
Kaempferol Suppresses the Activation of Mast Cells by Modulating the Expression of FcεRI and SHIP1

 , , , , and
, , , , and 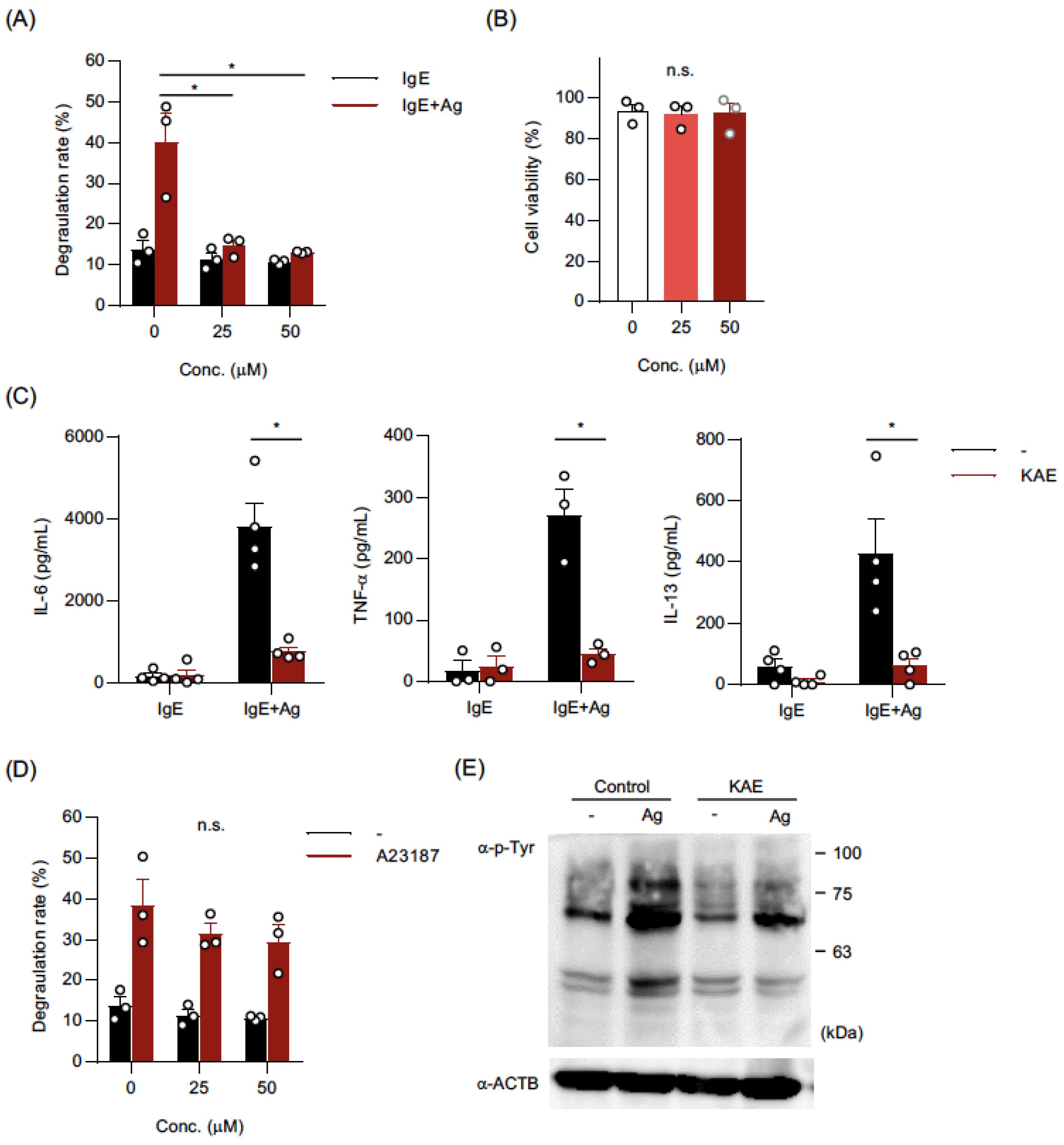{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Kaempferol Suppressed IgE-Mediated Activation of BMMCs
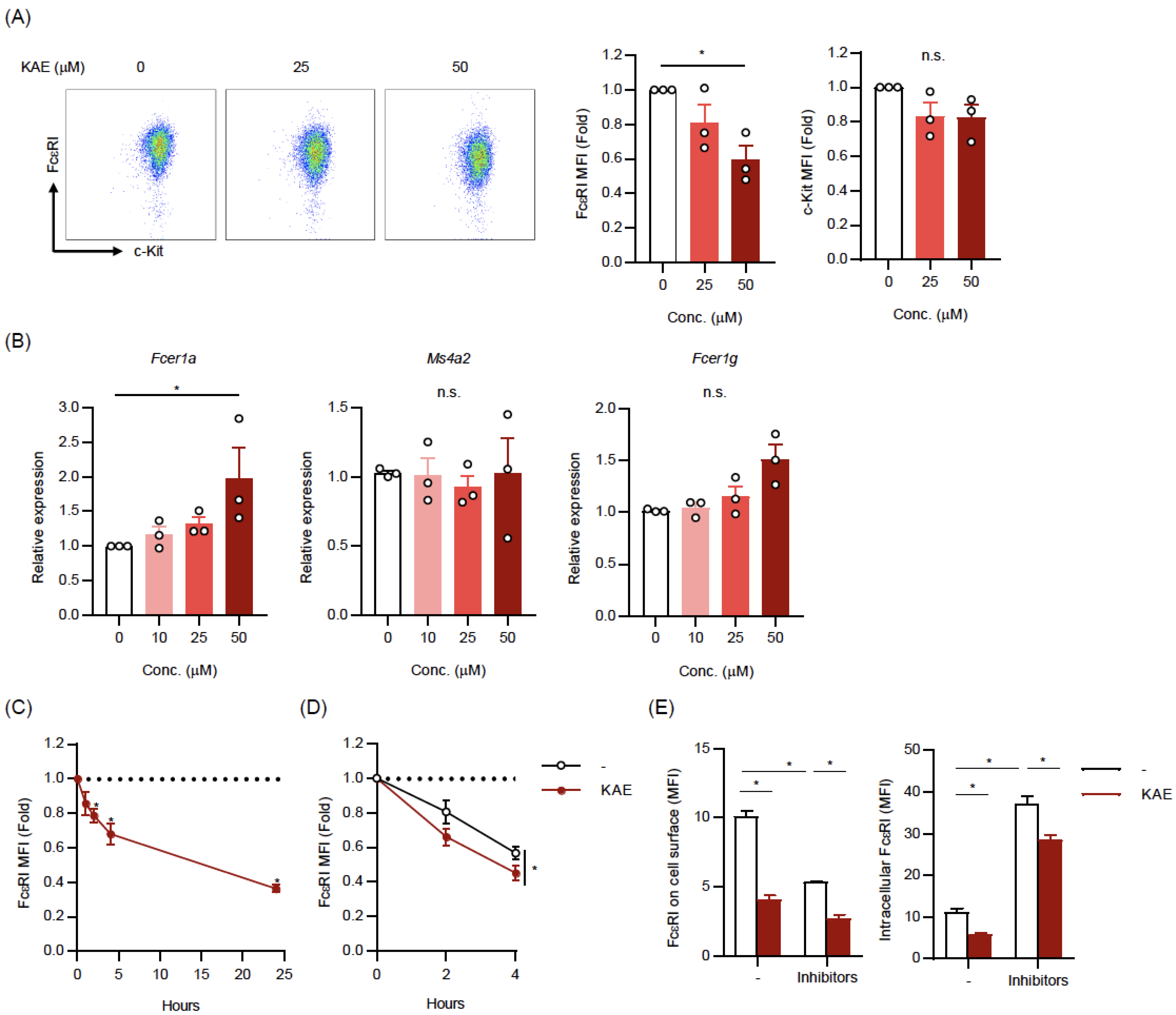
2.2. Kaempferol Reduced the Cell Surface Levels of FcεRI on BMMCs
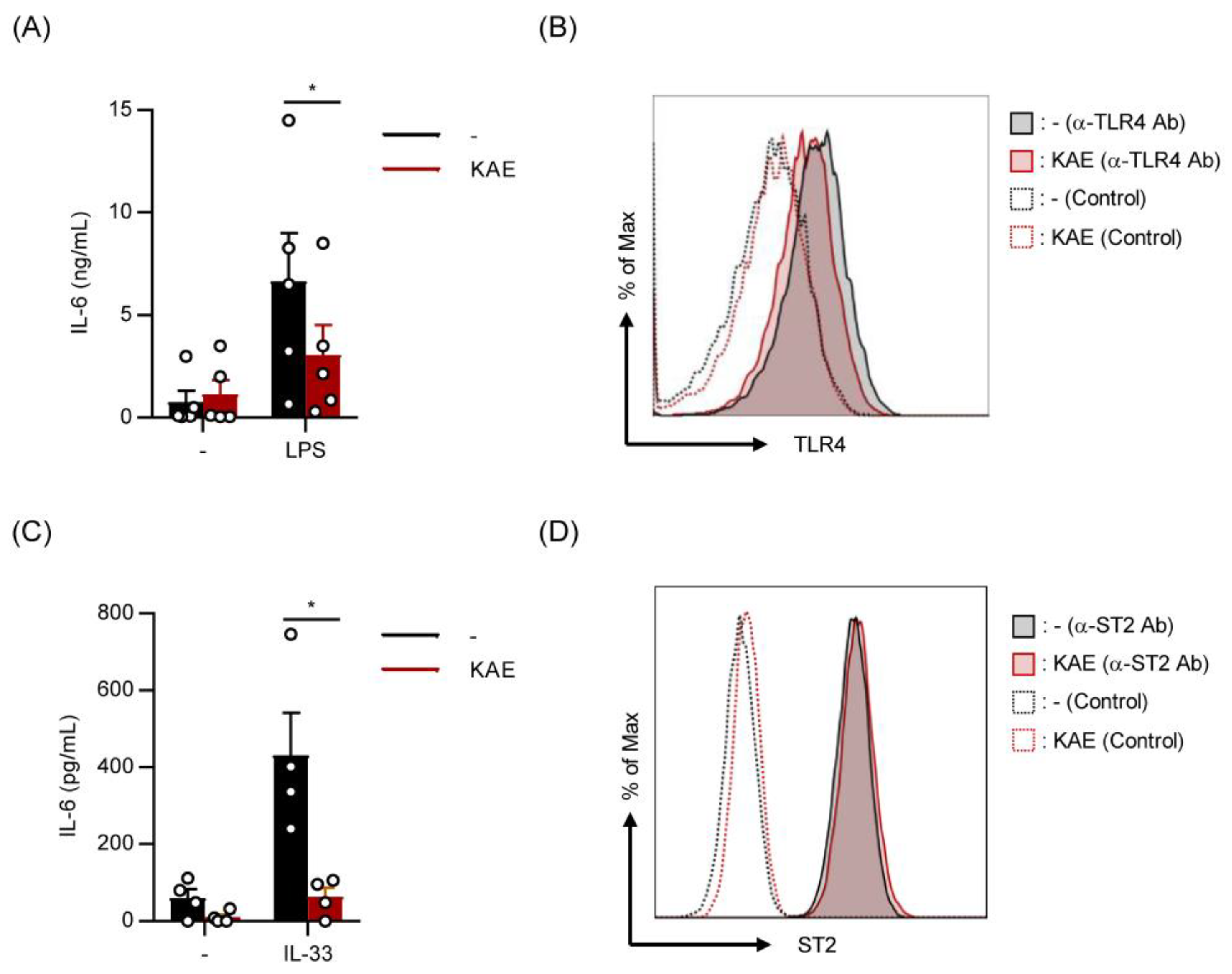
2.3. Kaempferol Suppressed LPS- and IL-33-Induced IL-6 Production without Affecting Cell Surface Levels of TLR4 and ST2 on BMMCs
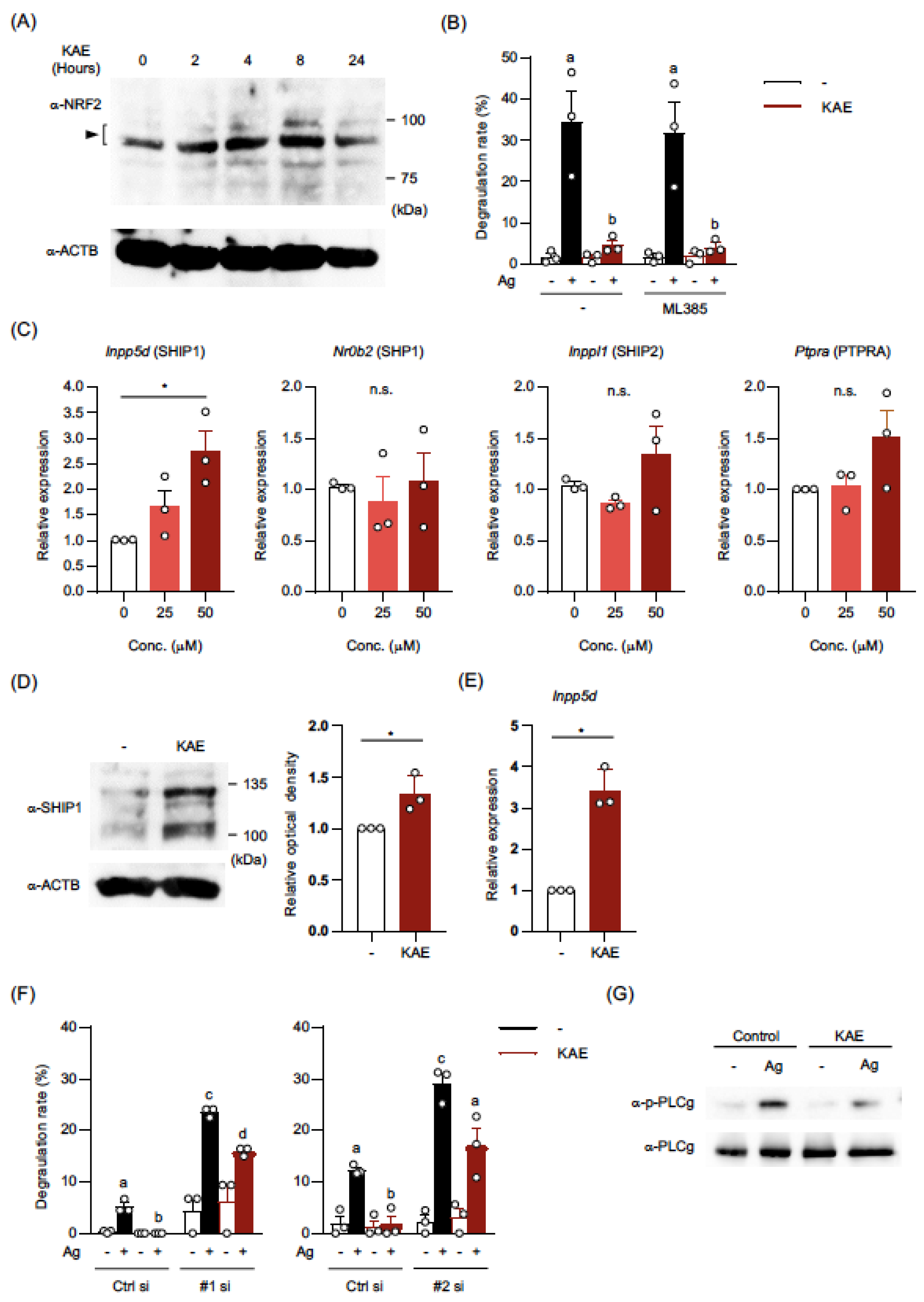
2.4. Kaempferol Treatment Activated NRF2 in BMMCs
2.5. The Levels of mRNA and Protein in SHIP1 were Upregulated in Kaempferol-Treated BMMCs
3. Discussion
4. Materials and Methods
4.1. Mice and Cells
4.2. Activation of MCs
4.3. ELISA
4.4. Western Blot Analysis
4.5. Flow Cytometry
4.6. Quantification of mRNA Using Real-Time PCR
4.7. siRNA Transfection
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maeda-Yamamoto, M.; Inagaki, N.; Kitaura, J.; Chikumoto, T.; Kawahara, H.; Kawakami, Y.; Sano, M.; Miyase, T.; Tachibana, H.; Nagai, H.; et al. O-methylated catechins from tea leaves inhibit multiple protein kinases in mast cells. J. Immunol. 2004, 172, 4486–4492. [Google Scholar] [CrossRef] [PubMed]
- Tokura, T.; Nakano, N.; Ito, T.; Matsuda, H.; Nagasako-Akazome, Y.; Kanda, T.; Ikeda, M.; Okumura, K.; Ogawa, H.; Nishiyama, C. Inhibitory effect of polyphenol-enriched apple extracts on mast cell degranulation in vitro targeting the binding between IgE and FcepsilonRI. Biosci. Biotechnol. Biochem. 2005, 69, 1974–1977. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Bae, J.Y.; Park, S.H.; Kim, Y.H.; Park, J.H.; Kang, Y.H. Resveratrol inhibits IgE-mediated basophilic mast cell degranulation and passive cutaneous anaphylaxis in mice. J. Nutr. 2013, 143, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, L.; Jiang, J.; Li, J.; Xu, C.; Jin, S.; Zhu, L.; Yan, G. Pterostilbene Inhibits FcεRI Signaling through Activation of the LKB1/AMPK Pathway in Allergic Response. J. Agric. Food Chem. 2020, 68, 3456–3465. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Hu, S.; Ge, S.; Jia, M.; Wang, N. Resveratrol inhibits MRGPRX2-mediated mast cell activation via Nrf2 pathway. Int. Immunopharmacol. 2021, 93, 107426. [Google Scholar] [CrossRef]
- Oh, H.A.; Han, N.R.; Kim, M.J.; Kim, H.M.; Jeong, H.J. Evaluation of the effect of kaempferol in a murine allergic rhinitis model. Eur. J. Pharm. 2013, 718, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Park, S.H.; Choi, Y.J.; Kim, Y.H.; Antika, L.D.; Habibah, N.U.; Kang, M.K.; Kang, Y.H. Dietary Compound Kaempferol Inhibits Airway Thickening Induced by Allergic Reaction in a Bovine Serum Albumin-Induced Model of Asthma. Int. J. Mol. Sci. 2015, 16, 29980–29995. [Google Scholar] [CrossRef]
- Cao, J.; Li, C.; Ma, P.; Ding, Y.; Gao, J.; Jia, Q.; Zhu, J.; Zhang, T. Effect of kaempferol on IgE-mediated anaphylaxis in C57BL/6 mice and LAD2 cells. Phytomedicine 2020, 79, 153346. [Google Scholar] [CrossRef]
- Kim, M.; Lim, S.J.; Kang, S.W.; Um, B.H.; Nho, C.W. Aceriphyllum rossii extract and its active compounds, quercetin and kaempferol inhibit IgE-mediated mast cell activation and passive cutaneous anaphylaxis. J. Agric. Food. Chem. 2014, 62, 3750–3758. [Google Scholar] [CrossRef]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.; Ha, J.H.; Lee, M.G.; Kim, J.E.; et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef]
- Lee, E.J.; Ji, G.E.; Sung, M.K. Quercetin and kaempferol suppress immunoglobulin E-mediated allergic inflammation in RBL-2H3 and Caco-2 cells. Inflamm. Res. 2010, 59, 847–854. [Google Scholar] [CrossRef]
- Shim, S.Y.; Choi, J.S.; Byun, D.S. Kaempferol isolated from Nelumbo nucifera stamens negatively regulates FcepsilonRI expression in human basophilic KU812F cells. J. Microbiol. Biotechnol. 2009, 19, 155–160. [Google Scholar] [CrossRef]
- Kempuraj, D.; Madhappan, B.; Christodoulou, S.; Boucher, W.; Cao, J.; Papadopoulou, N.; Cetrulo, C.L.; Theoharides, T.C. Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. Br. J. Pharm. 2005, 145, 934–944. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Bounab, Y.; Getahun, A.; Cambier, J.C.; Daëron, M. Phosphatase regulation of immunoreceptor signaling in T cells, B cells and mast cells. Curr. Opin. Immunol. 2013, 25, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Egner, P.A.; Agyeman, A.S.; Visvanathan, K.; Groopman, J.D.; Chen, J.G.; Chen, T.Y.; Fahey, J.W.; Talalay, P. Keap1-nrf2 signaling: A target for cancer prevention by sulforaphane. Top. Curr. Chem. 2013, 329, 163–177. [Google Scholar] [CrossRef]
- Qin, S.; Hou, D.X. Multiple regulations of Keap1/Nrf2 system by dietary phytochemicals. Mol. Nutr. Food Res. 2016, 60, 1731–1755. [Google Scholar] [CrossRef]
- Hirose, E.; Matsushima, M.; Takagi, K.; Ota, Y.; Ishigami, K.; Hirayama, T.; Hayashi, Y.; Nakamura, T.; Hashimoto, N.; Imaizumi, K.; et al. Involvement of heme oxygenase-1 in kaempferol-induced anti-allergic actions in RBL-2H3 cells. Inflammation 2009, 32, 99–108. [Google Scholar] [CrossRef]
- Yao, H.; Sun, J.; Wei, J.; Zhang, X.; Chen, B.; Lin, Y. Kaempferol Protects Blood Vessels From Damage Induced by Oxidative Stress and Inflammation in Association With the Nrf2/HO-1 Signaling Pathway. Front. Pharm. 2020, 11, 1118. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Piao, H.; Jiang, J.; Jin, G.; Zheng, M.; Yang, J.; Jin, X.; Sun, T.; Choi, Y.H.; Li, L.; et al. Polydatin inhibits mast cell-mediated allergic inflammation by targeting PI3K/Akt, MAPK, NF-κB and Nrf2/HO-1 pathways. Sci. Rep. 2017, 7, 11895. [Google Scholar] [CrossRef]
- Hu, B.; Zhao, S.; Huang, M.; Ren, J. Nuclear factor E2 related factor (NRF2) inhibits mast cell- mediated allergic inflammation via SIRT4-mediated mitochondrial metabolism. Ann. Palliat. Med. 2020, 9, 3839–3847. [Google Scholar] [CrossRef] [PubMed]
- Xian, Z.; Jin, G.; Li, H.; Jiang, J.; Wang, C.; Zhu, L.; Jin, Z.; Li, L.; Piao, H.; Zheng, M.; et al. Imperatorin Suppresses Anaphylactic Reaction and IgE-Mediated Allergic Responses by Inhibiting Multiple Steps of FceRI Signaling in Mast Cells: IMP Alleviates Allergic Responses in PCA. Biomed. Res. Int. 2019, 2019, 7823761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chiang, Y.J.; Hodes, R.J.; Siraganian, R.P. Inactivation of c-Cbl or Cbl-b differentially affects signaling from the high affinity IgE receptor. J. Immunol. 2004, 173, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Gustin, S.E.; Thien, C.B.; Langdon, W.Y. Cbl-b is a negative regulator of inflammatory cytokines produced by IgE-activated mast cells. J. Immunol. 2006, 177, 5980–5989. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Lantz, C.S.; Oettgen, H.C.; Katona, I.M.; Fleming, T.; Miyajima, I.; Kinet, J.P.; Galli, S.J. IgE enhances mouse mast cell Fc(epsilon)RI expression in vitro and in vivo: Evidence for a novel amplification mechanism in IgE-dependent reactions. J. Exp. Med. 1997, 185, 663–672. [Google Scholar] [CrossRef]
- Huber, M.; Helgason, C.D.; Damen, J.E.; Liu, L.; Humphries, R.K.; Krystal, G. The src homology 2-containing inositol phosphatase (SHIP) is the gatekeeper of mast cell degranulation. Proc. Natl. Acad. Sci. USA 1998, 95, 11330–11335. [Google Scholar] [CrossRef]
- Kalesnikoff, J.; Baur, N.; Leitges, M.; Hughes, M.R.; Damen, J.E.; Huber, M.; Krystal, G. SHIP negatively regulates IgE + antigen-induced IL-6 production in mast cells by inhibiting NF-kappa B activity. J. Immunol. 2002, 168, 4737–4746. [Google Scholar] [CrossRef]
- An, H.; Xu, H.; Zhang, M.; Zhou, J.; Feng, T.; Qian, C.; Qi, R.; Cao, X. Src homology 2 domain-containing inositol-5-phosphatase 1 (SHIP1) negatively regulates TLR4-mediated LPS response primarily through a phosphatase activity- and PI-3K-independent mechanism. Blood 2005, 105, 4685–4692. [Google Scholar] [CrossRef]
- Hochdörfer, T.; Kuhny, M.; Zorn, C.N.; Hendriks, R.W.; Vanhaesebroeck, B.; Bohnacker, T.; Krystal, G.; Huber, M. Activation of the PI3K pathway increases TLR-induced TNF-α and IL-6 but reduces IL-1β production in mast cells. Cell Signal. 2011, 23, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Haddon, D.J.; Antignano, F.; Hughes, M.R.; Blanchet, M.R.; Zbytnuik, L.; Krystal, G.; McNagny, K.M. SHIP1 is a repressor of mast cell hyperplasia, cytokine production, and allergic inflammation in vivo. J. Immunol. 2009, 183, 228–236. [Google Scholar] [CrossRef]
- Plum, T.; Wang, X.; Rettel, M.; Krijgsveld, J.; Feyerabend, T.B.; Rodewald, H.R. Human Mast Cell Proteome Reveals Unique Lineage, Putative Functions, and Structural Basis for Cell Ablation. Immunity 2020, 52, 404–416.e405. [Google Scholar] [CrossRef]
- Jiang, J.Z.; Ye, J.; Jin, G.Y.; Piao, H.M.; Cui, H.; Zheng, M.Y.; Yang, J.S.; Che, N.; Choi, Y.H.; Li, L.C.; et al. Asiaticoside Mitigates the Allergic Inflammation by Abrogating the Degranulation of Mast Cells. J. Agric. Food Chem. 2017, 65, 8128–8135. [Google Scholar] [CrossRef]
- Ciolino, H.P.; Daschner, P.J.; Yeh, G.C. Dietary flavonols quercetin and kaempferol are ligands of the aryl hydrocarbon receptor that affect CYP1A1 transcription differentially. Biochem. J. 1999, 340, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Kasakura, K.; Nagata, K.; Miura, R.; Iida, M.; Nakaya, H.; Okada, H.; Arai, T.; Kawakami, Y.; Kawakami, T.; Yashiro, T.; et al. Cooperative Regulation of the Mucosal Mast Cell-Specific Protease Genes. J. Immunol. 2020, 204, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Meurer, S.K.; Neß, M.; Weiskirchen, S.; Kim, P.; Tag, C.G.; Kauffmann, M.; Huber, M.; Weiskirchen, R. Isolation of Mature (Peritoneum-Derived) Mast Cells and Immature (Bone Marrow-Derived) Mast Cell Precursors from Mice. PLoS ONE 2016, 11, e0158104. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Kasakura, K.; Fujigaki, I.; Kageyama, A.; Okumura, K.; Ogawa, H.; Yashiro, T.; Nishiyama, C. The effect of PU.1 knockdown on gene expression and function of mast cells. Sci. Rep. 2018, 8, 2005. [Google Scholar] [CrossRef] [PubMed]
- Ashikari, T.; Hachisu, M.; Nagata, K.; Ando, D.; Iizuka, Y.; Ito, N.; Ito, K.; Ikeda, Y.; Matsubara, H.; Yashiro, T.; et al. Salicylaldehyde Suppresses IgE-Mediated Activation of Mast Cells and Ameliorates Anaphylaxis in Mice. Int. J. Mol. Sci. 2022, 23, 8826. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagata, K.; Araumi, S.; Ando, D.; Ito, N.; Ando, M.; Ikeda, Y.; Takahashi, M.; Noguchi, S.; Yasuda, Y.; Nakano, N.; et al. Kaempferol Suppresses the Activation of Mast Cells by Modulating the Expression of FcεRI and SHIP1. Int. J. Mol. Sci. 2023, 24, 5997. https://doi.org/10.3390/ijms24065997
Nagata K, Araumi S, Ando D, Ito N, Ando M, Ikeda Y, Takahashi M, Noguchi S, Yasuda Y, Nakano N, et al. Kaempferol Suppresses the Activation of Mast Cells by Modulating the Expression of FcεRI and SHIP1. International Journal of Molecular Sciences. 2023; 24(6):5997. https://doi.org/10.3390/ijms24065997
Chicago/Turabian StyleNagata, Kazuki, Sanae Araumi, Daisuke Ando, Naoto Ito, Miki Ando, Yuki Ikeda, Miki Takahashi, Sakura Noguchi, Yayoi Yasuda, Nobuhiro Nakano, and et al. 2023. "Kaempferol Suppresses the Activation of Mast Cells by Modulating the Expression of FcεRI and SHIP1" International Journal of Molecular Sciences 24, no. 6: 5997. https://doi.org/10.3390/ijms24065997
APA StyleNagata, K., Araumi, S., Ando, D., Ito, N., Ando, M., Ikeda, Y., Takahashi, M., Noguchi, S., Yasuda, Y., Nakano, N., Ando, T., Hara, M., Yashiro, T., Hachisu, M., & Nishiyama, C. (2023). Kaempferol Suppresses the Activation of Mast Cells by Modulating the Expression of FcεRI and SHIP1. International Journal of Molecular Sciences, 24(6), 5997. https://doi.org/10.3390/ijms24065997