- 1995–2013; NRRI Technical Report NRRI/TR-2013/36; University of Minnnesota: Duluth, MN, USA, 2013. [Google Scholar]
- Faccio, S.D.; Mitchell, B.R. Breeding Landbird Monitoring in the Northeast; Temperate Network, 2013 Summary Report; Natural Resource Data Series NPS/NETN/NRDS-2014/630; National Park Service: Fort Collins, CO, USA, 2014. [Google Scholar]
- Glennon, M.J.; Langdon, S.F.; Rubenstein, M.A.; Cross, M.S. Temporal changes in avian community composition in lowland conifer habitats at the southern edge of the boreal zone in the Adirondack Park, NY. PLoS ONE 2019, 14, e0220927. [Google Scholar] [CrossRef]
- Hill, J.M. The State of the Mountain Birds Report: 2020; Vermont Center for Ecostudies: White River Junction, VT, USA, 2020. [Google Scholar]
- Duclos, T.R.; DeLuca, W.V.; King, D.I. Direct and indirect effects of climate on bird abundance along elevational gradients in the Northern Appalachian Mountains. Divers. Distrib. 2019, 25, 1670–1683. [Google Scholar] [CrossRef]
- Ralston, J.; FitzGerald, A.M.; Scanga, S.E.; Kirchman, J.J. Observations of habitat associations in boreal forest birds and the geographic variation in bird community composition. Wilson J. Ornithol. 2019, 131, 12–23. [Google Scholar] [CrossRef]
- Glennon, M.J.; Langdon, S.F.; Rubenstein, M.A.; Cross, M.S. Relative contribution of climate and non-climate drivers in determining dynamic rates of boreal birds at the edge of their range. PLoS ONE 2019, 14, e0224308. [Google Scholar] [CrossRef]
- Niemi, G.J.; Hanowski, J.M.; Lima, A.R.; Nicholls, T.; Weiland, N. A critical analysis on the use of indicator species in management. J. Wildlife Manag. 1997, 61, 1240–1252. [Google Scholar] [CrossRef]
- Aubry, Y.; Desrochers, A.; Seutin, G. Regional patterns of habitat use by a threatened forest bird, the Bicknell’s Thrush (Catharus bicknelli), in Quebec. Can. J. Zool. 2016, 94, 301–309. [Google Scholar] [CrossRef]
- Chisholm, S.E.; Leonard, M.L. Effect of forest management on a rare habitat specialist, the Bicknell’s Thrush (Catharus bicknelli). Can. J. Zool. 2008, 86, 217–223. [Google Scholar] [CrossRef]
- Keith, S.A.; Newton, A.C.; Herbert, R.J.H.; Morecroft, M.D.; Bealey, C.E. Non-analogous community formation in response to climate change. J. Nat. Conserv. 2009, 17, 228–235. [Google Scholar] [CrossRef]
- MacArthur, R.H. Geographical Ecology: Patterns in the Distribution of Species; Harper and Row: New York, NY, USA, 1972. [Google Scholar]
- Brown, J.H.; Lomolino, M.V. Biogeography, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 1998. [Google Scholar]
- Freeman, B.G.; Montgomery, G. Interspecific aggression by the Swainson’s Thrush (Catharus ustulatus) may limit the distribution of the threatened Bicknell’s Thrush (Catharus bicknelli) in the Adirondack Mountains. Condor 2016, 118, 169–178. [Google Scholar] [CrossRef]
- Noon, B.R. The distribution of an avian guild along a temperate elevational gradient: The importance and expression of competition. Ecol. Monogr. 1981, 51, 105–124. [Google Scholar] [CrossRef]
- MacArthur, R.H. Population ecology of some warblers of northeastern coniferous forests. Ecology 1958, 39, 599–619. [Google Scholar] [CrossRef]
- Gill, F.B. Local cytonuclear extinction of the golden-winged warbler. Evolution 1997, 51, 519–525. [Google Scholar] [CrossRef]
- Vallender, R.; Van Wilgenburg, S.L.; Bulluck, L.P.; Roth, A.; Canterbury, R.; Larkin, J.; Fowlds, R.M.; Lovette, I.J. Extensive rangewide mitochondrial introgression indicated substantial cryptic hybridization in the golden-winged warbler (Vermivora chrysoptera). Avian Conserv. Ecol. 2009, 4, 4. [Google Scholar] [CrossRef]
- Kearns, A.M.; Restani, M.; Szabo, I.; Schrøder-Nielsen, A.; Kim, J.A.; Richardson, H.M.; Marzluff, J.M.; Fleischer, R.C.; Johnsen, A.; Omland, K.E. Genomic evidence of speciation reversal in ravens. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Rhymer, J.M.; Simberloff, D. Extinction by hybridization and introgression. Annu. Rev. Ecol. Syst. 1996, 27, 83–109. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Leary, R.F.; Spurell, P.; Wenburg, J.K. The problems with hybrids: Setting conservation guidelines. Trends Ecol. Evol. 2001, 16, 613–622. [Google Scholar] [CrossRef]
- Marshall, J.T. The Gray-Cheeked Thrush, Catharus minimus, and Its New England Subspecies, Bicknell’s thrush, Catharus minimus bicknelli. No. 28; Nuttall Ornithological Club: Cambridge, MA, USA, 2001. [Google Scholar]
- Morelli, T.L.; Hallworth, M.T.; Duclos, T.; Siren, A.; Ells, A.; Faccio, S.; McFarland, K.; Nislow, K.; Ralston, J.; Ratnaswamy, M.; et al. Habitat causes lags in response to climate change for a climate-vulnerable species. Glob. Ecol. Biogeog. Under review.
- Hallworth, M.; Siren, A.; DeLuca, W.V.; Duclos, T.; McFarland, K.P.; Hill, J.; Rimmer, C.C.; Morelli, T.L. Boom and bust: Pulsed resources drive distribution dynamics in forested food webs. Glob. Ecol. Biogeog. Under review.
- Whitaker, D.M.; Taylor, P.D.; Warkentin, I.G. Gray-cheeked Thrush (Catharus minimus minimus) distribution and habitat use in a montane forest landscape of western Newfoundland, Canada. Avian Conserv. Ecol. 2015, 10, 4. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ralston, J.; DeLuca, W.V. Conservation Lessons from the Study of North American Boreal Birds at Their Southern Periphery. Diversity 2020, 12, 257. https://doi.org/10.3390/d12060257
Ralston J, DeLuca WV. Conservation Lessons from the Study of North American Boreal Birds at Their Southern Periphery. Diversity. 2020; 12(6):257. https://doi.org/10.3390/d12060257
Chicago/Turabian StyleRalston, Joel, and William V. DeLuca. 2020. "Conservation Lessons from the Study of North American Boreal Birds at Their Southern Periphery" Diversity 12, no. 6: 257. https://doi.org/10.3390/d12060257
APA StyleRalston, J., & DeLuca, W. V. (2020). Conservation Lessons from the Study of North American Boreal Birds at Their Southern Periphery. Diversity, 12(6), 257. https://doi.org/10.3390/d12060257