The Leafless Vanilla Species-Complex from the South-West Indian Ocean Region: A Taxonomic Puzzle and a Model for Orchid Evolution and Conservation Research

 ,
,  ,
,
Abstract

1. Introduction
2. The Vanilla Genus and Leafless Species
2.1. Botany
2.2. Taxonomy
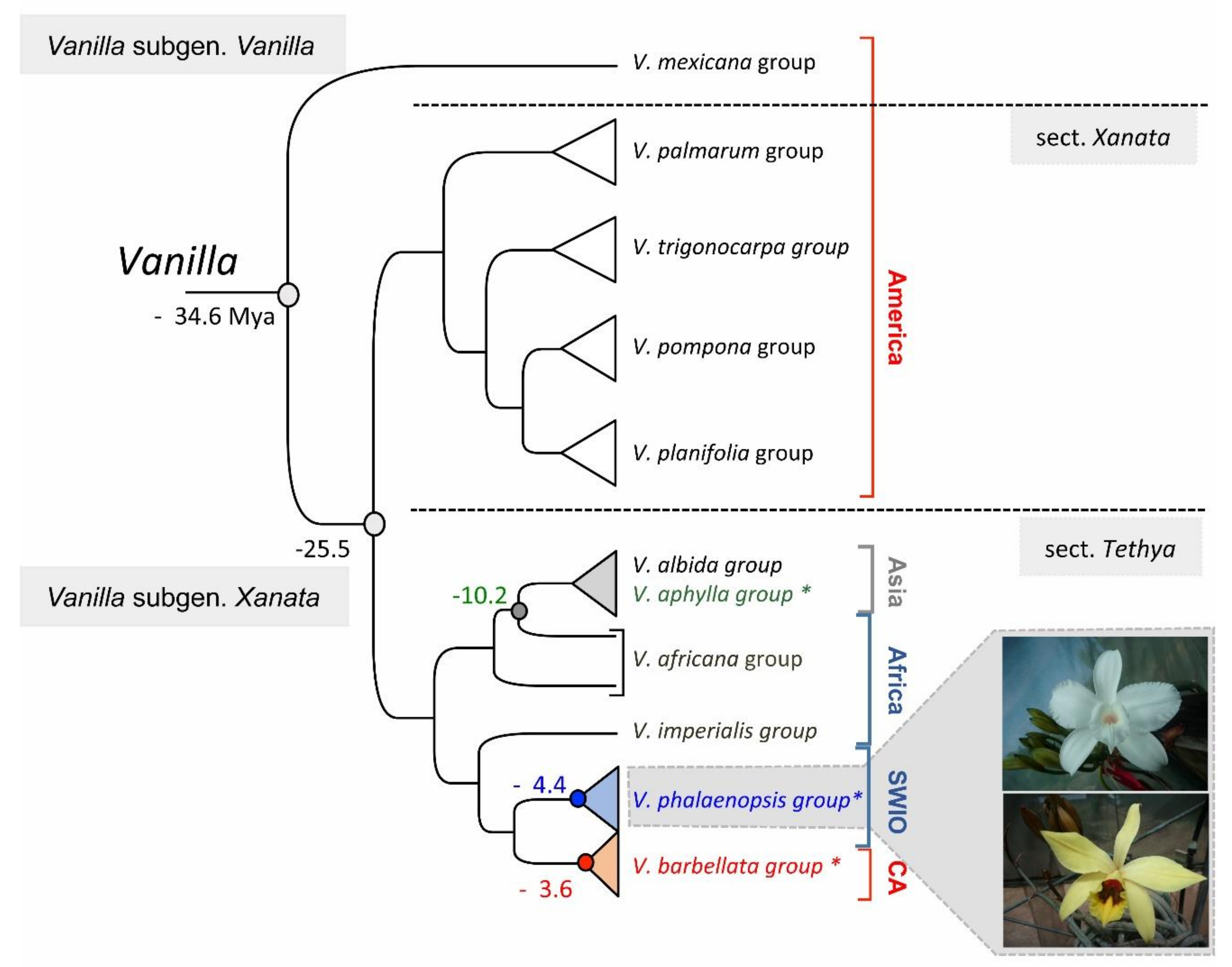
2.3. Evolution
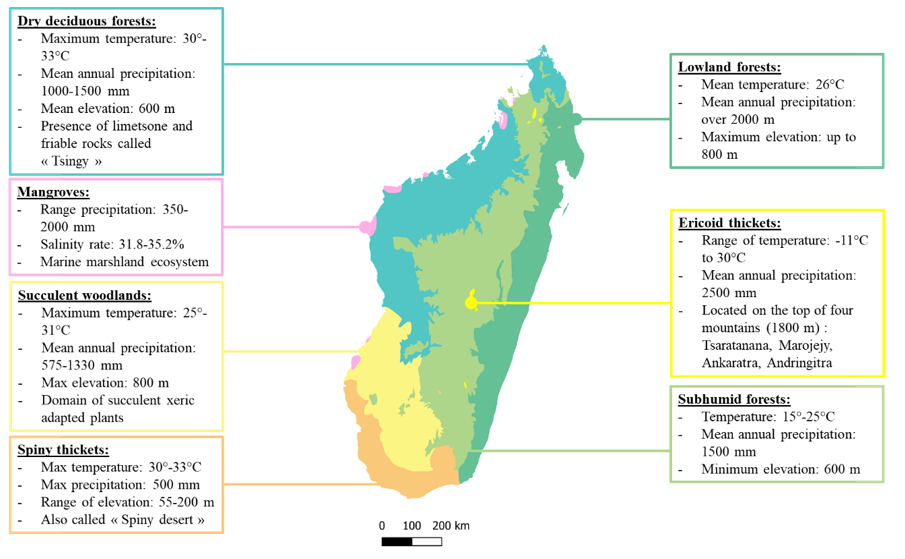
2.4. Ecology
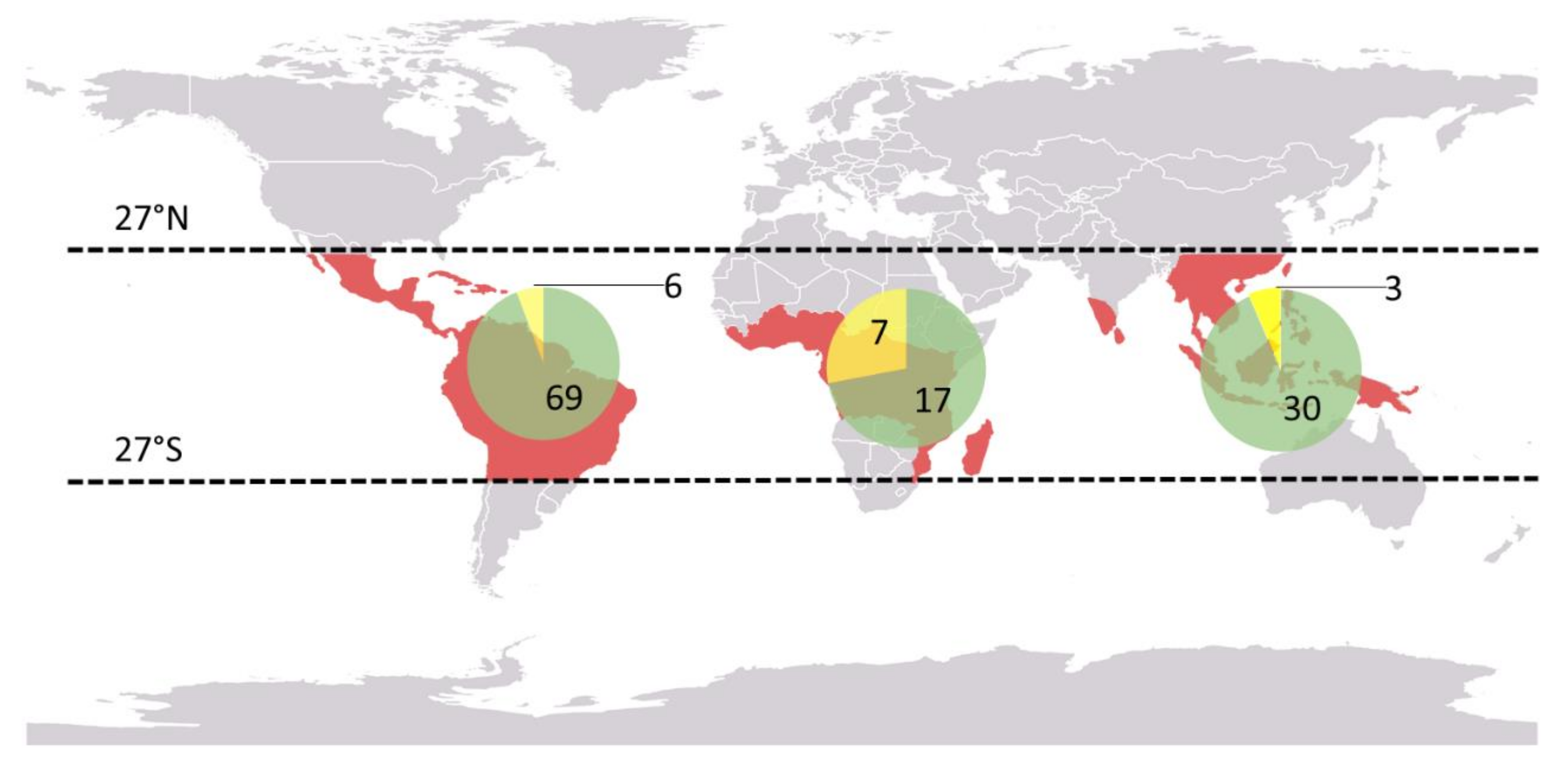
2.5. Species Diversity and Distribution
2.6. Reproduction Biology
2.7. Vanilla: A Taxonomic Complex Group
2.8. Ethnobotanic and Commercial Importance
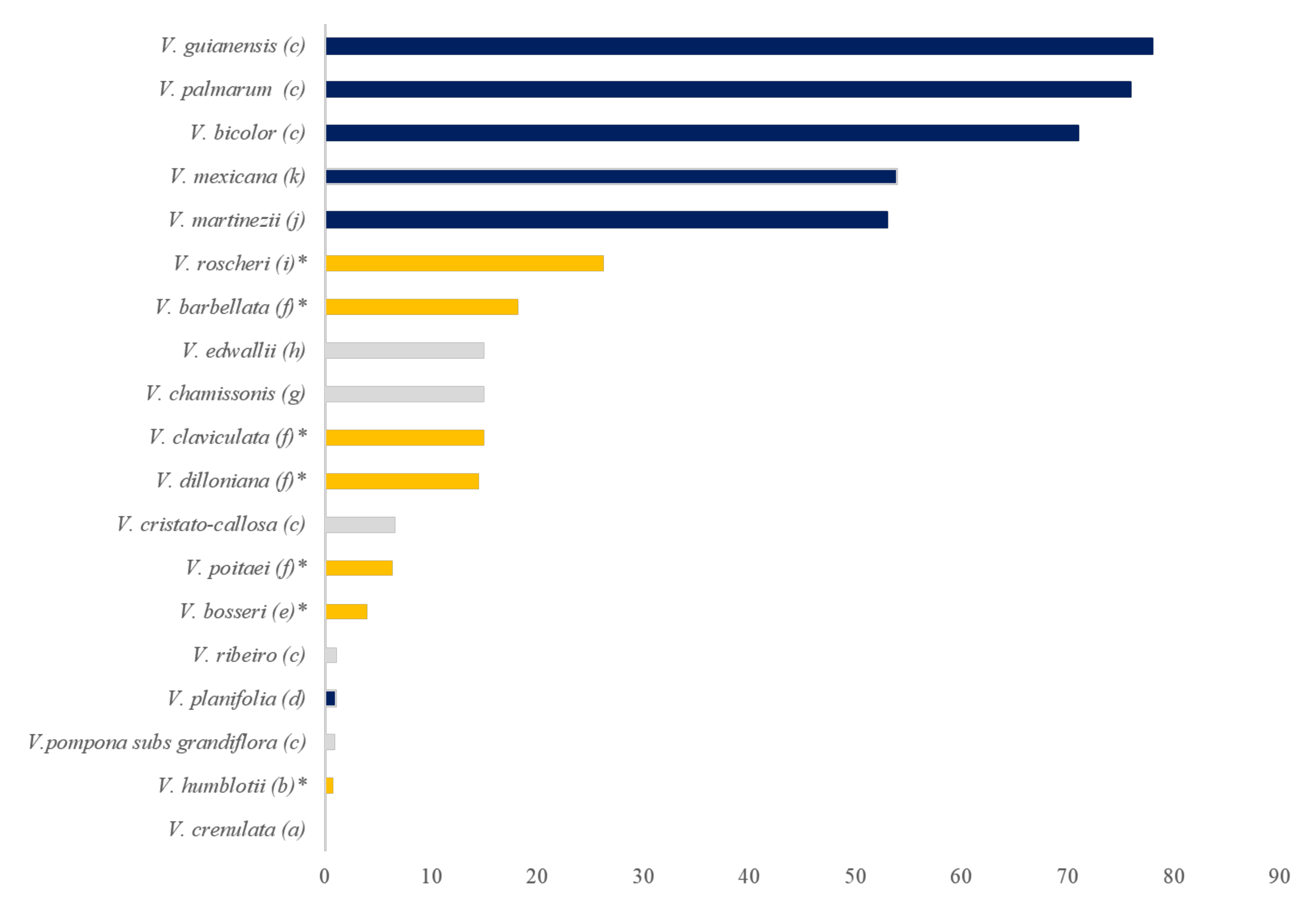
2.9. Conservation Issues
3. The Leafless Vanilla Clade from the SWIO Region: A Taxonomic Puzzle
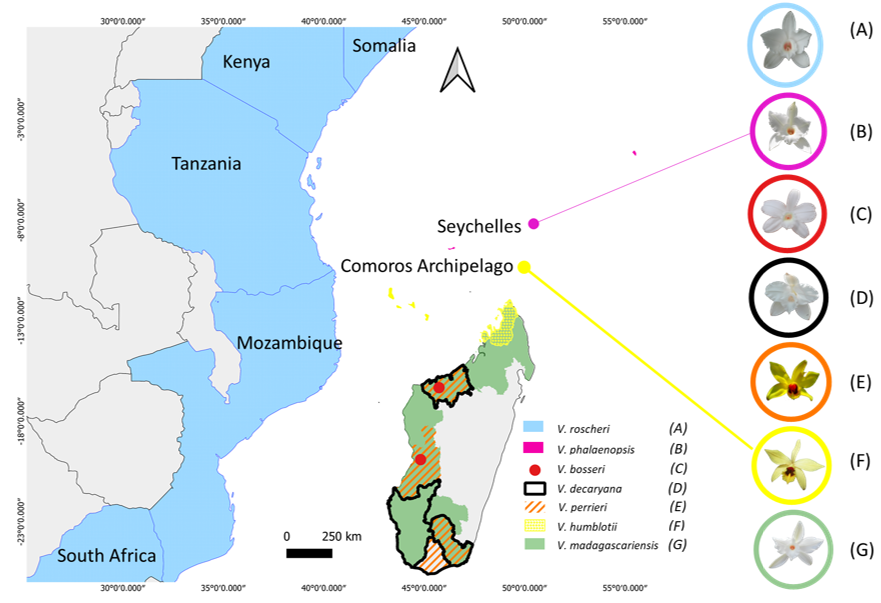
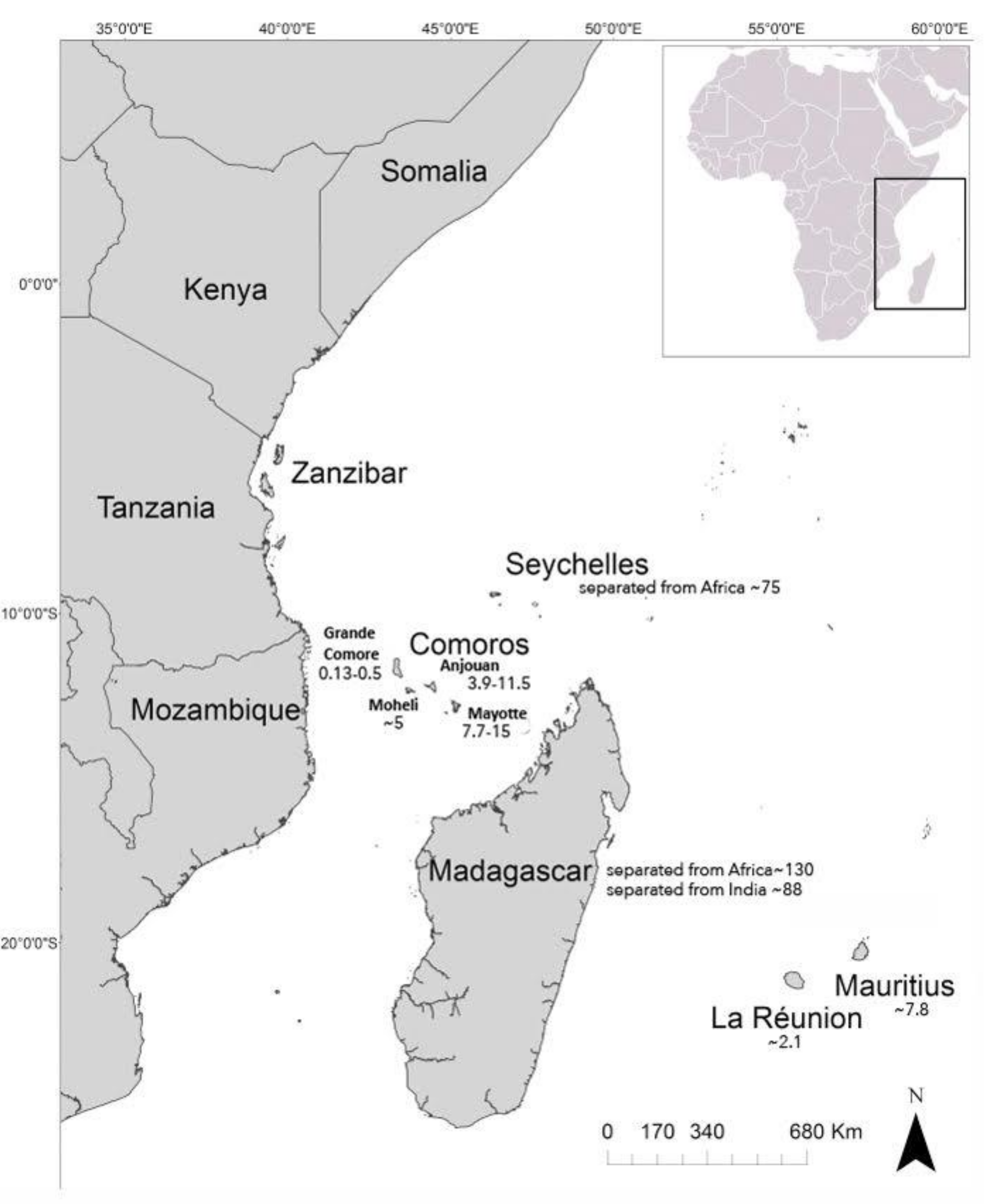
3.1. Distribution
3.2. Botanical Description
3.3. A Taxonomic Puzzle
3.4. Conclusion and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Fay, M.F.; Chase, M.W. Orchid biology: From Linnaeus via Darwin to the 21st century. Ann. Bot. 2009, 104, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Bouetard, A.; Lefeuvre, P.; Gigant, R.L.; Bory, S.; Pignal, M.; Besse, P.; Grisoni, M. Evidence of transoceanic dispersion of the genus Vanilla based on plastid DNA phylogenetic analysis. Mol. Phylogenetics Evol. 2010, 55, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, S.R.; Gravendeel, B.; Singer, R.B.; Marshall, C.R.; Pierce, N.E. Dating the origin of the Orchidaceae from a fossil orchid with its pollinator. Nature 2007, 448, 1042–1045. [Google Scholar] [CrossRef] [PubMed]
- Chinsamy, M.; Finnie, J.F.; Van Staden, J. The ethnobotany of South African medicinal orchids. S. Afr. J. Bot. 2011, 77, 2–9. [Google Scholar] [CrossRef]
- Pailler, T.; Andilyat, M.; Andrianarivo, C.; Baider, C.; Bytebier, B.; Filer, D.; Henze, F.; Morel, C. Guide des Orchidées des Îles de l’Océan Indien: Espèces Indigènes et Endémiques, 1st ed.; Commission de l’Océan Indien: La Réunion, France, 2018; pp. 8–163. [Google Scholar]
- Callmander, M.W.; Phillipson, P.B.; Schatz, G.E.; Andriambololonera, S.; Rabarimanarivo, M.; Rakotonirina, N.; Raharimampionona, J.; Chatelain, C.; Gautier, L.; Lowry, P.P. The endemic and non-endemic vascular flora of Madagascar updated. Plant Ecol. Evol. 2011, 144, 121–125. [Google Scholar] [CrossRef]
- Cribb, P.; Hermans, J. Field Guide to the Orchids of Madagascar; Royal Botanical Gardens: Kew, UK, 2009; ISBN 978-1-84246-158-7. [Google Scholar]
- Soto Arenas, M.Á.; Cribb, P. A new infrageneric classification and synopsis of the genus Vanilla Plum. Ex Mill. (Orchidaceae: Vanillinae). Lankesteriana 2010, 9, 355–398. [Google Scholar] [CrossRef]
- Portères, R. Le Genre Vanilla et ses Espèces. In Le Vanillier et la Vanille dans le Monde; Bouriquet, G., Ed.; Editions P. Lechevalier: Paris, France, 1954; pp. 94–920. [Google Scholar]
- De La Cruz Medina, J.; Rodriguez Jiménes, G.C.; Garcia, H.S.; Zarrabal, T.L.R.; Alvarado, M.A.G.; José, V. Vanilla: Post-Harvest Operations; Food and Agriculture Organization of United Nations: Veracruz, Mexico, 2009; pp. 1–25. [Google Scholar]
- Bosser, J.; Lecoufle, M. Les Orchidées de Madagascar; Parthénope; Biotope: Mèze, France, 2011; ISBN 2-36662-047-0. [Google Scholar]
- Descouvrières, P. Encyclopédie Des Orchidées Tropicales—1200 Espèces, Plus De 1000 Photographies|Rakuten; Ulmer: Paris, France, 2011; Volume 1, ISBN 978-2-84138-472-3. [Google Scholar]
- Soto Arenas, M.A.; Cameron, K.M. Vanilla. In Genera Orchidacearum: Orchidoideae; Pridgeon, A.M., Cribb, P., Chase, M.W., Rasmussen, F.N., Eds.; Oxford University Press: Oxford, UK, 2003; pp. 321–334. [Google Scholar]
- Botomanga, A. Stratégies D’adaptation à la Sècheresse et à la Dégradation de L’habitat des Racines de Vanilliers Aphylles. Master’s Thesis, Université d’Antananarivo, Antananarivo, Madagascar, 2018. [Google Scholar]
- Cameron, K.M. Vanilloid orchids systematics and evolution. In Vanilla; Odoux, E., Grisoni, M., Eds.; Medicinal and Aromatic Plants—Industrial Profiles; CRC Press: Boca Raton, FL, USA, 2011; Volume 47, pp. 1–12. ISBN 978-1-4200-8338-5. [Google Scholar]
- Griffiths, H.; Males, J. Succulent plants. Curr. Biol. 2017, 27, R890–R896. [Google Scholar] [CrossRef]
- Gigant, R.L.; Bory, S.; Grisoni, M.; Besse, P. Biodiversity and evolution in the Vanilla genus. In The Dynamical Processes of Biodiversity: Case Studies of Evolution and Spatial Distribution; IntechOpen: Rijeka, Croatia, 2011; pp. 1–26. ISBN 978-953-307-772-7. [Google Scholar]
- Gigant, R.L.; De Bruyn, A.; Church, B.; Humeau, L.; Gauvin-Bialecki, A.; Pailler, T.; Grisoni, M.; Besse, P. Active sexual reproduction but no sign of genetic diversity in range-edge populations of Vanilla roscheri Rchb. f. (Orchidaceae) in South Africa. Conserv. Genet. 2014, 15, 1403–1415. [Google Scholar] [CrossRef]
- Soto Arenas, M.A.; Dressler, R.L. A revision of the Mexican and central American species of Vanilla Plumier ex Miller with a characterization of their ITS region of the nuclear ribosomal DNA. Lankesteriana 2010, 9, 285–354. [Google Scholar] [CrossRef]
- Gigant, R.L.; De Bruyn, A.; M’sa, T.; Viscardi, G.; Gigord, L.; Gauvin-Bialecki, A.; Pailler, T.; Humeau, L.; Grisoni, M.; Besse, P. Combining pollination ecology and fine-scale spatial genetic structure analysis to unravel the reproductive strategy of an insular threatened orchid. S. Afr. J. Bot. 2016, 105, 25–35. [Google Scholar] [CrossRef]
- Nielsen, L.R.; Siegismund, H.R. Interspecific differentiation and hybridization in Vanilla species (Orchidaceae). Heredity 1999, 83, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, R.A. A revision of the genus Vanilla. Bot. J. Linn. Soc. 1896, 32, 439–478. [Google Scholar] [CrossRef]
- Besse, P. Guidelines for the choice of sequences for molecular plant taxonomy. In Molecular Plant Taxonomy; Methods in Molecular Biology; Besse, P., Ed.; Humana Press: Totowa, NJ, USA, 2014; Volume 1115, pp. 39–51. ISBN 978-1-62703-766-2. [Google Scholar]
- Azofeifa-Bolaños, J.B.; Gigant, R.L.; Nicolás-García, M.; Pignal, M.; Tavares-González, F.B.; Hágsater, E.; Salazar-Chávez, G.A.; Reyes-López, D.; Archila-Morales, F.L.; García-García, J.A.; et al. A new Vanilla species from Costa Rica closely related to V. planifolia (Orchidaceae). EJT 2017, 284, 1–26. [Google Scholar] [CrossRef]
- Cameron, K.M. Recent advances in the systematic biology of Vanilla and related orchids (Orchidaceae: Subfamily Vanilloideae). In Proceedings of the Vanilla: First International Congress, Princeton, NJ, USA, 11–12 November 2003; Allured: Carol Stream, IL, USA, 2005; pp. 89–93. [Google Scholar]
- Govaerts, R.; Bernet, P.; Kratochvil, K.; Gerlach, G.; Carr, G.; Alrich, P.; Pridgeon, A.M.; Pfahl, J.; Campacci, M.A.; Holland Baptista, D.; et al. World Checklist of Orchidaceae. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://wcsp.science.kew.org/ (accessed on 17 July 2020).
- IPNI International Plant Names Index. Available online: http://www.ipni.org (accessed on 17 July 2020).
- MBG Tropicos: Connecting the World to Botanical Data since 1982. Available online: https://www.tropicos.org/ (accessed on 17 July 2020).
- Pansarin, E.R.; Miranda, M.R. A new species of Vanilla (Orchidaceae: Vanilloideae) from Brazil. Phytotaxa 2016, 267, 84. [Google Scholar] [CrossRef]
- Barberena, F.F.V.A.; Sousa, T.D.S.; Ambrosio-Moreira, B.D.S.; Roque, N. What are the species of phorophytes of Vanilla palmarum (Orchidaceae) in Brazil? An assessment of emblematic specificity with palm tree species. Rodriguésia 2019, 70, e02732017. [Google Scholar] [CrossRef]
- Sambin, A.; Chiron, G.R. Deux Nouvelles Espèces de Vanilla (Orchidaceae) de Guyane Française. Richardiana 2015, XV, 306–316. [Google Scholar]
- Porras-Alfaro, A.; Bayman, P. Mycorrhizal fungi of Vanilla: Diversity, specificity and effects on seed germination and plant growth. Mycologia 2007, 99, 510–525. [Google Scholar] [CrossRef]
- Allorge-Boiteau, L. Les vanilles succulentes de Madagascar. Succulentes 2005, 2, 3–11. [Google Scholar]
- Allorge-Boiteau, L. Une nouvelle espèce de vanille à Madagascar. Hommes Et Plantes 2013, 85, 4–5. [Google Scholar]
- Soto Calvo, M.A.; Esperon, P.; Sauleda, R.P. A new species of Vanilla Miller is described for Cuba. New World Orchid Nomencl. Notes 2019, 56, 1–7. Available online: https://www.researchgate.net/profile/Ruben_Sauleda/publication/341056775_A_New_Species_of_Vanilla_Miller_is_Described_for_Cuba/links/5eab47c5299bf18b958a7273/A-New-Species-of-Vanilla-Miller-is-Described-for-Cuba.pdf (accessed on 20 July 2020).
- Bory, S.; Brown, S.; Duval, M.-F.; Besse, P. Evolutionary processes and diversification in the genus Vanilla. In Vanilla; Odoux, E., Grisoni, M., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 15–29. ISBN 978-1-4200-8338-5. [Google Scholar]
- Parizaca, A.D. Vanilla yanesha (Orchidaceae), a new species of the membranaceous-leaved group from the central rainforest of Peru. Willdenowia 2019, 49, 5. [Google Scholar] [CrossRef]
- Flanagan, N.S.; Ospina-Calderón, N.H.; Agapito, L.T.G.; Mendoza, M.; Mateus, H.A. A new species of Vanilla (Orchidaceae) from the North West Amazon in Colombia. Phytotaxa 2018, 364, 250. [Google Scholar] [CrossRef]
- Damian, A.; Mitidieri, N. Living in the clouds: A new high-elevation species of Vanilla (Orchidaceae, Vanilloideae) from Perú. Phytotaxa 2020, 451, 154–160. [Google Scholar] [CrossRef]
- Cameron, K.M.; Chase, M.W. Phylogenetic relationships of Pogoniinae (Vanilloideae, Orchidaceae): An herbaceous example of the Eastern North America-Eastern Asia phytogeographic disjunction. J. Plant Res. 1999, 112, 317–329. [Google Scholar] [CrossRef]
- Householder, E.; Janovec, J.; Mozambite, A.B.; Maceda, J.H.; Wells, J.; Valega, R. Diversity, natural history, and conservation of Vanilla (orchidaceae) in Amazonian wetlands of Madre De Dios, Peru. J. Bot. Res. Inst. Tex. 2010, 4, 227–243. [Google Scholar]
- Nielsen, L.R. Natural hybridization between Vanilla claviculata (W.Wright) Sw. and V. barbellata Rchb.f. (Orchidaceae): Genetic, morphological, and pollination experimental data. Bot. J. Linn. Soc. 2000, 133, 285–302. [Google Scholar] [CrossRef]
- Molineros Hurtado, F.; González Mina, R.T.; Flanagan, N.S.; Tupac Otero, J. Vanilla rivasii (orchidaceae), a new species from the Colombian pacific region. Lankesteriana 2014, 13. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: A spasmodic journey to diversification. Biol. J. Linn. Soc. 2005, 84, 1–54. [Google Scholar] [CrossRef]
- Gigant, R.L.; Rakotomanga, N.; Citadelle, G.; Silvestre, D.; Grisoni, M.; Besse, P. Microsatellite markers confirm self-pollination and autogamy in wild populations of Vanilla mexicana Mill. (syn. V. inodora) (Orchidaceae) in the island of Guadeloupe. In Microsatellite Markers; Abdurakhmonov, I., Ed.; IntechOpen: London, UK, 2016; pp. 73–93. ISBN 978-953-51-2797-0. [Google Scholar] [CrossRef]
- Micheneau, C.; Johnson, S.D.; Fay, M.F. Orchid pollination: From Darwin to the present day. Bot. J. Linn. Soc. 2009, 161, 1–19. [Google Scholar] [CrossRef]
- Peter, C.I.; Johnson, S.D. A pollinator shift explains floral divergence in an orchid species complex in South Africa. Ann. Bot. 2014, 113, 277–288. [Google Scholar] [CrossRef]
- Bembé, B. Functional morphology in male euglossine bees and their ability to spray fragrances (Hymenoptera, Apidae, Euglossini). Apidologie 2004, 35, 283–291. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, R.L. Trends in the pollination ecology of the Orchidaceae: Evolution and systematics. Can. J. Bot. 1992, 70, 642–650. [Google Scholar] [CrossRef]
- Pansarin, E.R.; Pansarin, L.M. Floral biology of two Vanilloideae (Orchidaceae) primarily adapted to pollination by euglossine bees. Plant Biol. J. 2014. [Google Scholar] [CrossRef]
- Petersson, L. Pollination Biology of the Endemic Orchid Vanilla bosseri in Madagascar. Master’s Thesis, Biology Education Center, Disciplinary Domain of Science and Technology, Uppsala University, Uppsala, Sweden, 2015. [Google Scholar]
- Ackerman, J.D.; Sabat, A.; Zimmerman, J.K. Seedling establishment in an epiphytic orchid: An experimental study of seed limitation. Oecologia 1996, 106, 192–198. [Google Scholar] [CrossRef]
- Macedo, R.C.A. Biologia Reprodutiva e Propagacao Vegetativa de Vanilla chamissonis Klotzsch: Subsidios para Manejo Sustentado. Master’s Thesis, Escola Superior de Agric Luiz de Queiroz, Piracicaba, Brazil, 2000. [Google Scholar]
- Lubinsky, P.; Van Dam, M.; Van Dam, A. Pollination of Vanilla and evolution in the Orchidaceae. Lindleyana 2006, 75, 926–929. [Google Scholar]
- Dressler, R.L. Phylogeny and Classification of the Orchid Family; Cambridge University Press: Cambridge, UK, 1993; ISBN 978-0-521-45058-4. [Google Scholar]
- Chaipanich, V.V.; Wanachantararak, P.; Hasin, S. Floral morphology and Potential Pollinator of Vanilla siamensis Rolfe ex Downie (Orchidaceae: Vanilloideae) in Thailand. Thail. Nat. Hist. Mus. J. 2020, 14, 1–14. [Google Scholar]
- Divakaran, M.; Babu, K.N.; Ravindran, P.N.; Peter, K.V. Interspecific hybridization in Vanilla and molecular characterization of hybrids and selfed progenies using RAPD and AFLP markers. Sci. Hortic. 2006, 108, 414–422. [Google Scholar] [CrossRef]
- Baack, E.; Melo, M.C.; Rieseberg, L.H.; Ortiz-Barrientos, D. The origins of reproductive isolation in plants. New Phytol. 2015, 207, 968–984. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Schlüter, P.M. Floral isolation, specialized pollination, and pollinator behavior in orchids. Annu. Rev. Entomol. 2009, 54, 425–446. [Google Scholar] [CrossRef]
- Scopece, G.; Widmer, A.; Cozzolino, S. Evolution of postzygotic reproductive isolation in a guild of deceptive orchids. Am. Nat. 2008, 171, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Schlüter, P.M.; Gross, K.; Schiestl, F.P. Floral isolation is the major reproductive barrier between a pair of rewarding orchid sister species. J. Evol. Biol. 2015, 28, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Minoo, D.; Jayakumar, V.N.; Veena, S.S.; Vimala, J.; Basha, A.; Saji, K.V.; Nirmal Babu, K.; Peter, K.V. Genetic variations and interrelationships in Vanilla planifolia and few related species as expressed by RAPD polymorphism. Genet. Resour. Crop. Evol. 2008, 55, 459–470. [Google Scholar] [CrossRef]
- Lubinsky, P.; Cameron, K.M.; Molina, M.C.; Wong, M.; Lepers-Andrzejewski, S.; Gomez-Pompa, A.; Kim, S.-C. Neotropical roots of a Polynesian spice: The hybrid origin of Tahitian Vanilla, Vanilla tahitensis (Orchidaceae). Am. J. Bot. 2008, 95, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Barona-Colmenares, A.A. Two new records in Orchidaceae (Vanillinae) from southernmost Colombian Amazonia: Vanilla javieri, a new species, and Vanilla appendiculata. Phytotaxa 2018, 375, 261. [Google Scholar] [CrossRef]
- Ennos, R.; French, G.; Hollingsworth, P. Conserving taxonomic complexity. Trends Ecol. Evol. 2005, 20, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Duval, M.-F.; Bory, S.; Andrzejewski, S.; Grisoni, M.; Besse, P.; Causse, S.; Charon, C.; Dron, M.; Odoux, E.; Wong, M. Diversité génétique des vanilliers dans leurs zones de dispersion secondaire. Les Actes du BRG 2006, 6, 181–196. [Google Scholar]
- Bory, S.; Catrice, O.; Brown, S.; Leitch, I.J.; Gigant, R.L.; Chiroleu, F.; Grisoni, M.; Duval, M.-F.; Besse, P. Natural polyploidy in Vanilla planifolia (Orchidaceae). Genome 2008, 51, 816–826. [Google Scholar] [CrossRef]
- Morales, F.A.; Menchaca, R.; Chiron, G.R. Notes on Mesoamerican orchids. II: Millenary use in the Q’eqchi communities of the lowlands, with a new Vanilla species. Richardiana 2019, 3, 100–108. [Google Scholar]
- Watteyn, C.; Fremout, T.; Karremans, A.P.; Huarcaya, R.P.; Azofeifa Bolaños, J.B.; Reubens, B.; Muys, B. Vanilla distribution modeling for conservation and sustainable cultivation in a joint land sparing/sharing concept. Ecosphere 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Fouché, J.G.; Jouve, L. Vanilla planifolia: History, botany and culture in Reunion island. Agronomie 1999, 19, 689–703. [Google Scholar] [CrossRef]
- Koyyappurath, S.; Conéjéro, G.; Dijoux, J.B.; Lapeyre-Montès, F.; Jade, K.; Chiroleu, F.; Gatineau, F.; Verdeil, J.L.; Besse, P.; Grisoni, M. Differential responses of Vanilla accessions to root rot and colonization by Fusarium oxysporum f. sp. radicis-vanillae. Front. Plant Sci. 2015, 6, 1125. [Google Scholar] [CrossRef] [PubMed]
- Bond, M.O.; Aregullin, M.A.; Laux, M.T. Antimicrobial, cytotoxic and antiproliferative properties of native and invasive orchids in the Dominican ethnobotany. Pennscience 2014, 12, 23–28. [Google Scholar]
- Hernandez-Hernandez, J. Vanilla diseases. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F.C., Eds.; Wiley-Blackwell: Chichester, UK, 2011; pp. 26–39. ISBN 978-1-4051-9325-2. [Google Scholar]
- Kannan, M.; Senthil Kumar, T.; Rao, M. Utilization of plant resources for non-medicinal purposes by Malayali tribes of Kalrayan hills of Salem District, Tamil Nadu, India. Int. J. Herb. Med. 2016, 4, 47–58. [Google Scholar]
- Balasubramanian, P.; Rajasekaran, A.; Prasad, S.N. Notes on the distribution and ethnobotany of some medicinal orchids in Nilgiri Biosphere Reserve. Zoos Print J. 2000, 15, 368. [Google Scholar] [CrossRef]
- Raveloarison, M.; Jeannoda, V.H.; Ramarosandratana, A.V. Traditional uses of wild Vanilla species in Madagascar. In Proceedings of the XXI AETFAT Congress, Nairobi, Kenya, 15–19 May 2017. [Google Scholar]
- Randrianarivony, T.N.; Ramarosandratana, A.V.; Andriamihajarivo, T.H.; Rakotoarivony, F.; Jeannoda, V.H.; Randrianasolo, A.; Bussmann, R.W. The most used medicinal plants by communities in Mahaboboka, Amboronabo, Mikoboka, Southwestern Madagascar. J. Ethnobiol. Ethnomed. 2017, 13, 19. [Google Scholar] [CrossRef]
- Rakotoarivelo, N.H.; Randrianarivony, T.N.; Rakotoarivony, F.; Randrianasolo, A. “Mangidy”: Malagasy folk beverages sold in Madagascar’s marketplaces. Ethnobot. Res. Appl. 2019, 18, 1–14. [Google Scholar] [CrossRef]
- Randriamiharisoa, M.N.; Kuhlman, A.R.; Jeannoda, V.; Rabarison, H.; Rakotoarivelo, N.; Randrianarivony, T.; Raktoarivony, F.; Randrianasolo, A.; Bussmann, R.W. Medicinal plants sold in the markets of Antananarivo, Madagascar. J. Ethnobiol. Ethnomed. 2015, 11, 60. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org (accessed on 17 September 2020).
- Fraga, C.N.D.; Couto, D.R.; Pansarin, E.R. Two new species of Vanilla (Orchidaceae) in the Brazilian Atlantic Forest. Phytotaxa 2017, 296, 63. [Google Scholar] [CrossRef]
- Pansarin, E.R. Vanilla dietschiana: An endangered species native to Brazil. Orchids 2010, 79, 106–109. [Google Scholar]
- Pansarin, E.R. Taxonomic notes on Vanilleae (Orchidaceae: Vanilloideae): Vanilla dietschiana, a Rare South American Taxon Transferred from Dictyophyllaria. Selbyana 2010, 30, 203–207. [Google Scholar]
- Raffi, A.; Nordin, F.A.; Abdullah, N.A.P.; Go, R. Vanilla norashikiniana R. Go et A. Raffi sp. nov., a new orchid species from Peninsular Malaysia. Malay. Nat. J. 2017, 69, 273–276. [Google Scholar]
- La Liste Rouge des Espèces Menacées en France: Flore Vasculaire de Mayotte. Available online: https://uicn.fr/wp-content/uploads/2016/06/Dossier_presse_Liste_rouge_Flore_vasculaire_de_Mayotte (accessed on 28 August 2020).
- Combrink, A.S.; Kyle, R. A Handbook on the Rare, Threatened and Endemic Species of the Greater St Lucia Wetland Park. A product of the Greater St Lucia Wetland Park-Rare, Threatened & Endemic Species Project, Unpublished Internal Report. Available online: https://www.globalnature.org/bausteine.net/f/6381/Rare%2CThreatenedandEndemicSpeciesReport_SouthAfrica.pdf?fd=2 (accessed on 29 August 2020).
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-521-87847-0. [Google Scholar]
- Flanagan, N.S.; Mosquera-Espinosa, A.T. An integrated strategy for the conservation and sustainable use of native Vanilla species in Colombia. Lankesteriana 2016, 16. [Google Scholar] [CrossRef]
- Van der Niet, T.; Peakall, R.; Johnson, S.D. Pollinator-driven ecological speciation in plants: New evidence and future perspectives. Ann. Bot. 2014, 113, 199–212. [Google Scholar] [CrossRef]
- Vences, M.; Wollenberg, K.C.; Vieites, D.R.; Lees, D.C. Madagascar as a model region of species diversification. Trends Ecol. Evol. 2009, 24, 456–465. [Google Scholar] [CrossRef]
- Burgess, N.D.; Hales, J.D.; Underwood, E.; Dinerstein, E.; Olson, D.; Itoua, I.; Schipper, J.; Ricketts, T.; Newman, K. Terrestrial Ecoregions of Africa and Madagascar: A Conservation Assessment; Island Press: Washington, DC, USA, 2004; ISBN 978-1-55963-364-2. [Google Scholar]
- Luke, W.R.Q. Rapid Assessment of Terrestrial Plant Diversity of Mnazi Bay Ruvuma Estuary Marine Park, Tanzania. Available online: https://portals.iucn.org/library/sites/library/files/documents/2004-098.pdf (accessed on 29 August 2020).
- Barley, A.J.; White, J.; Diesmos, A.C.; Brown, R.M. The challenge of species delimitation at the extremes: Diversification without morphological change in Philippine sun skinks. Evolution 2013, 67, 3556–3572. [Google Scholar] [CrossRef]
- Gottlieb, L.D. Rethinking classic examples of recent speciation in plants. New Phytol. 2003, 161, 71–82. [Google Scholar] [CrossRef]
- Shaffer, H.B.; Thomson, R.C. Delimiting species in recent radiations. Syst. Biol. 2007, 56, 896–906. [Google Scholar] [CrossRef]
- Rundell, R.J.; Price, T.D. Adaptive radiation, nonadaptive radiation, ecological speciation and nonecological speciation. Trends Ecol. Evol. 2009, 24, 394–399. [Google Scholar] [CrossRef]
- Grant, P.R.; Grant, B.R.; Petren, K. Hybridization in the recent past. Am. Nat. 2005, 166, 56–67. [Google Scholar] [CrossRef]
- Verma, P.C.; Chakrabarty, D.; Jena, S.N.; Mishra, D.K.; Singh, P.K.; Sawant, S.V.; Tuli, R. The extent of genetic diversity among Vanilla species: Comparative results for RAPD and ISSR. Ind. Crop. Prod. 2009, 29, 581–589. [Google Scholar] [CrossRef]
- Cameron, K.M. Vanilla Orchids: Natural History and Cultivation; Timber Press: Portland, OR, USA, 2011; ISBN 978-0-88192-989-8. [Google Scholar]
- Warren, B.H.; Bermingham, E.; Bowie, R.C.K.; Prys-Jones, R.P.; Thébaud, C. Molecular phylogeography reveals island colonization history and diversification of western Indian Ocean sunbirds (Nectarinia: Nectariniidae). Mol. Phylogenetics Evol. 2003, 29, 67–85. [Google Scholar] [CrossRef]
- Warren, B.H.; Bermingham, E.; Prys-Jones, R.P.; Thebaud, C. Tracking island colonization history and phenotypic shifts in Indian Ocean bulbuls (Hypsipetes: Pycnonotidae). Biol. J. Linn. Soc. 2005, 85, 271–287. [Google Scholar] [CrossRef]
- Kainulainen, K.; Razafimandimbison, S.G.; Wikström, N.; Bremer, B. Island hopping, long-distance dispersal and species radiation in the Western Indian Ocean: Historical biogeography of the Coffeeae alliance (Rubiaceae). J. Biogeogr. 2017, 44, 1966–1979. [Google Scholar] [CrossRef]
- Jaros, U.; Fischer, G.A.; Pailler, T.; Comes, H.P. Spatial patterns of AFLP diversity in Bulbophyllum occultum (Orchidaceae) indicate long-term refugial isolation in Madagascar and long-distance colonization effects in La Réunion. Heredity 2016, 116, 434–446. [Google Scholar] [CrossRef]
- Micheneau, C.; Carlsward, B.S.; Fay, M.F.; Bytebier, B.; Pailler, T.; Chase, M.W. Phylogenetics and biogeography of Mascarene angraecoid orchids (Vandeae, Orchidaceae). Mol. Phylogenetics Evol. 2008, 46, 908–922. [Google Scholar] [CrossRef]
- Andriananjamanantsoa, H.N. Systématique évolutive et biogéographie de Angraecum (Orchidaceae, Angraeinae) à Madagascar. Ph.D. Thesis, Université de Montréal, Montreal, QC, Canada, 2015. [Google Scholar]
- Warren, B.H.; Simberloff, D.; Ricklefs, R.E.; Aguilée, R.; Condamine, F.L.; Gravel, D.; Morlon, H.; Mouquet, N.; Rosindell, J.; Casquet, J.; et al. Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson. Ecol. Lett. 2015, 18, 200–217. [Google Scholar] [CrossRef]
- Darwin, C.M.A. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, 2nd ed.; John Murray: London, UK, 1859; pp. 10–394. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Blair, M.E.; Sterling, E.J.; Dusch, M.; Raxworthy, C.J.; Pearson, R.G. Ecological divergence and speciation between lemur (Eulemur) sister species in Madagascar. J. Evol. Biol. 2013, 26, 1790–1801. [Google Scholar] [CrossRef]
- Yoder, A.D.; Heckman, K.L. Mouse lemur phylogeography revises a model of ecogeographic constraint in Madagascar. In Primate Biogeography; Developments in Primatology: Progress and Prospects; Springer: Boston, MA, USA, 2006; pp. 255–268. ISBN 978-0-387-29871-9. [Google Scholar]
- Panitsa, M.; Koutsias, N.; Tsiripidis, I.; Zotos, A.; Dimopoulos, P. Species-based versus habitat-based evaluation for conservation status assessment of habitat types in the East Aegean islands (Greece). J. Nat. Conserv. 2011, 19, 269–275. [Google Scholar] [CrossRef]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Bortolus, A. Error cascades in the biological sciences: The unwanted consequences of using bad taxonomy in ecology. Ambio J. Hum. Environ. 2008, 37, 114–118. [Google Scholar] [CrossRef]
- Duminil, J.; Kenfack, D.; Viscosi, V.; Grumiau, L.; Hardy, O.J. Testing species delimitation in sympatric species complexes: The case of an African tropical tree, Carapa spp. (Meliaceae). Mol. Phylogenetics Evol. 2012, 62, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Rouhan, G.; Gaudeul, M. Plant taxonomy: A historical perspective, current challenges, and perspectives. In Molecular Plant Taxonomy; Methods in Molecular Biology; Besse, P., Ed.; Humana Press: Totowa, NJ, USA, 2014; Volume 1115, pp. 1–37. ISBN 978-1-62703-766-2. [Google Scholar]
- Besse, P.; Da Silva, D.; Grisoni, M. Plant DNA barcoding principles and limits: A case study in the genus Vanilla. In Molecular Plant Taxonomy Methods and Protocols; Besse, P., Ed.; Springer: New York, NY, USA, 2020; in press. [Google Scholar]
- Razafinarivo, N.J.; Guyot, R.; Davis, A.P.; Couturon, E.; Hamon, S.; Crouzillat, D.; Rigoreau, M.; Dubreuil-Tranchant, C.; Poncet, V.; De Kochko, A.; et al. Genetic structure and diversity of coffee (Coffea) across Africa and the Indian Ocean islands revealed using microsatellites. Ann. Bot. 2013, 111, 229–248. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stebbins, G.L. Variation and evolution in plants: Progress during the past twenty years. In Essays in Evolution and Genetics in Honor of Theodosius Dobzhansky: A Supplement to Evolutionary Biology; Hecht, M.K., Steere, W.C., Eds.; Springer: Boston, MA, USA, 1970; pp. 173–208. ISBN 978-1-4615-9585-4. [Google Scholar]
- Surveswaran, S.; Gowda, V.; Sun, M. Using an integrated approach to identify cryptic species, divergence patterns and hybrid species in Asian ladies’ tresses orchids (Spiranthes, Orchidaceae). Mol. Phylogenetics Evol. 2018, 124, 106–121. [Google Scholar] [CrossRef]
- Shafer, A.B.A.; Wolf, J.B.W. Widespread evidence for incipient ecological speciation: A meta-analysis of isolation-by-ecology. Ecol. Lett. 2013, 16, 940–950. [Google Scholar] [CrossRef]
- Ramírez-Barrera, S.M.; Velasco, J.A.; Orozco-Téllez, T.M.; Vázquez-López, A.M.; Hernández-Baños, B.E. What drives genetic and phenotypic divergence in the Red-crowned Ant tanager (Habia rubica, Aves: Cardinalidae), a polytypic species? Ecol. Evol. 2019, 9, 12339–12352. [Google Scholar] [CrossRef]
- Wang, I.J.; Bradburd, G.S. Isolation by environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Year | Continent | Group | Location |
|---|---|---|---|---|
| V. aphylla Blume. (V. calopogon) | 1825 | Asia | V. aphylla | Java, Malaysia, Philippines, Thailand |
| V. barbellata Rchb.f. (V. articulata, V. bakeri, V. parishii) | 1865 | America | V. barbellata | Caribbean Islands |
| V. bosseri L. Allorge. (V. montagnacii) | 2013 | Africa | V. phalaenopsis | Madagascar |
| V. claviculata Sw. | 1799 | America | V. barbellata | Caribbean Islands |
| V. decaryana H. Perrier. | 1934 | Africa | V. phalaenopsis | Madagascar |
| V. dilloniana Correll. | 1946 | America | V. barbellata | Caribbean Islands |
| V. humblotii Rchb.f. | 1885 | Africa | V. phalaenopsis | Comoros, Mayotte, Madagascar |
| V. madagascariensis Rolfe. | 1896 | Africa | V. phalaenopsis | Madagascar |
| V. marmoreisense Soto Calvo. | 2019 | America | V. barbellata | Cuba |
| V. penicillate Garay & Dunst. | 1965 | America | V. penicillata | Colombia, Suriname, Brazil |
| V. perrieri Schltr. | 1924 | Africa | V. phalaenopsis | Madagascar |
| V. phalaenopsis Rchb.f. | 1867 | Africa | V. phalaenopsis | Seychelles |
| V. poitaei Rchb.f. (V. correllii, V. eggersii) | 1876 | America | V. barbellata | Saint Domingue |
| V. roscheri Rchb.f. | 1876 | Africa | V. phalaenopsis | Tanzania, Kenya, Mozambique, South Africa |
| V. walkeriae Wight | 1845 | Asia | V. phalaenopsis | India, Sri Lanka |
| V. wightii Lindl. (V. wightiana) | 1845 | Asia | V. aphylla | India, Sri Lanka |
| Vanilla Species | Potential Pollinators | References |
|---|---|---|
| America | ||
| V. insignis | Eulaema polychrome | [20] |
| V. planifolia | Euglossa spp. | [20] |
| V. pompona subs grandiflora | Eulaema spp. | [42,56] |
| V. cribbiana | Eulaema sp. | [20] |
| V. pompona | Eulaema cingulata | [57] |
| V. trigonocarpa | Euglossa asarophora | [20] |
| V. edwallii | Epicharis affinis | [52] |
| Asia | ||
| V. siamensis | Thrinchostoma sp. | [58] |
| V. cf kaniensis | Aegilopa sp. | [14] |
| Africa | ||
| V. bosseri | Macrogalea ellioti, Liotrigona mahafalya, Liotrigona madecassa | [53] |
| V. roscheri | Allodapula variegate, Allodapa rufogastra, Anthophorini | [19] |
| V. humblotii | Allodape obscuripennis, Nectarinia coquerelli | [21] |
| Sympatric Species | Location | References |
|---|---|---|
| Leafy Species | ||
| V. hartii & V. cribbiana | Mexico | [20] |
| V. hartii & V. insignis | Mexico | [20] |
| V. hartii & V. odorata | Mexico | [20] |
| V. hartii & V. inodora | Mexico | [20] |
| V. rivasii & V. dressleri | Colombia | [44] |
| V. javieri & V. appendiculata | Colombia | [66] |
| V. yanesha & V. mexicana | Peru | [38] |
| Leafless Species | ||
| V. barbellata & V. claviculata | Puerto Rico | [43] |
| V. barbellata & V. dilloniana | Puerto Rico | [22] |
| V. dilloniana & V. claviculata | Puerto Rico | [22] |
| V. madagascariensis & V. perrieri | Madagascar | [8] 1 |
| V. madagascariensis & V. bosseri | Madagascar | 1 |
| Distribution | Flowering Period | Flower | Stem |
|---|---|---|---|
| V. bosseri L. Allorge, 2013 | |||
| Madagascar: Southwesten a Namoroka a Morondava a | September-November b,c | LS: 4.5 cm a C: white a La: pinkish throat with no pubescence and red protrusion between the crests a,b | C: grey in young stems and turns green in old stems a F: four longitudinal grooves a Tx: generally glabrous a,c |
| V. decaryana H. Perrier, 1934 | |||
| Madagascar d: South d Southwestern e Andohahela e | January d | LP: 3 cm d LS: 2.5 cm d Fl: 20–40 flowers d La: pinkish throat with two pubescent lines d | C: greenish-brown d F: two longitudinal grooves e Tx: glabrous c,d,e |
| V. humblotii Reichenbach f., 1885 | |||
| Comoros d, Mayotte d,g Madagascar f: North f Ankarana e Bemaraha e Andohahela e | November-April g | C: canary-yellow d,g LP: 5.5–7 cm d LS: 5.5–6.5 cm d La: very pubescent with velvety red or dark maroon throat d,h, papillosae h Fl: 40–50 flowers g | C: glaucous green d or bluish e F: one longitudinal groove e Tx: verrucose c,d,e |
| V.madagascariensis Rolfe, 1896 | |||
| Madagascar d: North (Sava, Diana, Ankarana) a Isalo a Ivohibe a | June-October a,d | C: white d LP: 5.5–6.5 cm a LS: 5.5–6.5 cm d Fl: 10–20 flowers a,d La: pubescent with red throat a | C: glaucous green d F: one longitudinal grooves e Tx: glabrous a,d,e |
| V. perrieri Schlechter, 1925 | |||
| Madagascar: Ankarafantsika d Western coast (Majunga, Morondava, Fort Dauphin, Ambovombe) e | November-December c | C: sharp yellow d LP: 5.5 cm d LS: 6.5 cm d La: pubescent with bright red or dark brownish throat d,h not papillosae h, extension of the hair on the back d | C: dark green e F: two longitudinal grooves e Tx: glabrous d,e, similar to V. madagascariensis, but sometimes verrucose c |
| V. phalaenopsis Reichenbach f., 1868 | |||
| Seychelles a | April i July-November j | C: white d LP: 5.5–6 cm d LS: 5.5–6 cm d Fl: corymbiform d La: two pubescent lines and yellow-orange desk d | C: green i F: one longitudinal groove d Tx: glabrous, similar to V. madagascariensis i |
| V. roscheri Reichenbach f., 1877 | |||
| East African: Mozambique d Zanzibar d Pemba Island d Dar-es-Salam d South Africa k | November-March k | C: white d LP: 7.5–8 cm d LS: 7.5–8 cm d Bud: brownish d La: two pubescent lines desk and pink throat d,k | C: reddish-brown d Tx: glabrous similar to V. madagascariensis i |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andriamihaja, C.F.; Ramarosandratana, A.V.; Grisoni, M.; Jeannoda, V.; Besse, P. The Leafless Vanilla Species-Complex from the South-West Indian Ocean Region: A Taxonomic Puzzle and a Model for Orchid Evolution and Conservation Research. Diversity 2020, 12, 443. https://doi.org/10.3390/d12120443
Andriamihaja CF, Ramarosandratana AV, Grisoni M, Jeannoda V, Besse P. The Leafless Vanilla Species-Complex from the South-West Indian Ocean Region: A Taxonomic Puzzle and a Model for Orchid Evolution and Conservation Research. Diversity. 2020; 12(12):443. https://doi.org/10.3390/d12120443
Chicago/Turabian StyleAndriamihaja, Cathucia F., Aro V. Ramarosandratana, Michel Grisoni, Vololoniaina Jeannoda, and Pascale Besse. 2020. "The Leafless Vanilla Species-Complex from the South-West Indian Ocean Region: A Taxonomic Puzzle and a Model for Orchid Evolution and Conservation Research" Diversity 12, no. 12: 443. https://doi.org/10.3390/d12120443
APA StyleAndriamihaja, C. F., Ramarosandratana, A. V., Grisoni, M., Jeannoda, V., & Besse, P. (2020). The Leafless Vanilla Species-Complex from the South-West Indian Ocean Region: A Taxonomic Puzzle and a Model for Orchid Evolution and Conservation Research. Diversity, 12(12), 443. https://doi.org/10.3390/d12120443