
Urinary Extracellular Vesicle Signatures as Biomarkers in Prostate Cancer Patients

 ,
,  ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Results
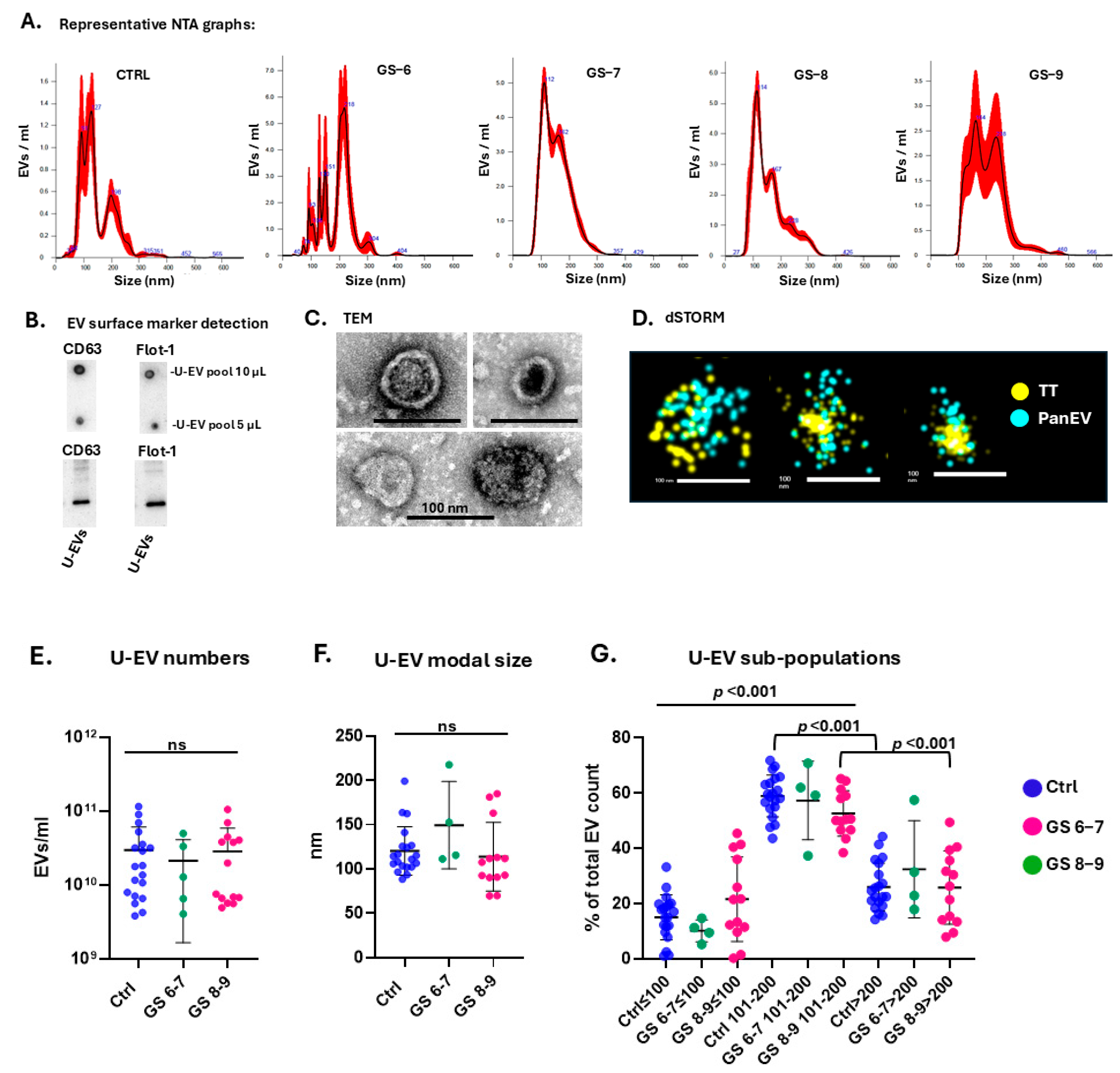
2.1. Characterisation, Quantification, and Size Profiling of Urinary EVs from Healthy Controls and Prostate Cancer Patients
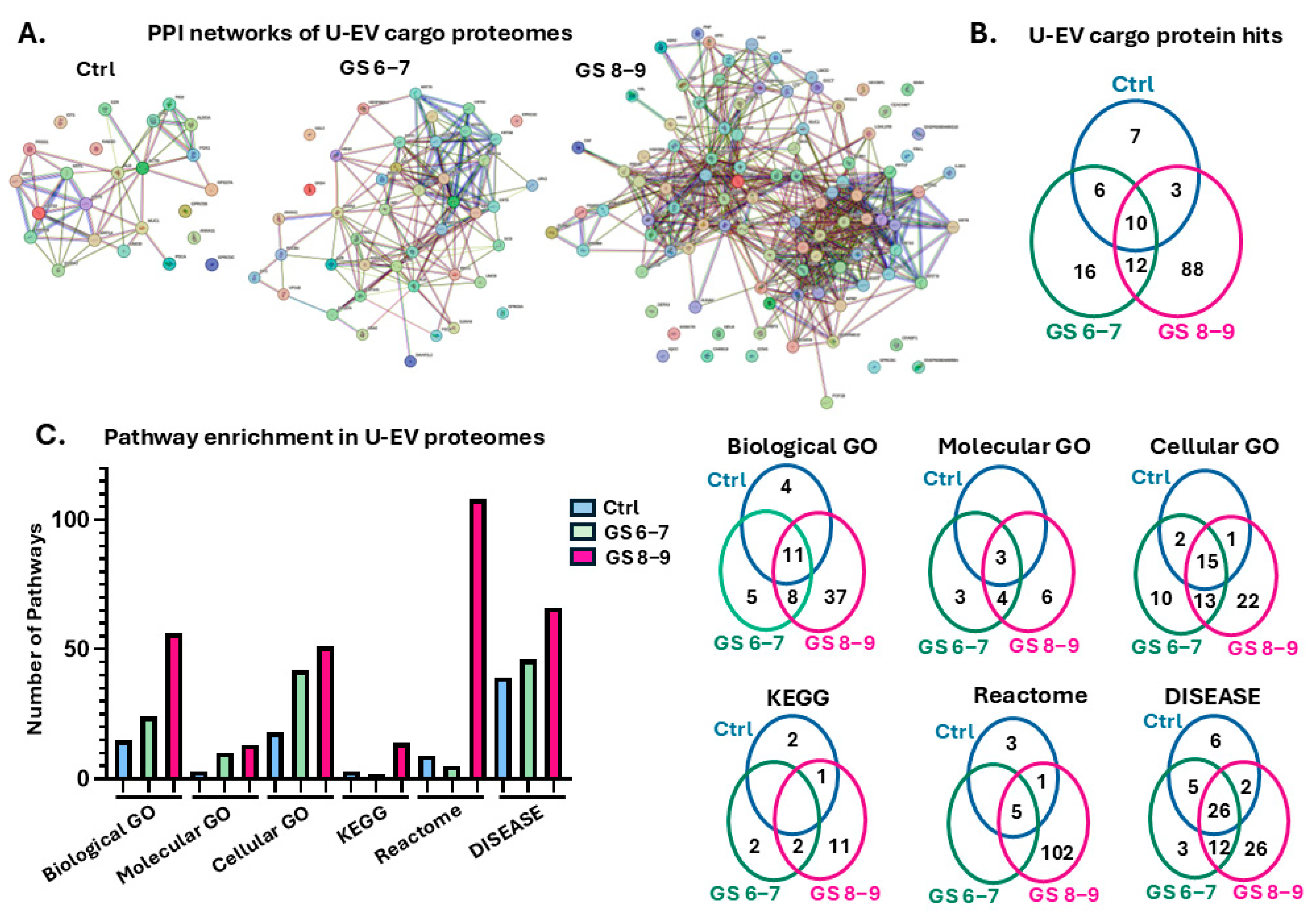
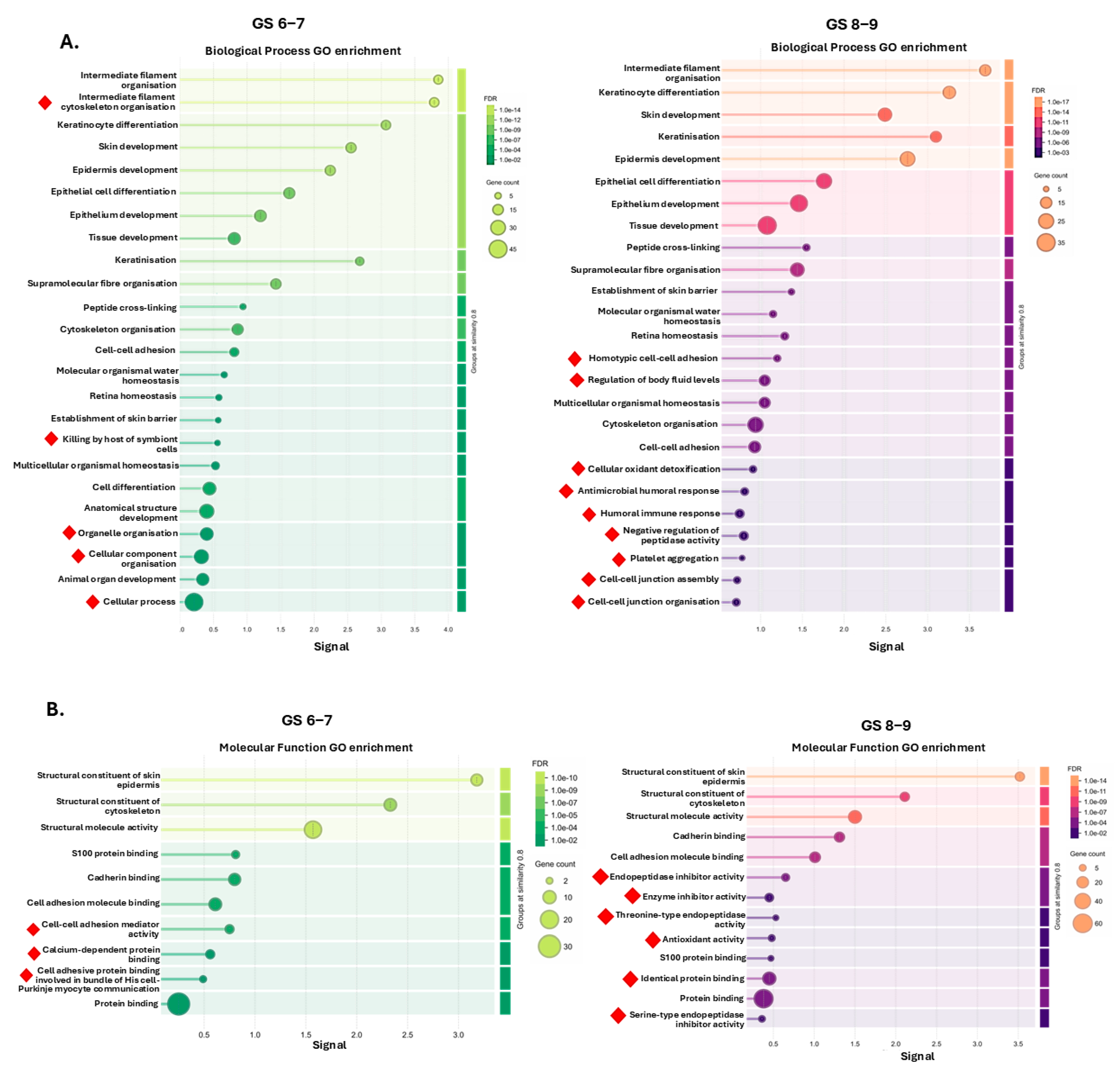
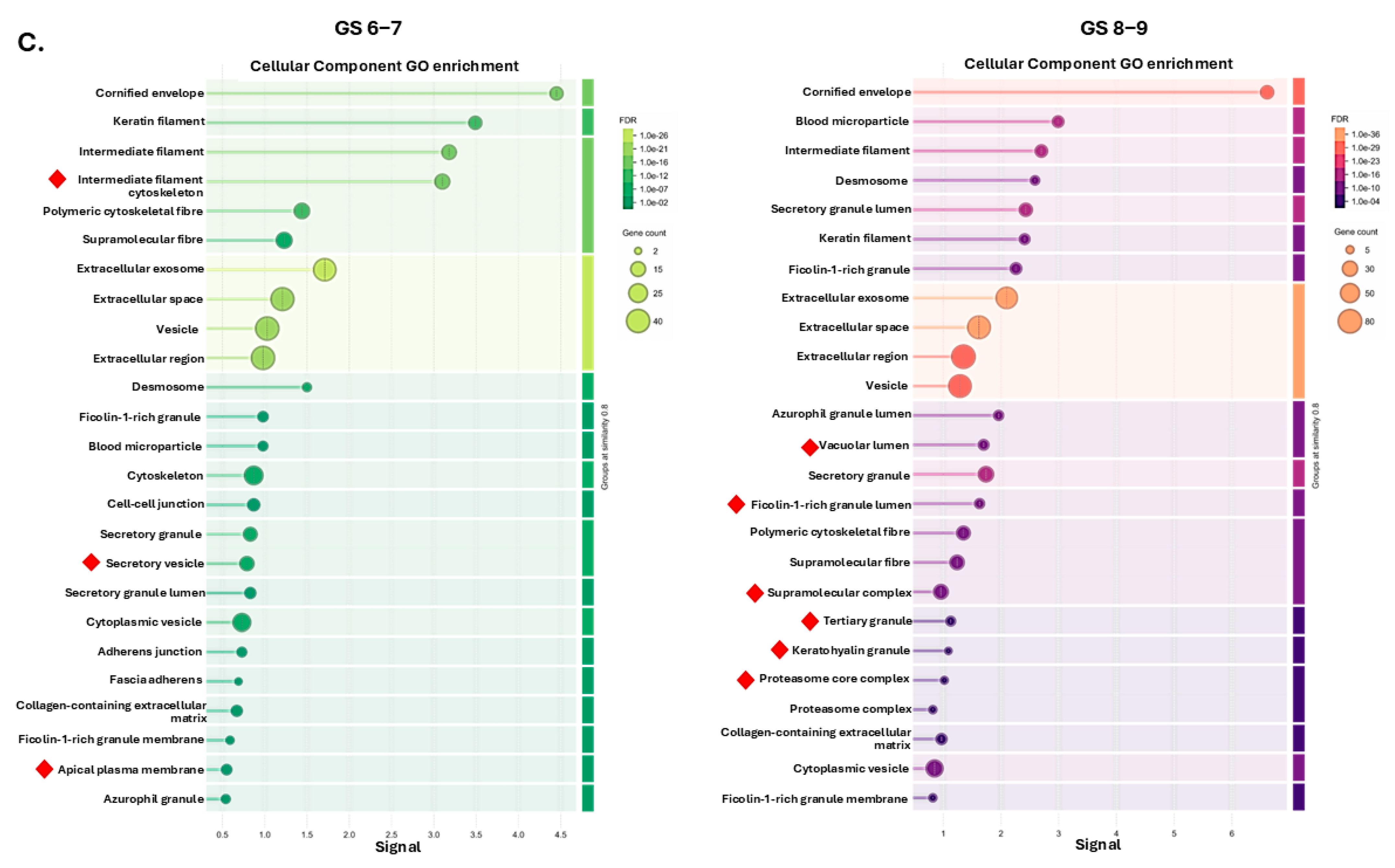
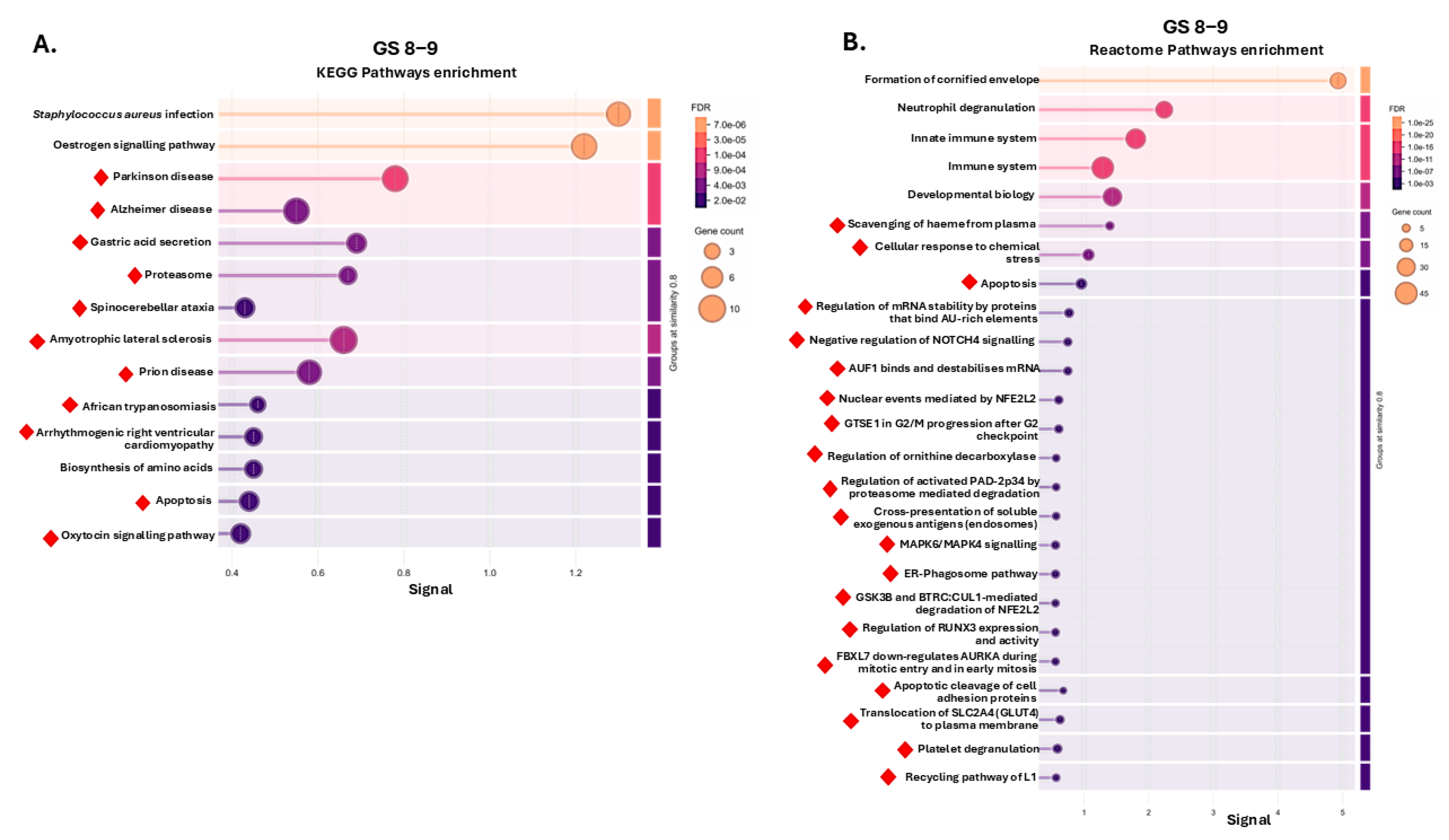
2.2. Protein–Protein Interaction (PPI) Network and Pathway Enrichment Analysis for Urinary EV Proteome Cargoes
3. Discussion
4. Materials and Methods
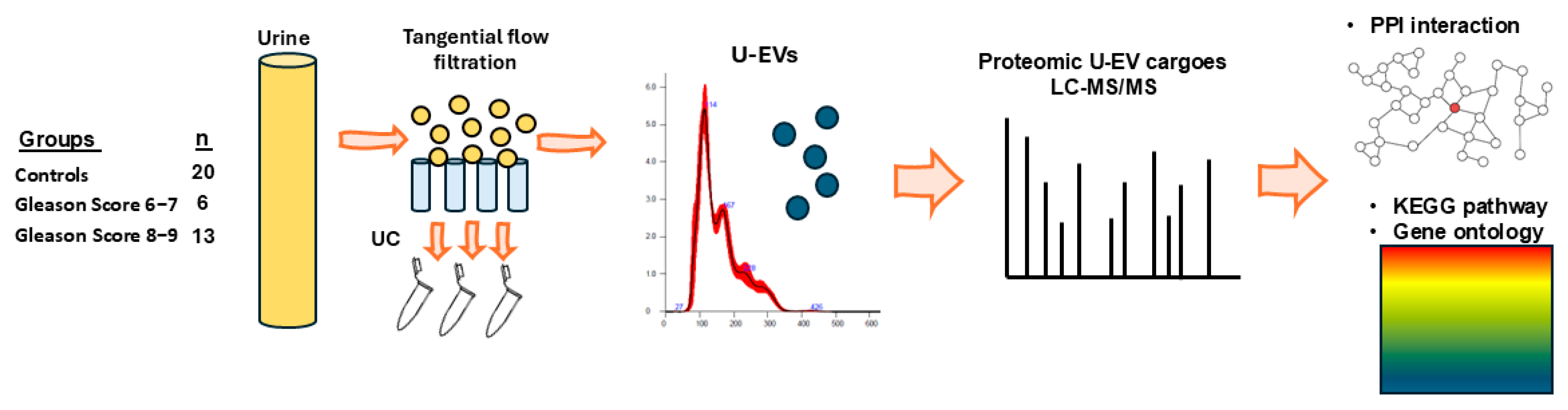
4.1. Urinary Samples from Patients
4.2. Isolation and Characterisation of Extracellular Vesicles from Urine
4.2.1. Nanoparticle Tracking Analysis
4.2.2. Western Blotting for EV Surface Markers
4.2.3. Transmission Electron Microscopy
4.2.4. Direct Stochastic Optical Reconstruction Microscopy (dSTORM) Imaging of U-EVs
4.3. Proteomic Cargo Analysis of U-EVs by Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS)
4.4. Protein–Protein Interaction (PPI) Networks and Functional Pathway Enrichment Analysis of U-EV Proteome Cargoes
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Descotes, J.L. Diagnosis of prostate cancer. Asian J. Urol. 2019, 6, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Erdbrügger, U.; Blijdorp, C.J.; Bijnsdorp, I.V.; Borràs, F.E.; Burger, D.; Bussolati, B.; Byrd, J.B.; Clayton, A.; Dear, J.W.; Falcón-Pérez, J.M.; et al. Urinary extracellular vesicles: A position paper by the Urine Task Force of the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2021, 10, e12093. [Google Scholar] [CrossRef] [PubMed]
- Bajo-Santos, C.; Brokāne, A.; Zayakin, P.; Endzeliņš, E.; Soboļevska, K.; Belovs, A.; Jansons, J.; Sperga, M.; Llorente, A.; Radoviča-Spalviņa, I.; et al. Plasma and urinary extracellular vesicles as a source of RNA biomarkers for prostate cancer in liquid biopsies. Front. Mol. Biosci. 2023, 10, 980433. [Google Scholar] [CrossRef] [PubMed]
- Steiner, L.; Eldh, M.; Offens, A.; Veerman, R.E.; Johansson, M.; Hemdan, T.; Netterling, H.; Huge, Y.; Abdul-Sattar Aljabery, F.; Alamdari, F.; et al. Protein profile in urinary extracellular vesicles is a marker of malignancy and correlates with muscle invasiveness in urinary bladder cancer. Cancer Lett. 2025, 609, 217352. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, K.P.; Rossi, I.; Abdullahi, M.; Ramirez, M.I.; Stratton, D.; Inal, J.M. Isolation and characterization of extracellular vesicles and future directions in diagnosis and therapy. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2023, 15, e1835. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.E.; Piedad, J.; Hemsworth, L.; West, A.; Johnston, I.D.; Dimov, N.; Inal, J.M.; Vasdev, N. Prostate cancer and microfluids. Urol. Oncol. 2021, 39, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Lu, F.; Chen, J.; Jiao, Y.; Qiu, Q.; Nian, X.; Qu, M.; Wang, Y.; Li, M.; Liu, F.; et al. UPCARE: Urinary Extracellular Vesicles-Derived Prostate Cancer Assessment for Risk Evaluation. J. Extracell. Vesicles 2024, 13, e12491. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.F.; Brewer, D.S.; Hurst, R.; Cooper, C.S. Applications of Urinary Extracellular Vesicles in the Diagnosis and Active Surveillance of Prostate Cancer. Cancers 2024, 16, 1717. [Google Scholar] [CrossRef] [PubMed]
- Khoo, A.; Govindarajan, M.; Qiu, Z.; Liu, L.Y.; Ignatchenko, V.; Waas, M.; Macklin, A.; Keszei, A.; Neu, S.; Main, B.P.; et al. Prostate cancer reshapes the secreted and extracellular vesicle urinary proteomes. Nat. Commun. 2024, 15, 5069. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Wang, Q.; Chen, H.; Liu, Z.; Han, M.; Gong, J.; Yue, L.; Ding, X.; Wang, S.; Yan, Z.; et al. Proteomic Identification of Small Extracellular Vesicle Proteins LAMB1 and Histone H4 for Prostate Cancer Diagnosis and Risk Stratification. Adv. Sci. 2024, 11, e2402509. [Google Scholar] [CrossRef] [PubMed]
- Bernardino, R.; Carvalho, A.S.; Hall, M.J.; Alves, L.; Leão, R.; Sayyid, R.; Pereira, H.; Beck, H.C.; Pinheiro, L.C.; Henrique, R.; et al. Profiling of urinary extracellular vesicle protein signatures from patients with cribriform and intraductal prostate carcinoma in a cross-sectional study. Sci. Rep. 2024, 14, 25065. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.I.; Egevad, L.; Amin, M.B.; Delahunt, B.; Srigley, J.R.; Humphrey, P.A.; Grading Committee. The 2014 International Society of Urological Pathology (ISUP) Consensus Conference on Gleason Grading of Prostatic Carcinoma: Definition of Grading Patterns and Proposal for a New Grading System. Am. J. Surg. Pathol. 2016, 40, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Randall, E.C.; Zadra, G.; Chetta, P.; Lopez, B.G.C.; Syamala, S.; Basu, S.S.; Agar, J.N.; Loda, M.; Tempany, C.M.; Fennessy, F.M.; et al. Molecular Characterization of Prostate Cancer with Associated Gleason Score Using Mass Spectrometry Imaging. Mol. Cancer Res. 2019, 17, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Jeanmard, N.; Bissanum, R.; Sriplung, H.; Charoenlappanit, S.; Roytrakul, S.; Navakanitworakul, R. Proteomic profiling of urinary extracellular vesicles differentiates breast cancer patients from healthy women. PLoS ONE 2023, 18, e0291574. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.; Jiang, W.G.; Mason, M.D. Cell-cell adhesion molecules and signaling intermediates and their role in the invasive potential of prostate cancer cells. J. Urol. 2000, 163, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liang, Y.; Deavers, M.T.; Kamat, A.M.; Matin, S.F.; Dinney, C.P.; Czerniak, B.; Guo, C.C. Uroplakin II is a more sensitive immunohistochemical marker than uroplakin III in urothelial carcinoma and its variants. Am. J. Clin. Pathol. 2014, 142, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, E.S. Mining The Cancer Genome Atlas gene expression data for lineage markers in distinguishing bladder urothelial carcinoma and prostate adenocarcinoma. Sci. Rep. 2021, 11, 6765. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.M.; Li, J.; Kang, A.D.; Huang, S.Q.; Liu, W.B.; Zhang, Y.; Liu, Z.H.; Zeng, L. Protein S100-A8: A potential metastasis-associated protein for breast cancer determined via iTRAQ quantitative proteomic and clinicopathological analysis. Oncol. Lett. 2018, 15, 5285–5293. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Cao, H.; Liu, C.; Sun, X.; Dai, S.; Liu, L.; Wang, Y.; Guo, C.; Wang, X.; Gao, Y.; et al. MAL2 reprograms lipid metabolism in intrahepatic cholangiocarcinoma via EGFR/SREBP-1 pathway based on single-cell RNA sequencing. Cell Death Dis. 2024, 15, 411. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Choi, Y.J.; Kim, S.G.; Nam, S.W.; Lee, J.Y.; Park, W.S. Programmed cell death 6 (PDCD6) as a prognostic marker for gastric cancers. Tumour Biol. 2012, 33, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Riddell, J.R.; Bshara, W.; Moser, M.T.; Spernyak, J.A.; Foster, B.A.; Gollnick, S.O. Peroxiredoxin 1 controls prostate cancer growth through Toll-like receptor 4-dependent regulation of tumor vasculature. Cancer Res. 2011, 71, 1637–1646. [Google Scholar] [CrossRef] [PubMed]
- Scaggiante, B.; Dapas, B.; Bonin, S.; Grassi, M.; Zennaro, C.; Farra, R.; Cristiano, L.; Siracusano, S.; Zanconati, F.; Giansante, C.; et al. Dissecting the expression of EEF1A1/2 genes in human prostate cancer cells: The potential of EEF1A2 as a hallmark for prostate transformation and progression. Br. J. Cancer 2012, 106, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Hui, Y.; Peng, H.; Zhang, H.; Li, M.; Song, L.; Li, F.; Cui, X. Identification of a PLCE1-regulated competing endogenous RNA regulatory network for esophageal squamous cell carcinoma. Oncol. Rep. 2021, 45, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Qiao, G.; Wu, A.; Chen, X.; Tian, Y.; Lin, X. Enolase 1, a Moonlighting Protein, as a Potential Target for Cancer Treatment. Int. J. Biol. Sci. 2021, 17, 3981–3992. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Chen, X.; Xu, H.; Zhi, Y.; Zhuo, X.; Yu, J.; Zhao, L. N6-methyladenosine RNA modified BAIAP2L2 facilitates extracellular vesicles-mediated chemoresistance transmission in gastric cancer. J. Transl. Med. 2025, 23, 320. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Fujimoto, J.; Men, T.; Ye, X.; Deng, J.; Lacroix, L.; Clifford, J.L.; Mao, L.; Van Pelt, C.S.; Lee, J.J.; et al. Identification of the retinoic acid-inducible Gprc5a as a new lung tumor suppressor gene. J. Natl. Cancer Inst. 2007, 99, 1668–1682. [Google Scholar] [CrossRef] [PubMed]
- Sundararaj, S.; Ravindran, A.; Casarotto, M.G. AHNAK: The quiet giant in calcium homeostasis. Cell Calcium. 2021, 96, 102403. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.A.; Smuczek, B.; Valadão, I.C.; Dzik, L.M.; Iglesia, R.P.; Cruz, M.C.; Zelanis, A.; de Siqueira, A.S.; Serrano, S.M.; Goldberg, G.S.; et al. AHNAK enables mammary carcinoma cells to produce extracellular vesicles that increase neighboring fibroblast cell motility. Oncotarget 2016, 7, 49998–50016. [Google Scholar] [CrossRef] [PubMed]
- Labrie, M.; Vladoiu, M.; Leclerc, B.G.; Grosset, A.A.; Gaboury, L.; Stagg, J.; St-Pierre, Y. A Mutation in the Carbohydrate Recognition Domain Drives a Phenotypic Switch in the Role of Galectin-7 in Prostate Cancer. PLoS ONE 2015, 10, e0131307. [Google Scholar] [CrossRef] [PubMed]
- Kohli, M.; Young, C.Y.; Tindall, D.J.; Nandy, D.; McKenzie, K.M.; Bevan, G.H.; Donkena, K.V. Whole blood defensin mRNA expression is a predictive biomarker of docetaxel response in castration-resistant prostate cancer. Oncol. Targets Ther. 2015, 8, 1915–1922. [Google Scholar] [CrossRef]
- Chen, G.; Yu, C.; Tang, Z.; Liu, S.; An, F.; Zhu, J.; Wu, Q.; Cao, J.; Zhan, Q.; Zhang, S. Metformin suppresses gastric cancer progression through calmodulin-like protein 3 secreted from tumor-associated fibroblasts. Oncol. Rep. 2019, 41, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Gentile, F.; La Civita, E.; Ventura, B.D.; Ferro, M.; Bruzzese, D.; Crocetto, F.; Tennstedt, P.; Steuber, T.; Velotta, R.; Terracciano, D. A Neural Network Model Combining [-2]proPSA, freePSA, Total PSA, Cathepsin D, and Thrombospondin-1 Showed Increased Accuracy in the Identification of Clinically Significant Prostate Cancer. Cancers 2023, 15, 1355. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.J.; Mondul, A.M.; Layne, T.M.; Yu, K.; Huang, J.; Stolzenberg-Solomon, R.Z.; Ziegler, R.G.; Purdue, M.P.; Huang, W.Y.; Abnet, C.C.; et al. Prediagnostic Serum Vitamin D, Vitamin D Binding Protein Isoforms, and Cancer Survival. JNCI Cancer Spectr. 2022, 6, pkac019. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Miyamoto, D.T.; Wittner, B.S.; Sullivan, J.P.; Aceto, N.; Jordan, N.V.; Yu, M.; Karabacak, N.M.; Comaills, V.; Morris, R.; et al. Expression of β-globin by cancer cells promotes cell survival during blood-borne dissemination. Nat. Commun. 2017, 8, 14344. [Google Scholar] [CrossRef] [PubMed]
- Albany, C.; Hahn, N.M. Heat shock and other apoptosis-related proteins as therapeutic targets in prostate cancer. Asian J. Androl. 2014, 16, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Stope, M.B.; Stender, C.; Schubert, T.; Peters, S.; Weiss, M.; Ziegler, P.; Zimmermann, U.; Walther, R.; Burchardt, M. Heat-shock protein HSPB1 attenuates microRNA miR-1 expression thereby restoring oncogenic pathways in prostate cancer cells. Anticancer Res. 2014, 34, 3475–3480. [Google Scholar] [PubMed]
- Yang, C.; Fischer-Kešo, R.; Schlechter, T.; Ströbel, P.; Marx, A.; Hofmann, I. Plakophilin 1-deficient cells upregulate SPOCK1: Implications for prostate cancer progression. Tumour Biol. 2015, 36, 9567–9577. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Reidenbach, S.; Schlechter, T.; Rothmann, A.C.; Will, R.; Hofmann, I. Plakophilin 1 deficiency in prostatic tumours is correlated with immune cell recruitment and controls the up-regulation of cytokine expression post-transcriptionally. FEBS J. 2023, 290, 1907–1919. [Google Scholar] [CrossRef] [PubMed]
- Breuninger, S.; Reidenbach, S.; Sauer, C.G.; Ströbel, P.; Pfitzenmaier, J.; Trojan, L.; Hofmann, I. Desmosomal plakophilins in the prostate and prostatic adenocarcinomas: Implications for diagnosis and tumor progression. Am. J. Pathol. 2010, 176, 2509–2519. [Google Scholar] [CrossRef] [PubMed]
- Ezponda, T.; Popovic, R.; Shah, M.Y.; Martinez-Garcia, E.; Zheng, Y.; Min, D.J.; Will, C.; Neri, A.; Kelleher, N.L.; Yu, J.; et al. The histone methyltransferase MMSET/WHSC1 activates TWIST1 to promote an epithelial-mesenchymal transition and invasive properties of prostate cancer. Oncogene 2013, 32, 2882–2890. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, L.; Li, H.; Jin, C.; Yu, Y.; Hou, L.; Liu, X.; Yu, Y.; Yan, R.; Xue, F. Identification of microenvironment related potential biomarkers of biochemical recurrence at 3 years after prostatectomy in prostate adenocarcinoma. Aging 2021, 13, 16024–16042. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Li, X.; Jia, Z.; Gu, C.; Jin, Z.; Wang, J.; Yuan, B.; Yang, J. Bioinformatics Analysis of GFAP as a Potential Key Regulator in Different Immune Phenotypes of Prostate Cancer. Biomed. Res. Int. 2021, 2021, 1466255. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.; Park, J.; Lee, J.S.; Yoe, J.; Park, G.Y.; Kim, E.; Jeon, H.; Cho, Y.M.; Roh, T.Y.; Lee, Y. miR-93/miR-106b/miR-375-CIC-CRABP1: A novel regulatory axis in prostate cancer progression. Oncotarget 2015, 6, 23533–23547. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.H.; Young, D.; Chen, Y.; Kuo, H.C.; Srinivasan, A.; Dobi, A.; Petrovics, G.; Cullen, J.; Mcleod, D.G.; Rosner, I.L.; et al. Prognostic features of Annexin A2 expression in prostate cancer. Pathology 2021, 53, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.D.; Lowrie, A.G.; Riddick, A.C.; Fearon, K.C.; Habib, F.K.; Ross, J.A. Dermcidin expression confers a survival advantage in prostate cancer cells subjected to oxidative stress or hypoxia. Prostate 2007, 67, 1308–1317. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, B.; Bräutigam, K.; Dartiguenave, F.; Martel, P.; Rakauskas, A.; Cesson, V.; Veit, M.; Oechslin, P.; Gu, A.; Hermanns, T.; et al. Urine biomarkers can predict prostate cancer and PI-RADS score prior to biopsy. Sci. Rep. 2024, 14, 18148. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.Z.; Miao, X.; Chang, J.; Zhou, H.; Zhang, J.J.; Mo, H.M.; Jia, Q. Smoking behavior associated upregulation of SERPINB12 promotes proliferation and metastasis via activating WNT signaling in NSCLC. J. Cardiothorac. Surg. 2024, 19, 141. [Google Scholar] [CrossRef] [PubMed]
- Clos-Garcia, M.; Loizaga-Iriarte, A.; Zuñiga-Garcia, P.; Sánchez-Mosquera, P.; Rosa Cortazar, A.; González, E.; Torrano, V.; Alonso, C.; Pérez-Cormenzana, M.; Ugalde-Olano, A.; et al. Metabolic alterations in urine extracellular vesicles are associated to prostate cancer pathogenesis and progression. J. Extracell. Vesicles 2018, 7, 1470442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, G.Y.; Su, W.C.; Chen, Y.T.; Liu, Y.F.; Wei, D.; Zhang, Y.X.; Tang, Q.Y.; Liu, Y.X.; Wang, S.Z.; et al. High Throughput Isolation and Data Independent Acquisition Mass Spectrometry (DIA-MS) of Urinary Extracellular Vesicles to Improve Prostate Cancer Diagnosis. Molecules 2022, 27, 8155. [Google Scholar] [CrossRef] [PubMed]
- Lafront, C.; Germain, L.; Campolina-Silva, G.H.; Weidmann, C.; Berthiaume, L.; Hovington, H.; Brisson, H.; Jobin, C.; Frégeau-Proulx, L.; Cotau, R.; et al. The estrogen signaling pathway reprograms prostate cancer cell metabolism and supports proliferation and disease progression. J. Clin. Investig. 2024, 134, e170809. [Google Scholar] [CrossRef] [PubMed]
- Federico, G.; Carotenuto, A.R.; Cutolo, A.; Palumbo, S.; Moccia, M.; Paladino, S.; Santoro, M.; Russo, T.; Fraldi, M.; Carlomagno, F. Ultrasound-induced mechanical damage of cancer cell cytoskeleton causes disruption of nuclear envelope and activation of cGAS-STING. Sci. Rep. 2025, 15, 18037. [Google Scholar] [CrossRef] [PubMed]
- Novysedlak, R.; Guney, M.; Al Khouri, M.; Bartolini, R.; Koumbas Foley, L.; Benesova, I.; Ozaniak, A.; Novak, V.; Vesely, S.; Pacas, P.; et al. The Immune Microenvironment in Prostate Cancer: A Comprehensive Review. Oncology 2024, 103, 521–545. [Google Scholar] [CrossRef] [PubMed]
- Pujana-Vaquerizo, M.; Bozal-Basterra, L.; Carracedo, A. Metabolic adaptations in prostate cancer. Br. J. Cancer 2024, 131, 1250–1262. [Google Scholar] [CrossRef] [PubMed]
- Achard, V.; Ceyzériat, K.; Tournier, B.B.; Frisoni, G.B.; Garibotto, V.; Zilli, T. Biomarkers to Evaluate Androgen Deprivation Therapy for Prostate Cancer and Risk of Alzheimer’s Disease and Neurodegeneration: Old Drugs, New Concerns. Front. Oncol. 2021, 11, 734881. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.; Roy, A.; Summers, C.; Ahles, T.; Li, C.R.; Chao, H.H. Androgen deprivation increases frontopolar cortical thickness in prostate cancer patients: An effect of early neurodegeneration? Am. J. Cancer Res. 2024, 14, 3652–3664. [Google Scholar] [CrossRef] [PubMed]
- Chaldakov, G.N.; Aloe, L.; Yanev, S.G.; Fiore, M.; Tonchev, A.B.; Vinciguerra, M.; Evtimov, N.T.; Ghenev, P.; Dikranian, K. Trackins (Trk-Targeting Drugs): A Novel Therapy for Different Diseases. Pharmaceuticals 2024, 17, 961. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, Y.; Numbere, N.; Wang, Y.; Miyamoto, H. Clinical Significance of Perineural Invasion by Prostate Cancer Detected on Needle Core Biopsy. Am. J. Clin. Pathol. 2023, 159, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Zareba, P.; Flavin, R.; Isikbay, M.; Rider, J.R.; Gerke, T.A.; Finn, S.; Pettersson, A.; Giunchi, F.; Unger, R.H.; Tinianow, A.M.; et al. Perineural Invasion and Risk of Lethal Prostate Cancer. Cancer Epidemiol. Biomarkers Prev. 2017, 26, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Koshiuka, K.; Elstner, E.; Williamson, E.; Said, J.W.; Tada, Y.; Koeffler, H.P. Novel therapeutic approach: Organic arsenical melarsoprol) alone or with all-trans-retinoic acid markedly inhibit growth of human breast and prostate cancer cells in vitro and in vivo. Br. J. Cancer. 2000, 82, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Bowden, C.J.; Figg, W.D.; Dawson, N.A.; Sartor, O.; Bitton, R.J.; Weinberger, M.S.; Headlee, D.; Reed, E.; Myers, C.E.; Cooper, M.R. A phase I/II study of continuous infusion suramin in patients with hormone-refractory prostate cancer: Toxicity and response. Cancer Chemother. Pharmacol. 1996, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Teng, M.; Yan, X.; Liu, Q. The role of serum interleukins in Cancer: A Multi-center Mendelian Randomization study. Int. Immunopharmacol. 2024, 137, 112520. [Google Scholar] [CrossRef] [PubMed]
- Melegh, Z.; Oltean, S. Targeting Angiogenesis in Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 2676. [Google Scholar] [CrossRef] [PubMed]
- Gurumurthy, S.; Vasudevan, K.M.; Rangnekar, V.M. Regulation of apoptosis in prostate cancer. Cancer Metastasis Rev. 2001, 20, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Qing, Y.; Wang, Y.; Hu, C.; Zhang, H.; Huang, Y.; Zhang, Z.; Ma, T.; Zhang, S.; Li, K. Evaluation of NOTCH family genes’ expression and prognostic value in prostate cancer. Transl. Androl. Urol. 2022, 11, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Dai, T.; Guo, M.; Zhao, X.; Chen, C.; Zhou, Y.; Qin, M.; Xu, L.; Zhao, J. A review of non-classical MAPK family member, MAPK4: A pivotal player in cancer development and therapeutic intervention. Int. J. Biol. Macromol. 2024, 271 Pt 2, 132686. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Thrasher, J.B.; Terranova, P. Glycogen synthase kinase-3: A potential preventive target for prostate cancer management. Urol. Oncol. 2015, 33, 456–463. [Google Scholar] [CrossRef]
- Chen, F.; Wang, M.; Bai, J.; Liu, Q.; Xi, Y.; Li, W.; Zheng, J. Role of RUNX3 in suppressing metastasis and angiogenesis of human prostate cancer. PLoS ONE 2014, 9, e86917. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ma, J.; Wang, X.; Zi, T.; Qian, D.; Li, C.; Xu, C. CCNB1 and AURKA are critical genes for prostate cancer progression and castration-resistant prostate cancer resistant to vinblastine. Front. Endocrinol. 2022, 13, 1106175. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Liu, C.; Shi, J.; Lai, C.; Gao, Z.; Luo, J.; Li, Z.; Tang, Z.; Li, K.; Xu, K. HMMR promotes prostate cancer proliferation and metastasis via AURKA/mTORC2/E2F1 positive feedback loop. Cell Death Discov. 2023, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.X.; Wang, Y.S.; Ren, Y.Y.; Dong, X.M.; Wu, P.; Xie, T.; Zhang, Q.; Zhou, J.L. Prostate cancer microenvironment: Multidimensional regulation of immune cells, vascular system, stromal cells, and microbiota. Mol. Cancer 2024, 23, 229. [Google Scholar] [CrossRef] [PubMed]
- Moshref, L.; Abidullah, M.; Czaykowski, P.; Chowdhury, A.; Wightman, R.; Hebbard, P. Prostate Cancer Metastasis to Stomach: A Case Report and Review of Literature. Curr. Oncol. 2023, 30, 3901–3914. [Google Scholar] [CrossRef] [PubMed]
- Odat, R.M.; Idrees, M.; Marsool, M.D.M.; Oglat, S.M.; Tbayshat, S.O.; Adnan, Z.I.; Alkhateeb, Y.A.; Aldamen, A.O.; Jain, H.; Nguyen, D.; et al. Stomach and duodenal ulcer as a cause of death in patients with cancer: A cohort study. Int. J. Emerg. Med. 2024, 17, 199. [Google Scholar] [CrossRef] [PubMed]
- Stojadinovic, M.; Stojadinovic, M.; Jankovic, S. Modifications to Prostate Cancer Diagnosis following COVID-19 and Following Models. Oncol. Res. Treat. 2025, 48, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Bril, F.; Elbert, A. Metabolic dysfunction-associated steatotic liver disease and urinary system cancers: Mere coincidence or reason for concern? Metabolism 2025, 162, 156066. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Wei, Y.; Li, X.; Pan, J.; Fang, B.; Zhang, T.; Lu, Y.; Ye, D.; Zhu, Y. From biology to the clinic—Exploring liver metastasis in prostate cancer. Nat. Rev. Urol. 2024, 21, 593–614. [Google Scholar] [CrossRef] [PubMed]
- Atci, T.; Ak, D.Y.; Baykal, C. Evaluation of secondary malignancies in a large series of mycosis fungoides. An. Bras. Dermatol. 2024, 99, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Heaf, J.G.; Hansen, A.; Laier, G.H. Quantification of cancer risk in glomerulonephritis. BMC Nephrol. 2018, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.S.; Wong, C.F.; Abraham, K.A. Membrano-proliferative glomerulonephritis associated with metastatic prostate carcinoma--should immunosuppressive therapy be considered? Nephrol. Dial. Transplant. 2008, 23, 777. [Google Scholar] [CrossRef]
- Wang, Q.; Pang, B.; Bucci, J.; Jiang, J.; Li, Y. The emerging role of extracellular vesicles and particles in prostate cancer diagnosis, and risk stratification. Biochim. Biophys. Acta Rev. Cancer 2024, 1879, 189210. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, G.; Wei, D.; Zhang, H.; Iliuk, A.; Xie, Z.; Gu, Y.; Gu, Z.; Zhang, Y.; Zhu, Y. One-Pot Sequential Enrichment of Urinary Extracellular Vesicle and miRNAs Identifies a Noninvasive Biomarker Panel for Prostate Cancer Diagnosis. Anal. Chem. 2024, 96, 19670–19677. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yu, C.; Jiang, K.; Yang, G.; Yang, S.; Tan, S.; Li, T.; Liang, H.; He, Q.; Wei, F.; et al. Unveiling potential: Urinary exosomal mRNAs as non-invasive biomarkers for early prostate cancer diagnosis. BMC Urol. 2024, 24, 163. [Google Scholar] [CrossRef] [PubMed]
- Jain, G.; Das, P.; Ranjan, P.; Neha; Valderrama, F.; Cieza-Borrella, C. Urinary extracellular vesicles miRNA-A new era of prostate cancer biomarkers. Front. Genet. 2023, 14, 1065757. [Google Scholar] [CrossRef] [PubMed]
- Brokāne, A.; Bajo-Santos, C.; Zayakin, P.; Belovs, A.; Jansons, J.; Lietuvietis, V.; Martens-Uzunova, E.S.; Jenster, G.W.; Linē, A. Validation of potential RNA biomarkers for prostate cancer diagnosis and monitoring in plasma and urinary extracellular vesicles. Front. Mol. Biosci. 2023, 10, 1279854. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kang, S.J.; Lee, J.; Park, K.H.; Rhee, W.J. Isolation and Characterization of Urinary Extracellular Vesicles from Healthy Donors and Patients with Castration-Resistant Prostate Cancer. Int. J. Mol. Sci. 2022, 23, 7134. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrügger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | n | Prostate Cancer Diagnosis | Gleason Score (GS) | Age Range |
|---|---|---|---|---|
| Controls | 20 | NO | NA | 55.15 (33, 65, 27, 30, 51, 54, 57, 39, 52, 50, 69, 49, 18, 79, 73, 84, 68, 76, 76, 53) |
| GS 6 | 1 | YES | 6 | 72 |
| GS 7 | 5 | YES | 7 | 71.4 (65, 66, 71, 73, 82) |
| GS 8 | 9 | YES | 8 | 64.7 (67, 77, 60, 53, 69, 53, 73, 60, 70) |
| GS 9 | 4 | YES | 9 | 69.8 (65, 68, 78, 68) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lange, S.; Bernstein, D.E.; Dimov, N.; Puttaswamy, S.; Johnston, I.; Kraev, I.; Needham, S.R.; Vasdev, N.; Inal, J.M. Urinary Extracellular Vesicle Signatures as Biomarkers in Prostate Cancer Patients. Int. J. Mol. Sci. 2025, 26, 6895. https://doi.org/10.3390/ijms26146895
Lange S, Bernstein DE, Dimov N, Puttaswamy S, Johnston I, Kraev I, Needham SR, Vasdev N, Inal JM. Urinary Extracellular Vesicle Signatures as Biomarkers in Prostate Cancer Patients. International Journal of Molecular Sciences. 2025; 26(14):6895. https://doi.org/10.3390/ijms26146895
Chicago/Turabian StyleLange, Sigrun, Darryl Ethan Bernstein, Nikolay Dimov, Srinivasu Puttaswamy, Ian Johnston, Igor Kraev, Sarah R. Needham, Nikhil Vasdev, and Jameel M. Inal. 2025. "Urinary Extracellular Vesicle Signatures as Biomarkers in Prostate Cancer Patients" International Journal of Molecular Sciences 26, no. 14: 6895. https://doi.org/10.3390/ijms26146895
APA StyleLange, S., Bernstein, D. E., Dimov, N., Puttaswamy, S., Johnston, I., Kraev, I., Needham, S. R., Vasdev, N., & Inal, J. M. (2025). Urinary Extracellular Vesicle Signatures as Biomarkers in Prostate Cancer Patients. International Journal of Molecular Sciences, 26(14), 6895. https://doi.org/10.3390/ijms26146895