

Cell Intrinsic IL-38 Affects B Cell Differentiation and Antibody Production
 , , and
, , and 
Abstract
1. Introduction
2. Results
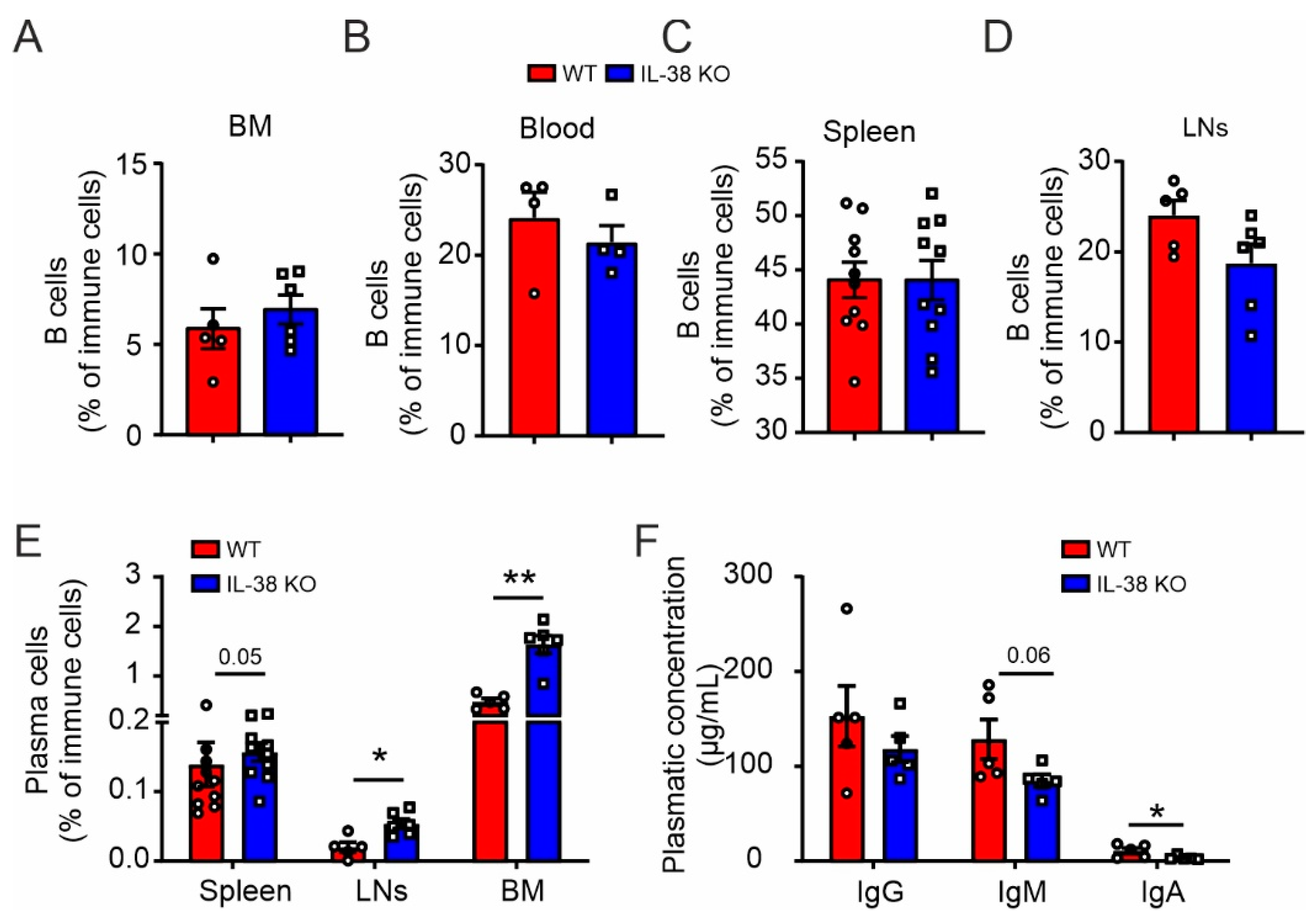
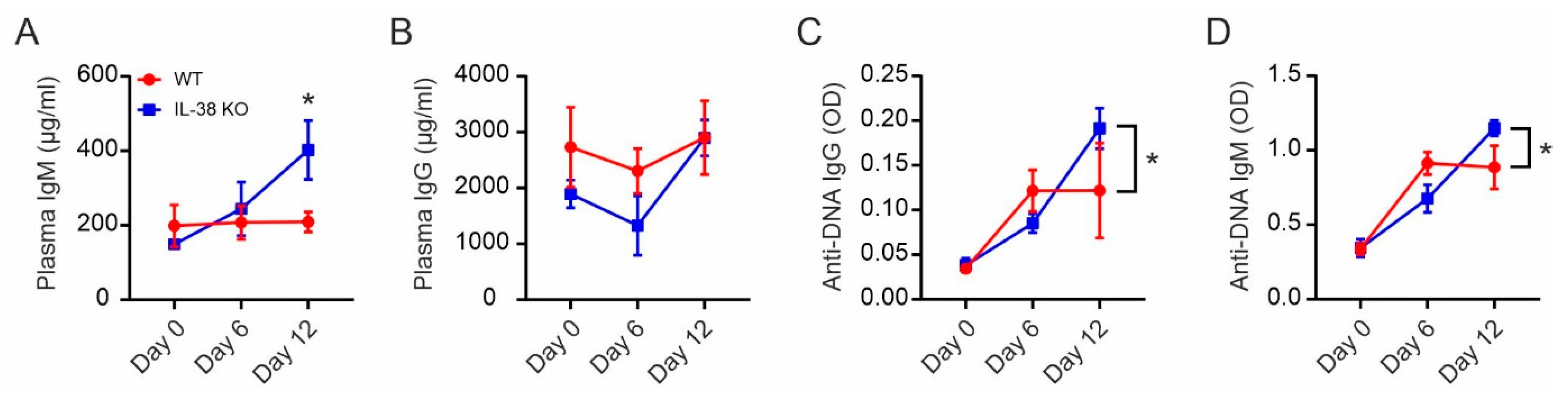
2.1. Impaired Plasma Cells Homeostasis in IL-38-Deficient Mice
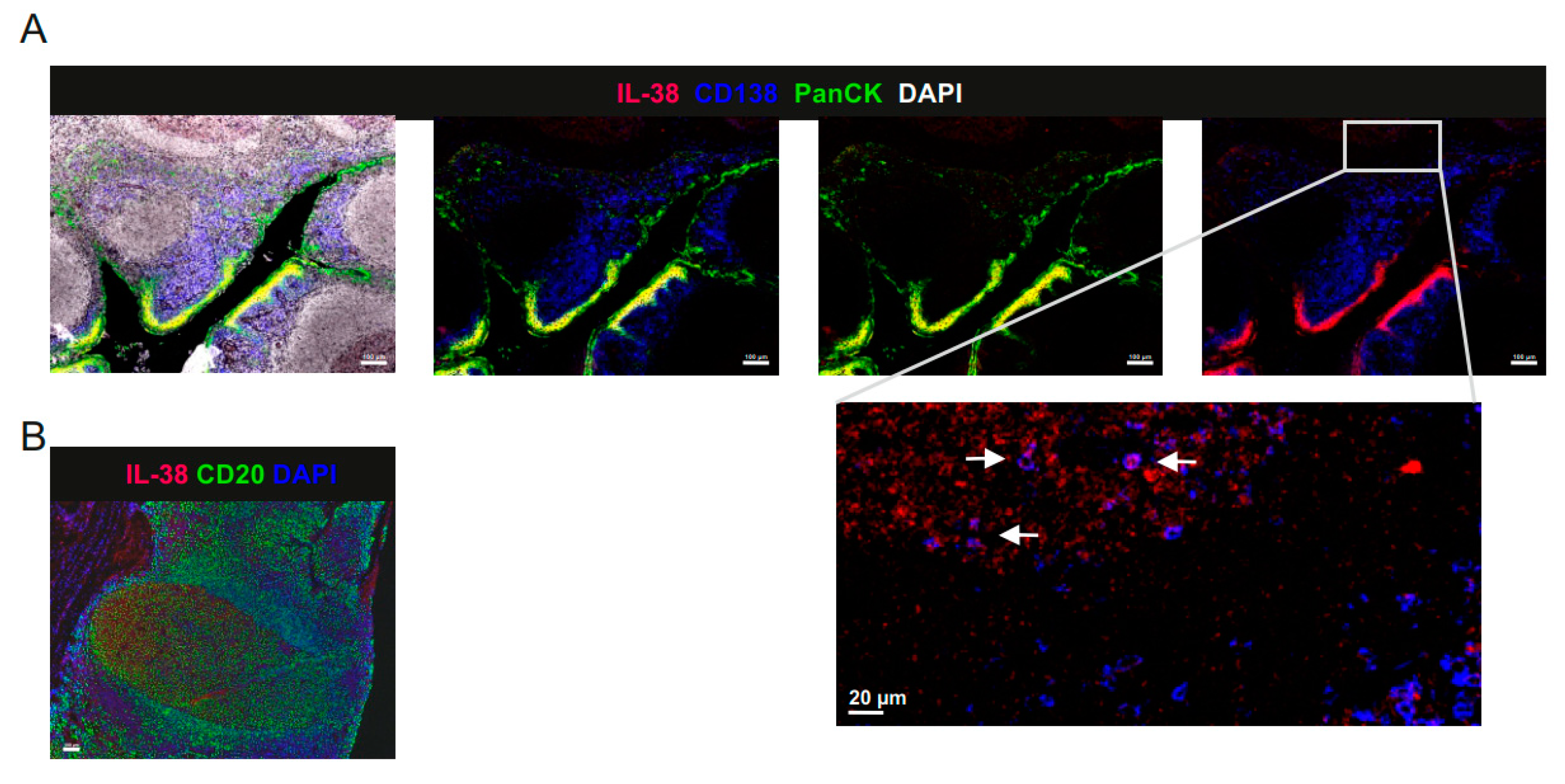
2.2. IL-38 Is Expressed in Tonsillar B Cells and Epithelium
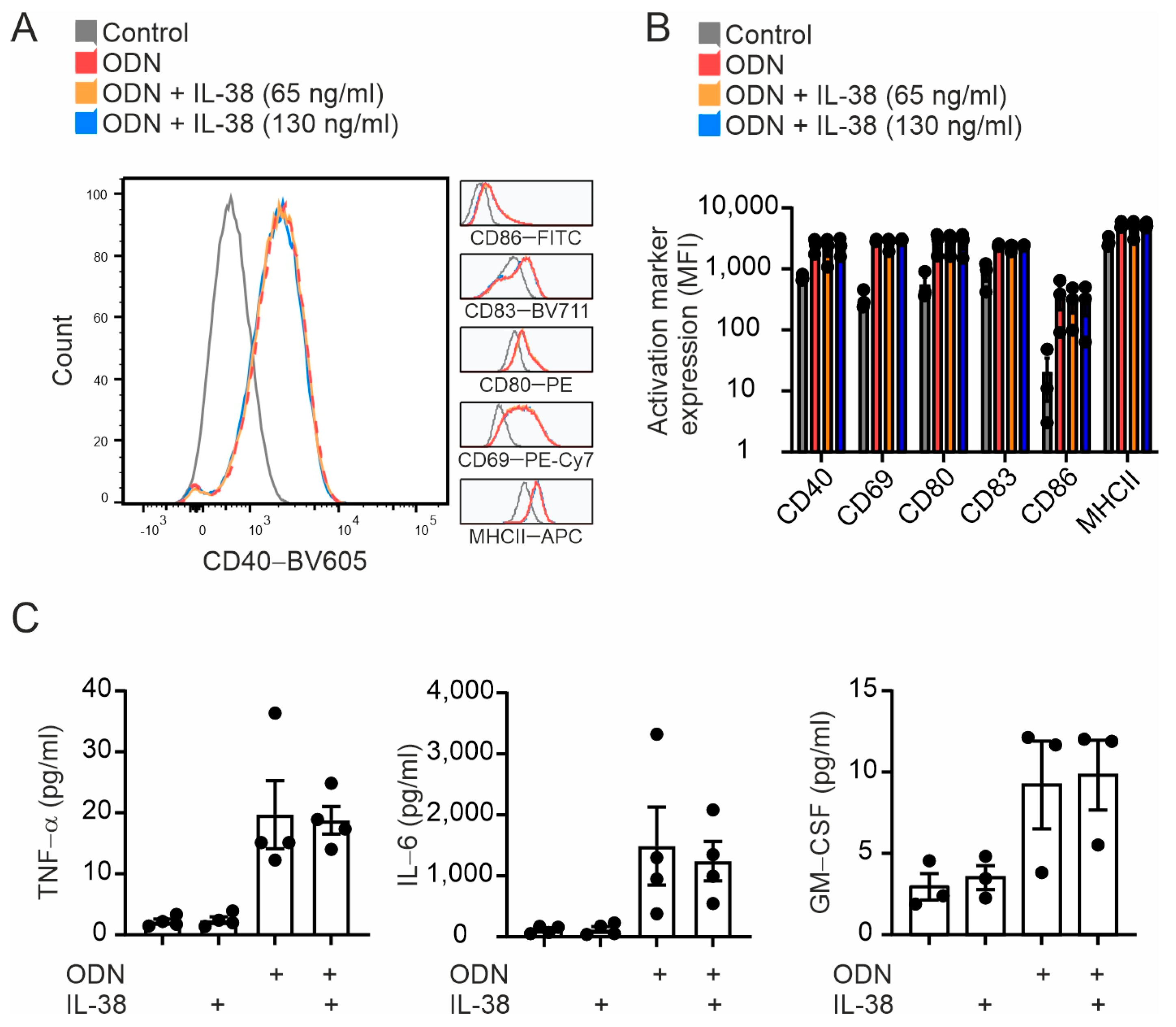
2.3. B Cell Activation Is Not Affected by Exogenous IL-38
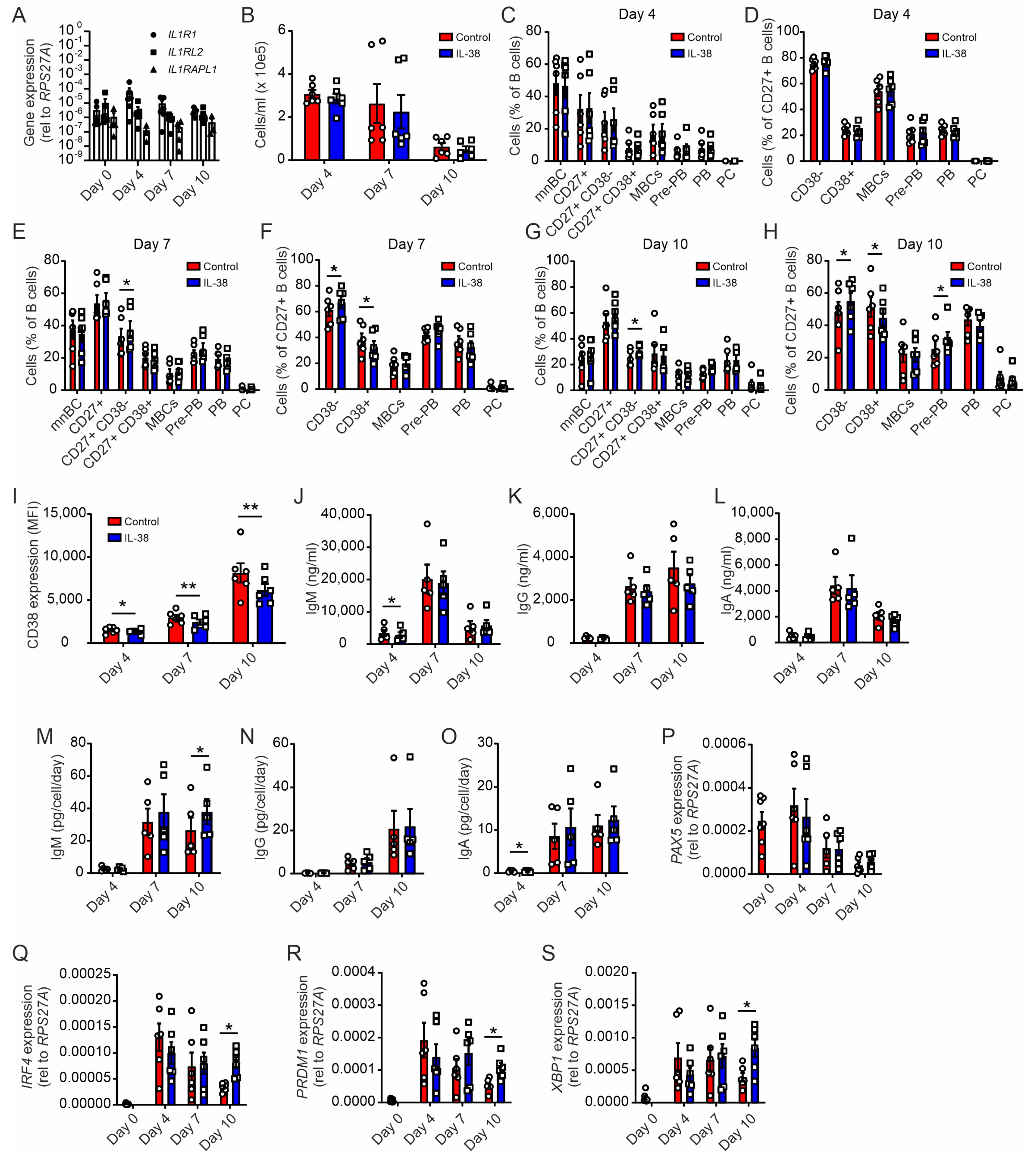
2.4. Plasma Cell Differentiation from Naïve B Cells Is Not Affected by Exogenous IL-38
2.5. Exogenous IL-38 Enhances IgM Production
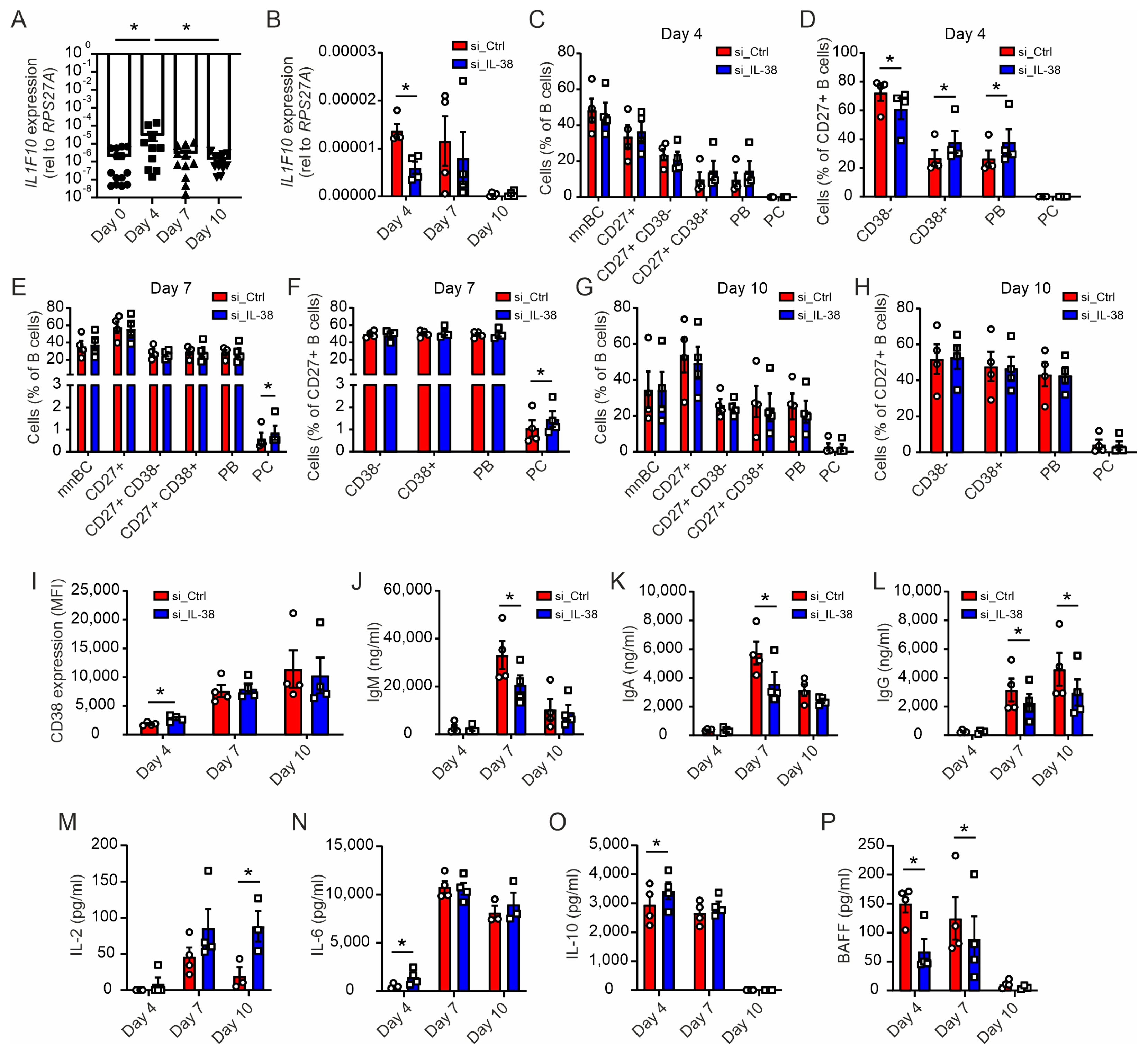
2.6. Early Silencing of IL-38 Promotes Plasma Cell Differentiation and Dampens Antibody Production
2.7. Absence of IL-38 Dysregulates Autoantibody Production
3. Discussion
4. Material and Methods
4.1. Animals
4.2. IL-18-Induced Autoantibody Production
4.3. Cells
4.4. B Cell Activation and Differentiation
4.5. B Cell Transfection
4.6. Flow Cytometry
4.7. Quantitative PCR
4.8. Cytometric Bead Array
4.9. LegendPlex
4.10. Histology
4.11. ELISA
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| B-CLL | B-cell chronic lymphocytic lymphoma |
| BCR | B cell receptor |
| BM | bone marrow |
| EAE | experimental autoimmune encephalomyelitis |
| GC | germinal center |
| IBD | inflammatory bowel diseases |
| IL | interleukin |
| IL-1R | interleukin-1 receptor |
| IL-1RAPL1 | X-linked IL-1 receptor accessory protein-like 1 |
| IMDM | Iscove’s modified Dulbecco’s Medium |
| KD | knockdown |
| LNs | lymph nodes |
| MBCs | memory B cells |
| mnBC | mature naïve B cells |
| PB | plasmablast |
| PC | plasma cells |
| Pre-PB | pre-plasmablasts |
| Ra | receptor antagonist |
| RA | rheumatoid arthritis |
| SLE | systemic-lupus-erythematosus |
References
- Lin, H.; Ho, A.S.; Haley-Vicente, D.; Zhang, J.; Bernal-Fussell, J.; Pace, A.M.; Hansen, D.; Schweighofer, K.; Mize, N.K.; Ford, J.E. Cloning and characterization of il-1hy2, a novel interleukin-1 family member. J. Biol. Chem. 2001, 276, 20597–20602. [Google Scholar] [CrossRef]
- Han, Y.; Mora, J.; Huard, A.; da Silva, P.; Wiechmann, S.; Putyrski, M.; Schuster, C.; Elwakeel, E.; Lang, G.; Scholz, A.; et al. Il-38 ameliorates skin inflammation and limits il-17 production from gammadelta t cells. Cell Rep. 2019, 27, 835–846.e5. [Google Scholar] [CrossRef]
- Mermoud, L.; Shutova, M.; Diaz-Barreiro, A.; Talabot-Ayer, D.; Drukala, J.; Wolnicki, M.; Kaya, G.; Boehncke, W.H.; Palmer, G.; Borowczyk, J. IL-38 orchestrates proliferation and differentiation in human keratinocytes. Exp. Dermatol. 2022, 31, 1699–1711. [Google Scholar] [CrossRef]
- Mercurio, L.; Morelli, M.; Scarponi, C.; Eisenmesser, E.Z.; Doti, N.; Pagnanelli, G.; Gubinelli, E.; Mazzanti, C.; Cavani, A.; Ruvo, M.; et al. Il-38 has an anti-inflammatory action in psoriasis and its expression correlates with disease severity and therapeutic response to anti-il-17a treatment. Cell Death Dis. 2018, 9, 1104. [Google Scholar] [CrossRef]
- van de Veerdonk, F.L.; Stoeckman, A.K.; Wu, G.; Boeckermann, A.N.; Azam, T.; Netea, M.G.; Joosten, L.A.; van der Meer, J.W.; Hao, R.; Kalabokis, V.; et al. Il-38 binds to the il-36 receptor and has biological effects on immune cells similar to il-36 receptor antagonist. Proc. Natl. Acad. Sci. USA 2012, 109, 3001–3005. [Google Scholar] [CrossRef]
- Mora, J.; Schlemmer, A.; Wittig, I.; Richter, F.; Putyrski, M.; Frank, A.C.; Han, Y.; Jung, M.; Ernst, A.; Weigert, A.; et al. Interleukin-38 is released from apoptotic cells to limit inflammatory macrophage responses. J. Mol. Cell Biol. 2016, 8, 426–438. [Google Scholar] [CrossRef]
- Han, Y.; Huard, A.; Mora, J.; da Silva, P.; Brune, B.; Weigert, A. Il-36 family cytokines in protective versus destructive inflammation. Cell. Signal. 2020, 75, 109773. [Google Scholar] [CrossRef]
- Boutet, M.A.; Bart, G.; Penhoat, M.; Amiaud, J.; Brulin, B.; Charrier, C.; Morel, F.; Lecron, J.C.; Rolli-Derkinderen, M.; Bourreille, A.; et al. Distinct expression of interleukin (il)-36α, βandγ, their antagonist il-36ra and il-38 in psoriasis, rheumatoid arthritis and crohn’s disease. Clin. Exp. Immunol. 2016, 184, 159–173. [Google Scholar] [CrossRef]
- Huard, A.; Do, H.N.; Frank, A.-C.; Sirait-Fischer, E.; Fuhrmann, D.; Hofmann, M.C.J.; Raue, R.; Palmer, G.; Brüne, B.; de Bruin, N.; et al. Il-38 ablation reduces local inflammation and disease severity in experimental autoimmune encephalomyelitis. J. Immunol. 2021, 206, 1058–1066. [Google Scholar] [CrossRef]
- de Graaf, D.M.; Jaeger, M.; van den Munckhof, I.C.L.; Ter Horst, R.; Schraa, K.; Zwaag, J.; Kox, M.; Fujita, M.; Yamauchi, T.; Mercurio, L.; et al. Reduced concentrations of the b cell cytokine interleukin 38 are associated with cardiovascular disease risk in overweight subjects. Eur. J. Immunol. 2021, 51, 662–671. [Google Scholar] [CrossRef]
- Xie, C.; Yan, W.; Quan, R.; Chen, C.; Tu, L.; Hou, X.; Fu, Y. Interleukin-38 is elevated in inflammatory bowel diseases and suppresses intestinal inflammation. Cytokine 2020, 127, 154963. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Imai, T.; Chatani, M.; Nishida, A.; Inatomi, O.; Kawahara, M.; Hoshino, T.; Andoh, A. The anti-inflammatory and protective role of interleukin-38 in inflammatory bowel disease. J. Clin. Biochem. Nutr. 2022, 70, 64–71. [Google Scholar] [CrossRef]
- Treanor, B. B-cell receptor: From resting state to activate. Immunology 2012, 136, 21–27. [Google Scholar] [CrossRef]
- Jellusova, J. Metabolic control of b cell immune responses. Curr. Opin. Immunol. 2020, 63, 21–28. [Google Scholar] [CrossRef]
- Victora, G.D.; Nussenzweig, M.C. Germinal centers. Annu. Rev. Immunol. 2012, 30, 429–457. [Google Scholar] [CrossRef]
- Hampe, C.S. B cells in autoimmune diseases. Scientifica 2012, 2012, 215308. [Google Scholar] [CrossRef]
- Rahman, P.; Sun, S.; Peddle, L.; Snelgrove, T.; Melay, W.; Greenwood, C.; Gladman, D. Association between the interleukin-1 family gene cluster and psoriatic arthritis. Arthritis Rheum. 2006, 54, 2321–2325. [Google Scholar] [CrossRef]
- Sims, A.M.; Timms, A.E.; Bruges-Armas, J.; Burgos-Vargas, R.; Chou, C.T.; Doan, T.; Dowling, A.; Fialho, R.N.; Gergely, P.; Gladman, D.D.; et al. Prospective meta-analysis of interleukin 1 gene complex polymorphisms confirms associations with ankylosing spondylitis. Ann. Rheum. Dis. 2008, 67, 1305–1309. [Google Scholar] [CrossRef]
- Guo, Z.S.; Li, C.; Lin, Z.M.; Huang, J.X.; Wei, Q.J.; Wang, X.W.; Xie, Y.Y.; Liao, Z.T.; Chao, S.Y.; Gu, J.R. Association of il-1 gene complex members with ankylosing spondylitis in chinese han population. Int. J. Immunogenet. 2010, 37, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Huang, Z.; Li, H.; Liu, X.; Zheng, S.; Su, W. Il-38: A new player in inflammatory autoimmune disorders. Biomolecules 2019, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Tsao, B.P.; Ohnishi, K.; Cheroutre, H.; Mitchell, B.; Teitell, M.; Mixter, P.; Kronenberg, M.; Hahn, B.H. Failed self-tolerance and autoimmunity in igg anti-DNA transgenic mice. J. Immunol. 1992, 149, 350–358. [Google Scholar] [CrossRef]
- Vlahakos, D.V.; Foster, M.H.; Adams, S.; Katz, M.; Ucci, A.A.; Barrett, K.J.; Datta, S.K.; Madaio, M.P. Anti-DNA antibodies form immune deposits at distinct glomerular and vascular sites. Kidney Int. 1992, 41, 1690–1700. [Google Scholar] [CrossRef]
- Han, X.; Uchida, K.; Jurickova, I.; Koch, D.; Willson, T.; Samson, C.; Bonkowski, E.; Trauernicht, A.; Kim, M.-O.; Tomer, G.; et al. Granulocyte-macrophage colony-stimulating factor autoantibodies in murine ileitis and progressive ileal crohn’s disease. Gastroenterology 2009, 136, 1261–1271.e3. [Google Scholar] [CrossRef] [PubMed]
- Onuma, E.K.; Amenta, P.S.; Ramaswamy, K.; Lin, J.J.; Das, K.M. Autoimmunity in ulcerative colitis (uc): A predominant colonic mucosal b cell response against human tropomyosin isoform 5. Clin. Exp. Immunol. 2000, 121, 466–471. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef]
- Ting, Y.T.; Petersen, J.; Ramarathinam, S.H.; Scally, S.W.; Loh, K.L.; Thomas, R.; Suri, A.; Baker, D.G.; Purcell, A.W.; Reid, H.H.; et al. The interplay between citrullination and hla-drb1 polymorphism in shaping peptide binding hierarchies in rheumatoid arthritis. J. Biol. Chem. 2018, 293, 3236–3251. [Google Scholar] [CrossRef]
- Lu, D.R.; McDavid, A.N.; Kongpachith, S.; Lingampalli, N.; Glanville, J.; Ju, C.H.; Gottardo, R.; Robinson, W.H. T cell-dependent affinity maturation and innate immune pathways differentially drive autoreactive b cell responses in rheumatoid arthritis. Arthritis Rheumatol. 2018, 70, 1732–1744. [Google Scholar] [CrossRef]
- Jourdan, M.; Caraux, A.; De Vos, J.; Fiol, G.; Larroque, M.; Cognot, C.; Bret, C.; Duperray, C.; Hose, D.; Klein, B. An in vitro model of differentiation of memory b cells into plasmablasts and plasma cells including detailed phenotypic and molecular characterization. Blood 2009, 114, 5173–5181. [Google Scholar] [CrossRef] [PubMed]
- Enoksson, S.L.; Grasset, E.K.; Hägglöf, T.; Mattsson, N.; Kaiser, Y.; Gabrielsson, S.; McGaha, T.L.; Scheynius, A.; Karlsson, M.C.I. The inflammatory cytokine il-18 induces self-reactive innate antibody responses regulated by natural killer t cells. Proc. Natl. Acad. Sci. USA 2011, 108, E1399–E1407. [Google Scholar] [CrossRef]
- Boutet, M.A.; Najm, A.; Bart, G.; Brion, R.; Touchais, S.; Trichet, V.; Layrolle, P.; Gabay, C.; Palmer, G.; Blanchard, F.; et al. Il-38 overexpression induces anti-inflammatory effects in mice arthritis models and in human macrophages in vitro. Ann. Rheum. Dis. 2017, 76, 1304–1312. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Yasue, T.; Miyake, K.; Kimoto, M.; Takatsu, K. Cd38 ligation induces tyrosine phosphorylation of bruton tyrosine kinase and enhanced expression of interleukin 5-receptor alpha chain: Synergistic effects with interleukin 5. Proc. Natl. Acad. Sci. USA 1995, 92, 11814–11818. [Google Scholar] [CrossRef] [PubMed]
- Pittner, B.T.; Shanafelt, T.D.; Kay, N.E.; Jelinek, D.F. Cd38 expression levels in chronic lymphocytic leukemia b cells are associated with activation marker expression and differential responses to interferon stimulation. Leukemia 2005, 19, 2264–2272. [Google Scholar] [CrossRef]
- Camponeschi, A.; Kläsener, K.; Sundell, T.; Lundqvist, C.; Manna, P.T.; Ayoubzadeh, N.; Sundqvist, M.; Thorarinsdottir, K.; Gatto, M.; Visentini, M.; et al. Human cd38 regulates b cell antigen receptor dynamic organization in normal and malignant b cells. J. Exp. Med. 2022, 219, e20220201. [Google Scholar] [CrossRef] [PubMed]
- Carriere, V.; Roussel, L.; Ortega, N.; Lacorre, D.-A.; Americh, L.; Aguilar, L.; Bouche, G.; Girard, J.-P. Il-33, the il-1-like cytokine ligand for st2 receptor, is a chromatin-associated nuclear factor in vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Werman, A.; Werman-Venkert, R.; White, R.; Lee, J.-K.; Werman, B.; Krelin, Y.; Voronov, E.; Dinarello, C.A.; Apte, R.N. The precursor form of il-1α is an intracrine proinflammatory activator of transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 2434–2439. [Google Scholar] [CrossRef]
- Bulau, A.-M.; Nold, M.F.; Li, S.; Nold-Petry, C.A.; Fink, M.; Mansell, A.; Schwerd, T.; Hong, J.; Rubartelli, A.; Dinarello, C.A.; et al. Role of caspase-1 in nuclear translocation of il-37, release of the cytokine, and il-37 inhibition of innate immune responses. Proc. Natl. Acad. Sci. USA 2014, 111, 2650–2655. [Google Scholar] [CrossRef]
- Talabot-Ayer, D.; Mermoud, L.; Borowczyk, J.; Drukala, J.; Wolnicki, M.; Modarressi, A.; Boehncke, W.H.; Brembilla, N.; Palmer, G. Interleukin-38 interacts with destrin/actin-depolymerizing factor in human keratinocytes. PLoS ONE 2019, 14, e0225782. [Google Scholar] [CrossRef]
- Lam, W.Y.; Bhattacharya, D. Metabolic links between plasma cell survival, secretion, and stress. Trends Immunol. 2018, 39, 19–27. [Google Scholar] [CrossRef]
- de Graaf, D.M.; Teufel, L.U.; Joosten, L.A.B.; Dinarello, C.A. Interleukin-38 in health and disease. Cytokine 2022, 152, 155824. [Google Scholar] [CrossRef]
- Diaz-Barreiro, A.; Huard, A.; Palmer, G. Multifaceted roles of il-38 in inflammation and cancer. Cytokine 2022, 151, 155808. [Google Scholar] [CrossRef]
- Maeda, K.; Mehta, H.; Drevets, D.A.; Coggeshall, K.M. Il-6 increases b-cell igg production in a feed-forward proinflammatory mechanism to skew hematopoiesis and elevate myeloid production. Blood 2010, 115, 4699–4706. [Google Scholar] [CrossRef] [PubMed]
- Hipp, N.; Symington, H.; Pastoret, C.; Caron, G.; Monvoisin, C.; Tarte, K.; Fest, T.; Delaloy, C. Il-2 imprints human naive b cell fate towards plasma cell through erk/elk1-mediated bach2 repression. Nat. Commun. 2017, 8, 1443. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Hirohata, S. The role of il-10 in human b cell activation, proliferation, and differentiation. J. Immunol. 1995, 154, 4341–4350. [Google Scholar] [CrossRef] [PubMed]
- Smulski, C.R.; Eibel, H. Baff and baff-receptor in b cell selection and survival. Front. Immunol. 2018, 9, 2285. [Google Scholar] [CrossRef] [PubMed]
- Nemazee, D. Mechanisms of central tolerance for b cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef]
- Rafiq, K.; Bergtold, A.; Clynes, R. Immune complex–mediated antigen presentation induces tumor immunity. J. Clin. Investig. 2002, 110, 71–79. [Google Scholar] [CrossRef]
- Shlomchik, M.J. Sites and stages of autoreactive b cell activation and regulation. Immunity 2008, 28, 18–28. [Google Scholar] [CrossRef]
- Hoyer, B.F.; Moser, K.; Hauser, A.E.; Peddinghaus, A.; Voigt, C.; Eilat, D.; Radbruch, A.; Hiepe, F.; Manz, R.A. Short-lived plasmablasts and long-lived plasma cells contribute to chronic humoral autoimmunity in nzb/w mice. J. Exp. Med. 2004, 199, 1577–1584. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Provider | Antibody | Clone | Provider |
|---|---|---|---|---|---|
| Mouse | Human | ||||
| CD45-Vioblue | 30F11 | Miltenyi | CD20-BV510 | 2H7 | BioLegend |
| CD11b-BV605 | M170 | BioLegend | IgD-FITC | IA6-2 | BioLegend |
| CD19-APC-Fire750 | 6D5 | BioLegend | CD40-BV605 | 5C3 | BioLegend |
| CD38-BV510 | 90 | BD | CD69-PE-Cy7 | FN50 | BioLegend |
| CD138-PE | 281-2 | BD | CD80-BV711 | L307.4 | BD |
| MHCII-APC | M5/114.15.2 | Miltenyi | CD83-PE | HB15 | BioLegend |
| CD3-PE-CF594 | 145-2C11 | BD | CD86-FITC | BU63 | Immunotools |
| MHCII-APC | L243 | BioLegend | |||
| CD27-BV711 | O323 | BioLegend | |||
| CD38-BV421 | HIT2 | BD | |||
| CD138-PE-Cy7 | MI15 | BioLegend |
| Primer | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| IL1R1 | ATGAAATTGATGTTCGTCCCTGT | ACCACGCAATAGTAATGTCCTG |
| IL1RL2 | TCCCGAAGAGTTGTGTTTTGG | TGAGTGTGTCAGTATGGCTTGA |
| IL1RAPL1 | ATGAAAGCTCCGATTCCACAC | TTTGGGCAAGGGAGTAATTTGT |
| PRDM1 | AAGCAACTGGATGCGCTATGT | GGGATGGGCTTAATGGTGTAGAA |
| XBP1 | CCCTCCAGAACATCTCCCCAT | ACATGACTGGGTCCAAGTTGT |
| PAX5 | ACTTGCTCATCAAGGTGTCAG | TCCTCCAATTACCCCAGGCTT |
| IRF4 | GCTGATCGACCAGATCGACAG | CGGTTGTAGTCCTGCTTGC |
| RPS27A | CTGGAAGATGGACGTACTTTGTC | CGACGAAGGCGACTAATTTTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huard, A.; Wilmes, C.; Kiprina, A.; Netzer, C.; Palmer, G.; Brüne, B.; Weigert, A. Cell Intrinsic IL-38 Affects B Cell Differentiation and Antibody Production. Int. J. Mol. Sci. 2023, 24, 5676. https://doi.org/10.3390/ijms24065676
Huard A, Wilmes C, Kiprina A, Netzer C, Palmer G, Brüne B, Weigert A. Cell Intrinsic IL-38 Affects B Cell Differentiation and Antibody Production. International Journal of Molecular Sciences. 2023; 24(6):5676. https://doi.org/10.3390/ijms24065676
Chicago/Turabian StyleHuard, Arnaud, Christian Wilmes, Anastasiia Kiprina, Christoph Netzer, Gaby Palmer, Bernhard Brüne, and Andreas Weigert. 2023. "Cell Intrinsic IL-38 Affects B Cell Differentiation and Antibody Production" International Journal of Molecular Sciences 24, no. 6: 5676. https://doi.org/10.3390/ijms24065676
APA StyleHuard, A., Wilmes, C., Kiprina, A., Netzer, C., Palmer, G., Brüne, B., & Weigert, A. (2023). Cell Intrinsic IL-38 Affects B Cell Differentiation and Antibody Production. International Journal of Molecular Sciences, 24(6), 5676. https://doi.org/10.3390/ijms24065676