Longitude, Forest Fragmentation, and Plant Size Influence Cycas micronesica Mortality Following Island Insect Invasions



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
3. Results
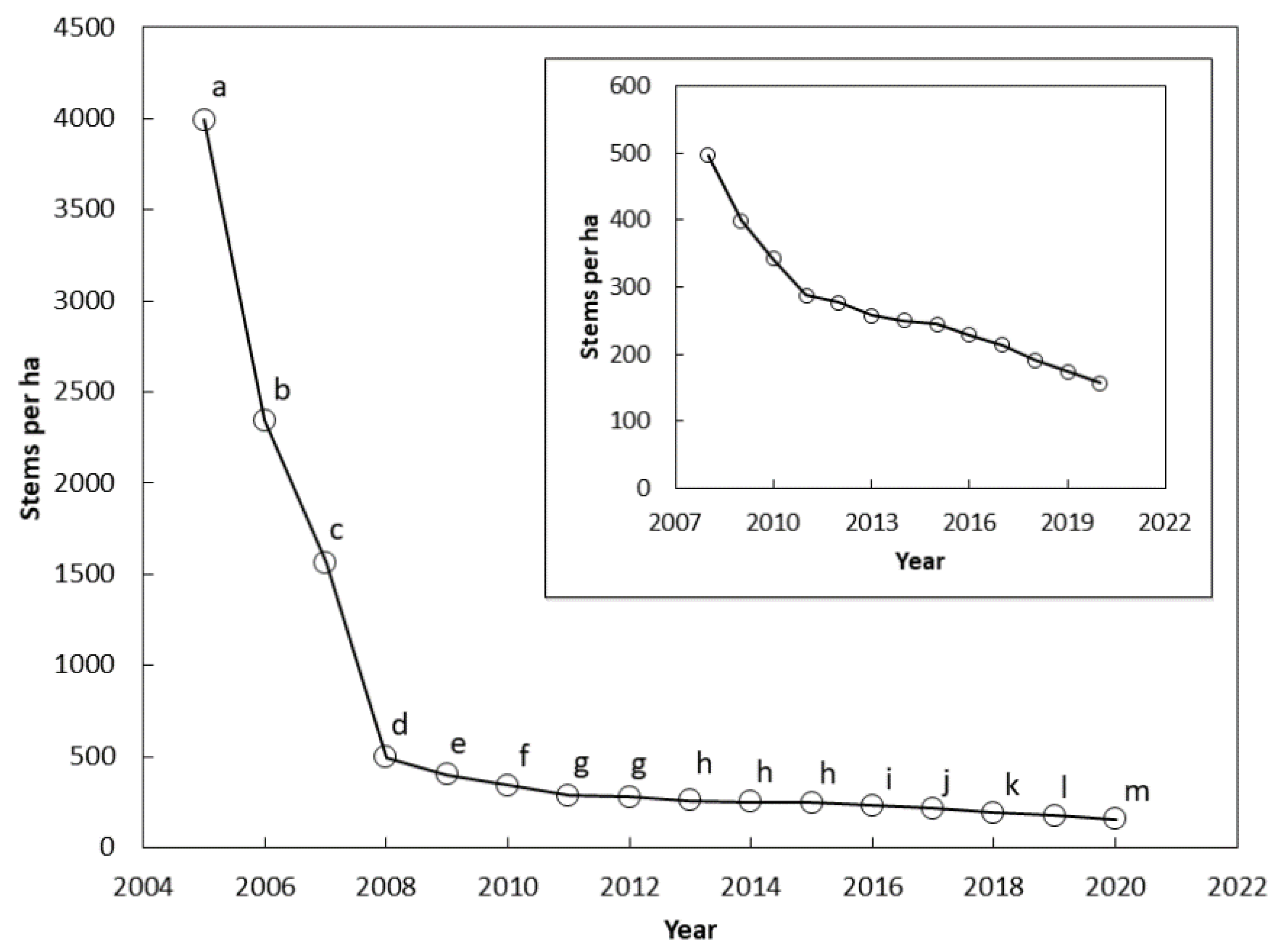
3.1. Guam Survival
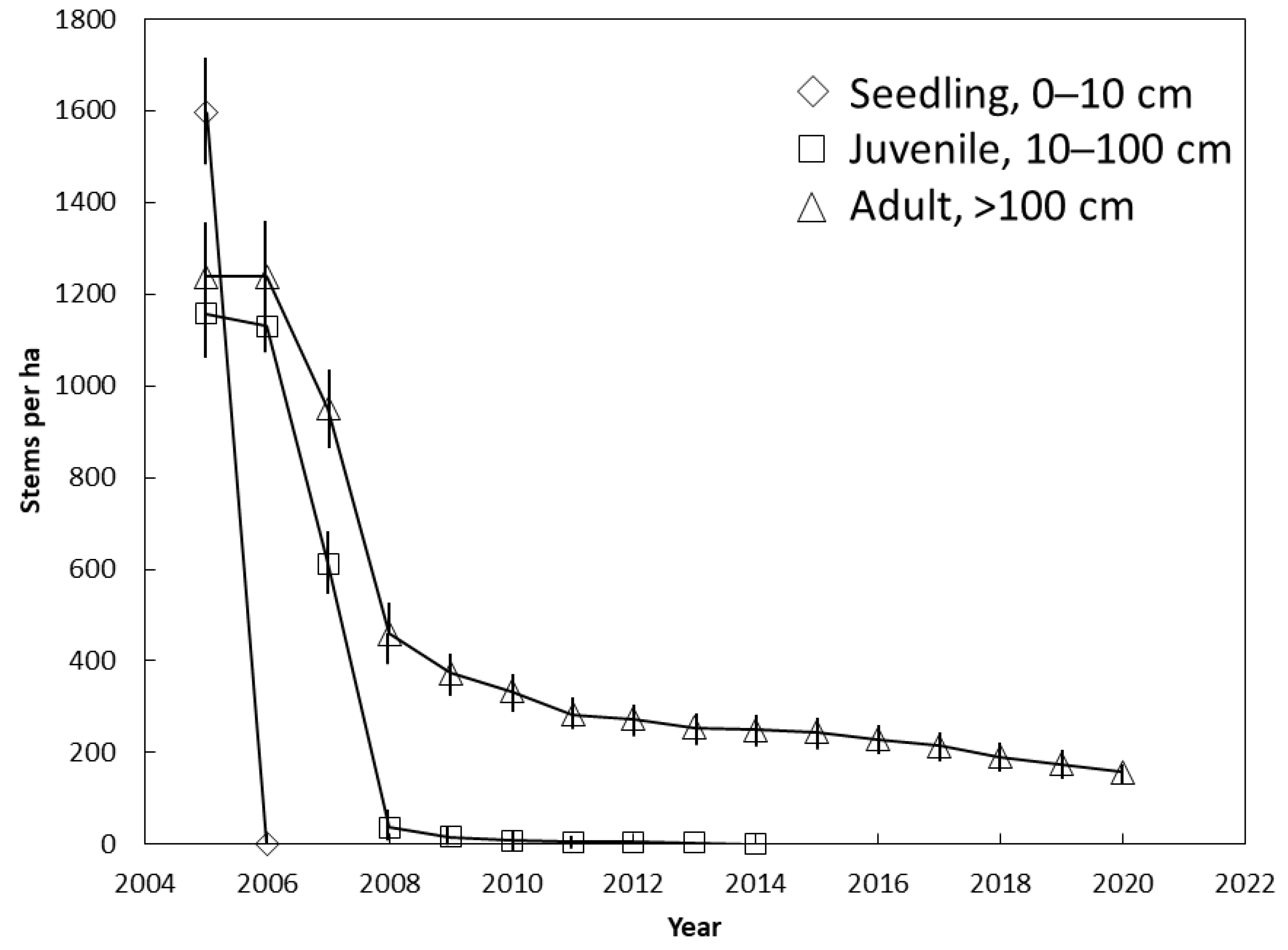
3.2. Guam Demography
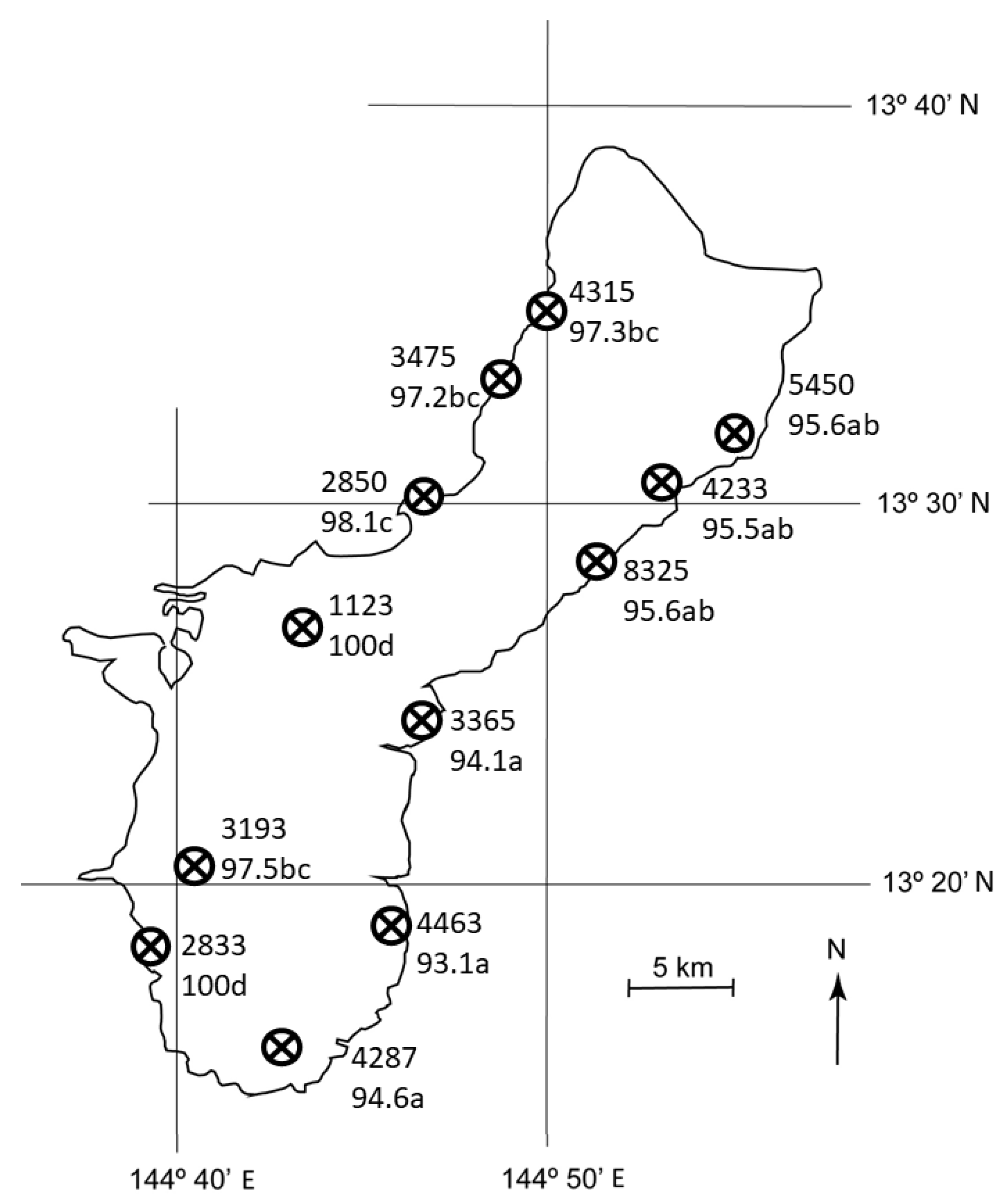
3.3. Guam Spatial Relations
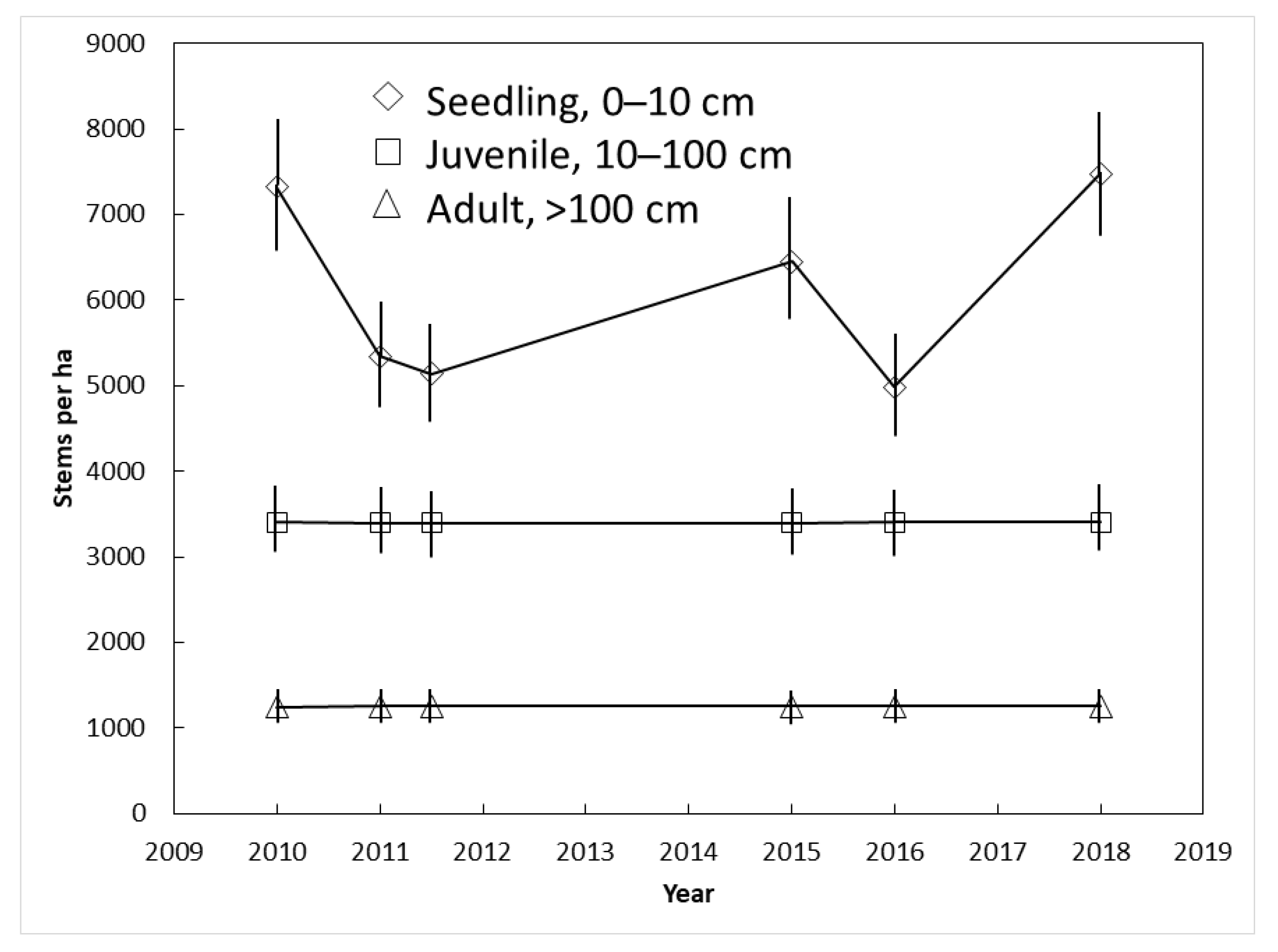
3.4. Yap Demography
4. Discussion
4.1. The Islands
4.2. Longitude and Fragmentation
4.3. Bigger Is Definitely Better
4.4. Looking Forward
4.4.1. Prognosis
4.4.2. Future Directions
4.5. Benchmarks
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McGeoch, M.A.; Butchart, S.H.M.; Spear, D.; Marais, E.; Kleynhans, E.J.; Symes, A.; Chanson, J.; Hoffmann, M. Global indicators of biological invasion: Species numbers, biodiversity impact and policy responses. Divers. Distrib. 2010, 16, 95–108. [Google Scholar] [CrossRef]
- Simberloff, D. How common are invasion-induced ecosystem impacts? Biol. Invasions 2011, 13, 1255–1268. [Google Scholar] [CrossRef]
- Sousa, R.; Morais, P.; Dias, E.; Antunes, C. Biological invasions and ecosystem functioning: Time to merge. Biol. Invasions 2011, 13, 1055–1058. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R. Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Davidson, C.B.; Gottschalk, K.W.; Johnson, J.E. Tree mortality following defoliation by the European gypsy moth (Lymantria dispar L.) in the United States: A review. Science 1999, 45, 74–84. [Google Scholar]
- Krasny, M.E.; DiGregorio, L.M. Gap dynamics in Allegheny northern hardwood forests in the presence of beech bark disease and gypsy moth disturbances. Ecol. Manag. 2001, 144, 265–274. [Google Scholar] [CrossRef]
- Jedlicka, J.; Vandermeer, J.; Aviles-Vazquez, K.; Barros, O.; Perfecto, I. Gypsy moth defoliation of oak trees and a positive response of red maple and black cherry: An example of indirect interaction. Am. Midl. Nat. 2004, 152, 231–236. [Google Scholar] [CrossRef]
- Orwig, D.A.; Cobb, R.C.; D’Amato, A.W.; Kizlinski, M.L.; Foster, D.R. Multi-year ecosystem response to hemlock woolly adelgid infestation in southern New England. Can. J. Res. 2008, 38, 834–843. [Google Scholar] [CrossRef]
- Kenis, M.; Auger-Rozenberg, M.-A.; Roques, A.; Timms, L.; Péré, C.; Cock, J.J.W.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Liebhold, A.M. Ecology of forest insect invasions. Biol. Invasions 2017, 19, 3141–3159. [Google Scholar] [CrossRef]
- Klooster, W.S.; Gandhi, K.J.K.; Long, L.C.; Perry, K.I.; Rice, K.B.; Herms, D.A. Ecological impacts of Emerald Ash Borer in forests at the epicenter of the invasion in North America. Forests 2018, 9, 250. [Google Scholar] [CrossRef]
- Kolka, R.K.; D’Amato, A.W.; Wagenbrenner, J.W.; Slesak, R.A.; Pypker, T.G.; Youngquist, M.B.; Grinde, A.R.; Palik, B.J. Review of ecosystem level impacts of Emerald Ash Borer on black ash wetlands: What does the future hold? Forests 2018, 9, 179. [Google Scholar] [CrossRef]
- Marler, T.E.; Muniappan, R. Pests of Cycas micronesica leaf, stem, and male reproductive tissues with notes on current threat status. Micronesica 2006, 39, 1–9. [Google Scholar]
- Marler, T.E. Cycad aulacaspis scale invades the Mariana Islands. Mem. N. Y. Bot. Gard. 2012, 106, 20–35. [Google Scholar]
- Marler, T.E. Temporal variations in leaf miner, butterfly, and stem borer infestations of Cycas micronesica in relation to Aulacaspis yasumatsui incidence. HortScience 2013, 48, 1334–1338. [Google Scholar] [CrossRef]
- Marler, T.E. Increased threat of island endemic tree’s extirpation via invasion-induced decline of intrinsic resistance to recurring tropical cyclones. Commun. Integr. Biol. 2013, 6, 22361. [Google Scholar] [CrossRef]
- Marler, T.E.; Lawrence, J.H.; Cruz, G.N. Topographic relief, wind direction, and conservation management decisions influence Cycas micronesica K.D. Hill population damage during tropical cyclone. J. Geogr. Nat. Disasters 2016, 6, 3. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N. Three invasive insects alter Cycas micronesica leaf chemistry and predict changes in biogeochemical cycling. Commun. Integr. Biol. 2016, 9, e1208324. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N. Do phytotoxic compounds in soil after scale-infested Cycas micronesica litter deposits explain reduced plant growth? HortScience 2013, 48, 1571–1573. [Google Scholar] [CrossRef]
- Watson, G.W.; Marler, T.E. Does cycad aulacaspis scale (Aulacaspis yasumatsui, Hemiptera: Diaspididae) play a direct role in causing soil phytotoxicity? Commun. Integr. Biol. 2014, 7, e27881. [Google Scholar] [CrossRef]
- Marler, T.E.; Cascasan, A.N.J. Carbohydrate depletion during lethal infestation of Aulacaspis yasumatsui on Cycas revoluta. Inter. J. Plant Sci. 2018, 179, 497–504. [Google Scholar] [CrossRef]
- Marler, T.E. Stem CO2 efflux of Cycas micronesica is reduced by chronic non-native insect herbivory. Plant Signal. Behav. 2020, 15, e1716160-2. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Lawrence, J.H. Demography of Cycas micronesica on Guam following introduction of the armoured scale Aulacaspis yasumatsui. J. Trop. Ecol. 2012, 28, 233–242. [Google Scholar] [CrossRef]
- Marler, T.E.; Lawrence, J.H. Canopy and knowledge gaps when invasive alien insects remove foundation species. Commun. Integr. Biol. 2013, 6, e22331. [Google Scholar] [CrossRef]
- Dueñas, M.-A.; Ruffhead, H.J.; Wakefield, N.H.; Roberts, P.D.; Hemming, D.J.; Diaz-Soltero, H. The role played by invasive species in interactions with endangered and threatened species in the United States: A systematic review. Biodiv. Conserv. 2018, 27, 3171–3183. [Google Scholar] [CrossRef]
- Marler, T.; Haynes, J.; Lindström, A. Cycas micronesica. IUCN 2012. IUCN Red List of Threatened Species. e.T61316A12462113. Available online: www.iucnredlist.org (accessed on 21 April 2020).
- United States Fish & Wildlife Service. Endangered and threatened wildlife and plants; endangered status for 16 species and threatened status for 7 species in Micronesia. Fed. Regist. 2015, 80, 59424–59497. [Google Scholar]
- Smith, C.W. Soil Survey of Islands of Yap, Federated States of Micronesia; USDA Soil Conservation Service: Washington, DC, USA, 1983. [Google Scholar]
- Young, F.J. Soil Survey of Territory of Guam; USDA: Washington, DC, USA, 1988; pp. 1–177. [Google Scholar]
- Moore, A.; Marler, T.; Miller, R.H.; Muniappan, R. Biological control of cycad aulacaspis scale on Guam. Cycad Newsl. 2005, 28, 6–8. [Google Scholar]
- Heu, R.A.; Chun, M.; Nagamine, W.T. Sago palm Scale, Aulacaspis yasumatsui Takagi (Homoptera: Diaspididae); State of Hawaii Department of Agriculture New Pest Advisory No. 99-01, Division of Plant Industry, Hawaii Department of Agriculture: Honolulu, HU, USA, 2003. [Google Scholar]
- Norstog, K.J.; Nicholls, T.J. The Biology of the Cycads; Cornell University Press: Ithaca, NY, USA, 1997. [Google Scholar]
- Marler, T.E. Vertical stratification in arthropod distribution research. Commun. Integr. Biol. 2013, 6, e25749. [Google Scholar] [CrossRef]
- Marler, T.E.; Miller, R.; Moore, A. Vertical stratification of predation on Aulacaspis yasumatsui infesting Cycas micronesica seedlings. HortScience 2013, 48, 60–62. [Google Scholar] [CrossRef]
- Marler, T.E.; Marler, P.N. Rhyzobius lophanthae behavior is influenced by cycad plant age providing odor samples in Y-tube olfactometer. Insects 2018, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Cibrián-Jaramillo, A.; Daly, A.C.; Brenner, E.; DeSalle, R.; Marler, T.E. When North and South don’t mix: Genetic connectivity of a recently endangered oceanic cycad, Cycas micronesica, in Guam using EST-microsatellites. Mol. Ecol. 2010, 19, 2364–2379. [Google Scholar] [CrossRef] [PubMed]
- Volis, S.; Deng, T. Importance of a single population demographic census as a first step of threatened species conservation planning. Biodiv. Conserv. 2020, 29, 527–543. [Google Scholar] [CrossRef]
- Marler, T.E.; Lee, V.; Shaw, C.A. Spatial variation of steryl glucosides in Cycas micronesica plants: Within- and among-plant sampling procedures. HortScience 2005, 40, 1607–1611. [Google Scholar] [CrossRef]
- Niklas, K.J.; Marler, T.E. Sex and population differences in the allometry of an endangered cycad species, Cycas micronesica (Cycadales). Int. J. Plant Sci. 2008, 169, 659–665. [Google Scholar] [CrossRef]
- Marler, T.E.; Niklas, K. Reproductive effort and success of Cycas micronesica K.D. Hill are affected by habitat. Int. J. Plant Sci. 2011, 172, 700–706. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N. Models to describe Cycas micronesica leaf and strobili development. HortScience 2011, 46, 1333–1337. [Google Scholar] [CrossRef]
- Hamada, T.; Terry, I.; Roemer, R.; Marler, T.E. Potential drift of pollen of Cycas micronesica on the island of Guam: A comparative study. HortScience 2015, 50, 1106–1117. [Google Scholar] [CrossRef]
- Hamada, T.; Terry, L.I.; Marler, T.E. Habitats, trade winds, and pollination of the endangered Cycas micronesica: Is there a role for wind as pollen vector on the island of Guam? Int. J. Plant Sci. 2015, 176, 525–543. [Google Scholar] [CrossRef]
- Dongol, N.; Marler, T.E. Season and frequency of Cycas micronesica leaf and reproductive events. Mem. N. Y. Bot. Gard. 2018, 117, 497–503. [Google Scholar]
- Lefkovitch, L.P. The study of population growth in organisms grouped by stages. Biometrika 1965, 35, 183–212. [Google Scholar] [CrossRef]
- Leslie, P.H. The use of matrices in certain population mathematics. Biometrika 1945, 33, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In Dynamics of Populations; Den Boer, P.J., Gradwell, G.R., Eds.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1971; pp. 298–312. [Google Scholar]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–529. [Google Scholar] [CrossRef]
- Marler, T.E.; Lindström, A.J. First, do no harm. Commun. Integr. Biol. 2017, 10, e1393593. [Google Scholar] [CrossRef]
- Keehley, P.; Abercrombie, N.N. Benchmarking in the Public and Nonprofit Sectors, 2nd ed.; Wiley: San Francisco, CA, USA, 2008. [Google Scholar]
- Sutherland, W.J.; Peel, M.J.S. Benchmarking as a means to improve conservation practice. Oryx 2010, 45, 56–59. [Google Scholar] [CrossRef][Green Version]
- Janzen, D.H. The uncertain future of the tropics. Nat. Hist. 1972, 81, 80–89. [Google Scholar]
- Janzen, D.H. The future of tropical ecology. Ann. Rev. Ecol. Syst. 1986, 17, 305–324. [Google Scholar] [CrossRef]
- Janzen, D.H. Tropical ecological and biocultural restoration. Science 1988, 239, 243–244. [Google Scholar] [CrossRef]
- Janzen, D.H. Tropical dry forests. In Biodiversity; Wilson, E.O., Peter, F.M., Eds.; National Academy Press: Washington, DC, USA, 1988; pp. 130–137. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marler, T.E.; Krishnapillai, M.V. Longitude, Forest Fragmentation, and Plant Size Influence Cycas micronesica Mortality Following Island Insect Invasions. Diversity 2020, 12, 194. https://doi.org/10.3390/d12050194
Marler TE, Krishnapillai MV. Longitude, Forest Fragmentation, and Plant Size Influence Cycas micronesica Mortality Following Island Insect Invasions. Diversity. 2020; 12(5):194. https://doi.org/10.3390/d12050194
Chicago/Turabian StyleMarler, Thomas E., and Murukesan V. Krishnapillai. 2020. "Longitude, Forest Fragmentation, and Plant Size Influence Cycas micronesica Mortality Following Island Insect Invasions" Diversity 12, no. 5: 194. https://doi.org/10.3390/d12050194
APA StyleMarler, T. E., & Krishnapillai, M. V. (2020). Longitude, Forest Fragmentation, and Plant Size Influence Cycas micronesica Mortality Following Island Insect Invasions. Diversity, 12(5), 194. https://doi.org/10.3390/d12050194