
Intrauterine Smoke Exposure, microRNA Expression during Human Lung Development, and Childhood Asthma

 , , , , , ,
, , , , , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Sample Characteristics
2.2. Differential Expression of miRNAs by IUS
2.3. Pathway Enrichment Analysis
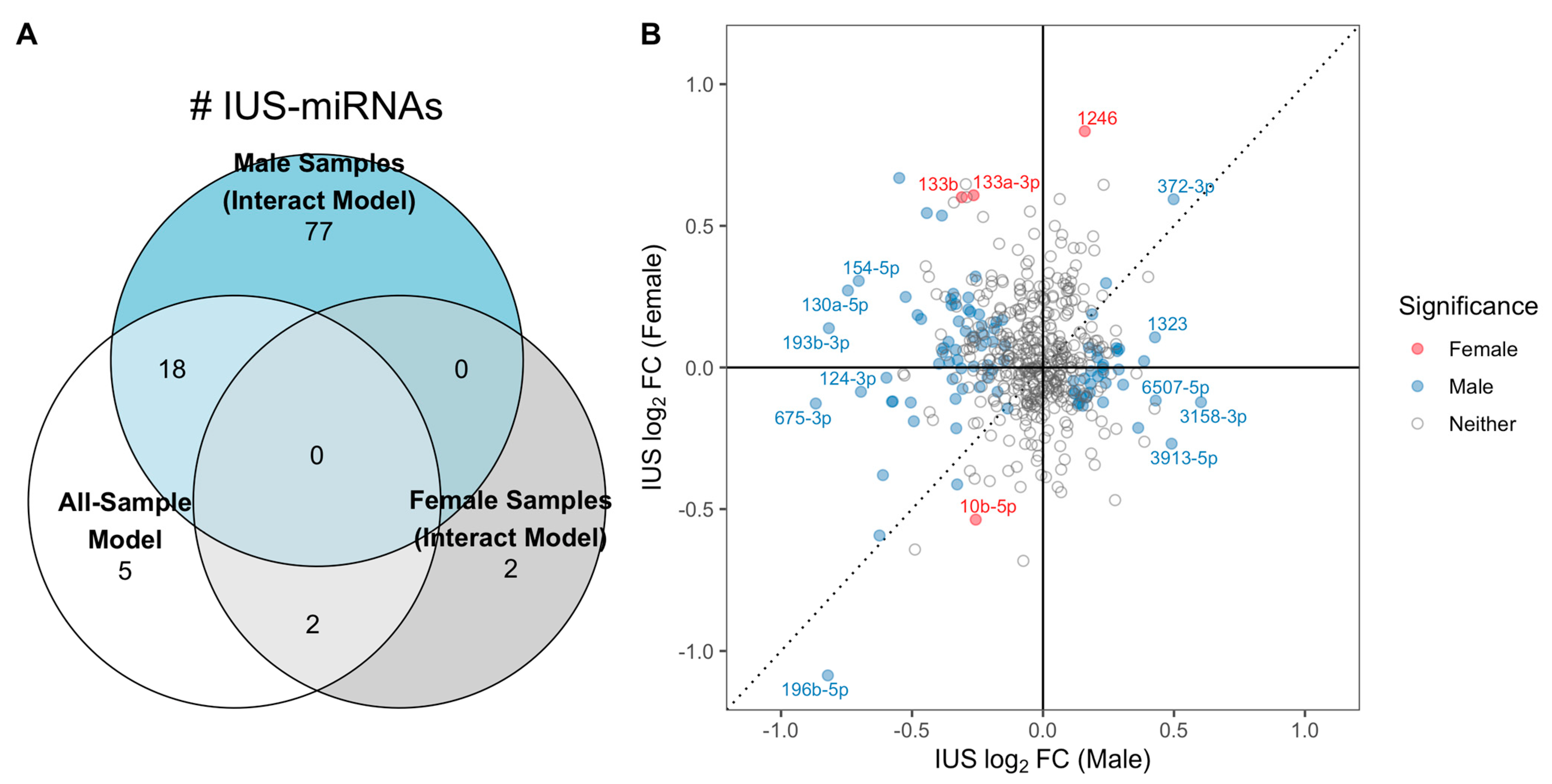
2.4. Sex-Specific Associations between IUS and miRNAs
2.5. Sex-Specific miRNA–mRNA Networks of IUS
2.6. Impact of IUS-Associated MicroRNAs on Asthma and Allergy Outcomes in a Childhood Asthma Cohort
3. Discussion
4. Materials and Methods
4.1. Fetal Lung Sample Acquisition and Metadata
4.2. miRNA Profiling
4.3. Differential miRNA Levels by IUS
4.4. Sex-Specific Effects of IUS
4.5. Predicted mRNA Targets and Pathway Analyses
4.6. miRNA–mRNA Correlations
4.7. IUS-miRNAs Associations with Asthma and Allergy Outcomes in a Childhood Asthma Cohort
4.8. Study Reproducibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sherman, C.B. Health effects of cigarette smoking. Clin. Chest Med. 1991, 12, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Benjamin-Garner, R.; Stotts, A. Impact of smoking exposure change on infant birth weight among a cohort of women in a prenatal smoking cessation study. Nicotine Tob. Res. 2013, 15, 685–692. [Google Scholar] [CrossRef]
- Tager, I.B.; Ngo, L.; Hanrahan, J.P. Maternal smoking during pregnancy. Effects on lung function during the first 18 months of life. Am. J. Respir. Crit. Care Med. 1995, 152, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, J.; Dockery, D.W.; Speizer, F.E. Maternal smoking during pregnancy as a predictor of lung function in children. Am. J. Epidemiol. 1994, 139, 1139–1152. [Google Scholar] [CrossRef] [PubMed]
- Moshammer, H.; Hoek, G.; Luttmann-Gibson, H.; Neuberger, M.A.; Antova, T.; Gehring, U.; Hruba, F.; Pattenden, S.; Rudnai, P.; Slachtova, H.; et al. Parental smoking and lung function in children: An international study. Am. J. Respir. Crit. Care Med. 2006, 173, 1255–1263. [Google Scholar] [CrossRef]
- Feleszko, W.; Ruszczynski, M.; Jaworska, J.; Strzelak, A.; Zalewski, B.M.; Kulus, M. Environmental tobacco smoke exposure and risk of allergic sensitisation in children: A systematic review and meta-analysis. Arch. Dis. Child 2014, 99, 985–992. [Google Scholar] [CrossRef]
- Kulig, M.; Luck, W.; Lau, S.; Niggemann, B.; Bergmann, R.; Klettke, U.; Guggenmoos-Holzmann, I.; Wahn, U. Effect of pre- and postnatal tobacco smoke exposure on specific sensitization to food and inhalant allergens during the first 3 years of life. Multicenter Allergy Study Group, Germany. Allergy 1999, 54, 220–228. [Google Scholar] [CrossRef]
- Lannero, E.; Wickman, M.; van Hage, M.; Bergstrom, A.; Pershagen, G.; Nordvall, L. Exposure to environmental tobacco smoke and sensitisation in children. Thorax 2008, 63, 172–176. [Google Scholar] [CrossRef]
- Raherison, C.; Penard-Morand, C.; Moreau, D.; Caillaud, D.; Charpin, D.; Kopferschmitt, C.; Lavaud, F.; Taytard, A.; Maesano, I.A. Smoking exposure and allergic sensitization in children according to maternal allergies. Ann. Allergy Asthma. Immunol. 2008, 100, 351–357. [Google Scholar] [CrossRef]
- Leung, D.Y.; Szefler, S.J.; Associate Editors of the JACI. In Utero smoke (IUS) exposure has been associated with increased prevalence of asthma and reduced lung function in healthy children. J. Allergy Clin. Immunol. 2010, 126, 481–482. [Google Scholar] [CrossRef]
- Goksör, E.; Amark, M.; Alm, B.; Gustafsson, P.M.; Wennergren, G. The impact of pre- and post-natal smoke exposure on future asthma and bronchial hyper-responsiveness. Acta Paediatr. 2007, 96, 1030–1035. [Google Scholar] [CrossRef]
- Gilliland, F.D.; Berhane, K.; Li, Y.F.; Rappaport, E.B.; Peters, J.M. Effects of early onset asthma and in utero exposure to maternal smoking on childhood lung function. Am. J. Respir. Crit. Care Med. 2003, 167, 917–924. [Google Scholar] [CrossRef]
- Bjerg, A.; Hedman, L.; Perzanowski, M.; Lundback, B.; Ronmark, E. A strong synergism of low birth weight and prenatal smoking on asthma in schoolchildren. Pediatrics 2011, 127, e905–e912. [Google Scholar] [CrossRef]
- Balte, P.; Karmaus, W.; Roberts, G.; Kurukulaaratchy, R.; Mitchell, F.; Arshad, H. Relationship between birth weight, maternal smoking during pregnancy and childhood and adolescent lung function: A path analysis. Respir. Med. 2016, 121, 13–20. [Google Scholar] [CrossRef]
- Wang, B.; Chen, H.; Chan, Y.L.; Wang, G.; Oliver, B.G. Why Do Intrauterine Exposure to Air Pollution and Cigarette Smoke Increase the Risk of Asthma? Front. Cell Dev. Biol. 2020, 8, 38. [Google Scholar] [CrossRef]
- Collins, M.H.; Moessinger, A.C.; Kleinerman, J.; Bassi, J.; Rosso, P.; Collins, A.M.; James, L.S.; Blanc, W.A. Fetal lung hypoplasia associated with maternal smoking: A morphometric analysis. Pediatr. Res. 1985, 19, 408–412. [Google Scholar] [CrossRef]
- Sekhon, H.S.; Jia, Y.; Raab, R.; Kuryatov, A.; Pankow, J.F.; Whitsett, J.A.; Lindstrom, J.; Spindel, E.R. Prenatal nicotine increases pulmonary alpha7 nicotinic receptor expression and alters fetal lung development in monkeys. J. Clin. Investig. 1999, 103, 637–647. [Google Scholar] [CrossRef]
- Sekhon, H.S.; Keller, J.A.; Benowitz, N.L.; Spindel, E.R. Prenatal nicotine exposure alters pulmonary function in newborn rhesus monkeys. Am. J. Respir. Crit. Care Med. 2001, 164, 989–994. [Google Scholar] [CrossRef]
- Sandberg, K.; Poole, S.D.; Hamdan, A.; Arbogast, P.; Sundell, H.W. Altered lung development after prenatal nicotine exposure in young lambs. Pediatr. Res. 2004, 56, 432–439. [Google Scholar] [CrossRef]
- Barker, D.J. Fetal growth and adult disease. Br. J. Obstet. Gynaecol. 1992, 99, 275–276. [Google Scholar] [CrossRef]
- Joss-Moore, L.A.; Albertine, K.H.; Lane, R.H. Epigenetics and the developmental origins of lung disease. Mol. Genet. Metab. 2011, 104, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Sohn, K.A.; Kim, J.H. MicroRNA-centric measurement improves functional enrichment analysis of co-expressed and differentially expressed microRNA clusters. BMC Genomics 2012, 13 (Suppl. S7), S17. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.C.; Tavazoie, M.; Doetsch, F. Stem cells: From epigenetics to microRNAs. Neuron 2005, 46, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Karp, X.; Ambros, V. Developmental biology. Encountering microRNAs in cell fate signaling. Science 2005, 310, 1288–1289. [Google Scholar] [CrossRef]
- Chen, C.; Tan, R.; Wong, L.; Fekete, R.; Halsey, J. Quantitation of microRNAs by real-time RT-qPCR. Methods Mol. Biol. 2011, 687, 113–134. [Google Scholar]
- Garzon, R.; Fabbri, M.; Cimmino, A.; Calin, G.A.; Croce, C.M. MicroRNA expression and function in cancer. Trends Mol. Med. 2006, 12, 580–587. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- Calin, G.A.; Liu, C.G.; Sevignani, C.; Ferracin, M.; Felli, N.; Dumitru, C.D.; Shimizu, M.; Cimmino, A.; Zupo, S.; Dono, M.; et al. MicroRNA profiling reveals distinct signatures in B cell chronic lymphocytic leukemias. Proc. Natl. Acad. Sci. USA 2004, 101, 11755–11760. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Ji, R.; Cheng, Y.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of MicroRNA in vascular neointimal lesion formation. Circ. Res. 2007, 100, 1579–1588. [Google Scholar] [CrossRef]
- Lu, Y.; Thomson, J.M.; Wong, H.Y.; Hammond, S.M.; Hogan, B.L. Transgenic over-expression of the microRNA miR-17-92 cluster promotes proliferation and inhibits differentiation of lung epithelial progenitor cells. Dev. Biol. 2007, 310, 442–453. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Wang, Y.; Zhang, H.; Weng, T.; Baviskar, P.; Guo, Y.; Gou, D.; Liu, L. MicroRNA-127 modulates fetal lung development. Physiol. Genomics 2009, 37, 268–278. [Google Scholar] [CrossRef]
- Boateng, E.; Krauss-Etschmann, S. miRNAs in Lung Development and Diseases. Int. J. Mol. Sci. 2020, 21, 2765. [Google Scholar] [CrossRef]
- Willinger, C.M.; Rong, J.; Tanriverdi, K.; Courchesne, P.L.; Huan, T.; Wasserman, G.A.; Lin, H.; Dupuis, J.; Joehanes, R.; Jones, M.R.; et al. MicroRNA Signature of Cigarette Smoking and Evidence for a Putative Causal Role of MicroRNAs in Smoking-Related Inflammation and Target Organ Damage. Circ. Cardiovasc. Genet. 2017, 10, e001678. [Google Scholar] [CrossRef]
- Dehmel, S.; Nathan, P.; Bartel, S.; El-Merhie, N.; Scherb, H.; Milger, K.; John-Schuster, G.; Yildirim, A.O.; Hylkema, M.; Irmler, M.; et al. Intrauterine smoke exposure deregulates lung function, pulmonary transcriptomes, and in particular insulin-like growth factor (IGF)-1 in a sex-specific manner. Sci. Rep. 2018, 8, 7547. [Google Scholar] [CrossRef]
- Vyhlidal, C.A.; Riffel, A.K.; Haley, K.J.; Sharma, S.; Dai, H.; Tantisira, K.G.; Weiss, S.T.; Leeder, J.S. Cotinine in human placenta predicts induction of gene expression in fetal tissues. Drug Metab. Dispos. 2013, 41, 305–311. [Google Scholar] [CrossRef]
- Sharma, S.; Tantisira, K.; Carey, V.; Murphy, A.J.; Lasky-Su, J.; Celedon, J.C.; Lazarus, R.; Klanderman, B.; Rogers, A.; Soto-Quiros, M.; et al. A role for Wnt signaling genes in the pathogenesis of impaired lung function in asthma. Am. J. Respir. Crit. Care Med. 2010, 181, 328–336. [Google Scholar] [CrossRef]
- Lin, N.W.; Liu, C.; Yang, I.V.; Maier, L.A.; DeMeo, D.L.; Wood, C.; Ye, S.; Cruse, M.H.; Smith, V.L.; Vyhlidal, C.A.; et al. Sex-Specific Differences in MicroRNA Expression During Human Fetal Lung Development. Front. Genet. 2022, 13, 762834. [Google Scholar] [CrossRef]
- Rodriguez Bauza, D.E.; Silveyra, P. Asthma, atopy, and exercise: Sex differences in exercise-induced bronchoconstriction. Exp. Biol. Med. 2021, 246, 1400–1409. [Google Scholar] [CrossRef]
- Pignataro, F.S.; Bonini, M.; Forgione, A.; Melandri, S.; Usmani, O.S. Asthma and gender: The female lung. Pharmacol. Res. 2017, 119, 384–390. [Google Scholar] [CrossRef]
- Svanes, C.; Omenaas, E.; Jarvis, D.; Chinn, S.; Gulsvik, A.; Burney, P. Parental smoking in childhood and adult obstructive lung disease: Results from the European Community Respiratory Health Survey. Thorax 2004, 59, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Iber, D. The control of lung branching morphogenesis. Curr. Top. Dev. Biol. 2021, 143, 205–237. [Google Scholar] [PubMed]
- Cushing, L.; Jiang, Z.; Kuang, P.; Lü, J. The roles of microRNAs and protein components of the microRNA pathway in lung development and diseases. Am. J. Respir. Cell Mol. Biol. 2015, 52, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Sly, P.D.; Holt, P.G. Role of innate immunity in the development of allergy and asthma. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 127–131. [Google Scholar] [CrossRef]
- Lee, A.; Leon Hsu, H.H.; Mathilda Chiu, Y.H.; Bose, S.; Rosa, M.J.; Kloog, I.; Wilson, A.; Schwartz, J.; Cohen, S.; Coull, B.A.; et al. Prenatal fine particulate exposure and early childhood asthma: Effect of maternal stress and fetal sex. J. Allergy Clin. Immunol. 2018, 141, 1880–1886. [Google Scholar] [CrossRef]
- Noakes, P.S.; Thomas, R.; Lane, C.; Mori, T.A.; Barden, A.E.; Devadason, S.G.; Prescott, S.L. Association of maternal smoking with increased infant oxidative stress at 3 months of age. Thorax 2007, 62, 714–717. [Google Scholar] [CrossRef]
- Macnee, W.; Rahman, I. Oxidants and antioxidants as therapeutic targets in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1999, 160, S58–S65. [Google Scholar] [CrossRef]
- Noakes, P.S.; Hale, J.; Thomas, R.; Lane, C.; Devadason, S.G.; Prescott, S.L. Maternal smoking is associated with impaired neonatal toll-like-receptor-mediated immune responses. Eur. Respir. J. 2006, 28, 721–729. [Google Scholar] [CrossRef]
- Strzelak, A.; Ratajczak, A.; Adamiec, A.; Feleszko, W. Tobacco Smoke Induces and Alters Immune Responses in the Lung Triggering Inflammation, Allergy, Asthma and Other Lung Diseases: A Mechanistic Review. Int. J. Environ. Res. Public Health 2018, 15, 1033. [Google Scholar] [CrossRef]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef]
- Schoepp, D.D.; Jane, D.E.; Monn, J.A. Pharmacological agents acting at subtypes of metabotropic glutamate receptors. Neuropharmacology 1999, 38, 1431–1476. [Google Scholar] [CrossRef]
- Patel, B.S.; Ravix, J.; Pabelick, C.; Prakash, Y.S. Class C GPCRs in the airway. Curr. Opin. Pharmacol. 2020, 51, 19–28. [Google Scholar] [CrossRef]
- Cross, A.J.; Anthenelli, R.; Li, X. Metabotropic Glutamate Receptors 2 and 3 as Targets for Treating Nicotine Addiction. Biol. Psychiatry 2018, 83, 947–954. [Google Scholar] [CrossRef]
- Niepiekło-Miniewska, W.; Matusiak, Ł.; Narbutt, J.; Lesiak, A.; Kuna, P.; Wiśniewski, A.; Piekarska, K.; Nowak, I.; Kuśnierczyk, P. Synergy of endoplasmic reticulum aminopeptidase 1 and 2 (ERAP1 and ERAP2) polymorphisms in atopic dermatitis: Effects on disease prevalence. Hum. Immunol. 2021, 82, 121–123. [Google Scholar] [CrossRef]
- He, H.; Wang, H.; Pei, F.; Jiang, M. MiR-543 Regulates the Development of Chronic Obstructive Pulmonary Disease by Targeting Interleukin-33. Clin. Lab. 2018, 64, 1199–1205. [Google Scholar] [CrossRef]
- Chan, B.C.L.; Lam, C.W.K.; Tam, L.S.; Wong, C.K. IL33: Roles in Allergic Inflammation and Therapeutic Perspectives. Front. Immunol. 2019, 10, 364. [Google Scholar] [CrossRef]
- Zimmermann, N.; Doepker, M.P.; Witte, D.P.; Stringer, K.F.; Fulkerson, P.C.; Pope, S.M.; Brandt, E.B.; Mishra, A.; King, N.E.; Nikolaidis, N.M.; et al. Expression and regulation of small proline-rich protein 2 in allergic inflammation. Am. J. Respir. Cell Mol. Biol. 2005, 32, 428–435. [Google Scholar] [CrossRef]
- Zakeri, A.; Russo, M. Dual Role of Toll-like Receptors in Human and Experimental Asthma Models. Front. Immunol. 2018, 9, 1027. [Google Scholar] [CrossRef]
- Bersimbaev, R.; Aripova, A.; Bulgakova, O.; Kussainova, A.; Akparova, A.; Izzotti, A. The Plasma Levels of hsa-miR-19b-3p, hsa-miR-125b-5p, and hsamiR- 320c in Patients with Asthma, COPD and Asthma-COPD Overlap Syndrome (ACOS). Microrna 2021, 10, 130–138. [Google Scholar] [CrossRef]
- Wang, W.; Sinha, A.; Lutter, R.; Yang, J.; Ascoli, C.; Sterk, P.J.; Nemsick, N.K.; Perkins, D.L.; Finn, P.W. Analysis of Exosomal MicroRNA Dynamics in Response to Rhinovirus Challenge in a Longitudinal Case-Control Study of Asthma. Viruses 2022, 14, 2444. [Google Scholar] [CrossRef]
- Rajput, C.; Walsh, M.P.; Eder, B.N.; Metitiri, E.E.; Popova, A.P.; Hershenson, M.B. Rhinovirus infection induces distinct transcriptome profiles in polarized human macrophages. Physiol. Genom. 2018, 50, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Cardenas, A.; Rifas-Shiman, S.L.; Hivert, M.F.; Gold, D.R.; Platts-Mills, T.A.; Lin, X.; Oken, E.; Baccarelli, A.A.; Litonjua, A.A.; et al. Epigenome-wide association study of total serum immunoglobulin E in children: A life course approach. Clin. Epigenetics 2018, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, C.; Vatrella, A.; Gallelli, L.; Lombardo, N.; Sciacqua, A.; Savino, R.; Pelaia, G. Role of p38 Mitogen-Activated Protein Kinase in Asthma and COPD: Pathogenic Aspects and Potential Targeted Therapies. Drug Des. Devel. Ther. 2021, 15, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Carascossa, S.; Dudek, P.; Cenni, B.; Briand, P.A.; Picard, D. CARM1 mediates the ligand-independent and tamoxifen-resistant activation of the estrogen receptor alpha by cAMP. Genes Dev. 2010, 24, 708–719. [Google Scholar] [CrossRef]
- Koo, H.K.; Morrow, J.; Kachroo, P.; Tantisira, K.; Weiss, S.T.; Hersh, C.P.; Silverman, E.K.; DeMeo, D.L. Sex-specific associations with DNA methylation in lung tissue demonstrate smoking interactions. Epigenetics 2021, 16, 692–703. [Google Scholar] [CrossRef]
- Kho, A.T.; Chhabra, D.; Sharma, S.; Qiu, W.; Carey, V.J.; Gaedigk, R.; Vyhlidal, C.A.; Leeder, J.S.; Tantisira, K.J.; Weiss, S.T. Age, Sexual Dimorphism, and Disease Associations in the Developing Human Fetal Lung Transcriptome. Am. J. Respir. Cell Mol. Biol. 2016, 54, 814–821. [Google Scholar] [CrossRef]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef]
- Russell, P.H.; Vestal, B.; Shi, W.; Rudra, P.D.; Dowell, R.; Radcliffe, R.; Saba, L.; Kechris, K. miR-MaGiC improves quantification accuracy for small RNA-seq. BMC Res. Notes 2018, 11, 296. [Google Scholar] [CrossRef]
- Waardenberg, A.J.; Field, M.A. consensusDE: An R package for assessing consensus of multiple RNA-seq algorithms with RUV correction. PeerJ 2019, 7, e8206. [Google Scholar] [CrossRef]
- Kern, F.; Fehlmann, T.; Solomon, J.; Schwed, L.; Grammes, N.; Backes, C.; Van Keuren-Jensen, K.; Craig, D.W.; Meese, E.; Keller, A. miEAA 2.0: Integrating multi-species microRNA enrichment analysis and workflow management systems. Nucleic Acids Res. 2020, 48, W521–W528. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Singh, A.; Shannon, C.P.; Gautier, B.; Rohart, F.; Vacher, M.; Tebbutt, S.J.; Le Cao, K.A. DIABLO: An integrative approach for identifying key molecular drivers from multi-omics assays. Bioinformatics 2019, 35, 3055–3062. [Google Scholar] [CrossRef]
- Celedon, J.C.; Soto-Quiros, M.E.; Silverman, E.K.; Hanson, L.; Weiss, S.T. Risk factors for childhood asthma in Costa Rica. Chest 2001, 120, 785–790. [Google Scholar] [CrossRef]
- Tse, S.M.; Gold, D.R.; Sordillo, J.E.; Hoffman, E.B.; Gillman, M.W.; Rifas-Shiman, S.L.; Fuhlbrigge, A.L.; Tantisira, K.G.; Weiss, S.T.; Litonjua, A.A. Diagnostic accuracy of the bronchodilator response in children. J. Allergy Clin. Immunol. 2013, 132, 554–559 e555. [Google Scholar] [CrossRef]
- Tiwari, A.; Li, J.; Kho, A.T.; Sun, M.; Lu, Q.; Weiss, S.T.; Tantisira, K.G.; McGeachie, M.J. COPD-associated miR-145-5p is downregulated in early-decline FEV(1) trajectories in childhood asthma. J. Allergy Clin. Immunol. 2021, 147, 2181–2190. [Google Scholar] [CrossRef]
- Rozowsky, J.; Kitchen, R.R.; Park, J.J.; Galeev, T.R.; Diao, J.; Warrell, J.; Thistlethwaite, W.; Subramanian, S.L.; Milosavljevic, A.; Gerstein, M. exceRpt: A Comprehensive Analytic Platform for Extracellular RNA Profiling. Cell Syst. 2019, 8, 352–357.e3. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No IUS Exposure | IUS-Exposure | All Samples | |
|---|---|---|---|
| N | 159 | 139 | 298 |
| Age (dpc) * | 87.0 (76.0, 98.0) | 89.0 (76.0, 96.0) | 87.0 (76.0, 96.0) |
| Placental Cotinine (ng/g) | 0.0 (0.0, 0.8) | 53.5 (27.1, 79.6) | 2.2 (0.0, 50.4) |
| Sex (Male) | 89 (56.0%) | 77 (55.4%) | 166 (55.7%) |
| miRNA | log2FC | lfSE | q-Val | Top Microarray Correlations | |
|---|---|---|---|---|---|
| Increases with IUS | hsa-miR-372-3p | 0.54 | 0.16 | 0.031 | VEGFC+, SLC27A6+, KRTAP20-2+, BACH2+, CYP4F2+, EHD2+, TOMM40L+, POMGNT2+ |
| hsa-miR-1246 | 0.45 | 0.15 | 0.059 | PTTG1+, HIST1H2BG+, USP34−, CNBP+, HIST2H2BE+, SUOX−, CYP51A1+, YTHDF3− | |
| hsa-miR-1323 | 0.28 | 0.07 | 0.003 | GNPAT+, PSMD13+, ORMDL3+, CAMLG+, ELOA+, FBXO5+, SERPINB1+, MDH1+ | |
| hsa-miR-431-5p | 0.27 | 0.08 | 0.031 | MUC1+, PNMA5−, SELENBP1+, KDM1B+, SERPINB2−, SLCO2B1+, CXCL16+, HIST1H3E− | |
| hsa-miR-320d | 0.19 | 0.07 | 0.059 | OTUD7A−, INSYN2A+, ADCY5+, ARSD+, AMN1+, AOC3+, NAALADL1+, BAIAP2L1+ | |
| hsa-miR-3613-5p | 0.18 | 0.07 | 0.072 | MCAM+, ABCF2+, SREK1IP1−, TTC38+, SLC25A11+, PRPS1+, RCL1+, NMT2+ | |
| hsa-miR-543 | 0.17 | 0.06 | 0.099 | TDRD12+, ERAP2−, OR2M5−, ACSL4−, AREG−, HCRTR2−, MAP7−, RBM11− | |
| hsa-miR-423-5p | 0.16 | 0.05 | 0.031 | CLOCK+, ENTPD7+, JRK−, IBA57+, LTV1+, MYL3+, CMSS1+, ADCY5+ | |
| hsa-let-7e-5p | 0.14 | 0.04 | 0.036 | GLMN+, ELSPBP1−, ZNF45−, CRH−, KCNH5−, INPP5A+, PDE8A+, CCDC9B+ | |
| Decreases with IUS | hsa-miR-148a-3p | −0.12 | 0.04 | 0.059 | HIST1H2BH−, OR52N2+, CATSPER3+, HIST1H2AL−, MMP16−, HIST1H2BO−, FOXC1−, HIST1H2BJ− |
| hsa-miR-200a-3p | −0.13 | 0.04 | 0.050 | PPP1R1A−, CCL2−, AKIRIN1−, PXK−, DGLUCY−, SNAI1−, S100A3+, GPR148+ | |
| hsa-miR-101-3p | −0.14 | 0.04 | 0.040 | EXOC7−, FRRS1L+, CAD−, NFATC1−, MTOR−, PPP4C−, WDR76−, CEP85L− | |
| hsa-miR-125b-5p | −0.17 | 0.05 | 0.040 | UBE4B−, VPS39−, KIF20A−, NCAPD2−, ST3GAL6−, PDLIM1−, FOXM1−, KLHDC3− | |
| hsa-miR-532-5p | −0.17 | 0.06 | 0.056 | EIF4G1−, SIPA1L2−, TRIM8−, NUP98−, ILF3−, RASGEF1A+, MTOR−, TTLL5− | |
| hsa-miR-221-5p | −0.17 | 0.07 | 0.092 | KHSRP−, KIRREL1−, PFAS−, SEC24D−, FMNL3−, ADGRA2−, SPTLC1+, EHD2− | |
| hsa-miR-324-5p | −0.17 | 0.06 | 0.076 | CHRNA1−, TSPAN15−, KCNN2−, ALDH18A1−, CHGB−, IL4R−, DCTN5−, PSMD13− | |
| hsa-miR-584-5p | −0.19 | 0.07 | 0.059 | DHRS9+, DLL4−, DHX32+, SAP130−, SDSL+, DAB2IP−, GABPA+, AGO1− | |
| hsa-miR-34a-5p | −0.19 | 0.06 | 0.040 | CDH7−, SCGB1D4+, GCN1−, THY1−, PSG4+, MRPL20−, MRPL20−, LZIC+ | |
| hsa-miR-19b-3p | −0.21 | 0.06 | 0.020 | CGA+, GCNT3+, SPRR2E+, TREH+, NAT8+, TLR3+, OR10S1+, CUEDC1− | |
| hsa-miR-140-3p | −0.37 | 0.11 | 0.031 | ENTPD7−, CYP19A1−, OGT+, FMN1+, NR4A2+, B3GNT5−, LIPG−, FAM20A− | |
| hsa-miR-10b-5p | −0.38 | 0.11 | 0.026 | STAB2+, PROX1+, RELN+, CCL21+, SMPD4−, IRS4−, PDE2A+, CGA− | |
| hsa-miR-29a-3p | −0.51 | 0.11 | 0.003 | BTG4+, DLK1−, ALDH8A1+, PCSK4−, C4BPB+, AKAP4+, EDN3+, GML+ | |
| hsa-miR-675-3p | −0.54 | 0.18 | 0.051 | KRTAP10−3+, ZNF263−, KRTAP10-12+, ASTN2+, FAM78B+, POLR2E−, TAF8−, BLVRB− | |
| hsa-miR-29c-3p | −0.60 | 0.16 | 0.020 | TRAF3IP2−, GJA4−, STX5−, TOM1−, TBC1D17−, PCIF1−, FTL−, DPF2− | |
| hsa-miR-196b-5p | −0.93 | 0.26 | 0.031 | UTS2R−, NOP10−, ALDH16A1+, OR14C36+, ORM1−, SFMBT1+, UBL5+, GNPAT− |
| miRNA | baseMean | log2FoldChange | FDR | Outcome * |
|---|---|---|---|---|
| hsa-miR-101-3p | 36,355.424 | −0.406 | 0.001828 | HDM |
| 36,896.186 | −0.255 | 0.045888 | BDR | |
| hsa-miR-10b-5p | 208,777.388 | 0.287 | 6.97 × 107 | IGE |
| 209,492.165 | 0.234 | 0.000494 | EOS | |
| 208,991.420 | 0.183 | 0.023477 | HDM | |
| 208,591.796 | 0.296 | 1.07 × 106 | FEV1PP | |
| 208,591.796 | −0.133 | 0.027242 | FEV1/FVC | |
| 210,699.398 | −0.247 | 0.000545 | BDR | |
| hsa-miR-140-3p | 21,185.162 | −0.564 | 5.61 × 105 | HDM |
| 21,724.957 | −0.372 | 0.004762 | BDR | |
| hsa-miR-19b-3p | 1272.550 | 0.507 | 0.002363 | IGE |
| 1264.854 | 0.419 | 0.03691 | EOS | |
| 1282.529 | 0.590 | 0.00032 | FEV1/FVC | |
| 1284.974 | −0.493 | 0.011979 | BDR | |
| hsa-miR-423-5p | 120,957.129 | −0.263 | 0.000638 | IGE |
| 120,792.953 | −0.318 | 0.000837 | HDM | |
| hsa-miR-543 | 124.481 | −0.783 | 0.000494 | EOS |
| 124.296 | −0.928 | 0.000569 | HDM | |
| 124.448 | −0.545 | 0.01383 | IGE | |
| 125.414 | 0.545 | 0.036505 | BDR | |
| hsa-miR-200a-3p | 440.055 | 0.286 | 0.02454 | FEV1/FVC |
| hsa-miR-221-5p | 2126.747 | 0.285 | 0.006463 | FEV1/FVC |
| hsa-miR-29a-3p | 6308.506 | 0.354 | 0.00032 | FEV1/FVC |
| hsa-miR-29c-3p | 353.824 | 0.507 | 0.002031 | FEV1/FVC |
| hsa-miR-532-5p | 783.174 | 0.438 | 0.006463 | FEV1/FVC |
| miRNA | baseMean | log2FoldChange | p-Value | FDR | Outcome * |
|---|---|---|---|---|---|
| Male | |||||
| hsa-miR-101-3p | 37,732.87 | −0.69266 | 2.37 × 106 | 1.5 × 105 | HDM |
| hsa-miR-101-3p | 37,495.54 | −0.48684 | 3.42 × 105 | 0.000163 | IGE |
| hsa-miR-10b-5p | 221,947.5 | −0.23555 | 0.004001 | 0.038013 | BDR |
| hsa-miR-10b-5p | 220,378.5 | 0.228005 | 0.003688 | 0.023355 | EOS |
| hsa-miR-10b-5p | 219,178.4 | −0.23061 | 0.001313 | 0.012474 | FEV1/FVC |
| hsa-miR-10b-5p | 219,247 | 0.313008 | 1.52 × 105 | 9.6 × 105 | IGE |
| hsa-miR-10b-5p | 219,178.4 | 0.502512 | 2.26 × 1011 | 4.29 × 1010 | FEV1PP |
| hsa-miR-140-3p | 21,853.83 | −0.98959 | 1.49 × 1010 | 2.83 × 109 | HDM |
| hsa-miR-140-3p | 21,885.73 | −0.70774 | 1.17 × 108 | 2.23 × 103 | IGE |
| hsa-miR-140-3p | 22,081.31 | −0.41309 | 0.001861 | 0.017682 | FEV1PP |
| hsa-miR-29a-3p | 5927.324 | −0.35357 | 0.001905 | 0.0181 | EOS |
| hsa-miR-423-5p | 130,667.4 | −0.30672 | 0.000622 | 0.011822 | FEV1/FVC |
| hsa-miR-423-5p | 130,778.8 | −0.5239 | 3.50 × 106 | 1.66 × 105 | HDM |
| hsa-miR-423-5p | 130,273.6 | −0.43617 | 1.22 × 106 | 1.16 × 105 | IGE |
| hsa-miR-543 | 142.4763 | 0.907437 | 0.000937 | 0.017796 | BDR |
| hsa-miR-543 | 140.9837 | −1.11925 | 2.20 × 105 | 0.000418 | EOS |
| hsa-miR-543 | 140.4037 | −1.54768 | 4.05 × 107 | 3.85 × 106 | HDM |
| hsa-miR-543 | 141.141 | −0.97293 | 7.37 × 105 | 0.00028 | IGE |
| Female | |||||
| hsa-miR-148a-3p | 13,627.06 | 0.325966 | 0.000904 | 0.01717 | IGE |
| hsa-miR-1246 | 190.8938 | -0.51509 | 0.001765 | 0.03354 | FEV1/FVC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosenberg, L.; Liu, C.; Sharma, R.; Wood, C.; Vyhlidal, C.A.; Gaedigk, R.; Kho, A.T.; Ziniti, J.P.; Celedón, J.C.; Tantisira, K.G.; et al. Intrauterine Smoke Exposure, microRNA Expression during Human Lung Development, and Childhood Asthma. Int. J. Mol. Sci. 2023, 24, 7727. https://doi.org/10.3390/ijms24097727
Rosenberg L, Liu C, Sharma R, Wood C, Vyhlidal CA, Gaedigk R, Kho AT, Ziniti JP, Celedón JC, Tantisira KG, et al. Intrauterine Smoke Exposure, microRNA Expression during Human Lung Development, and Childhood Asthma. International Journal of Molecular Sciences. 2023; 24(9):7727. https://doi.org/10.3390/ijms24097727
Chicago/Turabian StyleRosenberg, Lynne, Cuining Liu, Rinku Sharma, Cheyret Wood, Carrie A. Vyhlidal, Roger Gaedigk, Alvin T. Kho, John P. Ziniti, Juan C. Celedón, Kelan G. Tantisira, and et al. 2023. "Intrauterine Smoke Exposure, microRNA Expression during Human Lung Development, and Childhood Asthma" International Journal of Molecular Sciences 24, no. 9: 7727. https://doi.org/10.3390/ijms24097727
APA StyleRosenberg, L., Liu, C., Sharma, R., Wood, C., Vyhlidal, C. A., Gaedigk, R., Kho, A. T., Ziniti, J. P., Celedón, J. C., Tantisira, K. G., Weiss, S. T., McGeachie, M. J., Kechris, K., & Sharma, S. (2023). Intrauterine Smoke Exposure, microRNA Expression during Human Lung Development, and Childhood Asthma. International Journal of Molecular Sciences, 24(9), 7727. https://doi.org/10.3390/ijms24097727