
Review of Technological Challenges in Personalised Medicine and Early Diagnosis of Neurodegenerative Disorders

 , , , , and
, , , , and
Abstract
1. Introduction

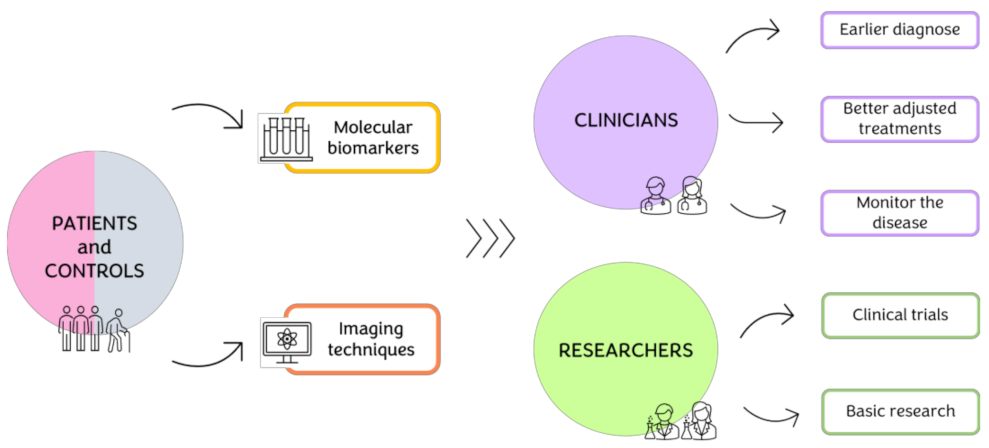{kind=link}
| Neurodegenerative Disease | Brain Origin | Physiological Characteristics | Most Vulnerable Neurons | Main Symptoms |
|---|---|---|---|---|
| Alzheimer’s disease | Brainstem nuclei, locus coeruleus, transentorhinal region, and olfactory bulb [10] | Hyperphosphorylated tau protein, β-amyloid plaques, neurofibrillary tangles, and neuritic plaques [11,12] | Cholinergic groups of neurons [13] | Noncognitive symptoms: aphasia, apraxia, agnosia, visuospatial deficits, difficulties in daily routine, and sleep disorders. Cognitive symptoms: loss of episodic memory, timing and spatial disorientation, and mood disorders [12,14] |
| Parkinson’s disease | Dorsal IX/X motor nucleus and olfactory bulb [12] | Misfolded α-synuclein, β -sheet amyloid aggregations, Lewy bodies, and Lewy neurites [15] | Noradrenergic neurons, dopaminergic neurons [12,15], especially in the substantia nigra pars compacta [16] | Nonmotor symptoms: constipation, visual defects, psychiatric symptoms, cognitive decline, and sleep disturbances. Motor symptoms: tremor, rigidity, bradykinesia/akinesia/hypokinesia, and gait disturbances [15] |
| Lewy body dementia | Dorsal IX/X motor nucleus, olfactory bulb, and striatum [16] | Hyperphosphorylated tau, Aβ aggregations, misfolded α-synuclein, βsheet amyloid aggregations, Lewy bodies, and Lewy neurites [15,16,17] | Dopaminergic neurons and cholinergic neurons [13,15,18] | Nonmotor symptoms: cognitive impairments, psychiatric symptoms, sleep disturbances, and hallucinations. Motor symptoms: bradykinesia, gait disturbances, and hyposmia [16] |
| Multiple system atrophy | Striatonigral and olivopontocerebellar neural and oligodendroglial nucleus [19] | α-synuclein inclusions in Papp–Lantos bodies and glial nuclear, neural cytoplasmic, neural nuclear, and astroglial cytoplasmic inclusions, astrogliosis, and reduced myelination [18] | Oligodendrocytes, dopaminergic neurons, and cerebellar neurons [18,19] | Autonomic dysfunctions: erectile dysfunction, lower urinary and cardiovascular symptoms, and dysphonia. Motor dysfunctions: tremor, ataxia, postural instability, and oculomotor abnormalities [18] |
| Amyotrophic lateral sclerosis | Brainstem, spinal cord, and skeletal muscles [20] | Aggregations of DNA-binding protein 43 (TDA-43) (in major cases), P-62 aggregations, and misfolded superoxide dismutase [20] | Upper and lower motor neurons [20] | Skeletal and motor cortex atrophy: weakness, cramps, muscle wasting; dysarthria, frontotemporal dementia, and cardiovascular failures [20] |
| Multiple sclerosis | Periventricular area, cortical area, infratentorial area, pons, and spinal cord [21] | Dysregulation of immunological T-cell response that leads to increased inflammation, demyelination, and axonal loss [22] | Oligodendrocytes, motor neurons [22] | Visual failures, spastic paraparesis, ataxia [22], urinary and/or faecal sphincter dysfunction, and tremor [21] |
| Huntington’s disease | Striatum (caudate and putamen), globus pallidus, and nucleus accumbens [23] | CAG triplet accumulations resulting in increasing misfolded huntingtin [23] | Medium spiny neurons of the striatum [24] | Chorea, myoclonus, tics, dystonia, gait, low emotional recognition, and impaired short-term memory, frontal/subcortical impairment [22] |
| Creutzfeldt–Jakob disease | Neocortex, striatum, thalamus, and cerebellar cortex [7] | Prion protein depositions, gliosis, neuropil rarefaction, neuronal loss, and spongiform alterations [7] | Neurons from the neocortex, striatum, thalamus, subiculum, quadrigeminal body, substantia nigra, pontine nucleus, and inferior olivary nucleus [7] | Memory disturbances, myoclonus, visual symptoms, akinetic mutism state, and cognitive impairment [7] |
1.1. Diagnostic Techniques
1.2. Alzheimer’s Disease and Parkinson’s Disease
2. Diagnostic Methods in Parkinson’s Disease and Alzheimer’s Disease
2.1. Neuroimaging Methods
2.1.1. Magnetic Resonance Imaging
Magnetic Resonance Imaging in Alzheimer’s Disease
- Structural Mangnetic Resonance Imaging
- Functional Magnetic Resonance Imaging
Magnetic Resonance Imaging in Parkinson’s Disease
- Structural Magnetic Resonance Imaging
2.1.2. Positron Emission Tomography
Positron Emission Tomography in Alzheimer’s Disease
- [18F] FDG-Positron Emission Tomography
- Amyloid beta-Positron Emission Tomography
- Tau-Positron Emission Tomography
Positron Emission Tomography in Parkinson’s Disease
- Dopamine Transporter Imaging
- Other Neurotransmitters
- Brain–Gut Denervation
- α-Synuclein
2.2. Molecular Methods
2.2.1. Biomarkers in Peripheral Fluids
Biomarkers in Alzheimer′s Disease
Biomarkers in Parkinson´s Disease
2.2.2. Inflammation in Neurodegenerative Diseases
2.2.3. Exosomes
Exosomes in Neurodegenerative Diseases
Exosomes as Biomarkers
2.2.4. Autoantibodies
Autoantibodies in Neurodegenerative Diseases
Autoantibodies as Biomarkers
3. Future Challenges in Personalised Diagnostics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kovacs, G.G. Concepts and Classification of Neurodegenerative Diseases, 1st ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2017; Volume 145, ISBN 9780128023952. [Google Scholar]
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Pritzkow, S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Sehar, U.; Rawat, P.; Reddy, A.P.; Kopel, J.; Reddy, P.H. Amyloid Beta in Aging and Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 12924. [Google Scholar] [CrossRef] [PubMed]
- Bisaglia, M.; Mammi, S.; Bubacco, L. Structural insights on physiological functions and pathological effects of a-synuclein. FASEB J. 2009, 23, 329–340. [Google Scholar] [CrossRef]
- Peng, C.; Trojanowski, J.Q.; Lee, V.M. Protein transmission in neurodegenerative disease. Nat. Rev. Neurol. 2020, 16, 199–212. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Jakob, A.M. Symposium: Fundamentals learned from diversity among typical and atypical appearances Creutzfeldt-Jakob disease. Neuropathology 2016, 37, 174–188. [Google Scholar] [CrossRef]
- Armstrong, R. What causes neurodegenerative disease? Folia Neuropathogica 2020, 58, 93–112. [Google Scholar] [CrossRef]
- Mumtaz, S.; Rana, J.N.; Choi, E.H.; Han, I. Microwave Radiation and the Brain: Mechanisms, Current Status, and Future Prospects. Int. J. Mol. Sci. 2022, 23, 9288. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Analysis of knee infrared image based on sample entropy algorithm. In Proceedings of the 2015 8th International Conference on Intelligent Networks and Intelligent Systems (ICINIS), Tianjin, China, 1–3 November 2015. [Google Scholar] [CrossRef]
- Soria-Lopez, J.A.; Gonzalez, H.M.; Leger, G.C. Alzheimer’s disease. In Handbook of Clinical Neurology; Elsevier B.V.: Amsterdam, The Netherlands, 2019; Volume 167, ISBN 9780128047668. [Google Scholar]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. The preclinical phase of the pathological process underlying sporadic Alzheimer’s disease. Brain 2015, 138, 2814–2833. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. Potencial Pathways of Abnormal Tau and a-synuclein dissemination in Sporadic Alzheimer’s and Parkinson´s Diseases. Cold Spring Harb. Perspect. Biol. 2016, 8, a023630. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Jellinger, K.A.; Korczyn, A.D. Are dementia with Lewy bodies and Parkinson’s disease dementia the same disease ? BMC Med. 2018, 16, 34. [Google Scholar] [CrossRef]
- Taylor, J.; Burn, D.J. New evidence on the management of Lewy body dementia. Lancet Neurol. 2020, 19, 157–169. [Google Scholar] [CrossRef]
- Jellinger, K.A. Multiple System Atrophy: An Oligodendroglioneural Synucleinopathy. J. Alzheimer’s Dis. 2018, 62, 1141–1179. [Google Scholar] [CrossRef]
- Fanciulli, A.; Stankovic, I.; Krismer, F.; Seppi, K.; Levin, J.; Wenning, G.K. Multiple System Atrophy; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 149. [Google Scholar]
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J.; Simmons, Z.; van den Berg, L. Amyotrophic lateral sclerosis. In Xpharm: The Comprehensive Pharmacology Reference; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–5. [Google Scholar] [CrossRef]
- Oh, J.; Vidal-jordana, A.; Montalban, X. Multiple sclerosis: Clinical aspects. Curr. Opin. Neurol. 2018, 31, 752–759. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G.; Road, W.; Unit, P.N.; Square, C. Multiple Sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef]
- Ghosh, R.; Tabrizi, S.J. Huntington Disease, 1st ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; Volume 147, ISBN 9780444632333. [Google Scholar]
- McColgan, P.; Tabrizi, S.J. Huntington’s disease: A clinical review. Eur. J. Neurol. 2018, 38, 42–49. [Google Scholar] [CrossRef]
- Jeromin, A.; Bowser, R. Biomarkers in Neurodegenerative Diseases. In Neurodegeneratives Diseases; Springer: Berlin/Heidelberg, Germany, 2017; pp. 491–528. ISBN 9783319571935. [Google Scholar]
- Erkkinen, M.G.; Kim, M.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef]
- Annavarapu, R.N.; Kathi, S.; Krishna, V.V. Non-Invasive Imaging Modalities to Study Neurodegenerative Diseases of Aging Brain. J. Chem. Neuroanat. 2018, 95, 54–69. [Google Scholar] [CrossRef]
- Kamagata, K.; Andica, C.; Kato, A.; Saito, Y.; Uchida, W.; Hatano, T.; Lukies, M.; Ogawa, T.; Takeshige-amano, H.; Akashi, T.; et al. Diffusion Magnetic Resonance Imaging-Based Biomarkers for Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 5216. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.J.; Kirilina, E.; Otaduy, M.C.G.; Ivanov, D.; Acosta-Cabronero, J.; Callaghan, M.F.; Lambert, C.; Cardenas-Blanco, A.; Pine, K.; Passamonti, L.; et al. Locus coeruleus imaging as a biomarker for noradrenergic dysfunction in neurodegenerative diseases. Brain 2019, 142, 2558–2571. [Google Scholar] [CrossRef] [PubMed]
- Doroszkiewicz, J.; Groblewska, M. Molecular Biomarkers and Their Implications for the Early Diagnosis of Selected Neurodegenerative —Diseases. Int. J. Mol. Sci. 2022, 23, 4610. [Google Scholar] [CrossRef] [PubMed]
- Clarimon, J.; Moreno-grau, S.; Cervera-carles, L.; Dols-icardo, O.; Suenchez-juan, P.; Ruiz, A. Genetic architecture of neurodegenerative dementias. Neuropharmacology 2020, 168, 108014. [Google Scholar] [CrossRef]
- Blauwendraat, C.; Nalls, M.A.; Singleton, A.B. The genetic architecture of Parkinson’s disease. Lancet Neurol. 2020, 19, 170–178. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease . Molecules 2020, 25, 5789. [Google Scholar] [CrossRef]
- Young, P.N.E.; Estarellas, M.; Coomans, E.; Srikrishna, M.; Beaumont, H.; Maass, A.; Venkataraman, A.V.; Lissaman, R.; Jiménez, D.; Betts, M.J.; et al. Imaging biomarkers in neurodegeneration: Current and future practices. Alzheimer’s Res. Ther. 2020, 12, 49. [Google Scholar] [CrossRef]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef]
- Ridha, B.H.; Barnes, J.; Bartlett, J.W.; Godbolt, A.; Pepple, T.; Rossor, M.N.; Fox, N.C. Tracking atrophy progression in familial Alzheimer’s disease: A serial MRI study. Lancet Neurol. 2006, 5, 828–834. [Google Scholar] [CrossRef]
- Dickerson, B.C.; Bakkour, A.; Salat, D.H.; Feczko, E.; Pacheco, J.; Greve, D.N.; Grodstein, F.; Wright, C.I.; Blacker, D.; Rosas, H.D.; et al. The cortical signature of Alzheimer’s disease: Regionally specific cortical thinning relates to symptom severity in very mild to mild AD dementia and is detectable in asymptomatic amyloid-positive individuals. Cereb. Cortex 2009, 19, 497–510. [Google Scholar] [CrossRef]
- Wang, J.; Ekin, A.; De Haan, G. Shape analysis of brain ventricles for improved classification of alzheimer’s patients. In Proceedings of the 2008 15th IEEE International Conference on Image Processing, San Diego, CA, USA, 12–15 October 2008; pp. 2252–2255. [Google Scholar] [CrossRef]
- Shim, H.S.; Ly, M.J.; Tighe, S.K. Brain Imaging in the Differential Diagnosis of Young-Onset Dementias. Psychiatr. Clin. N. Am. 2015, 38, 281–294. [Google Scholar] [CrossRef]
- Hohenfeld, C.; Werner, C.J.; Reetz, K. Resting-state connectivity in neurodegenerative disorders: Is there potential for an imaging biomarker? NeuroImage Clin. 2018, 18, 849–870. [Google Scholar] [CrossRef]
- Bookheimer, S.Y.; Strojwas, M.H.; Cohen, M.S.; Saunders, A.M.; Pericak-Vance, M.A.; Mazziotta, J.C.; Small, G.W. Patterns of Brain Activation in People at Risk for Alzheimer’s Disease. N. Engl. J. Med. 2000, 343, 450–456. [Google Scholar] [CrossRef]
- Foster, C.M.; Kennedy, K.M.; Morn, M.M.; Hoagey, D.A.; Rodrigue, K.M. Both hyper- and hypo-activation to cognitive challenge are associated with increased beta-amyloid deposition in healthy aging: A nonlinear effect. Neuroimage 2017, 43, 909–932. [Google Scholar] [CrossRef]
- Chau, M.T.; Todd, G.; Wilcox, R.; Agzarian, M.; Bezak, E. Diagnostic accuracy of the appearance of Nigrosome-1 on magnetic resonance imaging in Parkinson’s disease: A systematic review and meta-analysis. Park. Relat. Disord. 2020, 78, 12–20. [Google Scholar] [CrossRef]
- Martin-Bastida, A.; Pietracupa, S.; Piccini, P. Neuromelanin in parkinsonian disorders: An update. Int. J. Neurosci. 2017, 127, 1116–1123. [Google Scholar] [CrossRef]
- Ofori, E.; Pasternak, O.; Planetta, P.; Burciu, R.; Snyder, A.; Febo, M.; Golde, T.; Okun, M.; Vaillancourt, D. Increased free-water in the substantia nigra of Parkinson´s disease: A single-site and multi-site study. Neurobiol. Aging 2015, 36, 1097–1104. [Google Scholar] [CrossRef]
- Pavese, N.; Tai, Y.F. Nigrosome Imaging and Neuromelanin Sensitive MRI in Diagnostic Evaluation of Parkinsonism. Mov. Disord. Clin. Pract. 2018, 5, 131–140. [Google Scholar] [CrossRef]
- Mangesius, S.; Hussl, A.; Krismer, F.; Mahlknecht, P.; Reiter, E.; Tagwercher, S.; Djamshidian, A.; Schocke, M.; Esterhammer, R.; Wenning, G.; et al. MR planimetry in neurodegenerative parkinsonism yields high diagnostic accuracy for PSP. Park. Relat. Disord. 2018, 46, 47–55. [Google Scholar] [CrossRef]
- Nestor, P.J.; Altomare, D.; Festari, C.; Drzezga, A.; Rivolta, J.; Walker, Z.; Bouwman, F.; Orini, S.; Law, I.; Agosta, F.; et al. Clinical utility of FDG-PET for the differential diagnosis among the main forms of dementia. Eur. J. Nucl. Med. Mol. Imaging 2018, 45, 1509–1525. [Google Scholar] [CrossRef]
- Kawasaki, K.; Ishii, K.; Saito, Y.; Oda, K.; Kimura, Y.; Ishiwata, K. Influence of mild hyperglycemia on cerebral FDG distribution patterns calculated by statistical parametric mapping. Ann. Nucl. Med. 2008, 22, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol. Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef] [PubMed]
- Klunk, W.E.; Koeppeb, R.A.; Pricec, J.C.; Benzingerd, T.; Devous, M.D., Sr.; Jagust, W.J.; Johnson, K.A.; Mathis, C.A.; Minhas, D.; Pontecorvo, M.J.; et al. The Centiloid Project: Standardizing Quantitative Amyloid Plaque Estimation by PET William. Alzheimers Dement. 2015, 11, 1–15.e4. [Google Scholar] [CrossRef] [PubMed]
- Ikonomovic, M.D.; Klunk, W.E.; Abrahamson, E.E.; Mathis, C.A.; Price, J.C.; Tsopelas, N.D.; Lopresti, B.J.; Ziolko, S.; Bi, W.; Paljug, W.R.; et al. Post-mortem correlates of in vivo PiB-PET amyloid imaging in a typical case of Alzheimer’s disease. Brain 2008, 131, 1630–1645. [Google Scholar] [CrossRef] [PubMed]
- Ossenkoppele, R.; Schonhaut, D.R.; Schöll, M.; Lockhart, S.N.; Ayakta, N.; Baker, S.L.; O’Neil, J.P.; Janabi, M.; Lazaris, A.; Cantwell, A.; et al. Tau PET patterns mirror clinical and neuroanatomical variability in Alzheimer’s disease. Brain 2016, 139, 1551–1567. [Google Scholar] [CrossRef]
- Ossenkoppele, R.; Rabinovici, G.D.; Smith, R.; Cho, H.; Scholl, M.; Strandberg, O.; Palmqvist, S.; Mattsson, N.; Janelidze, S.; Santillo, A.; et al. Discriminative accuracy of [18F]flortaucipir positron emission tomography for Alzheimer disease vs other neurodegenerative disorders. JAMA 2018, 320, 1151–1162. [Google Scholar] [CrossRef]
- Marquié, M.; Normandin, M.D.; Meltzer, A.C.; Tick, M.S.; Andrea, N.V.; Antón-fernández, A.; Klunk, W.E.; Mathis, C.A.; Ikonomovic, M.D.; Debnath, M.; et al. Pathologic correlations of [F-18]-AV-1451 imaging in non-Alzheimer tauopathies. Ann. Neurol. 2018, 81, 117–128. [Google Scholar] [CrossRef]
- Miyamoto, T.; Miyamoto, M.; Numahata, K.; Onoue, H.; Akaiwa, Y.; Sairenchi, T. Reduced dopamine transporter binding predicts early transition to Lewy body disease in Japanese patients with idiopathic rapid eye movement sleep behavior disorder. J. Neurol. Sci. 2020, 414, 116821. [Google Scholar] [CrossRef]
- Prange, S.; Theis, H.; Banwinkler, M.; van Eimeren, T. Molecular Imaging in Parkinsonian Disorders—What’s New and Hot? Brain Sci. 2022, 12, 1146. [Google Scholar] [CrossRef]
- Horsager, J.; Andersen, K.B.; Knudsen, K.; Skjærbæk, C.; Fedorova, T.D.; Okkels, N.; Schaeffer, E.; Bonkat, S.K.; Geday, J.; Otto, M.; et al. Brain-first versus body-first Parkinson’s disease: A multimodal imaging case-control study. Brain 2020, 143, 3077–3088. [Google Scholar] [CrossRef]
- Capotosti, F.; Vokali, E.; Molette, J.; Ravache, M.; Delgado, C.; Kocher, J.; Pittet, L.; Dimitrakopoulos, I.K.; Di-Bonaventura, I.; Touilloux, T.; et al. The development of [18 F]ACI-12589, a high affinity and selective alpha-synuclein radiotracer, as a biomarker for Parkinson’s disease and other synucleinopathies. Alzheimer’s Dement. 2021, 17, 17–18. [Google Scholar] [CrossRef]
- Teunissen, C.E.; Verberk, I.M.W.; Thijssen, E.H.; Vermunt, L.; Hansson, O.; Zetterberg, H.; van der Flier, W.M.; Mielke, M.M.; del Campo, M. Blood-based biomarkers for Alzheimer’s disease: Towards clinical implementation. Lancet Neurol. 2022, 21, 66–77. [Google Scholar] [CrossRef]
- West, T.; Kirmess, K.M.; Meyer, M.R.; Holubasch, M.S.; Knapik, S.S.; Hu, Y.; Contois, J.H.; Jackson, E.N.; Harpstrite, S.E.; Bateman, R.J.; et al. A blood-based diagnostic test incorporating plasma Aβ42/40 ratio, ApoE proteotype, and age accurately identifies brain amyloid status: Findings from a multi cohort validity analysis. Mol. Neurodegener. 2021, 16, 30. [Google Scholar] [CrossRef]
- Stockmann, J.; Verberk, I.M.W.; Timmesfeld, N.; Denz, R.; Budde, B.; Lange-Leifhelm, J.; Scheltens, P.; van der Flier, W.M.; Nabers, A.; Teunissen, C.E.; et al. Correction to: Amyloid-β misfolding as a plasma biomarker indicates risk for future clinical Alzheimer’s disease in individuals with subjective cognitive decline. Alzheimer’s Res. Ther. 2021, 13, 25. [Google Scholar] [CrossRef]
- Karikari, T.K.; Benedet, A.L.; Ashton, N.J.; Lantero Rodriguez, J.; Snellman, A.; Suárez-Calvet, M.; Saha-Chaudhuri, P.; Lussier, F.; Kvartsberg, H.; Rial, A.M.; et al. Diagnostic performance and prediction of clinical progression of plasma phospho-tau181 in the Alzheimer’s Disease Neuroimaging Initiative. Mol. Psychiatry 2021, 26, 429–442. [Google Scholar] [CrossRef]
- Thijssen, E.H.; Verberk, I.M.W.; Vanbrabant, J.; Koelewijn, A.; Heijst, H.; Scheltens, P.; van der Flier, W.; Vanderstichele, H.; Stoops, E.; Teunissen, C.E. Highly specific and ultrasensitive plasma test detects Abeta(1–42) and Abeta(1–40) in Alzheimer’s disease. Sci. Rep. 2021, 11, 9736. [Google Scholar] [CrossRef]
- Mielke, M.M.; Hagen, C.E.; Xu, J.; Chai, X.; Vemuri, P.; Lowe, V.J.; Airey, D.C.; Knopman, D.S.; Roberts, R.O.; Machulda, M.M.; et al. Plasma phospho-tau181 increases with Alzheimer’s disease clinical severity and is associated with tau-PET and amyloid-PET. Alzheimers Dement. 2018, 14, 989–997. [Google Scholar] [CrossRef]
- Thijssen, E.H.; La Joie, R.; Wolf, A.; Storm, A.; Wang, P.; Laccarino, L.; Bourakova, V.; Cobigo, Y.; Heuer, H.; Spina, S.; et al. Diagnostic value of plasma phosphorylated tau181 in Alzheimer’s disease and frontotemporal lobar degeneration. Nat. Med. 2020, 26, 387–397. [Google Scholar] [CrossRef]
- Moscoso, A.; Grothe, M.J.; Ashton, N.J.; Karikari, T.K.; Lantero-Rodriguez, J.; Snellman, A.; Suarez-Calvet, M.; Blennow, K.; Zetterberg, H.; Schöll, M. Longitudinal Associations of Blood Phosphorylated Tau181 and Neurofilamente Light Chain with Neurodegeneration in Alzheimer Disease. JAMA Neurol. 2021, 78, 396–406. [Google Scholar] [CrossRef]
- Karikari, T.K.; Pascoal, T.A.; Ashton, N.J.; Janelidze, S.; Benedet, A.L.; Rodriguez, J.L.; Chamoun, M.; Savard, M.; Kang, M.S.; Therriault, J.; et al. Blood phosphorylated tau 181 as a biomarker for Alzheimer’s disease: A diagnostic performance and prediction modelling study using data from four prospective cohorts. Lancet Neurol. 2020, 19, 422–433. [Google Scholar] [CrossRef]
- Janelidze, S.; Mattsson, N.; Palmqvist, S.; Smith, R.; Beach, T.G.; Serrano, G.E.; Chai, X.; Proctor, N.K.; Eichenlaub, U.; Zetterberg, H.; et al. Plasma P-tau181 in Alzheimer’s disease: Relationship to other biomarkers, differential diagnosis, neuropathology and longitudinal progression to Alzheimer’s dementia. Nat. Med. 2020, 26, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Benussi, A.; Karikari, T.K.; Ashton, N.; Gazzina, S.; Premi, E.; Benussi, L.; Ghidoni, R.; Rodriguez, J.L.; Emeršič, A.; Simrén, J.; et al. Diagnostic and prognostic value of serum NfL and p-Tau181 in frontotemporal lobar degeneration. J. Neurol. Neurosurg. Psychiatry 2020, 91, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Lantero, J.; Thomas, R.; Marc, K.K.; Calvet, S.; Troakes, C.; King, A. Plasma p-tau181 accurately predicts Alzheimer’s disease pathology at least 8 years prior to post-mortem and improves the clinical characterisation of cognitive decline. Acta Neuropathol. 2020, 140, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, S.; Tideman, P.; Cullen, N.; Zetterberg, H.; Blennow, K.; Dage, J.L.; Stomrud, E.; Janelidze, S.; Mattsson-Carlgren, N.; Hansson, O. Prediction of future Alzheimer’s disease dementia using plasma phospho-tau combined with other accessible measures. Nat. Med. 2021, 27, 1034–1042. [Google Scholar] [CrossRef]
- Barth, N.R. Blood plasma phosphorylated-tau isoforms track CNS change in Alzheimer’s disease. J. Exp. Med. 2020, 217, e20200861. [Google Scholar]
- Janelidze, S.; Berron, D.; Smith, R.; Strandberg, O.; Proctor, N.K.; Dage, J.L.; Stomrud, E.; Palmqvist, S.; Mattson-Carlgren, N.; Hansson, O. Associations of Plasma Phospho-Tau217 Levels With Tau Positron Emission Tomograohy in Early Alzheimer Disease. JAMA Neurol. 2021, 78, 149–156. [Google Scholar] [CrossRef]
- Ashton, N.J.; Pascoal, T.A.; Karikari, T.K.; Benedet, A.L. Plasma p—tau231: A new biomarker for incipient Alzheimer’s disease pathology. Acta Neuropathol. 2021, 141, 709–724. [Google Scholar] [CrossRef]
- Mielke, M.M.; Frank, R.D.; Dage, J.L.; Jeromin, A.; Aston, N.; Blennow, K.; Karikari, T.K.; Vanmechelen, E.; Zetterberg, H.; Algeciras-Schimnich, A.; et al. Comparison of Plasma Phosphorylated Tau Species With Amyloid and Tau Positron Emission Tomography, Neurodegeneration, Vascular Pathology, and Cognitive Outcomes. JAMA Neurol. 2021, 78, 1108–1117. [Google Scholar] [CrossRef]
- Mattsson-carlgren, N.; Janelidze, S.; Palmqvist, S.; Cullen, N.; Svenningsson, A.L.; Strandberg, O.; Mengel, D.; Walsh, D.M.; Stomrud, E.; Dage, J.L.; et al. Longitudinal plasma p-tau217 is increased in early stages of Alzheimer’s disease. Brain 2020, 143, 3234–3241. [Google Scholar] [CrossRef]
- Gisslén, M.; Price, R.W.; Andreasson, U.; Norgren, N.; Nilsson, S.; Hagberg, L.; Fuchs, D.; Spudich, S.; Blennow, K.; Zetterberg, H. Plasma Concentration of the Neuro fi lament Light Protein (NFL) is a Biomarker of CNS Injury in HIV Infection: A Cross-Sectional Study. EBioMedicine 2016, 3, 135–140. [Google Scholar] [CrossRef]
- Bridel, C.; Van Wieringen, W.N.; Zetterberg, H.; Tijms, B.M.; Teunissen, C.E. Diagnostic Value of Cerebrospinal Fluid Neurofilament Light Protein in Neurology. JAMA Neurol. 2019, 76, 1035–1048. [Google Scholar] [CrossRef]
- Zetterberg, H.; Skillbäck, T.; Mattson, N.; Trojanowski, J.Q.; Portelius, E.; Shaw, L.M.; Weiner, M.W.; Blennow, K. Association of Cerebrospinal Fluid Neurofilament Light Concentration With Alzheimer Disease Progression. JAMA Neurol. 2016, 73, 60–67. [Google Scholar] [CrossRef]
- Mattsson-Carlgreen, N.; Cullen, N.C.; Andreassson, U.; Zetterberg, H.; Blennow, K. Association Between Longitudinal Plasma Neurofilament Light and Neurodegeneration in Patients With Alzheimer Disease. JAMA Neurol. 2019, 76, 791–799. [Google Scholar] [CrossRef]
- Verberk, I.M.W.; Thijssen, E.; Koelewijn, J.; Mauroo, K.; Vanbrabant, J.; De Wilde, A.; Zwan, M.D.; Verfaillie, S.C.J.; Ossenkoppele, R.; Barkhof, F.; et al. Combination of plasma amyloid protein strongly associates with cerebral amyloid pathology. Alzheimer’s Res. Ther. 2020, 7, 118. [Google Scholar] [CrossRef]
- Chatterjee, P.; Pedrini, S.; Stoops, E.; Goozee, K.; Villemagne, V.L.; Asih, P.R. Plasma glial fi brillary acidic protein is elevated in cognitively normal older adults at risk of Alzheimer’s disease. Transl. Psychiatry 2021, 11, 27. [Google Scholar] [CrossRef]
- Oeckl, P.; Halbgebauer, S.; Anderl-Straub, S.; Steinacker, P.; Huss, A.M.; Neugebauer, H.; von Arnim, C.A.; Diehl-Schmid, J.; Grimmer, T.; Kornhuber, J.; et al. Glial Fibrillary Acidic Protein in Serum is Increased in Alzheimer’s Disease and Correlates with Cognitive Impairment. J. Alzheimer’s Dis. 2019, 67, 481–488. [Google Scholar] [CrossRef]
- Swarbrick, S.; Wragg, N.; Ghosh, S.; Stolzing, A. Systematic Review of miRNA as Biomarkers in Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 6156–6167. [Google Scholar] [CrossRef]
- Magalhães, P.; Lashuel, H.A. Opportunities and challenges of alpha-synuclein as a potential biomarker for Parkinson’s disease and other synucleinopathies. NPJ Park. Dis. 2022, 8, 9. [Google Scholar] [CrossRef]
- Parnetti, L.; Gaetani, L.; Eusebi, P.; Paciotti, S.; Hansson, O.; El-Agnaf, O.; Mollenhauer, B.; Blennow, K.; Calabresi, P. CSF and blood biomarkers for Parkinson’s disease. Lancet Neurol. 2019, 18, 573–586. [Google Scholar] [CrossRef]
- Mollenhauer, B.; Bowman, F.D.B.; Drake, D.; Duong, J.; Blennow, K.; El-Agnaf, O.; Shaw, L.M.; Masucci, J.; Taylor, P.; Umek, R.M.; et al. Antibody-based methods for the measurement of α-synuclein concentration in human cerebrospinal fluid—Method comparison and round robin study. J. Neurochem. 2019, 149, 126–138. [Google Scholar] [CrossRef]
- Hall, S.; Surova, Y.; Öhrfelt, A.; Zetterberg, H.; Lindqvist, D.; Hansson, O. CSF biomarkers and clinical progression of Parkinson disease. Neurology 2015, 84, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Fairfoul, G.; McGuire, L.I.; Pal, S.; Ironside, J.W.; Neumann, J.; Christie, S.; Joachim, C.; Esiri, M.; Evetts, S.G.; Rolinski, M.; et al. Alpha-synuclein RT-QuIC in the CSF of patients with alpha-synucleinopathies. Ann. Clin. Transl. Neurol. 2016, 3, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Shahnawaz, M.; Tokuda, T.; Waraga, M.; Mendez, N.; Ishii, R.; Trenkwalder, C.; Mollenhauer, B.; Soto, C. Development of a biochemical diagnosis of Parkinson disease by detection of α-synuclein misfolded aggregates in cerebrospinal fluid. JAMA Neurol. 2017, 74, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Majbour, N.K.; Vaikath, N.N.; Van Dijk, K.D.; Ardah, M.T.; Varghese, S.; Vesterager, L.B.; Montezinho, L.P.; Poole, S.; Safieh-Garabedian, B.; Tokuda, T.; et al. Oligomeric and phosphorylated alpha-synuclein as potential CSF biomarkers for Parkinson’s disease. Mol. Neurodegener. 2016, 11, 7. [Google Scholar] [CrossRef]
- Foulds, P.G.; Diggle, P.; Mitchell, J.D.; Parker, A.; Hasegawa, M.; Masuda-Suzukake, M.; Mann, D.M.A.; Allsop, D. A longitudinal study on a-synuclein in blood plasma as a biomarker for Parkinson’s disease. Sci. Rep. 2013, 3, 2540. [Google Scholar] [CrossRef]
- Cariulo, C.; Martufi, P.; Verani, M.; Azzollini, L.; Bruni, G.; Weiss, A.; Deguire, S.M.; Lashuel, H.A.; Scaricamazza, E.; Sancesario, G.M.; et al. Phospho-S129 Alpha-Synuclein Is Present in Human Plasma but Not in Cerebrospinal Fluid as Determined by an Ultrasensitive Immunoassay. Front. Neurosci. 2019, 13, 889. [Google Scholar] [CrossRef]
- Bougea, A.; Stefanis, L.; Paraskevas, G.P.; Emmanouilidou, E.; Vekrelis, K.; Kapaki, E. Plasma alpha-synuclein levels in patients with Parkinson’s disease: A systematic review and meta-analysis. Neurol. Sci. 2019, 40, 929–938. [Google Scholar] [CrossRef]
- Del Tredici, K.; Hawkes, C.H.; Ghebremedhin, E.; Braak, H. Lewy pathology in the submandibular gland of individuals with incidental Lewy body disease and sporadic Parkinson’s disease. Acta Neuropathol. 2010, 119, 703–713. [Google Scholar] [CrossRef]
- Bougea, A.; Koros, C.; Stefanis, L. Salivary alpha-synuclein as a biomarker for Parkinson’s disease: A systematic review. J. Neural Transm. 2019, 126, 1373–1382. [Google Scholar] [CrossRef]
- Al-Nimer, M.S.M.; Mshatat, S.F.; Abdulla, H.I. Saliva α-synuclein and a high extinction coeffi cient protein: A novel approach in assessment biomarkers of parkinson’s disease. N. Am. J. Med. Sci. 2014, 6, 633–637. [Google Scholar] [CrossRef]
- Devic, I.; Hwang, H.; Edgar, J.S.; Izutsu, K.; Presland, R.; Pan, C.; Goodlett, D.R.; Wang, Y.; Armaly, J.; Tumas, V.; et al. Salivary α-synuclein and DJ-1: Potential biomarkers for Parkinson’s disease. Brain 2011, 134, e178. [Google Scholar] [CrossRef]
- Goldman, J.G.; Andrews, H.; Amara, A.; Naito, A.; Alcalay, R.N.; Shaw, L.M.; Taylor, P.; Xie, T.; Tuite, P.; Henchcliffe, C.; et al. Cerebrospinal fluid, plasma, and saliva in the BioFIND study: Relationships among biomarkers and Parkinson’s disease Features. Mov. Disord. 2018, 33, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Vivacqua, G.; Suppa, A.; Mancinelli, R.; Belvisi, D.; Fabbrini, A.; Costanzo, M.; Formica, A.; Onori, P.; Fabbrini, G.; Berardelli, A. Salivary alpha-synuclein in the diagnosis of Parkinson’s disease and Progressive Supranuclear Palsy. Park. Relat. Disord. 2019, 63, 143–148. [Google Scholar] [CrossRef]
- Stewart, T.; Sui, Y.-T.; Gonzalez-Cuyar, L.F.; Wong, D.T.W.; Akin, D.M.; Tumas, V.; Aasly, J.; Ashmore, E.; Aro, P.; Ginghina, C.; et al. Cheek cell-derived a-synuclein and DJ-1 do not differentiate Parkinson´s disease from control. Neurobiol. Aging 2014, 35, 418–420. [Google Scholar] [CrossRef][Green Version]
- Kang, W.; Chen, W.; Yang, Q.; Zhang, L.; Zhang, L.; Wang, X.; Dong, F.; Zhao, Y.; Chen, S.; Quinn, T.J.; et al. Salivary total α-synuclein, oligomeric α-synuclein and SNCA variants in Parkinson’s disease patients. Sci. Rep. 2016, 6, 28143. [Google Scholar] [CrossRef]
- Goh, S.Y.; Chao, Y.X.; Dheen, S.T.; Tay, S.S.-W. Role of MicroRNAs in Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5649. [Google Scholar] [CrossRef]
- Bartels, A.L.; Willemsen, A.T.M.; Doorduin, J.; De Vries, E.F.J.; Dierckx, R.A.; Leenders, K.L. Parkinsonism and Related Disorders [11 C] -PK11195 PET: Quantification of neuroinflammation and a monitor of anti-inflammatory treatment in Parkinson’s disease? Park. Relat. Disord. 2010, 16, 57–59. [Google Scholar] [CrossRef]
- Brodacki, B.; Staszewski, J.; Toczyłowska, B.; Kozłowska, E. Neuroscience Letters Serum interleukin (IL-2, IL-10, IL-6, IL-4), TNF ␣, and INF ␥ concentrations are elevated in patients with atypical and idiopathic parkinsonism. Neurosci. Lett. 2008, 441, 158–162. [Google Scholar] [CrossRef]
- Reale, M.; Iarlori, C.; Thomas, A.; Gambi, D.; Perfetti, B.; Di Nicola, M.; Onofrj, M. Brain, Behavior, and Immunity Peripheral cytokines profile in Parkinson’s disease. Brain Behav. Immun. 2009, 23, 55–63. [Google Scholar] [CrossRef]
- Barnum, C.J.; Chen, X.; Chung, J.; Williams, M.; Grigoryan, N.; Tesi, R.J.; Tansey, M.G. Peripheral administration of the selective inhibitor of soluble Tumor Necrosis Factor (TNF) XPro1595 attenuates nigral cell loss and glial activation in 6-OHDA hemiparkinsonian rats. J. Park. Dis. 2014, 4, 349–360. [Google Scholar] [CrossRef]
- Mccoy, M.K.; Ruhn, K.A.; Martinez, T.N.; Mcalpine, F.E.; Tansey, M.G. Intranigral lentiviral delivery of dominant negative TNF attenuates neurodegeneration and behavioral deficits in hemiparkinsonian rats. Mol. Ther. 2008, 16, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.S.; Barnum, C.J.; Ruhn, K.A.; Varghese, S.; Treviño, I.; Blesch, A. Delayed Dominant-Negative TNF Gene Therapy Halts Progressive Loss of Nigral Dopaminergic Neurons in a Rat Model of Parkinson’s Disease. Mol. Ther. 2010, 19, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A.; Marxreiter, F. Th17 Lymphocytes Induce Neuronal Cell Death in a Human iPSC-Based Model of Parkinson’s Disease Short Article Th17 Lymphocytes Induce Neuronal Cell Death in a Human iPSC-Based Model of Parkinson’s Disease. Cell Stem Cell 2018, 23, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Alcalay, R.N.; Garretti, F.; Cote, L.; Kanter, E.; Agin-Liebes, J.P.; Liong, C.; McMurtrey, C.; Hildebrand, W.H.; Mao, X.; et al. T cells of Parkinson´s disease patients recognize a-synuclein peptides. Nature 2017, 546, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Yang, W.; Wei, H.; Dean, M.N.; Standaert, D.G. Dysregulation of the Adaptive Immune System in Patients With Early-Stage Parkinson Disease. Neurol. Neuroimmunol. Nueroinflamm. 2021, 8, e1036. [Google Scholar] [CrossRef]
- Thounaojam, M.C.; Kaushik, D.K.; Basu, A. MicroRNAs in the Brain: It’s Regulatory Role in Neuroinflammation. Mol. Neurobiol. 2013, 47, 1034–1044. [Google Scholar] [CrossRef]
- Nagele, R.G.; Wegiel, J.; Venkataraman, V.; Imaki, H.; Wang, K.; Wegiel, J. Contribution of glial cells to the development of amyloid plaques in Alzheimer’s disease. Neurobiol. Aging 2004, 25, 663–674. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Olabarria, M.; Noristani, H.N.; Yeh, C.; Rodriguez, J.J. Astrocytes in Alzheimer’s Disease. Neurother. J. Am. Soc. Exp. Neurother. 2010, 7, 399–412. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Mi, S.; Li, L. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Kim, J.-H. A Comprehensive Review on Factors Influences Biogenesis, Functions, Therapeutic and Clinical Implications of Exosomes. Int. J. Nanomed. 2021, 16, 1281–1312. [Google Scholar] [CrossRef]
- Simons, M.; Raposo, G. Exosomes—Vesicular carriers for intercellular communication. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef]
- Rashed, M.H.; Bayraktar, E.; Helal, G.K.; Abd-ellah, M.F. Exosomes: From Garbage Bins to Promising Therapeutic Targets. Int. J. Mol. Sci. 2017, 18, 538. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Desai, N.N.; Zahid, M.; Rubert, D.; Librelotto, N.; Luisa, M.; Bishayee, A.; Nabavi, S.M. Exosome biogenesis, bioactivities and functions as new delivery systems of natural compounds. Biotechnol. Adv. 2017, 36, 328–334. [Google Scholar] [CrossRef]
- Soria, F.N.; Pampliega, O.; Bourdenx, M.; Meissner, W.G.; Hill, A.F. Exosomes, an Unmasked Culprit in Neurodegenerative Diseases. Front. Neurosci. 2017, 11, 26. [Google Scholar] [CrossRef]
- Tamkovich, S.N.; Tutanov, O.S.; Laktionov, P.P. Exosomes: Generation, Structure, Transport, Biological Activity, and Diagnostic Application. Biol. Membr. 2016, 10, 163–173. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, Y.; Zhou, J.; Cao, K. Adipose-derived mesenchymal stem cell exosomes: A novel pathway for tissues repair. Cell Tissue Bank. 2019, 20, 153–161. [Google Scholar] [CrossRef]
- Wan, R.; Hussain, A.; Behfar, A.; Moran, S.L.; Zhao, C. The Therapeutic Potential of Exosomes in Soft Tissue Repair and Regeneration. Int. J. Mol. Sci. 2022, 23, 3869. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, M.; Tian, W. Physiological and pathological impact of exosomes of adipose tissue. Cell Prolif. 2016, 49, 3–13. [Google Scholar] [CrossRef]
- Desdin-Mico, G.; Mittelbrunn, M. Role of exosomes in the protection of cellular homeostasis. Cell Adh. Migr. 2017, 11, 127–134. [Google Scholar] [CrossRef]
- Olejarz, W.; Kubiak-Tomaszewska, G.; Chrzanowska, A.; Lorenc, T. Exosomes in Angiogenesis and Anti-angiogenic Therapy in Cancers. Int. J. Mol. Sci. 2020, 21, 5840. [Google Scholar] [CrossRef]
- Zhong, D.; Cao, Y.; Li, C.; Li, M.; Rong, Z.; Jiang, L.; Guo, Z.; Lu, H.; Hu, J. Highlight article Neural stem cell-derived exosomes facilitate spinal cord functional recovery after injury by promoting angiogenesis. Exp. Biol. Med. 2020, 245, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Aslan, C.; Maralbashi, S.; Salari, F.; Kahroba, H.; Sigaroodi, F.; Kazemi, T.; Kharaziha, P. Tumor—Derived exosomes: Implication in angiogenesis and antiangiogenesis cancer therapy. Cell. Physiol. 2019, 234, 16885–16903. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, S.; Israel, S.; Nagy, C.; Turecki, G. The emerging role of exosomes in mental disorders. Transl. Psychiatry 2019, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front. Cell. Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef]
- Jafari, A.; Babajani, A.; Abdollahpour, M.; Nayebali, A. Exosomes and cancer: From molecular mechanisms to clinical applications. Med. Oncol. 2021, 38, 45. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, D. Exosomes in cancer development, metastasis, and immunity. Biochim. Biophys. Acta Rev. Cancer 2020, 2, 455–468. [Google Scholar] [CrossRef]
- Jan, A.T.; Malik, M.A.; Rahman, S.; Yeo, H.R.; Lee, E.J.; Abdullah, T.S.; Choi, I. Perspective Insights of Exosomes in Neurodegenerative Diseases: A Critical Appraisal. Front. Aging Neurosci. 2017, 9, 317. [Google Scholar] [CrossRef]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome Theranostics: Biology and Translational Medicine. Theranostics 2018, 8, 237–255. [Google Scholar] [CrossRef]
- Anca, M.D.; Fenoglio, C.; Serpente, M.; Arosio, B.; Cesari, M.; Scarpini, E.A.; Galimberti, D. Exosome Determinants of Physiological Aging and Age-Related Neurodegenerative Diseases. Front. Aging Neurosci. 2019, 11, 232. [Google Scholar] [CrossRef]
- Liu, W.; Bai, X.; Zhang, A.; Huang, J.; Xu, S. Role of Exosomes in Central Nervous System Diseases. Front. Mol. Neurosci. 2019, 12, 240. [Google Scholar] [CrossRef]
- Pinnell, J.R.; Cui, M.; Tieu, K. Exosomes in Parkinson Disease. J. Neurochem. 2021, 157, 413–428. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Li, D.; Wu, W.; Xie, Z.; Liu, Q. Physiological and pathological insights into exosomes in the brain. Zool. Res. 2020, 41, 365–372. [Google Scholar] [CrossRef]
- Pascual, M.; Ibáñez, F.; Guerri, C. Exosomes as mediators of neuron-glia communication in neuroinflammation. Neural Regen. Res. 2018, 15, 796–801. [Google Scholar] [CrossRef]
- Li, K.; Huang, H.; Ren, H.; Yang, X. Role of exosomes in the pathogenesis of inflammation in Parkinson’s disease. Neural Regen. Res. 2022, 17, 1898–1906. [Google Scholar] [CrossRef]
- Zagrean, A.; Hermann, D.M.; Opris, I.; Zagrean, L.; Popa-wagner, A. Multicellular Crosstalk Between Exosomes and the Neurovascular Unit After Cerebral Ischemia. Therapeutic Implications. Front. Neurosci. 2018, 12, 811. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.; Jeyaraj, M.; Qasim, M.; Kim, J. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, W.; Zhang, H.; Zhang, F.; Chen, L.; Ma, L.; Leon, M.; Chen, S.; Liu, N.; Zhao, Q.; et al. Progress, opportunity, and perspective on exosome isolation—Efforts for efficient exosome-based theranostics. Theranostics 2020, 10, 3684–3707. [Google Scholar] [CrossRef]
- Jia, L.; Qiu, Q.; Zhang, H.; Chu, L.; Du, Y.; Zhang, J.; Zhou, C.; Liang, F.; Shi, S.; Wang, S.; et al. Concordance between the assessment of Aβ42, T-tau, and P-T181-tau in peripheral blood neuronal-derived exosomes and cerebrospinal fluid. Alzheimer’s Dement. 2019, 15, 1071–1080. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, Y.; Gao, Q.; Ping, D.; Wang, Y.; Wu, W.; Lin, X.; Fang, Y.; Zhang, J.; Shao, A. The Role of Exosomal microRNAs and Oxidative Stress in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 3232869. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Hornung, S.; Kruayatidee, A.; Maina, K.N.; del Rosario, I.; Paul, K.C.; Wong, D.Y.; Duarte Folle, A.; Markovic, D.; Palma, J.A.; et al. α-Synuclein in blood exosomes immunoprecipitated using neuronal and oligodendroglial markers distinguishes Parkinson’s disease from multiple system atrophy. Acta Neuropathol. 2021, 142, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Wu, Y.; Liu, G.; Jiang, Y.; Wang, X.; Wang, Z.; Zhang, J.; Feng, T. α-Synuclein in salivary extracellular vesicles as a potential biomarker of Parkinson’s disease. Neurosci. Lett. 2019, 696, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Rani, K.; Mukherjee, R.; Singh, E.; Kumar, S.; Sharma, V.; Vishwakarma, P.; Bharti, P.S.; Nikolajeff, F.; Dinda, A.K.; Goyal, V.; et al. Neuronal exosomes in saliva of Parkinson’s disease patients: A pilot study. Park. Relat. Disord. 2019, 67, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Quezada, J.; Ayala-Mar, S.; González-Valdez, J. The role of lipids in exosome biology and intercellular communication: Function, analytics and applications. Traffic 2021, 22, 204–220. [Google Scholar] [CrossRef]
- Ma, H.; Murphy, C.; Loscher, C.E.; Kennedy, R.O. Autoantibodies—Enemies, and/or potential allies? Front. Immunol. 2022, 13, 953726. [Google Scholar] [CrossRef]
- Lacroix-Desmazes, S.; Kaveri, S.V.; Mouthon, L.; Ayouba, A.; Malanchere, E.; Coutinho, A.; Kazatchkine, M.D. Self-reactive antibodies Ž natural autoantibodies. in healthy individuals. J. Immunol. Methods 1998, 216, 117–137. [Google Scholar] [CrossRef]
- Yadav, S.; Kashaninejad, N.; Masud, M.K.; Yamauchi, Y. Autoantibodies as diagnostic and prognostic cancer biomarker: Detection techniques and approaches. Biosens. Bioelectron. 2019, 139, 111315. [Google Scholar] [CrossRef]
- Volkov, M.; Coppola, M.; Huizinga, R.; Eftimov, F. Comprehensive overview of autoantibody isotype and subclass distribution. J. Allerg. Clin. Immunol. 2022, 150, 999–1010. [Google Scholar] [CrossRef]
- Ludwig, R.J.; Vanhoorelbeke, K.; Leypoldt, F.; Kaya, Z.; Bieber, K.; McLachlan, S.M.; Komorowski, L.; Luo, J.; Cabral-Marques, O.; Hammers, C.M.; et al. Mechanisms of Autoantibody- induced Pathology. Front. Immunol. 2017, 8, 603. [Google Scholar] [CrossRef]
- Meffre, E.; Connor, K.C.O. Impaired B cell tolerance checkpoints promote the development of autoimmune diseases and pathogenic autoantibodies. Immunol. Rev. 2022, 292, 90–101. [Google Scholar] [CrossRef]
- Ding, T.; Su, R.; Wu, R.; Xue, H.; Wang, Y.; Su, R.; Gao, C.; Li, X.; Wang, C. Frontiers of Autoantibodies in Autoimmune Disorders: Crosstalk Between Tfh/Tfr and Regulatory B Cells. Front. Immunol. 2021, 12, 641013. [Google Scholar] [CrossRef]
- Gibson, L.L.; Mckeever, A.; Cullen, A.E.; Nicholson, T.R.; Aarsland, D.; Zandi, M.S.; Pollak, T.A. Neuronal surface autoantibodies in dementia: A systematic review and meta—Analysis. J. Neurol. 2021, 268, 2769–2779. [Google Scholar] [CrossRef]
- Wotton, C.J.; Goldacre, M.J. Associations between speci fi c autoimmune diseases and subsequent dementia: Retrospective record-linkage cohort study, UK. BMJ 2017, 71, 576–583. [Google Scholar] [CrossRef]
- Witolear, A.; Jansen, I.E.; Wang, Y.; Desikan, R.S.; Gibbs, R.J.; Blauwendraat, C.; Schork, A.J.; Betella, F.; Ellinghaus, D.; Franke, A. Genome-wide Pleiotropy Between Parkinson Disease and Autoimmune Diseases. JAMA Neurol. 2017, 74, 780–792. [Google Scholar] [CrossRef]
- Sim, K.; Im, K.C.; Park, S. The Functional Roles and Applications of Immunoglobulins in Neurodegenerative Disease. Int. J. Cancer 2020, 21, 5295. [Google Scholar] [CrossRef]
- De Virgilio, A.; Greco, A.; Fabbrini, G.; Inghilleri, M.; Rizzo, M.I.; Gallo, A.; Conte, M.; Rosato, C.; Ciniglio Appiani, M.; de Vicentiis, M. Parkinson´s disease: Autoimmunity and neuroinflammation. Autoimmun. Rev. 2016, 15, 1005–1011. [Google Scholar] [CrossRef]
- Prüss, H. Autoantibodies in neurological disease. Nature 2021, 21, 798–813. [Google Scholar] [CrossRef]
- Marquetand, J.; Van Lessen, M.; Bender, B.; Reimold, M.; Elsen, G.; Stoecker, W.; Synofzik, M.; Medizinische, E. Slowly progressive LGI1 encephalitis with isolated late-onset cognitive dysfunction: A treatable mimic of Alzheimer’s disease. Eur. J. Neurol. 2015, 23, 28–29. [Google Scholar] [CrossRef]
- Mckeon, A.; Marnane, M.; Stack, J.P.; Kelly, P.J.; Lynch, T. Potassium Channel Antibody–Associated Encephalopathy Presenting With a Frontotemporal Dementia–like Syndrome. Arch. Neurol. 2007, 64, 1528–1530. [Google Scholar] [CrossRef]
- Reintjes, W.; Romijn, M.D.M.; Hollander, D.; Bruggen, J.P.; Marum, R.J. Van Reversible Dementia: Two Nursing Home Patients With Voltage-Gated Potassium Channel Antibody-Associated Limbic Encephalitis. J. Am. Med. Dir. Assoc. 2015, 16, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Tomczak, A.; Su, E.; Tugizova, M.; Carlson, A.M.; Kipp, L.B.; Feng, H.; Han, M.H. A case of GFAP-astroglial autoimmunity presenting with reversible parkinsonism. Mult. Scler. Relat. Disord. 2020, 39, 101900. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, H.; Meyer, T.; Prüss, H. GABA B receptor encephalitis in a patient diagnosed with amyotrophic lateral sclerosis. BMC Neurol. 2019, 19, 41. [Google Scholar] [CrossRef] [PubMed]
- Sabater, L.; Gaig, C.; Gelpi, E.; Bataller, L.; Lewerenz, J.; Torres-Vega, E.; Contreras, A.; Giometto, B.; Compta, Y.; Embid, C.; et al. A novel NREM and REM parasomnia with sleep breathing disorder associated with antibodies against IgLON5: A case series, pathological features, and characterization of the antigen. Lancet Neurol. 2014, 13, 575–586. [Google Scholar] [CrossRef]
- Holmdahl, R.; Nimmerjahn, F.; Ludwig, R.J. Editorial: Autoantibodies. Front. Immunol. 2019, 10, 484. [Google Scholar] [CrossRef]
- Sieghart, D.; Platzer, A.; Studenic, P.; Alasti, F.; Grundhuber, M.; Swiniarski, S.; Horn, T.; Haslacher, H.; Blüml, S.; Smolen, J.; et al. Determination of autoantibody isotypes increases the sensitivity of serodiagnostics in rheumatoid arthritis. Front. Immunol. 2018, 9, 876. [Google Scholar] [CrossRef]
- Giurdanella, F.; Nijenhuis, A.M.; Diercks, G.F.H.; Jonkman, M.F.; Pas, H.H. Keratinocyte binding assay identifies anti-desmosomal pemphigus antibodies where other tests are negative. Front. Immunol. 2018, 9, 839. [Google Scholar] [CrossRef]
- Scharf, M.; Miske, R.; Kade, S.; Hahn, S.; Denno, Y.; Begemann, N.; Rochow, N.; Radzimski, C.; Brakopp, S.; Probst, C.; et al. A spectrum of neural autoantigens, newly identified by histo-immunoprecipitation, mass spectrometry, and recombinant cell-based indirect immunofluorescence. Front. Immunol. 2018, 9, 1447. [Google Scholar] [CrossRef]
- Wen, L.; Krauss-Etschmann, S.; Petersen, F.; Yu, X. Autoantibodies in chronic obstructive pulmonary disease. Front. Immunol. 2018, 9, 66. [Google Scholar] [CrossRef]

| DIAGNOSTIC TOOLS IN NEURODEGENERATIVE DISORDERS | ||||
|---|---|---|---|---|
| MOLECULAR BIOMARKERS | IMAGING TECHNIQUES | |||
| Biomarkers in peripheric fluids | Alpha-synuclein in Parkinson’s Disease (PD) | Magnetic Resonance Imaging (MRI) | Structural MRI | |
| Amyloid-β and tau in Alzheimer´s Disease (AD) | ||||
| Functional MRI | ||||
| Inflammation markers | ||||
| Positron Emission Tomography (PET) | PD | DAT imaging | ||
| Exosomes | Neurotransmitters | |||
| Brain-gut denervation | ||||
| Alpha-synuclein | ||||
| Autoantibodies | AD | [18F] FDG–PET | ||
| Amyloid-beta | ||||
| Tau | ||||
| Molecular Biomarkers | Neuroimaging Techniques |
|---|---|
| Strengths | |
|
|
| Weaknesses | |
|
|
| Opportunities | |
|
|
| Challenges | |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Fernández, C.; Egiguren-Ortiz, J.; Razquin, J.; Gómez-Galán, M.; De las Heras-García, L.; Paredes-Rodríguez, E.; Astigarraga, E.; Miguélez, C.; Barreda-Gómez, G. Review of Technological Challenges in Personalised Medicine and Early Diagnosis of Neurodegenerative Disorders. Int. J. Mol. Sci. 2023, 24, 3321. https://doi.org/10.3390/ijms24043321
Domínguez-Fernández C, Egiguren-Ortiz J, Razquin J, Gómez-Galán M, De las Heras-García L, Paredes-Rodríguez E, Astigarraga E, Miguélez C, Barreda-Gómez G. Review of Technological Challenges in Personalised Medicine and Early Diagnosis of Neurodegenerative Disorders. International Journal of Molecular Sciences. 2023; 24(4):3321. https://doi.org/10.3390/ijms24043321
Chicago/Turabian StyleDomínguez-Fernández, Celtia, June Egiguren-Ortiz, Jone Razquin, Margarita Gómez-Galán, Laura De las Heras-García, Elena Paredes-Rodríguez, Egoitz Astigarraga, Cristina Miguélez, and Gabriel Barreda-Gómez. 2023. "Review of Technological Challenges in Personalised Medicine and Early Diagnosis of Neurodegenerative Disorders" International Journal of Molecular Sciences 24, no. 4: 3321. https://doi.org/10.3390/ijms24043321
APA StyleDomínguez-Fernández, C., Egiguren-Ortiz, J., Razquin, J., Gómez-Galán, M., De las Heras-García, L., Paredes-Rodríguez, E., Astigarraga, E., Miguélez, C., & Barreda-Gómez, G. (2023). Review of Technological Challenges in Personalised Medicine and Early Diagnosis of Neurodegenerative Disorders. International Journal of Molecular Sciences, 24(4), 3321. https://doi.org/10.3390/ijms24043321