Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy


Abstract

1. Introduction
2. Materials and Methods
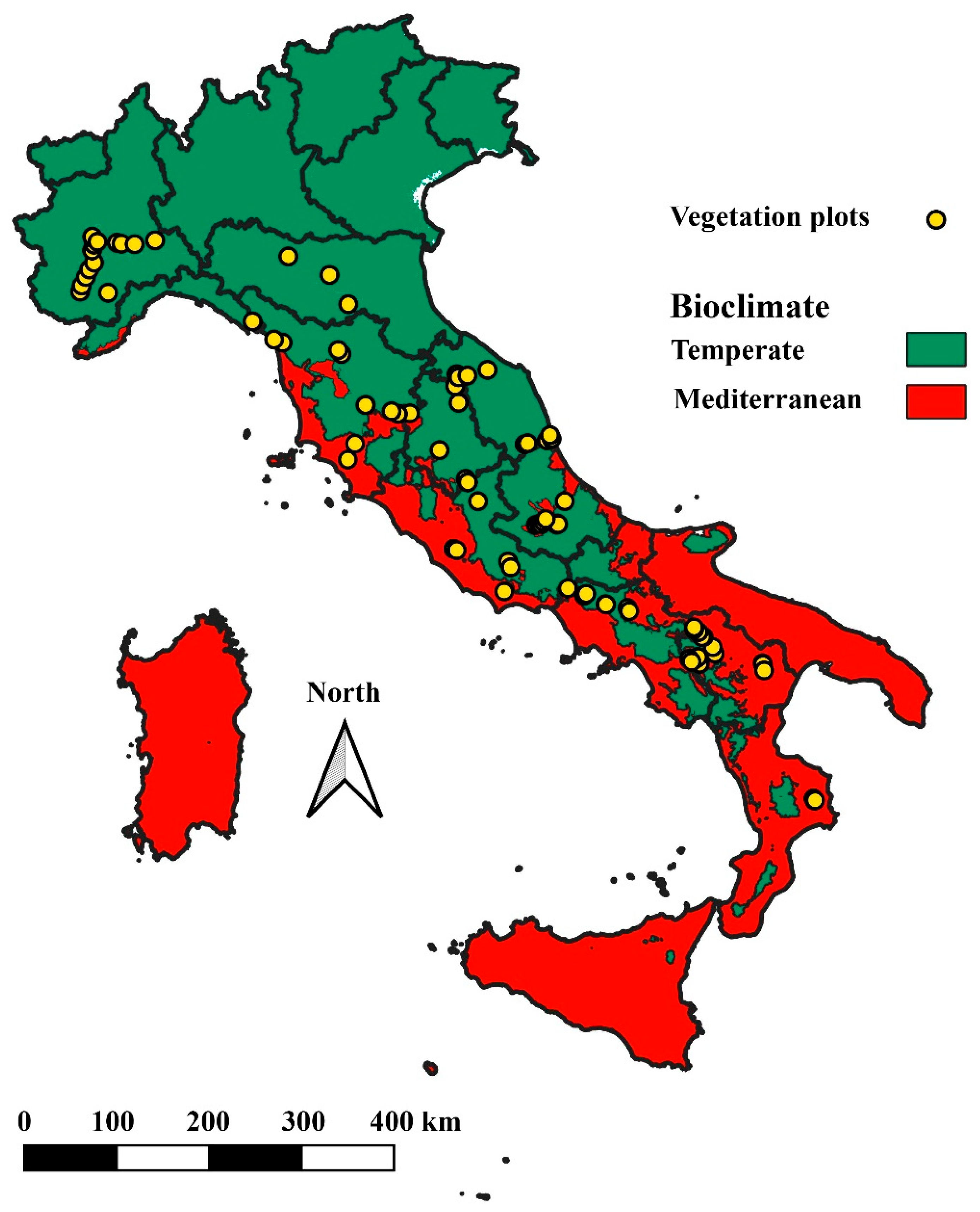
2.1. Study Area and Sampling Design
2.2. Dataset Preparation and Statistical Analyses
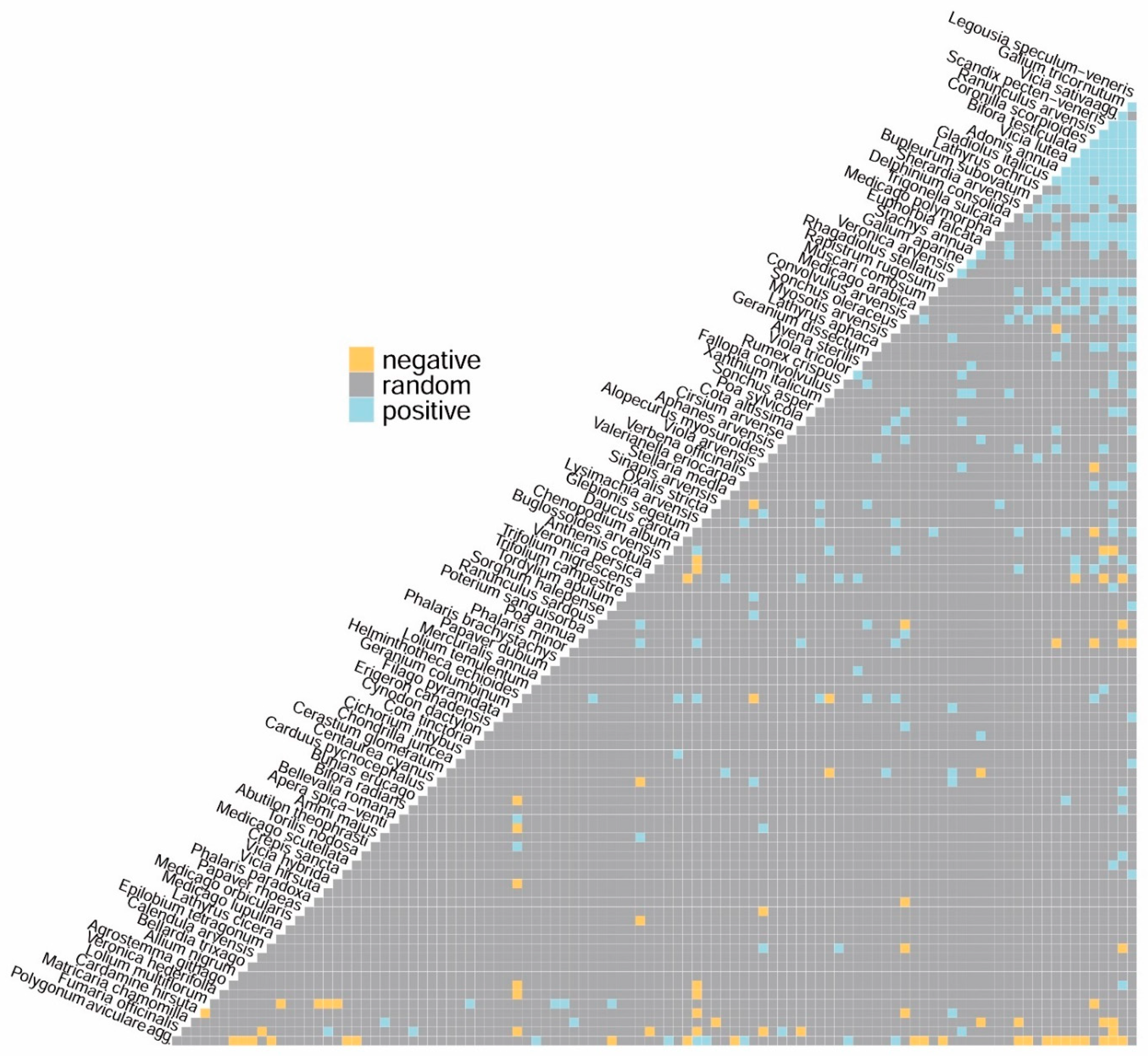
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological assembly rules in plant communities—Approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D.; Vandermast, D.B.; Kuppinger, D.M.; Manthey, M.; Peet, R.K. Co-occurrence based assessment of habitat generalists and specialists: A new approach for the measurement of niche width. J. Ecol. 2007, 95, 707–722. [Google Scholar] [CrossRef]
- Pannek, A.; Manthey, M.; Diekmann, M. Comparing resource-based and co-occurrence-based methods for estimating species niche breadth. J. Veg. Sci. 2016, 27, 596–605. [Google Scholar] [CrossRef]
- Bellanger, S.; Darmency, H.; Guillemin, J.P. Relationships between weed diversity and Centaurea cyanus. In Proceedings of the XIIIème Colloque International sur la Biologie des Mauvaises Herbes, Dijon, France, 8–10 September 2009; pp. 8–15. [Google Scholar]
- Bellanger, S.; Guillemin, J.P.; Bretagnolle, V.; Darmency, H. Centaurea cyanus as a biological indicator of segetal species richness in arable fields. Weed Res. 2012, 52, 551–563. [Google Scholar] [CrossRef]
- Petit, S.; Fried, G. Patterns of weed co-occurrence at the field and landscape level. J. Veg. Sci. 2012, 23, 1137–1147. [Google Scholar] [CrossRef]
- Holzner, W. Weed species and weed communities. Vegetatio 1978, 38, 13–20. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Bretagnolle, V.; Gaba, S. Weeds for bees? A review. Agron Sustain Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef]
- Blaix, C.; Moonen, A.C.; Dostatny, D.F.; Izquierdo, J.; Le Corff, J.; Morrison, J.; Von Redwitz, C.; Schumacher, M.; Westerman, P.R. Quantification of regulating ecosystem services provided by weeds in annual cropping systems using a systematic map approach. Weed Res. 2018, 58, 151–164. [Google Scholar] [CrossRef]
- Weisberger, D.; Nichols, V.; Liebman, M. Does diversifying crop rotations suppress weeds? A meta-analysis. PLoS ONE 2019, 14, e0219847. [Google Scholar] [CrossRef]
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Šilc, U.; Vrbničanin, S.; Božić, D.; Čarni, A.; Dajić Stevanović, Z. Weed vegetation in the north-western Balkans: Diversity and species composition. Weed Res. 2009, 49, 602–612. [Google Scholar] [CrossRef]
- Mahaut, L.; Gaba, S.; Fried, G. A functional diversity approach of crop sequences reveals that weed diversity and abundance show different responses to environmental variability. J. Appl. Ecol. 2019, 56, 1400–1409. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Petit, S.; Dessaint, F.; Rosati, L.; Abbate, G. Species composition, richness, and diversity of weed communities of winter arable land in relation to geo-environmental factors: A gradient analysis in mainland Italy. Botany 2020. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Cimalová, S.; Kropáč, Z.; Otýpková, Z.; Pyšek, P.; Tichý, L. Weed vegetation of arable land in central Europe: Gradients of diversity and species composition. J. Veg. Sci. 2004, 15, 415–422. [Google Scholar] [CrossRef]
- Fried, G.; Norton, R.L.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Nowak, A.; Nowak, S.; Nobis, M.; Nobis, A. Crop type and altitude are the main drivers of species composition of arable weed vegetation in Tajikistan. Weed Res. 2015, 55, 525–536. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 3–264. [Google Scholar] [CrossRef]
- Zohary, M. Plant Life of Palestine, Israel and Jordan; Ronald: New York, NY, USA, 1962. [Google Scholar]
- Scholz, H. Questions about indigenous plants and anecophytes. Taxon 2007, 56, 1255–1260. [Google Scholar] [CrossRef]
- Brullo, S.; Guarino, R. The Mediterranean weedy vegetation and its origin. Ann. di Bot. 2007, 7, 101–110. [Google Scholar]
- Abbate, G.; Cicinelli, E.; Iamonico, D.; Iberite, M. Floristic analysis of the weed communities in wheat and corn crops: A case study in western-central Italy. Ann. di Bot. 2013, 3, 97–105. [Google Scholar]
- Latini, M.; Fanfarillo, E.; De Luca, E.; Iberite, M.; Abbate, G. The weed vegetation of the bean “Fagiolo Cannellino di Atina” and the red pepper “Peperone di Pontecorvo” PDO crops (Latium, central Italy). Plant Sociol. 2020, 57, 1–10. [Google Scholar]
- Janssen, J.A.M.; Rodwell, J.S.; García Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Ssymank, A.; et al. European Red List of Habitats—Part 2: Terrestrial and Freshwater Habitats; Publication Office of the European Union: Luxembourg, 2016; pp. 1–38. [Google Scholar]
- Fanfarillo, E.; Latini, M.; Iberite, M.; Bonari, G.; Nicolella, G.; Rosati, L.; Salerno, G.; Abbate, G. The segetal flora of winter cereals and allied crops in Italy: Species inventory with chorological, structural and ecological features. Plant Biosyst. 2020. [Google Scholar] [CrossRef]
- Albrecht, H. Suitability of arable weeds as indicator organism to evaluate species conservation effects of management in agricultural ecosystems. Agr. Ecosyst. Environ. 2003, 98, 201–211. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reboud, X. A functional analysis of large-scale temporal shifts from 1970 to 2000 in weed assemblages of sunflower crops in France. J. Veg. Sci. 2009, 20, 49–58. [Google Scholar] [CrossRef]
- Richner, N.; Holderegger, R.; Linder, H.P.; Walter, T. Reviewing change in the arable flora of Europe: A meta-analysis. Weed Res. 2015, 55, 1–13. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Kasperski, A.; Giuliani, A.; Abbate, G. Shifts of arable plant communities after agricultural intensification: A floristic and ecological diachronic analysis in maize fields of Latium (central Italy). Bot. Lett. 2019, 166, 356–365. [Google Scholar] [CrossRef]
- Hilbig, W. Preservation of agrestal weeds. In Biology and Ecology of Weeds; Holzner, W., Numata, M., Eds.; Dr W. Junk Publishers: The Hague, The Netherlands, 1982; pp. 57–59. [Google Scholar]
- Aboucaya, A.; Jauzein, P.; Vinciguerra, L.; Virevaire, M. Plan National d’action pour la Conservation des Plantes Messicoles. Rapport Final rédigé à la Demande du Ministère de l’Aménagement du Territoire et de l’Environnement, Direction De La Nature Et Des Paysages; Conservatoire Botanique National Méditerranéen de Porquerolles: Hyères, France, 2000. [Google Scholar]
- Cambecèdes, J.; Largier, G.; Lombard, A. Plan National d’actions en Faveur des Plantes Messicoles; Conservatoire Botanique National des Pyrénées et de Midi-Pyrénées: Bagnères de Bigorre, France, 2012. [Google Scholar]
- Storkey, J.; Meyer, S.; Still, K.S.; Leuschner, C. The impact of agricultural intensification and land-use change on the European arable flora. Proc. R. Soc. B. 2012, 279, 1421–1429. [Google Scholar] [CrossRef]
- Nowak, A.; Nowak, S.; Nobis, M.; Nobis, A. A report on the conservation status of segetal weeds in Tajikistan. Weed Res. 2014, 54, 635–648. [Google Scholar] [CrossRef]
- Albrecht, H.; Cambecèdes, J.; Lang, M.; Wagner, M. Management options for the conservation of rare arable plants in Europe. Bot Lett. 2016, 163, 389–415. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Scoppola, A.; Lososová, Z.; Abbate, G. Segetal plant communities of traditional agroecosystems: A phytosociological survey in central Italy. Phytocoenologia 2019, 49, 165–183. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Latini, M.; Iberite, M.; Abbate, G. Segetal flora of Italy. Version 1.5. Laboratory of Systematic Botany and Floristics—Department of Environmental Biology, Sapienza University, 2020. Occurrence dataset. Available online: https://doi.org/10.15468/44yxfz (accessed on 30 April 2020).
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Costantini, E.A.C.; Barbetti, R.; Fantappiè, M.; L’Abate, G.; Lorenzetti, R.; Magini, S. Pedodiversity. In The Soils of Italy; Dazzi, C., Ed.; Springer: Berlin, Germany, 2013. [Google Scholar]
- Chytrý, M.; Otýpková, Z. Plot sizes used for phytosociological sampling of European vegetation. J. Veg. Sci. 2003, 14, 563–570. [Google Scholar] [CrossRef]
- Güler, B.; Jentsch, A.; Apostolova, I.; Bartha, S.; Bloor, J.M.G.; Campetella, G.; Canullo, R.; Házi, J.; Kreyling, J.; Poittier, J.; et al. How plot shape and spatial arrange-affect plant species richness counts: Implications for sampling design and rarefaction analyses. J. Veg. Sci. 2016, 27, 692–703. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie; Springer: Vienna, Austria, 1964. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole-New Business Media: Bologna, Italy, 2017; Volume 2, pp. 1065–2260. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Galasso, G.; Bartolucci, F.; Peruzzi, L.; Ardenghi, N.M.G.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; Bandini Mazzanti, M.; Barberis, G.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Fried, G.; Petit, S.; Reboud, X. A specialist-generalist classification of the arable flora and its response to changes in agricultural practices. BMC Ecol. 2010, 10, 20. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Bioindicazione attraverso le piante vascolari. Valori di indicazione secondo Ellenberg (zeigerwerte) per le specie della Flora d’Italia. Braun-Blanquetia 2005, 39, 97. [Google Scholar]
- Griffith, M.D.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 12 May 2020).
- Veech, J.A. A Probabilistic Model for Analysing Species Co-Occurrence: Probabilistic Model. Global Ecol. Biogeogr. 2013, 22, 252–260. [Google Scholar] [CrossRef]
- Isbell, F.; Reich, P.B.; Tilman, D.; Hobbie, S.E.; Polasky, S.; Binder, S. Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. PNAS 2013, 110, 11911–11916. [Google Scholar] [CrossRef] [PubMed]
- Chytrý, M. Phytosociological data give biased estimates of species richness. J. Veg. Sci. 2001, 12, 439–444. [Google Scholar] [CrossRef]
- Perrino, E.V.; Calabrese, G. Endangered segetal species in southern Italy: Distribution, conservation status, trends, actions and ethnobotanical notes. Genet. Resour. Crop. Evol. 2018, 65, 2107–2134. [Google Scholar] [CrossRef]
- Condit, R.S.; Ashton, P.S.; Baker, P.J.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, Z.; Ye, W.; Cao, H.; Wei, S.; Wang, Z.; Lian, J.; Sun, I.-F.; Ma, K.; He, F. Spatial distributions of tree species in a subtropical forest of China. Oikos 2009, 118, 495–502. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Rare and Threatened Taxa | Positively Associated Taxa | Negatively Associated Taxa |
|---|---|---|
| Adonis annua | Aphanes arvensis; Bifora testiculata; Bupleurum subovatum; Coronilla scorpioides; Cota altissima; Delphinium consolida; Euphorbia falcata; Galium tricornutum; Geranium dissectum; Gladiolus italicus; Legousia speculum-veneris; Medicago polymorpha; Muscari comosum; Ranunculus arvensis; Scandix pecten-veneris; Vicia sativa agg.; Vicia lutea | Poa annua*; Polygonum aviculare agg.; Sonchus oleraceus* |
| Agrostemma githago | Galium tricornutum | Helminthotheca echioides; Lysimachia arvensis |
| Bifora radians | Galium tricornutum | |
| Centaurea cyanus | Veronica arvensis; Viola arvensis | Veronica persica |
| Filago pyramidata | Legousia speculum-veneris | |
| Galium tricornutum | Adonis annua; Agrostemma githago; Bifora radians; Bifora testiculata; Buglossoides arvensis; Bupleurum subovatum; Carduus pycnocephalus; Convolvulus arvensis; Coronilla scorpioides; Euphorbia falcata; Gladiolus italicus; Lathyrus aphaca; Lathyrus ochrus; Legousia speculum-veneris; Medicago arabica; Medicago polymorpha; Muscari comosum; Ranunculus arvensis; Rhagadiolus stellatus; Scandix pecten-veneris; Sonchus asper*; Torilis nodosa; Trigonella sulcata; Valerianella eriocarpa; Vicia sativa agg.; Vicia lutea | Epilobium tetragonum; Poa annua*; Polygonum aviculare agg.; Ranunculus sardous; Veronica persica |
| Lathyrus aphaca | Alopecurus myosuroides*; Galium tricornutum; Legousia speculum-veneris; Ranunculus arvensis; Scandix pecten-veneris | |
| Legousia speculum-veneris | Adonis annua; Anthemis cotula; Avena sterilis; Bifora testiculata; Bupleurum subovatum; Chondrilla juncea; Convolvulus arvensis; Coronilla scorpioides; Cota altissima; Crepis sancta; Delphinium consolida; Euphorbia falcata; Fallopia convolvulus; Filago pyramidata; Galium tricornutum; Gladiolus italicus; Glebionis segetum; Lathyrus aphaca; Lathyrus ochrus; Medicago polymorpha; Muscari comosum; Myosotis arvensis; Ranunculus arvensis; Rhagadiolus stellatus; Scandix pecten-veneris; Stachys annua; Tordylium apulum; Trigonella sulcata; Vicia lutea; Viola arvensis | Poa annua* |
| Lolium temulentum | Scandix pecten-veneris | |
| Ranunculus arvensis | Adonis annua; Anthemis cotula; Bifora testiculata; Bupleurum subovatum; Convolvulus arvensis; Coronilla scorpioides; Delphinium consolida; Euphorbia falcata; Fallopia convolvulus; Galium tricornutum; Helminthotheca echioides; Lathyrus aphaca; Lathyrus ochrus; Legousia speculum-veneris; Medicago polymorpha; Rhagadiolus stellatus; Scandix pecten-veneris; Stachys annua; Trigonella sulcata; Valerianella eriocarpa; Vicia sativa agg.; Vicia lutea | Glebionis segetum; Lolium multiflorum; Poa annua*; Viola arvensis |
| Scandix pecten-veneris | Adonis annua; Aphanes arvensis; Bifora testiculata; Buglossoides arvensis; Bupleurum subovatum; Coronilla scorpioides; Delphinium consolida; Euphorbia falcata; Galium tricornutum; Gladiolus italicus; Medicago arabica; Medicago polymorpha; Muscari comosum; Lathyrus aphaca; Lathyrus ochrus; Legousia speculum-veneris; Lolium temulentum; Ranunculus arvensis; Rhagadiolus stellatus; Sherardia arvensis; Trigonella sulcata; Vicia lutea; Vicia sativa agg. | Chenopodium album*; Matricaria chamomilla; Polygonum aviculare agg.; Veronica persica |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanfarillo, E.; Latini, M.; Abbate, G. Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy. Diversity 2020, 12, 195. https://doi.org/10.3390/d12050195
Fanfarillo E, Latini M, Abbate G. Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy. Diversity. 2020; 12(5):195. https://doi.org/10.3390/d12050195
Chicago/Turabian StyleFanfarillo, Emanuele, Marta Latini, and Giovanna Abbate. 2020. "Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy" Diversity 12, no. 5: 195. https://doi.org/10.3390/d12050195
APA StyleFanfarillo, E., Latini, M., & Abbate, G. (2020). Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy. Diversity, 12(5), 195. https://doi.org/10.3390/d12050195