
Extract of Aster koraiensis Nakai Leaf Ameliorates Memory Dysfunction via Anti-inflammatory Action

 ,
,
Abstract
1. Introduction
2. Results
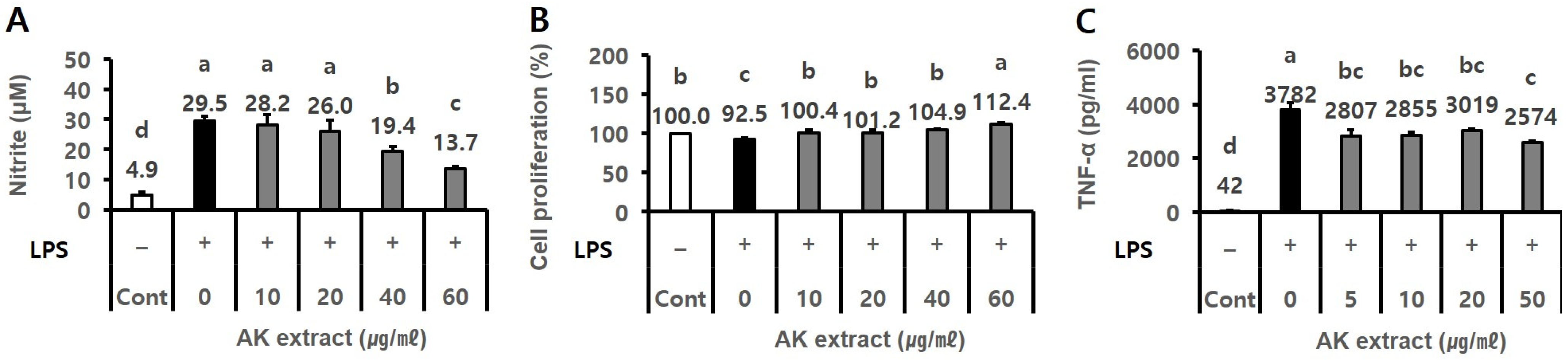
2.1. AK Extract Had Anti-Inflammatory Activity In Vitro
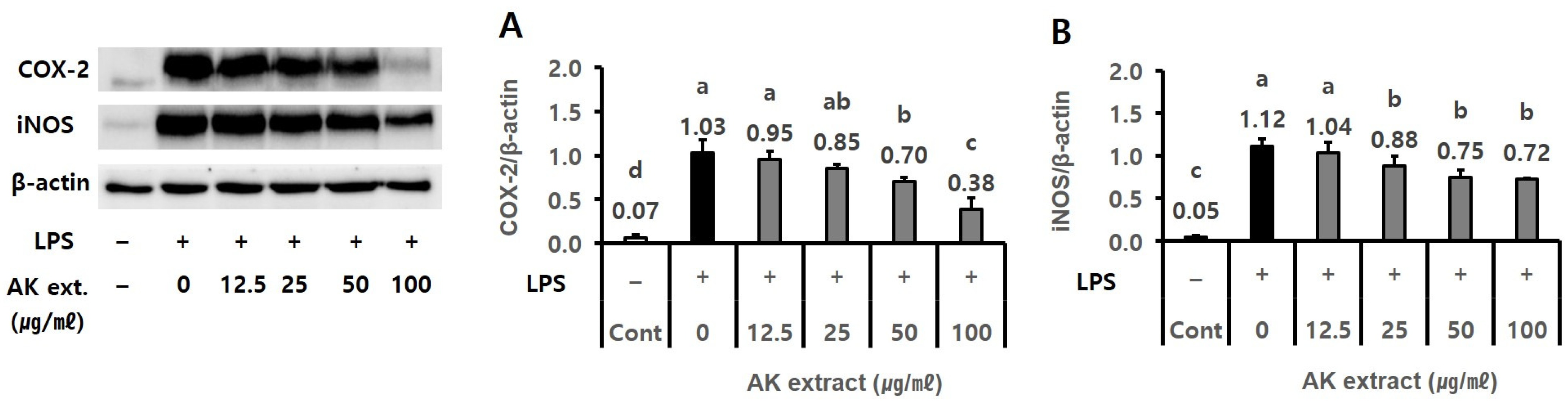
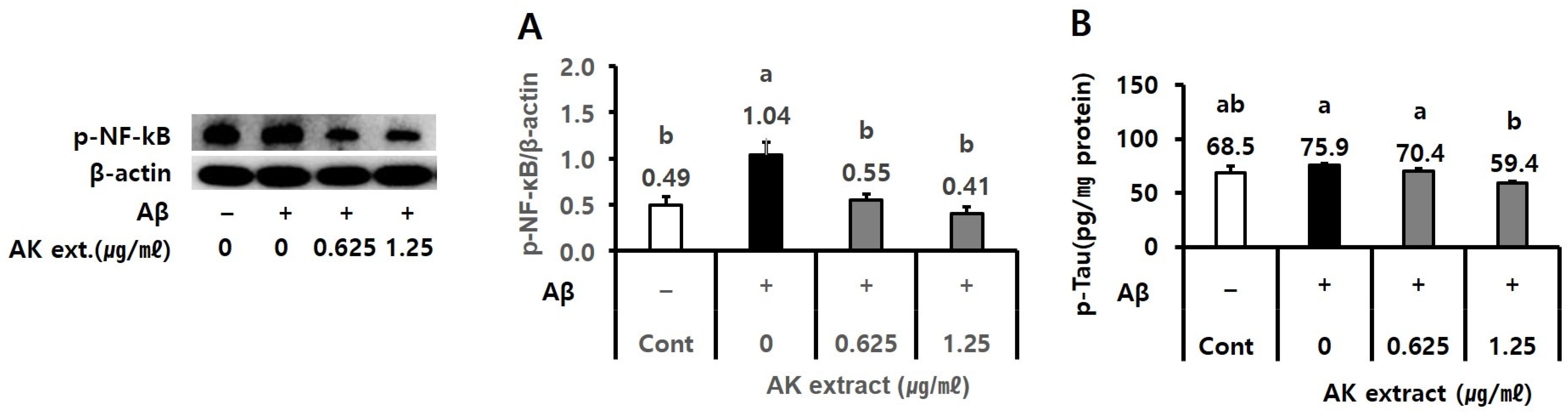
2.2. AK Extract Has Inhibitory Effects on the Expressions of Inflammatory Protein and the Production of P-Tau in Cells
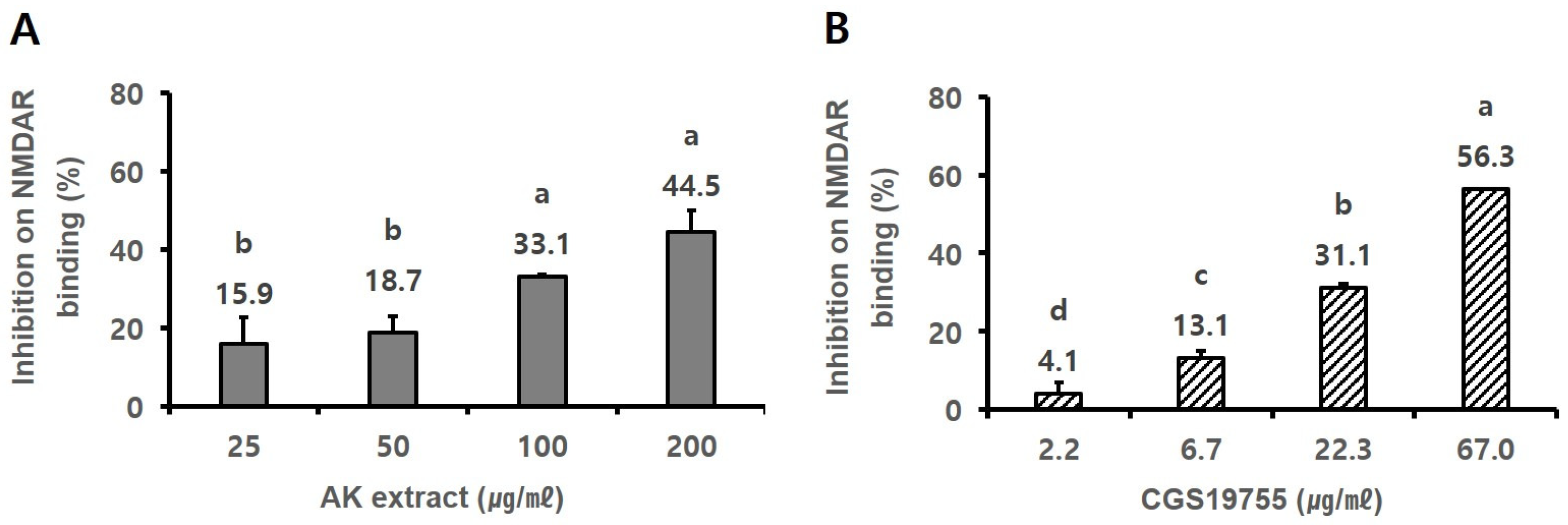
2.3. AK Extract Had In Vitro Binding Capacity on NMDA Receptor
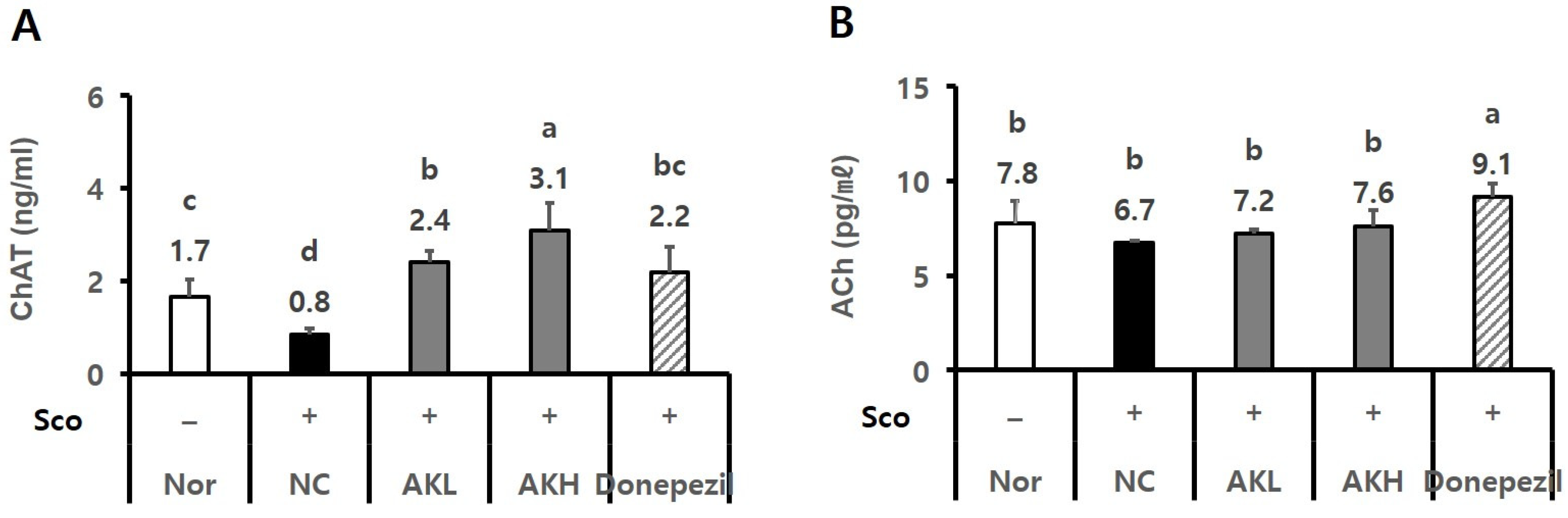
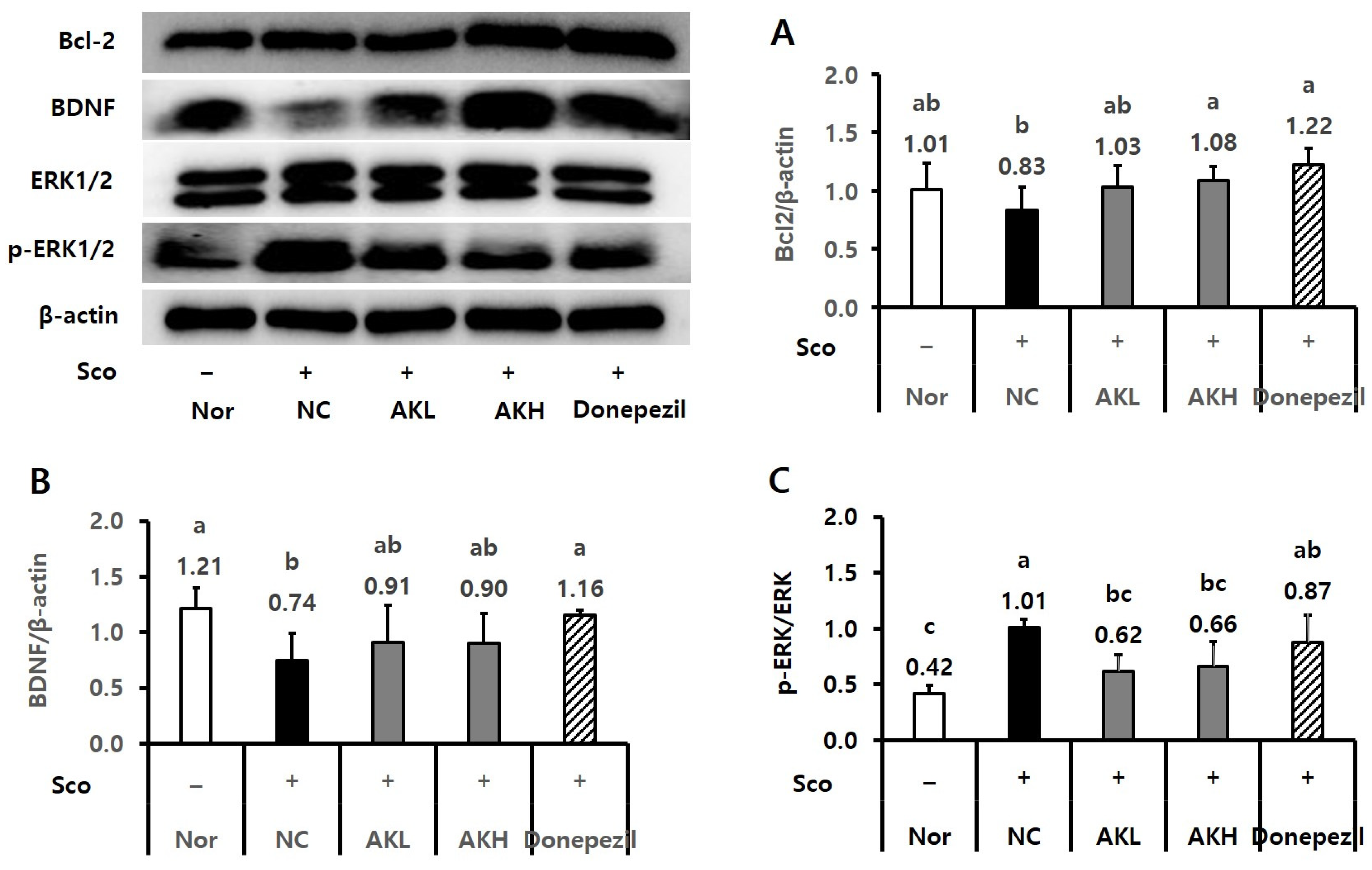
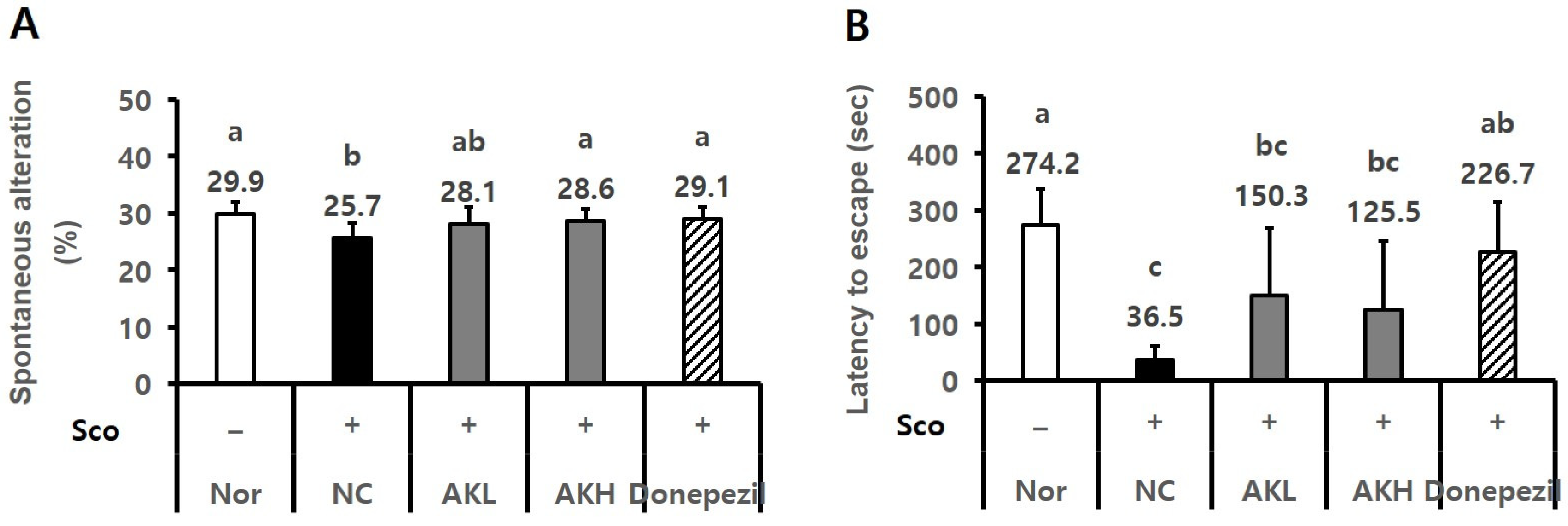
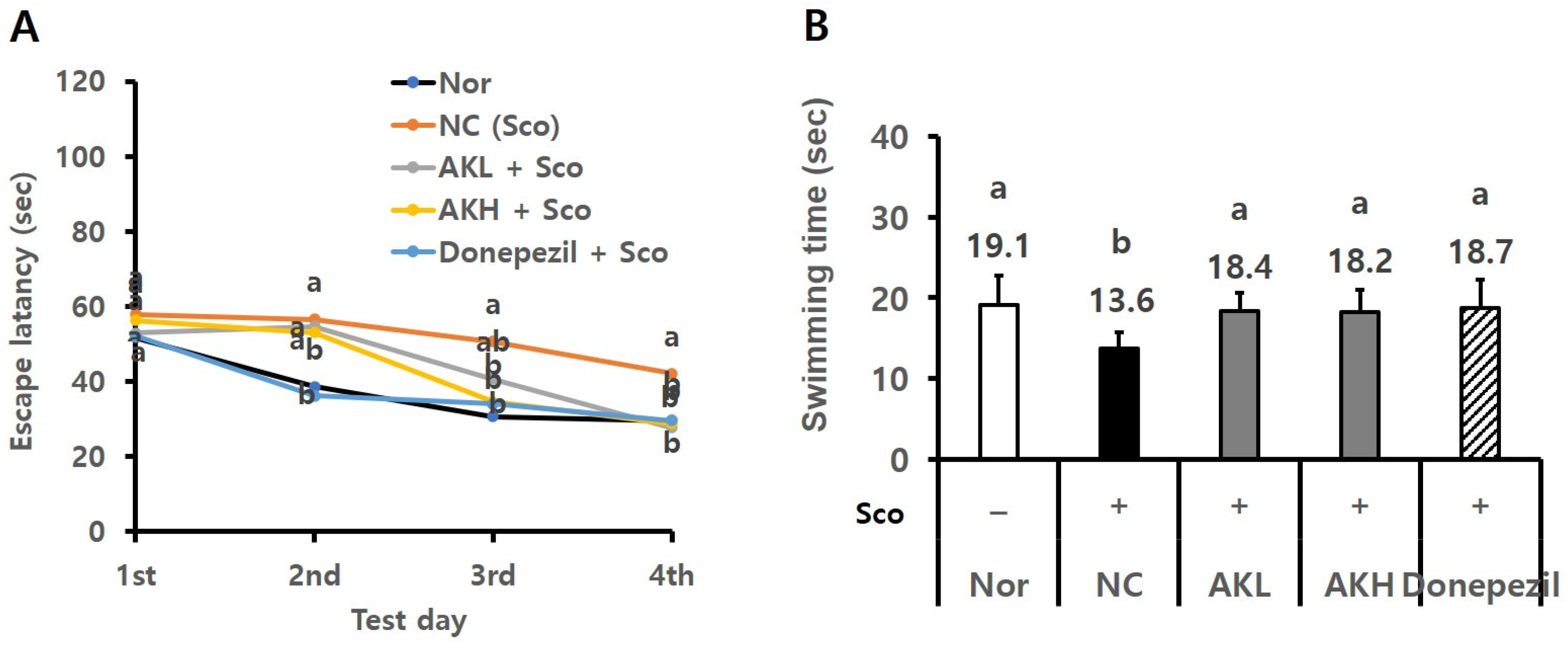
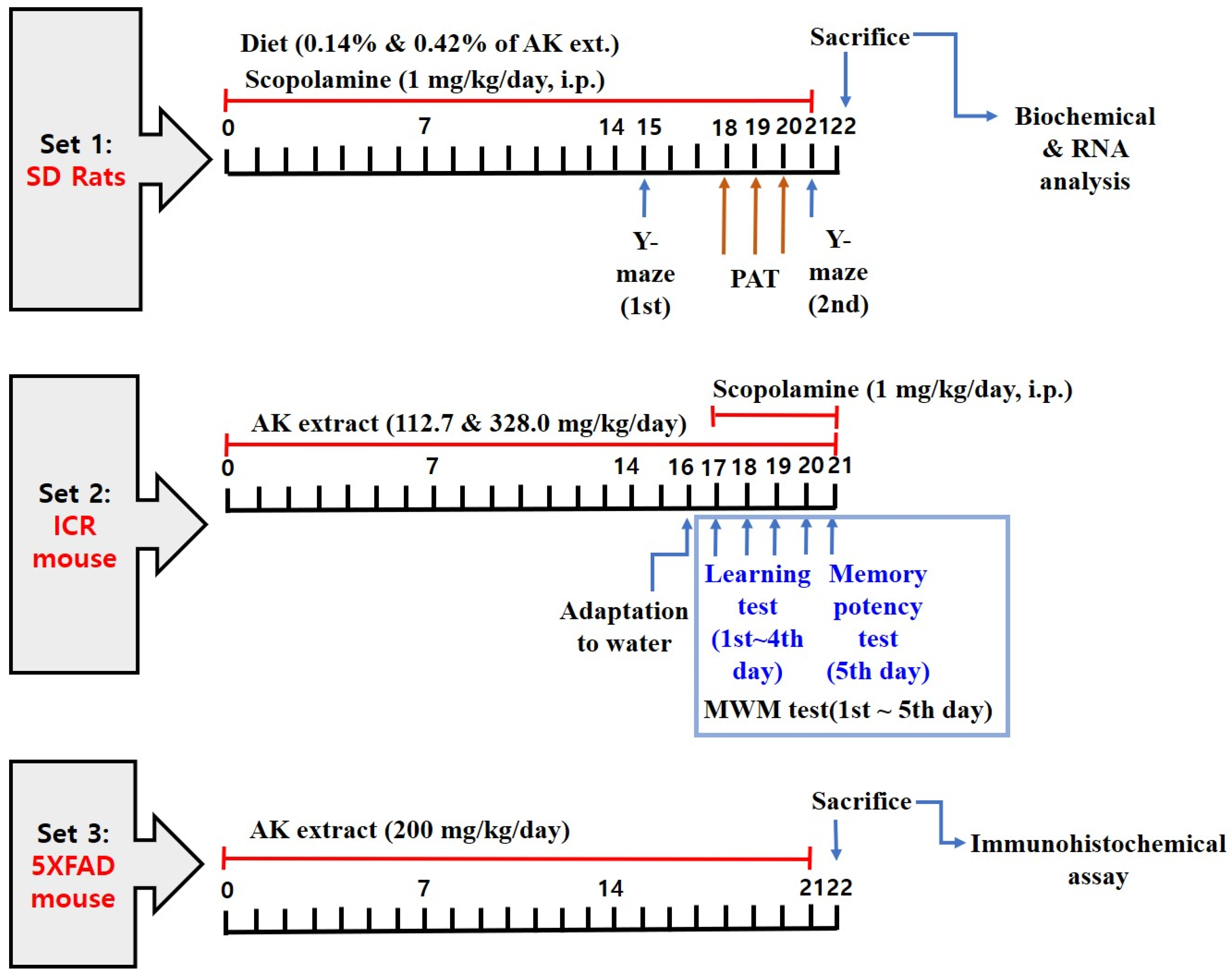
2.4. AK Extract Improved Memory in Rats Chronically Treated with Scopolamine
2.5. AK Extract Improved Memory in Rats Acutely Treated with Scopolamine
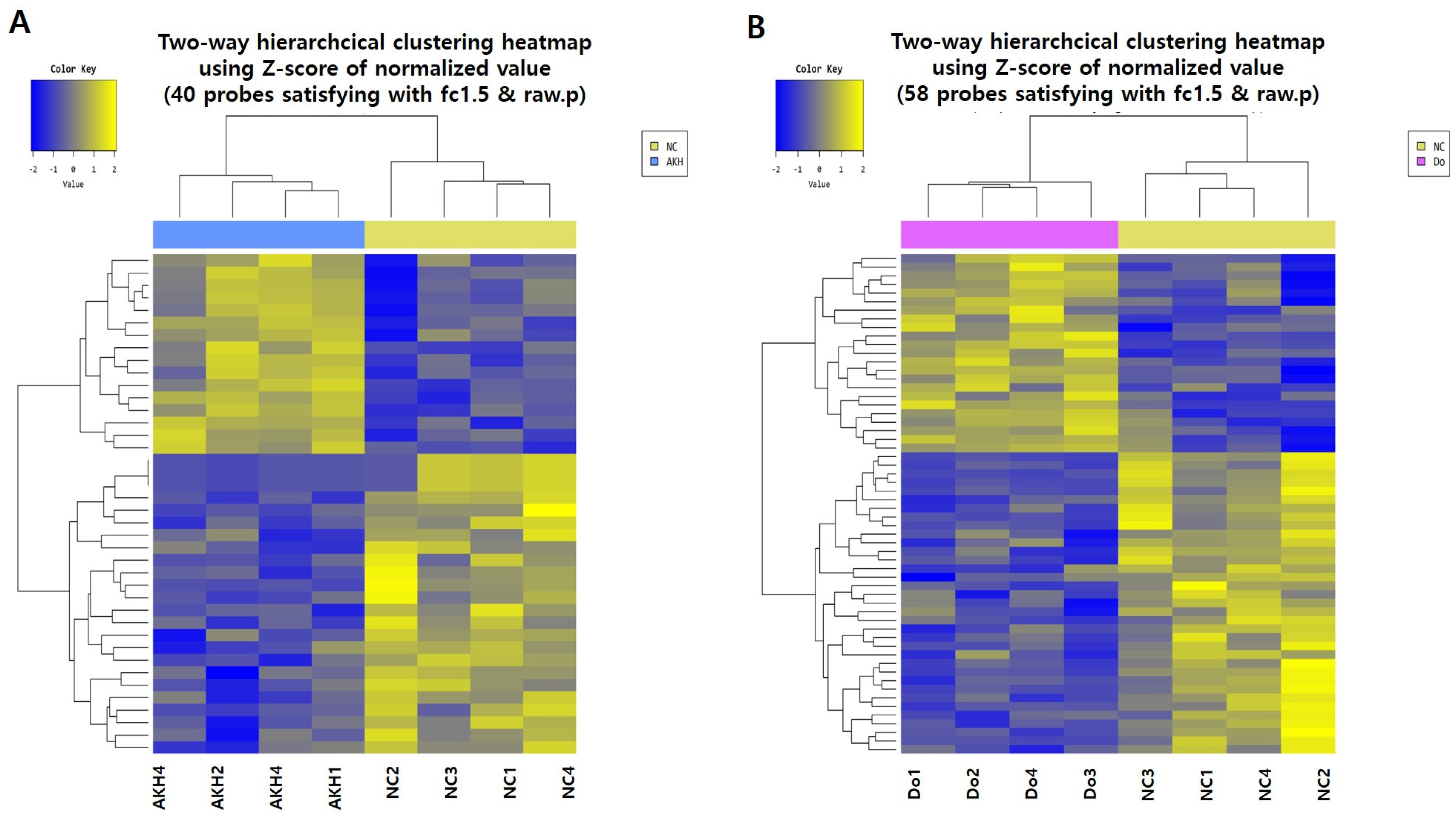
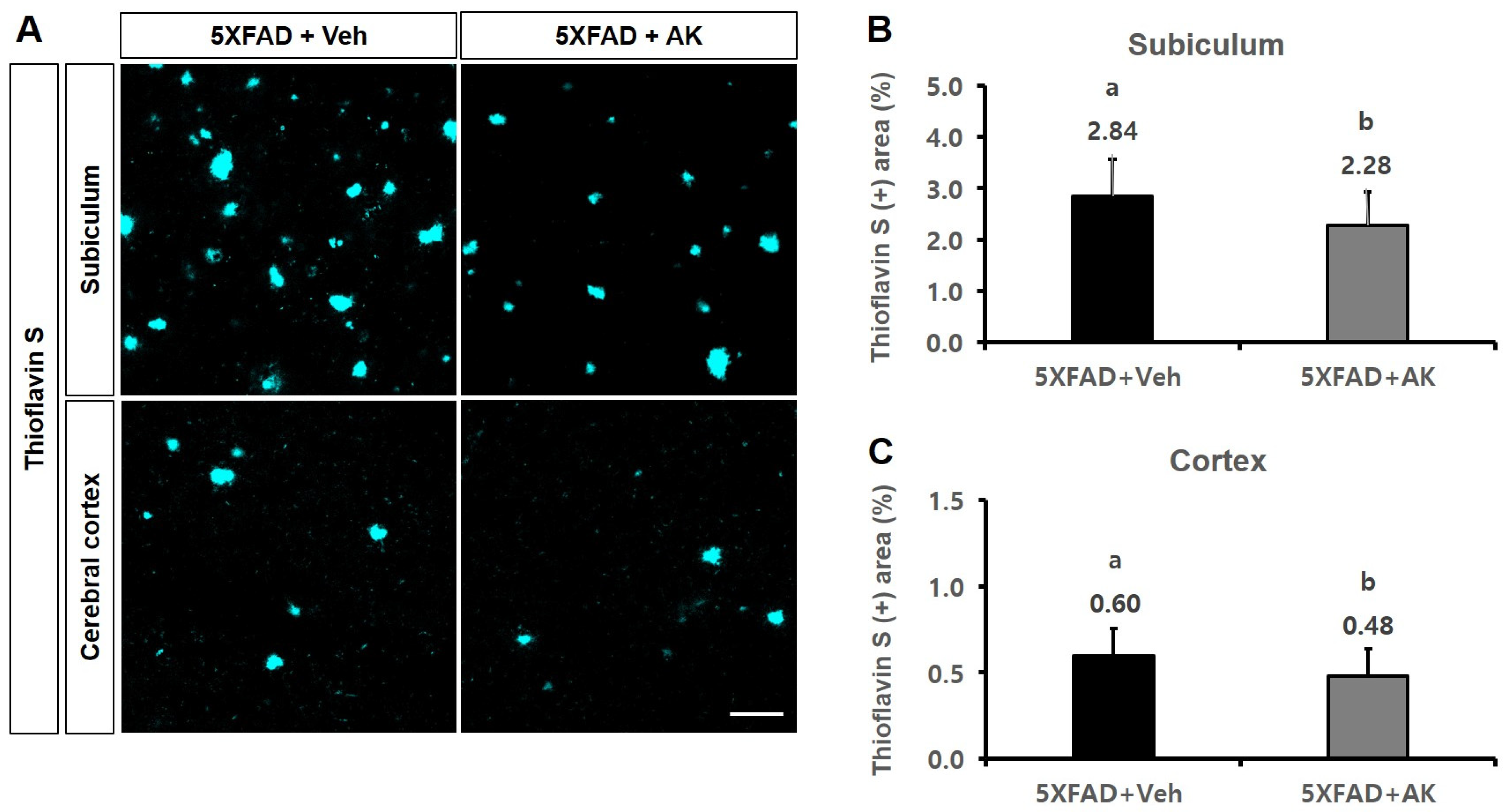
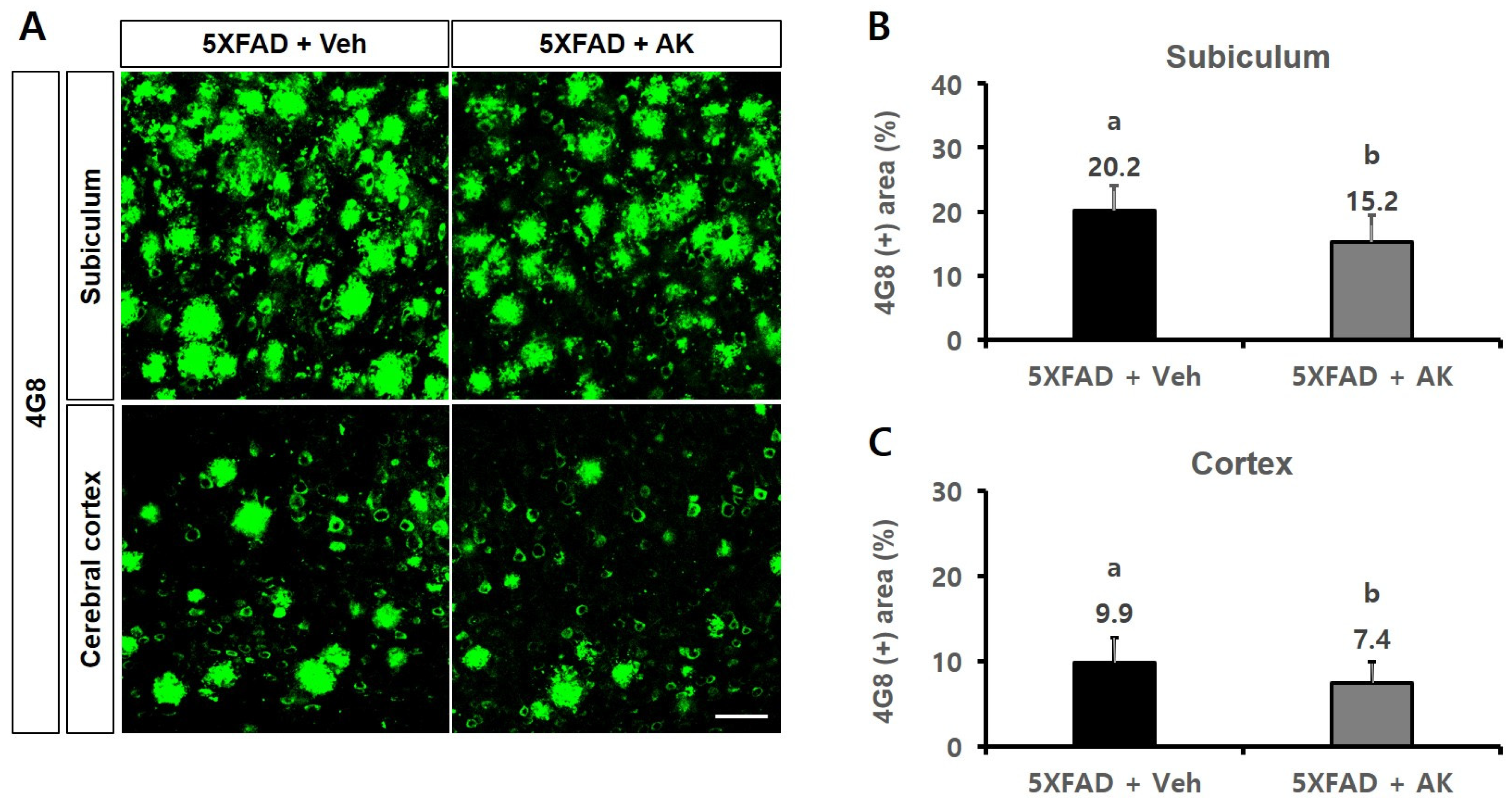
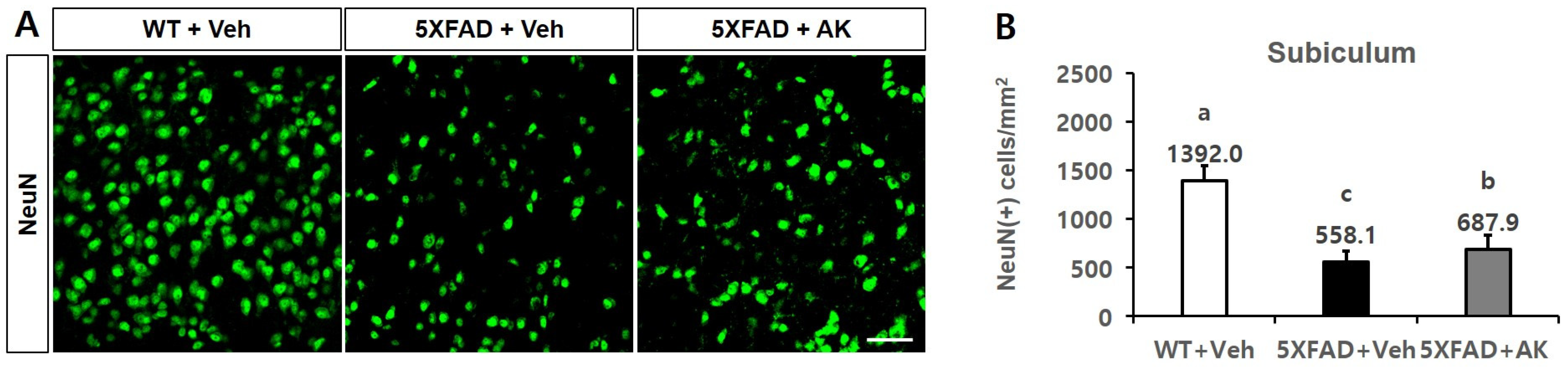
2.6. AK Extract Reduced Aβ Accumulation and Protected Neuronal Cells in the Brains of Transgenic Mice
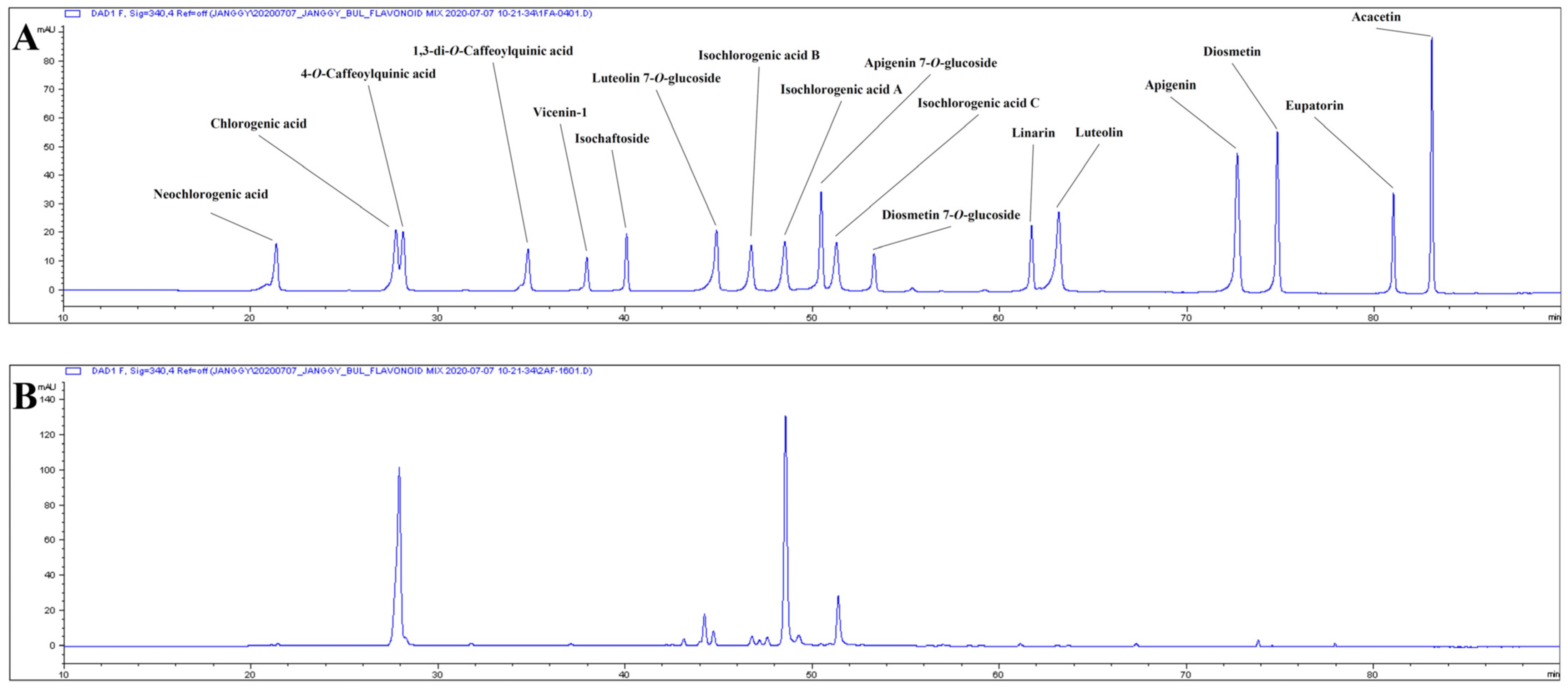
2.7. Phenolic Components of AK Extract
3. Discussion
4. Materials and Methods
4.1. Plant Resource and Extract Preparation
4.2. Chemicals
4.3. Cell Culture
4.4. Analysis of Inflammatory Activities in BV2 and RAW 264.7 Cells
4.5. Analysis of Protein Levels Related to Inflammatory Pathway and p-Tau Inhibition Activity in SH-SY5Y Cells
4.6. Analysis of Binding Capacity on NMDA Receptor
4.7. Analysis of the Effects on Scopolamine-Treated Animals
4.7.1. Experiments in Rats Chronically Treated with Scopolamine
4.7.2. Analysis of Biomarkers in Animals Chronically Treated with Scopolamine
4.7.3. Gene Expression Assay in Animals Chronically Treated with Scopolamine
4.7.4. Experiment in Mice Acutely Treated with Scopolamine
4.8. Analysis of the Effects of AK Extracts on Transgenic Mouse AD Model
4.9. Analysis of the Phenolic Composition of AK Extract
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alzheimer’s Disease International. World Alzheimer Report 2018: The state of the art of dementia research: New Front. 2018. Available online: https://www.alzint.org/u/WorldAlzheimerReport2018.pdf (accessed on 5 August 2022).
- Donev, R.; Kolev, M.; Millet, B.; Thome, J. Neuronal death in Alzheimer’s disease and therapeutic opportunities. J. Cell. Mol. Med. 2009, 13, 4329–4348. [Google Scholar] [CrossRef]
- He, Y.; Chen, Z.; Gong, G.; Evans, A. Neuronal networks in Alzheimer’s disease. Neuroscientist 2009, 15, 333–350. [Google Scholar] [CrossRef]
- Rahman, H.; Akter, R.; Bhattacharya, T.; Abdel-Daim, M.M.; Alkahtani, S.; Arafah, M.W.; Al-Johani, N.S.; Alhoshani, N.M.; Alkeraishan, N.; Alhenaky, A.; et al. Resveratrol and neuroprotection: Impact and its therapeutic potential in Alzheimer’s disease. Front. Pharmacol. 2020, 11, 619024. [Google Scholar] [CrossRef]
- Singhal, G.; Jaehne, E.J.; Corrigan, F.; Toben, C.; Baune, B.T. Inflammasomes in neuroinflammation and changes in brain function: A focused review. Front. Neurosci. 2014, 8, 315. [Google Scholar] [CrossRef]
- Muñoz-Moreno, E.; Tudela, R.; López-Gil, X.; Soria, G. Early brain connectivity alterations and cognitive impairment in a rat model of Alzheimer’s disease. Alzheimer’s Res. Ther. 2018, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Salinaro, A.T.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Crea, R.; Salvatore Cuzzocrea, S.; Paola, R.D.; et al. Hidrox® Roles in Neuroprotection: Biochemical Links between Traumatic Brain Injury and Alzheimer’s Disease. Antioxidants 2021, 10, 818. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Salinaro, A.T.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Cuzzocrea, S.; Paola, R.D.; Fusco, R.; et al. Key Mechanisms and Potential Implications of Hericium erinaceus in NLRP3 Inflammasome Activation by Reactive Oxygen Species during Alzheimer’s Disease. Antioxidants 2021, 10, 1664. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Ozben, T.; Ozben, S. Neuro-inflammation and anti-inflammatory treatment options for Alzheimer’s disease. Clin. Biochem. 2019, 72, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Hou, J.; Ping, J.; Cai, D. Advances in developing novel therapeutic strategies for Alzheimer’s disease. Mol. Neurodegener. 2018, 13, 64. [Google Scholar] [CrossRef]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar] [CrossRef]
- Moilanen, E.; Vapaatalo, H. Nitric oxde in inflammation and respose. Ann. Med. 1995, 27, 359–367. [Google Scholar] [CrossRef]
- Hosking, H. Nitric oxide and the immune system: A literature review. Plymouth Stud. Sci. 2009, 2, 270–278. Available online: http://hdl.handle.net/10026.1/13880 (accessed on 25 June 2022).
- Boje, K.M.; Arora, P.K. Microglial-produced nitric oxide and reactive nitrogen oxides mediate neuronal cell death. Brain Res. 1992, 587, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, K.R.; Brim, B.L.; Das, S.R. Selective vulnerabilities of N-methyl-D-aspartate (NMDA) receptors during brain aging. Front. Aging Neurosci. 2010, 2, 11. [Google Scholar] [CrossRef]
- Sharma, V.K.; Singh, T.G.; Garg, N.; Dhiman, S.; Gupta, S.; Rahman, H.; Najda, A.; Walasek-Janusz, M.; Kamel, M.; Albadrani, G.M.; et al. Dysbiosis and Alzheimer’s disease: A role for chronic stress? Biomolecules 2021, 11, 678. [Google Scholar] [CrossRef] [PubMed]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J. Neurol. Neurosurg. Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef]
- Bajo, R.; Pusil, S.; López, M.E.; Canuet, L.; Pereda, E.; Osipova, D.; Maestú, F.; Pekkonen, E. Scopolamine effects on functional brain connectivity: A pharmacological model of Alzheimer’s disease. Sci. Rep. 2015, 5, 9748. [Google Scholar] [CrossRef] [PubMed]
- Carlesimo, G.A.; Piras, F.; Orfei, M.D.; Iorio, M.; Caltagirone, C.; Spalletta, G. Atrophy of presubiculum and subiculum is the earliest hippocampal anatomical marker of Alzheimer’s disease. Alzheimers Dement. 2015, 1, 24–32. [Google Scholar] [CrossRef]
- Hyman, B.T.; Van Hoesen, G.W.; Damasio, A.R.; Barnes, C.L. Alzheimer’s disease: Cell-specific pathology isolates the hippocampal formation. Science 1984, 225, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Kim, K.J.; Kim, S.J.; Kim, S. Novel protease from the leaves of edible medicinal plant Aster koraiensis Nakai with antithrombotic activity: Purification and partial characterization. J. Food Biochem. 2017, 41, e12334. [Google Scholar] [CrossRef]
- Hong, C.H.; Hur, S.K.; Oh, O.; Kim, S.S.; Nam, K.A.; Lee, S.K. Evaluation of natural products on inhibition of inducible cyclooxygenase (COX-2) and nitric oxide synthase (iNOS) in cultured mouse macrophage cells. J. Ethnopharmacol. 2002, 83, 153–159. [Google Scholar] [CrossRef]
- Lee, J.; Lee, Y.M.; Lee, B.W.; Kim, J.H.; Kim, J.S. Chemical constituents from the aerial parts of Aster koraiensis with protein glycation and aldose reductase inhibitory activities. J. Nat. Prod. 2012, 75, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Kim, C.S.; Kim, J.S. Identification and quantification of human neutrophil elastase inhibitory caffeoylquinic acids in the leaves of Aster koraiensis. Nat. Prod. Commun. 2018, 13, 1519–1522. [Google Scholar] [CrossRef]
- Kwon, J.; Ko, K.; Zhang, L.; Zhao, D.; Yang, H.O.; Kwon, H.C. An autophagy inducing triterpene saponin derived from Aster koraiensis. Molecules 2019, 24, 4489. [Google Scholar] [CrossRef]
- Nhoek, P.; Ahn, J.; Chae, H.S.; Pel, P.; Kim, Y.M.; Lee, S.E.; Lee, J.H.; Kim, J.; Choi, Y.C.; Lee, K.; et al. Isolation of polyacetylenes with proprotein convertase/kexin type 9 downregulating activity and two new sesquiterpenes from the aerial parts of Aster koraiensis. Tetrahedron Lett. 2020, 61, 1519575. [Google Scholar] [CrossRef]
- Zhang, L.; Park, J.Y.; Zhao, D.; Kwon, H.C.; Yang, H.O. Neuroprotective effect of astersaponin I against Parkinson’s disease through autophagy induction. Biomol. Ther. 2021, 29, 615–629. [Google Scholar] [CrossRef]
- Sohn, E.; Kim, J.; Kim, C.S.; Kim, Y.S.; Jang, D.S.; Kim, J.S. Extract of the aerial parts of Aster koraiensis reduced development of diabetic nephropathy via anti-apoptosis of podocytes in streptozotocin-induced diabetic rats. Biochem. Biophys. Res. Commun. 2010, 391, 733–738. [Google Scholar] [CrossRef]
- Park, S.H.; Sim, Y.B.; Kim, S.M.; Kang, Y.J.; Lee, J.K.; Lim, S.S.; Kim, J.K.; Suh, H.W. Antinociceptive profiles and mechanisms of orally administered Aster koraiensis extract in the mouse. J. Med. Plant Res. 2011, 5, 6267–6272. [Google Scholar] [CrossRef]
- Kim, J.; Jo, K.; Kim, C.S.; Kim, J.S. Aster koraiensis extract prevents diabetes induced retinal vascular dysfunction in spontaneously diabetic Torii rats. BMC Complement. Altern. Med. 2017, 17, 497. [Google Scholar] [CrossRef]
- Kim, J.; Lee, Y.M.; Jung, W.; Park, S.B.; Kim, C.S.; Kim, J.S. Aster koraiensis extract and chlorogenic acid inhibit retinal angiogenesis in a mouse model of oxygen-induced retinopathy. Evid. Based Complement. Altern. Med. 2018, 2018, 6402650. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.W.; Kim, J.; Jo, K.; Kim, J.S.; Kim, C.S. Aster koraiensis extract improves impaired skin wound healing during hyperglycemia. Integr. Med. Res. 2018, 7, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hyun, S.W.; Lee, I.S.; Jo, K.; Kim, Y.S.; Kim, J.S.; Kim, C.S. Aster koraiensis extract lowers postprandial glucose in normoglycemic and high-fat-diet-induced obese mice. Food Sci. Biotechnol. 2019, 28, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.C.; Ha, J.H.; Lee, J.K.; Jung, S.H.; Kim, J.C. In vivo anti-inflammation potential of Aster koraiensis extract for dry eye syndrome by the protection of ocular surface. Nutrients 2020, 12, 3245. [Google Scholar] [CrossRef]
- Cameron, B.; Landreth, G.E. Inflammation, microglia and Alzheimer’s disease. Neurobiol. Dis. 2010, 37, 503–509. [Google Scholar] [CrossRef]
- Serpente, M.; Bonsi, R.; Scarpini, E.; Galimberti, D. Innate immune system and inflammation in Alzheimer’s disease; from pathogenesis to treatment. Neuroimmunomodulation 2014, 21, 79–87. [Google Scholar] [CrossRef]
- Akter, R.; Rahman, H.; Behl, T.; Chowdhury, M.A.R.; Manirujjaman, M.; Bulbul, I.J.; Elshenaw, S.E.; Tit, D.M.; Bungau, S. Prospective role of polyphenolic compounds in the treatment of neurodegenerative diseases. CNS Neurol. Disord. Drug Targets 2020, 20, 430–450. [Google Scholar] [CrossRef]
- Cai, Z.; Hussain, M.D.; Yan, L.J. Microglia, neuroinflammation, and beta-amyloid protein in Alzheimer’s disease. Int. J. Neurosci. 2014, 124, 307–321. [Google Scholar] [CrossRef]
- Viatour, P.; Merville, M.P.; Bours, V.; Alain Chariot, A. Phosphorylation of NF-kappaB and IkappaB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef]
- Carvajal, F.J.; Mattison, H.A.; Cerpa, W. Role of NMDA receptor-mediated glutamatergic signaling in chronic and acute neuropathologies. Neural Plast. 2016, 2016, 2701526. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.P.; Tarozzo, G.; Reggiani, A.; Piomelli, S.; Cavalli, A. Galantamine potentiates the neuroprotective effect of memantine against NMDA-induced excitotoxicity. Brain Behav. 2013, 3, 67–74. [Google Scholar] [CrossRef]
- Parsons, C.G.; Stoffler, A.; Danysz, W. Memantine: A NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system-too little activation is bad, too much is even worse. Neuropharmacology 2007, 53, 699–723. [Google Scholar] [CrossRef]
- Tabet, N. Acetylcholinesterase inhibitors for Alzheimer’s disease: Anti-inflammatories in acetylcholine clothing. Age Ageing 2006, 35, 336–338. [Google Scholar] [CrossRef]
- Lane, R.M.; Potkin, S.G.; Enz, A. Targeting acetylcholinesterase and butyrylcholinesterase in dementia. Int. J. Neuropsychopharmacol. 2006, 9, 101–124. [Google Scholar] [CrossRef]
- DeKosky, S.T.; Ikonomovic, M.D.; Styren, S.D.; Beckett, L.; Wisniewski, S.; Bennett, D.A.; Cochran, E.J.; Kordower, J.H.; Mufson, E.J. Upregulation of choline acetyltransferase activity in hippocampus and frontal cortex of elderly subjects with mild cognitive impairment. Ann. Neurol. 2002, 51, 145–155. [Google Scholar] [CrossRef]
- Wang, S.; Rosengren, L.E.; Franlund, M.; Hamberger, A.; Kenneth, G.; Haglid, K.G. Bcl-2 expression regulates cell sensitivity to S100b-mediated apoptosis. Mol. Brain Res. 1999, 70, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Song, H.; Park, M.N.; Kim, S.H.; Shim, B.S.; Kim, B. Ethanol extract of Oldenlandia diffusa herba attenuates scopolamine-induced cognitive impairments in mice via activation of BDNF, P-CREB and inhibition of acetylcholinesterase. Int. J. Mol. Sci. 2018, 19, 363. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nan, G. The extracellular signal-regulated kinase 1/2 pathway neurological diseases: A potential therapeutic target (Review). Int. J. Mol. Med. 2017, 39, 1338–1346. [Google Scholar] [CrossRef]
- Arning, L.; Stock, A.K.; Kloster, E.; Epplen, J.T.; Beste, C. NPY2-receptor variation modulates iconic memory processes. Eur. Neuropsychopharmacol. 2014, 24, 1298–1302. [Google Scholar] [CrossRef]
- Tohda, M.; Nomura, M.; Nomura, Y. Molecular pathopharmacology of 5-HT2c receptors and the RNA editing in the brain. J. Pharmacol. Sci. 2006, 100, 427–432. [Google Scholar] [CrossRef]
- Tanaka, K.F.; Samuels, B.A.; Hen, R. Serotonic receptor expression along the dorsal-ventral axis of mouse hippocampus. Philos. Trans. R. Soc. B 2012, 367, 2395–2401. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Koh, S.Q.; Guadagna, S.; Fracis, P.T.; Esiri, M.M.; Chen, C.P.; Wong, P.T.H.; Dawe, G.S.; Lai, M.K.P. Altered relaxin family receptors RXFP1 and RXFP3 in the neocortex of depressed Alzheimer’s disease patients. Psychopharmacology 2016, 233, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Sakata, K.; Overacre, A.E. Promoter Ⅳ-BDNF deficiency disturbs cholinergic gene expression of CHRNA5, CHRM2, and CHRM5: Effect of drug and environmental treatments. J. Neurochem. 2017, 143, 49–64. [Google Scholar] [CrossRef]
- Razani, E.; Pourbagheri-Sigaroodi, A.; Safaroghli-Azar, A.; Zoghi, A.; Shanaki-Bavarsad, M.; Bashash, D. The PI3K/Akt signaling axis in Alzheimer’s disease: A valuable target to stimulate or suppress? Cell Stress Chaperones 2021, 26, 871–887. [Google Scholar] [CrossRef]
- Sárvári, M.; Kalló, I.; Hrabovszky, E.; Solymosi, N.; Rodolosse, A.; Vastagh, C.; Auer, H.; Liposits, Z. Hippocampal Gene Expression Is Highly Responsive to Estradiol Replacement in Middle-Aged Female Rats. Endocrinology 2015, 156, 2632–2645. [Google Scholar] [CrossRef]
- Kenedy, P.G.E. Human African trypanosomiasis of the CNS: Current issues and challenges. J. Clin. Investig. 2004, 113, 496–504. [Google Scholar] [CrossRef]
- Dzikowski, R.; Templeton, T.J.; Deitsch, K. Variant antigen gene expression in malaria. Cell. Microbiol. 2006, 8, 1371–1381. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.J.; Jeong, Y.; Jeon, S.G.; Kim, S.; Lee, S.; Choi, H.S.; Im, C.S.; Kim, S.H.; Kim, S.H.; Park, J.H.; et al. Uncaria rhynchophylla ameliorates amyloid beta deposition and amyloid beta-mediated pathology in 5XFAD mice. Neurochem. Int. 2018, 121, 114–124. [Google Scholar] [CrossRef]
- Park, Y.H.; Shin, S.J.; Kim, H.S.; Hong, S.B.; Kim, S.; Nam, Y.; Kim, J.J.; Lim, K.; Kim, J.S.; Kim, J.; et al. Omega-3 fatty acid-type docosahexaenoic acid protects against Aβ-mediated mitochondrial deficits and pathomechanisms in Alzheimer’s disease-related animal model. Int. J. Mol. Sci. 2020, 21, 3879. [Google Scholar] [CrossRef]
- Saitou, K.; Ochiai, R.; Kozuma, K.; Sato, H.; Koikeda, T.; Osaki, N.; Katsuragi, Y. Effect of chlorogenic acids on cognitive function: A randomized, double-blind, placebo-controlled trial. Nutrients 2018, 10, 1337. [Google Scholar] [CrossRef] [PubMed]
- Miyamae, Y.; Kurisu, M.; Murakami, K.; Han, J.; Isoda, H.; Irie, K.; Shigemori, H. Protective effects of caffeoylquinic acids on the aggregation and neurotoxicity of the 42-residue amyloid β-protein. Bioorg. Med. Chem. 2012, 20, 5844–5849. [Google Scholar] [CrossRef]
- Gao, L.; Li, X.; Meng, S.; Ma, T.; Wan, L.; Xu, S. Chlorogenic acid alleviates Aβ25-35-induced autophagy and cognitive impairment via the mTOR/TFEB signaling pathway. Drug Des. Devel. Ther. 2020, 14, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Ochiai, R.; Kozuma, K.; Sato, H.; Katsuragi, Y. Effect of chlorogenic acid intake on cognitive function in the elderly: A pilot study. Evid. Based Complement. Altern. Med. 2018, 2018, 8608497. [Google Scholar] [CrossRef]
- Liu, W.; Li, J.; Zhang, X.; Zu, Y.; Yang, Y.; Liu, W.; Xu, Z.; Gao, H.; Sun, X.; Jiang, X.; et al. Current advances in naturally occurring caffeoylquinic acids: Structure, bioactivity, and synthesis. J. Agric. Food Chem. 2020, 68, 10489–10516. [Google Scholar] [CrossRef] [PubMed]
- Sills, M.A.; Fagg, G.; Pozza, M.; Angst, C.; Brundish, D.E.; Hurt, S.D.; Wilusz, E.J.; Williams, M. [3H]CGP 39653: A new N-methyl-D-aspartate antagonist radioligand with low nanomolar affinity in rat brain. Eur. J. Pharmacol. 1991, 192, 19–24. [Google Scholar] [CrossRef]
- Malekzadeh, S.; Edalatmanesh, M.A.; Mehrabani, D.; Shariati, M. Drugs induced Alzheimer’s disease in animal model. Galen Med. J. 2017, 6, 185–196. [Google Scholar] [CrossRef]
- Flood, J.F.; Cherkin, A. Scopolamine effects on memory retention in mice: A model of dementia? Behav. Neural Biol. 1986, 45, 169–184. [Google Scholar] [CrossRef]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine-induced impairment of spontaneous alteration behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology 1988, 94, 491–495. [Google Scholar] [CrossRef]
- Kim, H.G.; Moon, M.; Choi, J.G.; Park, G.; Kim, A.J.; Hur, J.; Lee, K.T.; Oh, M.S. Donepezil inhibits the amyloid oligomer-induced microglial activation in vitro and in vivo. Neurotoxicology 2014, 40, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Lee, S.; Chung, E.S.; Lim, S.; Kim, W.S.; Yoon, H.; Kim, S.K.; Ahn, K.S.; Jang, Y.P.; Bae, H. Neuroprotective effects of Cuscutae semen in a mouse model of Parkinson’s disease. Evid. Based Complement. Altern. Med. 2014, 2014, 150153. [Google Scholar] [CrossRef]
- Sadeghi, L.; Babadi, V.Y.; Tanwir, F. Improving effects of Echium amoenum aqueous extract on rat model of Alzheimer’s disease. J. Integr. Neurosci. 2018, 17, 661–669. [Google Scholar] [CrossRef]
- Saleem, U.; Raza, Z.; Anwar, F.; Ahmad, B.; Hira, S.; Ali, T. Experimental and computational studies to characterize and evaluate the therapeutic effect of Albizia lebbeck (L.) seeds in Alzheimer’s disease. Medicina 2019, 55, 184. [Google Scholar] [CrossRef] [PubMed]
- Elrod, K.; Buccafusco, J.J. An evaluation of the mechanism of scopolamine-induced impairment in two passive avoidance protocols. Pharmacol. Biochem. Behav. 1988, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kehr, J.; Yoshitake, S.; Ijiri, S.; Koch, E.; Nödner, M.; Yoshitake, T. Ginkgo biloba leaf extract (EGb 761®) and its specific acylated flavonol constituents increase dopamine and acetylcholine levels in the rat medial prefrontal cortex: Possible implications for the cognitive enhancing properties of EGb 761®. Int. Psychogeriatr. 2012, 24, 525–534. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
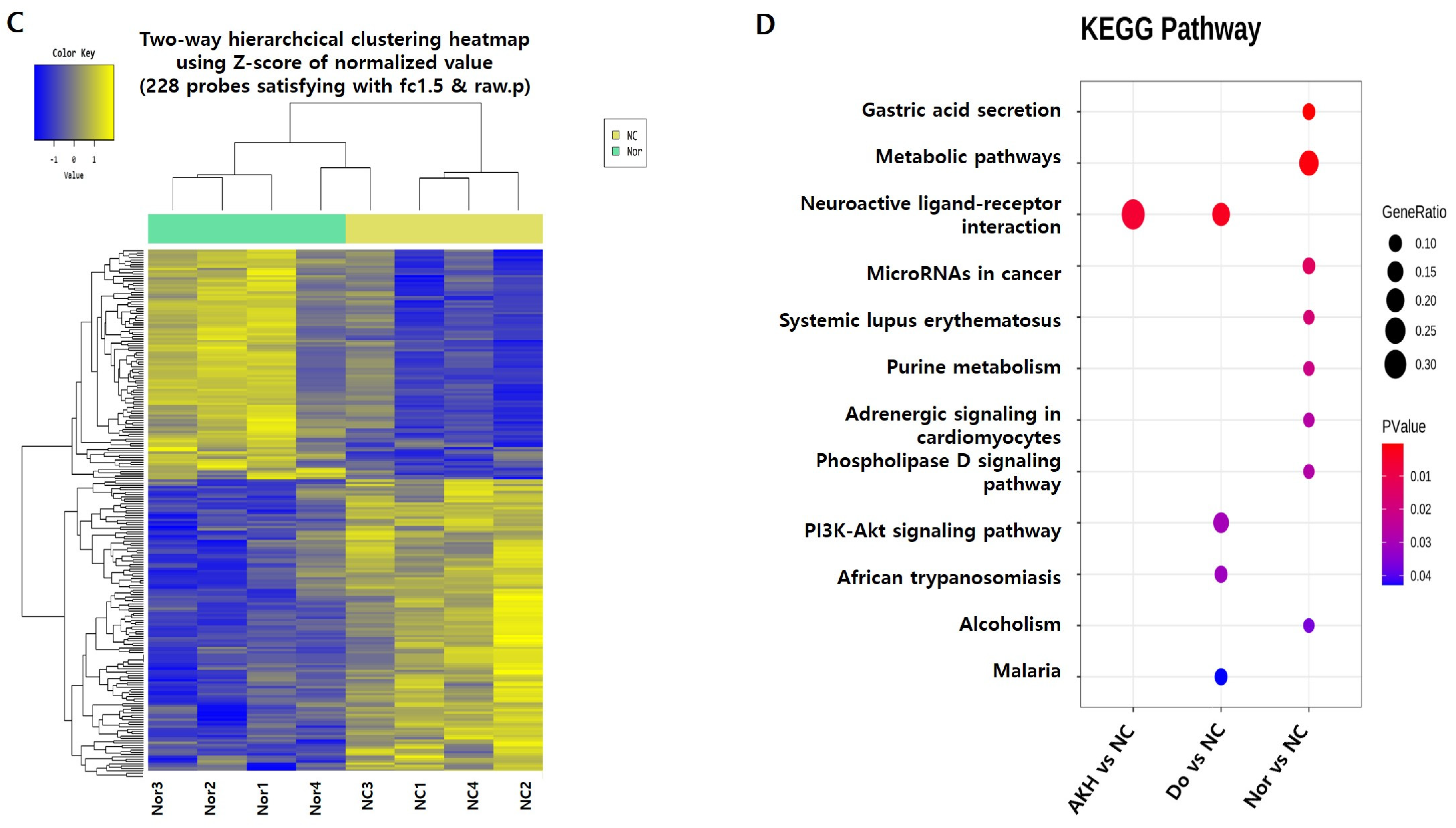
| MapName | p Value | Bonferroni | FDR | Gene Symbol | Fold Change | Raw p Value | |
|---|---|---|---|---|---|---|---|
| (A) AKH vs. NC | Neuroactive ligand -receptor interaction | 0.00582612 | 0.081565736 | 0.081565736 | Npy2r | 2.590162 | 0.023796771 |
| 0.00582612 | 0.081565736 | 0.081565736 | Npy2r | 2.578688 | 0.036323573 | ||
| 0.00582612 | 0.081565736 | 0.081565736 | Htr2c | 1.819431 | 0.048359603 | ||
| 0.00582612 | 0.081565736 | 0.081565736 | Rxfp1 | −1.940098 | 0.048331939 | ||
| (B) Donepezil vs. NC | Neuroactive ligand-receptor interaction | 0.00308635 | 0.138885974 | 0.138885974 | Npy2r | 2.200639 | 0.04181401 |
| 0.00308635 | 0.138885974 | 0.138885974 | Chrm5 | 1.583830 | 0.0307853 | ||
| 0.00308635 | 0.138885974 | 0.138885974 | Trhr | 1.605675 | 0.04482668 | ||
| 0.00308635 | 0.138885974 | 0.138885974 | Rxfp1 | −2.124701 | 0.03873565 | ||
| PI3K-Akt signalling pathway | 0.03017463 | 1 | 0.469178641 | Sgk1 | 1.650502 | 0.01282808 | |
| 0.03017463 | 1 | 0.469178641 | Igf2 | −3.045132 | 0.01642387 | ||
| 0.03017463 | 1 | 0.469178641 | Col1a2 | −1.967161 | 0.03576157 | ||
| African trypanosomiasis | 0.03127858 | 1 | 0.469178641 | LOC689064 | −2.205598 | 0.01463965 | |
| 0.03127858 | 1 | 0.469178641 | Hbb-b1 | −1.572961 | 0.04611988 | ||
| Malaria | 0.04276662 | 1 | 0.481124465 | LOC689064 | −2.205598 | 0.01463965 | |
| 0.04276662 | 1 | 0.481124465 | Hbb-b1 | −1.572961 | 0.04611988 |
| RT (min) | Phenolics | Content (mg/g extract, d.b.) |
|---|---|---|
| 21.40 | Neochlorogenic acid | 0.544 ± 0.009 |
| 27.88 | Chlorogenic acid | 54.539 ± 0.938 |
| 28.25 | 4-O-caffeoylquinic acid | 1.793 ± 0.042 |
| 40.11 | Isochaftoside | 0.112 ± 0.003 |
| 44.92 | Luteolin 7-O-glucoside | 2.895 ± 0.206 |
| 46.78 | Isochlorogenic acid B | 0.299 ± 0.010 |
| 48.58 | Isochlorogenic acid A | 65.983 ± 0.891 |
| 50.46 | Apigenin 7-O-glucoside | 0.377 ± 0.006 |
| 51.38 | Isochlorogenic acid C | 15.47 ± 0.521 |
| 63.16 | Luteolin | 2.752 ± 0.196 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-E.; Park, S.; Jang, G.Y.; Lee, J.; Moon, M.; Ji, Y.-J.; Jung, J.W.; Nam, Y.; Shin, S.J.; Lee, Y.; et al. Extract of Aster koraiensis Nakai Leaf Ameliorates Memory Dysfunction via Anti-inflammatory Action. Int. J. Mol. Sci. 2023, 24, 5765. https://doi.org/10.3390/ijms24065765
Lee S-E, Park S, Jang GY, Lee J, Moon M, Ji Y-J, Jung JW, Nam Y, Shin SJ, Lee Y, et al. Extract of Aster koraiensis Nakai Leaf Ameliorates Memory Dysfunction via Anti-inflammatory Action. International Journal of Molecular Sciences. 2023; 24(6):5765. https://doi.org/10.3390/ijms24065765
Chicago/Turabian StyleLee, Seung-Eun, Saetbyeol Park, Gwi Yeong Jang, Jeonghoon Lee, Minho Moon, Yun-Jeong Ji, Ji Wook Jung, Yunkwon Nam, Soo Jung Shin, Yunji Lee, and et al. 2023. "Extract of Aster koraiensis Nakai Leaf Ameliorates Memory Dysfunction via Anti-inflammatory Action" International Journal of Molecular Sciences 24, no. 6: 5765. https://doi.org/10.3390/ijms24065765
APA StyleLee, S.-E., Park, S., Jang, G. Y., Lee, J., Moon, M., Ji, Y.-J., Jung, J. W., Nam, Y., Shin, S. J., Lee, Y., Choi, J., & Kim, D. H. (2023). Extract of Aster koraiensis Nakai Leaf Ameliorates Memory Dysfunction via Anti-inflammatory Action. International Journal of Molecular Sciences, 24(6), 5765. https://doi.org/10.3390/ijms24065765