
In Vitro and In Vivo Effects of IGF-1 Delivery Strategies on Tendon Healing: A Review


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
3. IGF-1 and Its Receptor IGF1R: Biological Characteristics and Mechanism of Action
4. PART A: Effects of IGF-1 Delivery In Vitro
4.1. IGF-1 Supplemented Culture Media
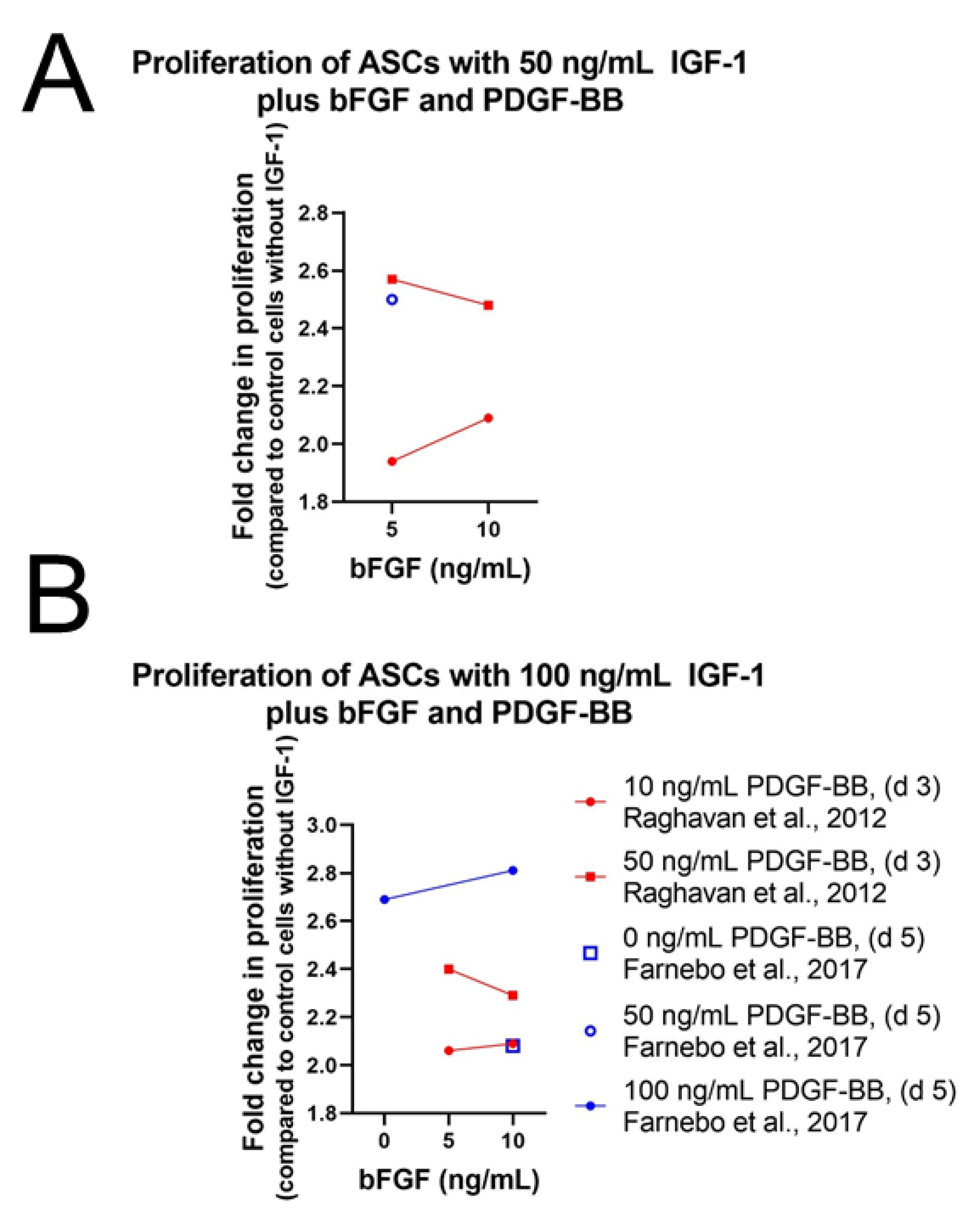
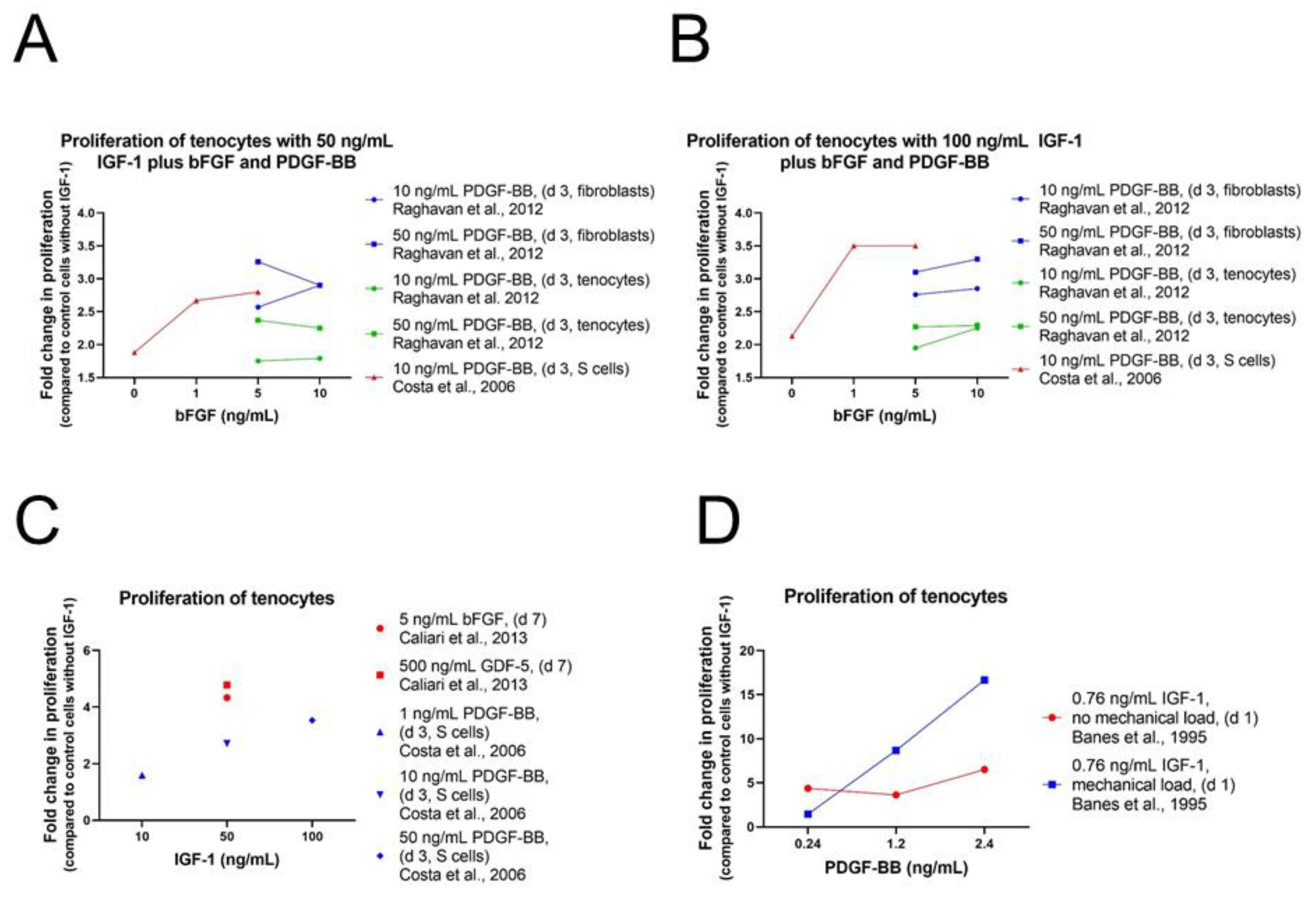
4.1.1. Proliferation and Migration
Stem Cells
Tenocytes and Fibroblasts
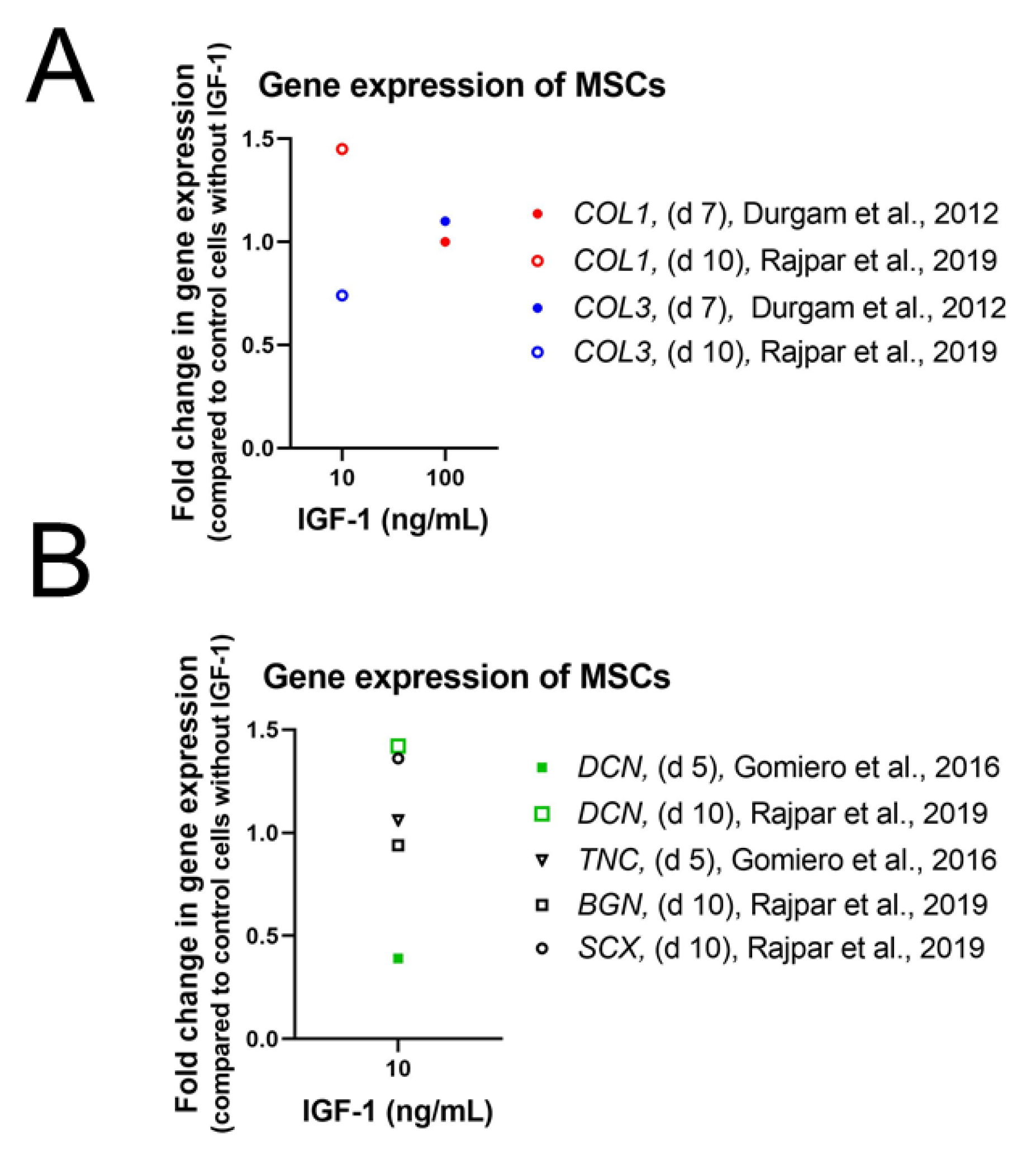
4.1.2. Differentiation and Tendon Markers
Stem Cells
Tenocytes and Fibroblasts
4.1.3. Morphology and ECM
Stem Cells
Tenocytes and Fibroblasts
4.1.4. Differential Effects of IGF-1 Applied as a Single Factor to Different Cell Types—Association to Signalling Pathways
4.2. IGF-1 in Combination with Other Growth Factors
4.2.1. Proliferation and Migration
Stem Cells
Tenocytes and Fibroblasts
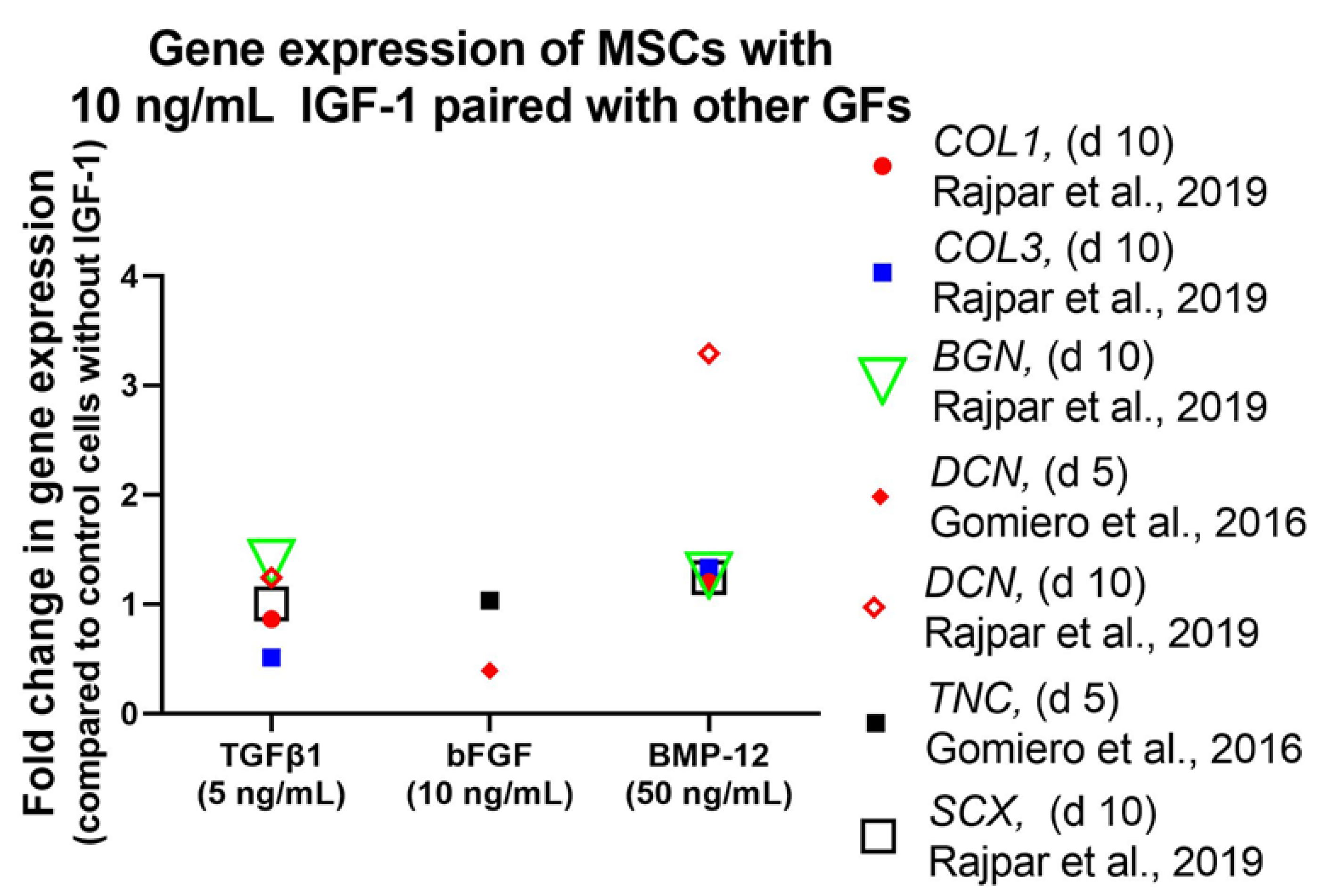
4.2.2. Differentiation and Tendon Markers
Stem Cells
Tenocytes and Fibroblasts
4.2.3. Morphology and ECM
Stem Cells
Tenocytes and Fibroblasts
4.3. Experiments Using Biological Supplementation Containing IGF-1
5. PART B: Effects of IGF-1 Delivery In Vivo
5.1. IGF-1 Injections
5.2. GH Injections
5.3. Fibrin Sealing Gel
5.4. Platelet Rich Plasma
5.5. Matrix Incorporation of IGF-1
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | ascorbic acid |
| AbAm | antibiotic antimycotic |
| ACAN | aggrecan |
| ALP | alkaline phosphatase |
| Akt = PKB | protein kinase B; collective name of a set of three serine/threonine-specific protein kinases |
| ASCs | adipose-derived stem cells, adipose tissue-derived stromal cells |
| α-MEM | α-MEM medium |
| BAD | cell death protein |
| BAPN | beta-aminopropionitrile |
| BCL-2 | antagonist of cell death BAD |
| bFGF | basic fibroblast growth factor |
| BGN | biglycan |
| BMP | bone morphogenetic protein |
| BMP-12 = GDF-7 | growth and differentiation factor-7 |
| BMP-13 = GDF-6 | growth and differentiation factor-6 |
| BMP-14 = GDF-5 | growth and differentiation factor-5 |
| BMSC | bone marrow-derived stromal cells |
| BSA | bovine serum albumin |
| CG | collagen-glycosaminoglycan |
| CM | Conditioned media |
| col I | collagen I protein |
| col III | collagen III protein |
| COL1A1 | collagen 1A1 gene |
| COL3A1 | collagen 3A1 gene |
| COMP | cartilage oligomeric matrix protein |
| CSA | cross-sectional area |
| CTGF | connective tissue growth factor |
| dACM | dehydrated amnion/chorion membrane |
| DCN | Decorin |
| dePRF | denatured PRF = denatured platelet rich fibrin |
| DMEM | Dubelco’s modified Eagle Medium |
| DNA | desoxy ribonucleic acid |
| ECM | Extracellular matrix |
| EGF | epidermal growth factor |
| EGR 1 | early growth response protein 1 |
| FBS | fetal bovine serum |
| FCS | fetal calf serum |
| FGF | fibroblast growth factors |
| FOXO | forkhead box transcription factor |
| FSR | fractional synthesis rates |
| GAG | glycosaminoglycan |
| GDF | growth and differentiation factor |
| GH | growth hormone |
| Grb2 | growth factor receptor-bound protein 2, an adapter protein |
| GTPase Ras | small GTPase (hydrolase enzymes for guanosine triphosphate) |
| Ham’s F12 | Ham’s F12 cell culture medium |
| H&E | Hematoxylin&Eosin |
| HSAM | hypothermically stored amniotic membrane |
| HSR | heavy slow resistance |
| IGF | insulin-like growth factor |
| IGFBP | insulin-like growth factor binding protein |
| IGF-1Ea | insulin-like growth factor isoform Ea |
| IGF-1Eb | insulin-like growth factor isoform Eb |
| IGF-1Ec | insulin-like growth factor isoform Ec |
| IGF1R | insulin-like growth factor-1 receptor |
| IP3 | inositol trisphosphate |
| IRS= | insulin receptor substrates |
| LLLT | low-level laser therapy |
| LR3-IGF-1 | variant form of IGF-1 which binds with low affinity to circulating binding proteins |
| m | mimic |
| M-199 | name of medium Mki67, proliferation Marker Ki67 |
| MAPK | mitogen-activated protein kinase |
| MGF | mechano growth factor = insulin-like growth factor isoform Ec |
| Mki67 | proliferation Marker Ki67 |
| MKX | Mohawk |
| mRNA | messenger ribonucleic acid |
| MSC | mesenchymal stem cells |
| mTOR | mammalian target of Rapamycin |
| mTORC2 | mammalian target of Rapamycin Complex 2 |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide… |
| OD | optical density |
| ON | Osteonectin |
| p85 | regulatory subunit of PI3K |
| PDGF-BB | platelet-derived growth factor-BB |
| PDK1 | Phosphoinositide-dependent Kinase-1 |
| PEG-IGF-1m | pegylated IGF-1 mimic |
| PI3K | phosphatidylinositol-3 kinase |
| PINP | procollagen type I N-terminal propeptide |
| PIP2 | phosphatidylinositol diphosphate |
| PIP3 | phosphatidylinositol triphosphate |
| PLC | phospholipase C |
| PRF | platelet rich fibrin |
| PRP | platelet rich plasma |
| Raf | best characterized Ras effector |
| Ras | small GTPase |
| rhGH | recombinant human growth factors |
| rhIGF-1 | recombinant human IGF-1 |
| RTKs | receptor tyrosine kinases |
| RUNX | runt-related transcription factor |
| SCX | Scleraxis |
| SDF-1 | stromal cell-derived factor 1 |
| Sos | guanine nucleotide |
| SOX9 | SRY-box transcription factor 9 |
| SVF | stromal vascular fraction |
| TGFβ | transforming growth factor β |
| TGFβ1 | transforming growth factor β1 |
| TGFβ2 | transforming growth factor β2 |
| TGFβ3 | transforming growth factor β3 |
| TIF | tendon internal fibroblasts |
| TNC | Tenascin-C |
| TNMD | Tenomodulin |
| TSC | tendon-derived stem cells |
| VEGF | vascular endothelial growth factor |
| VISA-P | Victorian Institute of Sport Assessment-Patella |
References
- Chartier, C.; ElHawary, H.; Baradaran, A.; Vorstenbosch, J.; Xu, L.; Efanov, J.I. Tendon: Principles of Healing and Repair. Semin. Plast. Surg. 2021, 35, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.L.; Nödl, M.T.; Galloway, J.L. Bringing tendon biology to heel: Leveraging mechanisms of tendon development, healing, and regeneration to advance therapeutic strategies. Dev. Dyn. 2021, 250, 393–413. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, E.; Tuna, Z.; Oksuz, C. Getting Better Results in Flexor Tendon Surgery and Therapy. Plast. Reconstr. Surg. Glob. Open 2021, 9, e3432. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar]
- Smith, A.H.; Capin, J.J.; Zarzycki, R.; Snyder-Mackler, L. Athletes With Bone-Patellar Tendon-Bone Autograft for Anterior Cruciate Ligament Reconstruction Were Slower to Meet Rehabilitation Milestones and Return-to-Sport Criteria Than Athletes With Hamstring Tendon Autograft or Soft Tissue Allograft: Secondary Analysis From the ACL-SPORTS Trial. J. Orthop. Sports Phys. Ther. 2020, 50, 259–266. [Google Scholar]
- Eliasson, P.; Agergaard, A.S.; Couppé, C.; Svensson, R.; Hoeffner, R.; Warming, S.; Warming, N.; Holm, C.; Jensen, M.H.; Krogsgaard, M.; et al. The Ruptured Achilles Tendon Elongates for 6 Months After Surgical Repair Regardless of Early or Late Weightbearing in Combination With Ankle Mobilization: A Randomized Clinical Trial. Am. J. Sport. Med. 2018, 46, 2492–2502. [Google Scholar] [CrossRef]
- Gulotta, L.V.; Rodeo, S.A. Growth Factors for Rotator Cuff Repair. Clin. Sport. Med. 2009, 28, 13–23. [Google Scholar] [CrossRef]
- Bolt, P.; Clerk, A.N.; Luu, H.H.; Kang, Q.; Kummer, J.L.; Deng, Z.L.; Olson, K.; Primus, F.; Montag, A.G.; He, T.C.; et al. BMP-14 gene therapy increases tendon tensile strength in a rat model of Achilles tendon injury. J. Bone Joint Surg. Am. 2007, 89, 1315–1320. [Google Scholar] [CrossRef]
- Morita, W.; Snelling, S.J.; Dakin, S.G.; Carr, A.J. Profibrotic mediators in tendon disease: A systematic review. Arthritis Res. Ther. 2016, 18, 269. [Google Scholar] [CrossRef]
- Evrova, O.; Buschmann, J. In vitro and in vivo effects of PDGF-BB delivery strategies on tendon healing: A review. Eur. Cells Mater. 2017, 34, 15–39. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed]
- Disser, N.P.; Sugg, K.B.; Talarek, J.R.; Sarver, D.C.; Rourke, B.J.; Mendias, C.L. Insulin-like growth factor 1 signaling in tenocytes is required for adult tendon growth. FASEB J. 2019, 33, 12680–12695. [Google Scholar] [CrossRef] [PubMed]
- Banes, A.J.; Tsuzaki, M.; Hu, P.Q.; Brigman, B.; Brown, T.; Almekinders, L.; Lawrence, W.T.; Fischer, T. PDGF-BB, IGF-I and mechanical load stimulate DNA synthesis in avian tendon fibroblasts in vitro. J. Biomech. 1995, 28, 1505–1513. [Google Scholar] [CrossRef]
- Kurtz, C.A.; Loebig, T.G.; Anderson, D.D.; DeMeo, P.J.; Campbell, P.G. Insulin-like growth factor I accelerates functional recovery from Achilles tendon injury in a rat model. Am. J. Sport. Med. 1999, 27, 363–369. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Chen, X.; Li, G.; Chan, K.M.; Heng, B.C.; Yin, Z.; Ouyang, H.W. Concise Review: Stem Cell Fate Guided By Bioactive Molecules for Tendon Regeneration. Stem Cells Transl. Med. 2018, 7, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Ehirchiou, D.; Kilts, T.M.; Inkson, C.A.; Embree, M.C.; Sonoyama, W.; Li, L.; Leet, A.I.; Seo, B.-M.; Zhang, L.; et al. Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche. Nat. Med. 2007, 13, 1219–1227. [Google Scholar] [CrossRef]
- Goncalves, A.I.; Rodrigues, M.T.; Lee, S.-J.; Atala, A.; Yoo, J.J.; Reis, R.L.; Gomes, M.E. Understanding the Role of Growth Factors in Modulating Stem Cell Tenogenesis. PLoS ONE 2013, 8, e83734. [Google Scholar] [CrossRef]
- Pfäffle, R.; Kiess, W. GH and IGF-1 Replacement in Children. Handb. Exp. Pharmacol. 2020, 261, 67–86. [Google Scholar]
- Nicholls, A.R.; Holt, R.I. Growth Hormone and Insulin-Like Growth Factor-1. Front. Horm. Res. 2016, 47, 101–114. [Google Scholar]
- Werner, H.; Weinstein, D.; Bentov, I. Similarities and differences between insulin and IGF-I: Structures, receptors, and signalling pathways. Arch. Physiol. Biochem. 2008, 114, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, F.; Barberi, L.; Dobrowolny, G.; Villa Nova Bacurau, A.; Nicoletti, C.; Rizzuto, E.; Rosenthal, N.; Scicchitano, B.M.; Musarò, A. Effects of IGF-1 isoforms on muscle growth and sarcopenia. Aging Cell 2019, 18, e12954. [Google Scholar] [CrossRef]
- Dai, Z.; Wu, F.; Yeung, E.W.; Li, Y. IGF-IEc expression, regulation and biological function in different tissues. Growth Horm. IGF Res. 2010, 20, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Tunç, B.S.; Toprak, F.; Toprak, S.F.; Sozer, S. In vitro investigation of growth factors including MGF and IGF-1 in neural stem cell activation, proliferation, and migration. Brain Res. 2021, 1759, 147366. [Google Scholar] [CrossRef]
- Luo, Q.; Wu, K.; Zhang, B.; Song, G. Mechano growth factor E peptide promotes rat bone marrow-derived mesenchymal stem cell migration through CXCR4-ERK1/2. Growth Factors 2015, 33, 210–219. [Google Scholar] [PubMed]
- Bach, L.A. IGF-binding proteins. J. Mol. Endocrinol. 2018, 61, T11–T28. [Google Scholar] [CrossRef]
- Ranke, M.B. Insulin-like growth factor binding-protein-3 (IGFBP-3). Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 701–711. [Google Scholar] [CrossRef]
- Olesen, J.L.; Heinemeier, K.M.; Langberg, H.; Magnusson, S.P.; Kjaer, M.; Flyvbjerg, A. Expression, content, and localization of insulin-like growth factor I in human achilles tendon. Connect. Tissue Res. 2006, 47, 200–206. [Google Scholar] [CrossRef]
- Dahlgren, L.A.; Nixon, A.J. Cloning and expression of equine insulin-like growth factor binding proteins in normal equine tendon. Am. J. Vet. Res. 2005, 66, 300–306. [Google Scholar] [CrossRef]
- Li, T.; Forbes, M.E.; Fuller, G.N.; Li, J.; Yang, X.; Zhang, W. IGFBP2: Integrative hub of developmental and oncogenic signaling network. Oncogene 2020, 39, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Bruick, R.K.; Yu, Y. Secreted IGFBP5 mediates mTORC1-dependent feedback inhibition of IGF-1 signalling. Nat. Cell Biol. 2016, 18, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Abbah, S.A.; Thomas, D.; Browne, S.; O’Brien, T.; Pandit, A.; Zeugolis, D.I. Co-transfection of decorin and interleukin-10 modulates pro-fibrotic extracellular matrix gene expression in human tenocyte culture. Sci. Rep. 2016, 6, 20922. [Google Scholar] [CrossRef] [PubMed]
- Trenker, R.; Jura, N. Receptor tyrosine kinase activation: From the ligand perspective. Curr. Opin. Cell Biol. 2020, 63, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.A.; Guo, S. Insulin receptor substrate signaling controls cardiac energy metabolism and heart failure. J. Endocrinol. 2017, 233, R131–R143. [Google Scholar] [CrossRef]
- Józefiak, A.; Larska, M.; Pomorska-Mól, M.; Ruszkowski, J.J. The IGF-1 Signaling Pathway in Viral Infections. Viruses 2021, 13, 1488. [Google Scholar] [CrossRef]
- Wang, T.; Thien, C.; Wang, C.; Ni, M.; Gao, J.; Wang, A.; Jiang, Q.; Tuan, R.S.; Zheng, Q.; Zheng, M.H. 3D uniaxial mechanical stimulation induces tenogenic differentiation of tendon-derived stem cells through a PI3K/AKT signaling pathway. FASEB J. 2018, 32, 4804–4814. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Pinto, M.C.; Kihara, A.H.; Goulart, V.A.; Tonelli, F.M.; Gomes, K.N.; Ulrich, H.; Resende, R.R. Calcium signaling and cell proliferation. Cell Signal. 2015, 27, 2139–2149. [Google Scholar] [CrossRef]
- Saran, U.; Humar, B.; Kolly, P.; Dufour, J.F. Hepatocellular carcinoma and lifestyles. J. Hepatol. 2016, 64, 203–214. [Google Scholar] [CrossRef]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, D.M.; Kang, J.H.; Choudhury, M.; Andrianifahanana, M.; Yin, X.; Limper, A.H.; Leof, E.B. IPF pathogenesis is dependent upon TGFβ induction of IGF-1. FASEB J. 2020, 34, 5363–5388. [Google Scholar] [CrossRef] [PubMed]
- Reif, K.; Buday, L.; Downward, J.; Cantrell, D.A. SH3 domains of the adapter molecule Grb2 complex with two proteins in T cells: The guanine nucleotide exchange protein Sos and a 75-kDa protein that is a substrate for T cell antigen receptor-activated tyrosine kinases. J. Biol. Chem. 1994, 269, 14081–14087. [Google Scholar] [CrossRef] [PubMed]
- Kulhanek, K.R.; Roose, J.P.; Rubio, I. Regulation of the Small GTPase Ras and Its Relevance to Human Disease. Methods Mol. Biol. 2021, 2262, 19–43. [Google Scholar] [PubMed]
- Terrell, E.M.; Morrison, D.K. Ras-Mediated Activation of the Raf Family Kinases. Cold Spring Harb. Perspect. Med. 2019, 9, a033746. [Google Scholar] [CrossRef]
- Lavoie, H.; Therrien, M. Regulation of RAF protein kinases in ERK signalling. Nat. Rev. Mol. Cell Biol. 2015, 16, 281–298. [Google Scholar] [CrossRef]
- Poulsen, R.C.; Carr, A.J.; Hulley, P.A. Cell proliferation is a key determinant of the outcome of FOXO3a activation. Biochem. Biophys. Res. Commun. 2015, 462, 78–84. [Google Scholar] [CrossRef]
- Roy, S.K.; Srivastava, R.K.; Shankar, S. Inhibition of PI3K/AKT and MAPK/ERK pathways causes activation of FOXO transcription factor, leading to cell cycle arrest and apoptosis in pancreatic cancer. J. Mol. Signal. 2010, 5, 10. [Google Scholar] [CrossRef]
- Sasako, T.; Ueki, K. Insulin/IGF-1 signaling and aging. Nihon Rinsho 2016, 74, 1435–1440. [Google Scholar]
- Villarruel, C.; Aguilar, P.S.; Ponce Dawson, S. High rates of calcium-free diffusion in the cytosol of living cells. Biophys. J. 2021, 120, 3960–3972. [Google Scholar] [CrossRef]
- Paupe, V.; Prudent, J. New insights into the role of mitochondrial calcium homeostasis in cell migration. Biochem. Biophys. Res. Commun. 2018, 500, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, D.R. Involvement of insulin-like growth factor-I in the control of glucose homeostasis. Curr. Opin. Pharmacol. 2006, 6, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Simpson, H.L.; Jackson, N.C.; Shojaee-Moradie, F.; Jones, R.H.; Russell-Jones, D.L.; Sönksen, P.H.; Dunger, D.B.; Umpleby, A.M. Insulin-like growth factor I has a direct effect on glucose and protein metabolism, but no effect on lipid metabolism in type 1 diabetes. J. Clin. Endocrinol. Metab. 2004, 89, 425–432. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Yakar, S. Mechanisms of disease: Metabolic effects of growth hormone and insulin-like growth factor 1. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Zenobi, P.D.; Graf, S.; Ursprung, H.; Froesch, E.R. Effects of insulin-like growth factor-I on glucose tolerance, insulin levels, and insulin secretion. J. Clin. Investig. 1992, 89, 1908–1913. [Google Scholar] [CrossRef] [PubMed]
- Bianda, T.L.; Hussain, M.A.; Keller, A.; Glatz, Y.; Schmitz, O.; Christiansen, J.S.; Alberti, K.G.; Froesch, E.R. Insulin-like growth factor-I in man enhances lipid mobilization and oxidation induced by a growth hormone pulse. Diabetologia 1996, 39, 961–969. [Google Scholar] [CrossRef]
- Aguirre, G.A.; De Ita, J.R.; de la Garza, R.G.; Castilla-Cortazar, I. Insulin-like growth factor-1 deficiency and metabolic syndrome. J. Transl. Med. 2016, 14, 3. [Google Scholar] [CrossRef]
- Kasprzak, A. Insulin-Like Growth Factor 1 (IGF-1) Signaling in Glucose Metabolism in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 6434. [Google Scholar] [CrossRef] [PubMed]
- Natoli, R.M.; Athanasiou, K.A. P188 reduces cell death and IGF-I reduces GAG release following single-impact loading of articular cartilage. J. Biomech. Eng. 2008, 130, 041012. [Google Scholar] [CrossRef]
- Hui, W.; Rowan, A.D.; Cawston, T. Insulin-like growth factor 1 blocks collagen release and down regulates matrix metalloproteinase-1, -3, -8, and -13 mRNA expression in bovine nasal cartilage stimulated with oncostatin M in combination with interleukin 1alpha. Ann. Rheum. Dis. 2001, 60, 254–261. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Chubinskaya, S.; Schoeberl, B.; Florine, E.; Kopesky, P.; Grodzinsky, A.J. Effects of insulin-like growth factor-1 and dexamethasone on cytokine-challenged cartilage: Relevance to post-traumatic osteoarthritis. Osteoarthr. Cartil. 2015, 23, 266–274. [Google Scholar] [CrossRef]
- Lo, M.Y.; Kim, H.T. Chondrocyte apoptosis induced by collagen degradation: Inhibition by caspase inhibitors and IGF-1. J. Orthop. Res. 2004, 22, 140–144. [Google Scholar] [CrossRef]
- Mueller, R.V.; Hunt, T.K.; Tokunaga, A.; Spencer, E.M. The effect of insulinlike growth factor I on wound healing variables and macrophages in rats. Arch. Surg. 1994, 129, 262–265. [Google Scholar] [CrossRef]
- Suh, D.Y.; Hunt, T.K.; Spencer, E.M. Insulin-like growth factor-I reverses the impairment of wound healing induced by corticosteroids in rats. Endocrinology 1992, 131, 2399–2403. [Google Scholar] [CrossRef]
- Abrahamsson, S.O.; Lundborg, G.; Lohmander, L.S. Recombinant human insulin-like growth factor-I stimulates in vitro matrix synthesis and cell proliferation in rabbit flexor tendon. J. Orthop. Res. 1991, 9, 495–502. [Google Scholar] [CrossRef]
- Murphy, D.J.; Nixon, A.J. Biochemical and site-specific effects of insulin-like growth factor I on intrinsic tenocyte activity in equine flexor tendons. Am. J. Vet. Res. 1997, 58, 103–109. [Google Scholar]
- Tsuzaki, M.; Brigman, B.E.; Yamamoto, J.; Lawrence, W.T.; Simmons, J.G.; Mohapatra, N.K.; Lund, P.K.; Van Wyk, J.; Hannafin, J.A.; Bhargava, M.M.; et al. Insulin-like growth factor-I is expressed by avian flexor tendon cells. J. Orthop. Res. 2000, 18, 546–556. [Google Scholar] [CrossRef]
- Hansson, H.A.; Dahlin, L.B.; Lundborg, G.; Löwenadler, B.; Paleus, S.; Skottner, A. Transiently increased insulin-like growth factor I immunoreactivity in tendons after vibration trauma. An immunohistochemical study on rats. Scand. J. Plast. Reconstr. Surg. Hand Surg. 1988, 22, 1–6. [Google Scholar]
- Abrahamsson, S.O.; Lohmander, S. Differential effects of insulin-like growth factor-I on matrix and DNA synthesis in various regions and types of rabbit tendons. J. Orthop. Res. 1996, 14, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Hansson, H.A.; Engström, A.M.; Holm, S.; Rosenqvist, A.L. Somatomedin C immunoreactivity in the Achilles tendon varies in a dynamic manner with the mechanical load. Acta Physiol. Scand. 1988, 134, 199–208. [Google Scholar] [CrossRef]
- Abrahamsson, S.O.; Lundborg, G.; Lohmander, L.S. Long-term explant culture of rabbit flexor tendon: Effects of recombinant human insulin-like growth factor-I and serum on matrix metabolism. J. Orthop. Res. 1991, 9, 503–515. [Google Scholar] [CrossRef]
- Caliari, S.R.; Harley, B.A.C. The effect of anisotropic collagen-GAG scaffolds and growth factor supplementation on tendon cell recruitment, alignment, and metabolic activity. Biomaterials 2011, 32, 5330–5340. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, X.; Lin, S.; Li, X.; Zhang, S.; Song, Y.H. Insulin-like growth factor 1 enhances the migratory capacity of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2007, 356, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Ponte, A.L.; Marais, E.; Gallay, N.; Langonné, A.; Delorme, B.; Hérault, O.; Charbord, P.; Domenech, J. The in vitro migration capacity of human bone marrow mesenchymal stem cells: Comparison of chemokine and growth factor chemotactic activities. Stem Cells 2007, 25, 1737–1745. [Google Scholar] [CrossRef]
- Ozaki, Y. Comprehensive Analysis of Chemotactic Factors for Bone Marrow Mesenchymal Stem Cells. Stem Cells Dev. 2007, 16, 119–130. [Google Scholar] [CrossRef]
- Durgam, S.S.; Stewart, A.A.; Pondenis, H.C.; Gutierrez-Nibeyro, S.M.; Evans, R.B.; Stewart, M.C. Comparison of equine tendon- and bone marrow-derived cells cultured on tendon matrix with or without insulin-like growth factor-I supplementation. Am. J. Vet. Res. 2012, 73, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, S.S.; Woon, C.Y.L.; Kraus, A.; Megerle, K.; Hung, P.; Chang, J. Optimization of Human Tendon Tissue Engineering: Synergistic Effects of Growth Factors for Use in Tendon Scaffold Repopulation. Plast. Reconstr. Surg. 2012, 129, 479–489. [Google Scholar] [CrossRef]
- Holladay, C.; Abbah, S.A.; O’Dowd, C.; Pandit, A.; Zeugolis, D.I. Preferential tendon stem cell response to growth factor supplementation. J. Tissue Eng. Regen. Med. 2016, 10, 783–798. [Google Scholar] [CrossRef] [PubMed]
- Farnebo, S.; Farnebo, L.; Kim, M.; Woon, C.; Pham, H.; Chang, J. Optimized Repopulation of Tendon Hydrogel: Synergistic Effects of Growth Factor Combinations and Adipose-Derived Stem Cells. Hand 2017, 12, 68–77. [Google Scholar] [CrossRef]
- Musson, D.S.; Tay, M.L.; Chhana, A.; Pool, B.; Coleman, B.; Naot, D.; Cornish, J. Lactoferrin and parathyroid hormone are not harmful to primary tenocytes in vitro, but PDGF may be. Muscles Ligaments Tendons J. 2017, 7, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.A.; Wu, C.; Pham, B.V.; Chong, A.K.S.; Pham, H.M.; Chang, J. Tissue engineering of flexor tendons: Optimization of tenocyte proliferation using growth factor supplementation. Tissue Eng. 2006, 12, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Caliari, S.R.; Harley, B.A.C. Composite Growth Factor Supplementation Strategies to Enhance Tenocyte Bioactivity in Aligned Collagen-GAG Scaffolds. Tissue Eng. Part A 2013, 19, 1100–1112. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, L.A.; MohamMed, H.O.; Nixon, A.J. Temporal expression of growth factors and matrix molecules in healing tendon lesions. J. Orthop. Res. 2005, 23, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wang, X.; Zhang, Y.; Carr, A.J.; Zhu, L.; Xia, Z.; Sabokbar, A. Development of a refined tenocyte expansion culture technique for tendon tissue engineering. J. Tissue Eng. Regen. Med. 2014, 8, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Kang, E.S. Ideal concentration of growth factors in rabbit’s flexor tendon culture. Yonsei Med. J. 1999, 40, 26–29. [Google Scholar] [CrossRef]
- Dahlgren, L.A.; Nixon, A.J.; Brower-Toland, B.D. Effects of beta-aminopropionitrile on equine tendon metabolism in vitro and on effects of insulin-like growth factor-I on matrix production by equine tenocytes. Am. J. Vet. Res. 2001, 62, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Gomiero, C.; Bertolutti, G.; Martinello, T.; Van Bruaene, N.; Broeckx, S.Y.; Patruno, M.; Spaas, J.H. Tenogenic induction of equine mesenchymal stem cells by means of growth factors and low-level laser technology. Vet. Res. Commun. 2016, 40, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Rajpar, I.; Barrett, J.G. Optimizing growth factor induction of tenogenesis in three-dimensional culture of mesenchymal stem cells. J. Tissue Eng. 2019, 10, 2041731419848776. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.R.; Buhrmann, C.; Mobasheri, A.; Matis, U.; Shakibaei, M. Three-dimensional high-density co-culture with primary tenocytes induces tenogenic differentiation in mesenchymal stem cells. J. Orthop. Res. 2011, 29, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Stanco, D.; Caprara, C.; Ciardelli, G.; Mariotta, L.; Gola, M.; Minonzio, G.; Soldati, G. Tenogenic differentiation protocol in xenogenic-free media enhances tendon-related marker expression in ASCs. PLoS ONE 2019, 14, e0212192. [Google Scholar] [CrossRef]
- Herchenhan, A.; Bayer, M.L.; Eliasson, P.; Magnusson, S.P.; Kjaer, M. Insulin-like growth factor I enhances collagen synthesis in engineered human tendon tissue. Growth Horm. IGF Res. 2015, 25, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Bestwick, C.S.; Bestwick, L.A.; Maffulli, N.; Aspden, R.M. Phenotypic drift in human tenocyte culture. Tissue Eng. 2006, 12, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Evrova, O.; Kellenberger, D.; Calcagni, M.; Vogel, V.; Buschmann, J. Supporting Cell-Based Tendon Therapy: Effect of PDGF-BB and Ascorbic Acid on Rabbit Achilles Tenocytes in Vitro. Int. J. Mol. Sci. 2020, 21, 458. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.P.; Chan, K.M.; Maffulli, N.; Webb, S.; Lee, K.K. Effect of basic fibroblast growth factor. An in vitro study of tendon healing. Clin. Orthop. Relat. Res. 1997, 239–247. [Google Scholar]
- Chen, M.H.; Huang, Y.C.; Sun, J.S.; Chao, Y.H.; Chen, M.H. Second messengers mediating the proliferation and collagen synthesis of tenocytes induced by low-level laser irradiation. Lasers Med. Sci. 2015, 30, 263–272. [Google Scholar] [CrossRef]
- Wolfman, N.M.; Hattersley, G.; Cox, K.; Celeste, A.J.; Nelson, R.; Yamaji, N.; Dube, J.L.; DiBlasio-Smith, E.; Nove, J.; Song, J.J.; et al. Ectopic induction of tendon and ligament in rats by growth and differentiation factors 5, 6, and 7, members of the TGF-beta gene family. J. Clin. Investig. 1997, 100, 321–330. [Google Scholar] [CrossRef]
- Chhabra, A.; Tsou, D.; Clark, R.T.; Gaschen, V.; Hunziker, E.B.; Mikic, B. GDF-5 deficiency in mice delays Achilles tendon healing. J. Orthop. Res. 2003, 21, 826–835. [Google Scholar] [CrossRef]
- Roberts, A.B.; Sporn, M.B. (Eds.) The Transforming Growth Factor-βs. In Peptide Growth Factors and Their Receptors I; Springer: New York, NY, USA, 1991; pp. 419–472. [Google Scholar]
- Shah, M.; Foreman, D.M.; Ferguson, M.W. Neutralisation of TGF-beta 1 and TGF-beta 2 or exogenous addition of TGF-beta 3 to cutaneous rat wounds reduces scarring. J. Cell Sci. 1995, 108(Pt. 3), 985–1002. [Google Scholar] [CrossRef]
- Qiu, Y.W.; Wang, X.; Zhang, Y.N.; Carr, A.J.; Zhu, L.W.; Xia, Z.D.; Sabokbar, A. In vitro two-dimensional and three-dimensional tenocyte culture for tendon tissue engineering. J. Tissue Eng. Regen. Med. 2016, 10, E216–E226. [Google Scholar] [CrossRef]
- Violini, S.; Ramelli, P.; Pisani, L.F.; Gorni, C.; Mariani, P. Horse bone marrow mesenchymal stem cells express embryo stem cell markers and show the ability for tenogenic differentiation by in vitro exposure to BMP-12. BMC Cell Biol. 2009, 10, 29. [Google Scholar] [CrossRef]
- Fu, S.C.; Wong, Y.P.; Chan, B.P.; Pau, H.M.; Cheuk, Y.C.; Lee, K.M.; Chan, K.M. The roles of bone morphogenetic protein (BMP) 12 in stimulating the proliferation and matrix production of human patellar tendon fibroblasts. Life Sci. 2003, 72, 2965–2974. [Google Scholar] [CrossRef] [PubMed]
- Orfei, C.P.; Vigano, M.; Pearson, J.R.; Colombini, A.; De Luca, P.; Ragni, E.; Santos-Ruiz, L.; de Girolamo, L. In Vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGF3, BMP-12 and Ascorbic Acid Stimulation. Int. J. Mol. Sci. 2019, 20, 149. [Google Scholar] [CrossRef] [PubMed]
- MooRes, J. Vitamin C: A wound healing perspective. Br. J. Community Nurs. 2013, 18, S8–S11. [Google Scholar]
- Lui, P.P.; Wong, O.T.; Lee, Y.W. Transplantation of tendon-derived stem cells pre-treated with connective tissue growth factor and ascorbic acid in vitro promoted better tendon repair in a patellar tendon window injury rat model. Cytotherapy 2016, 18, 99–112. [Google Scholar] [CrossRef]
- Lee, C.H.; Lee, F.Y.; Tarafder, S.; Kao, K.; Jun, Y.; Yang, G.; Mao, J.J. Harnessing endogenous stem/progenitor cells for tendon regeneration. J. Clin. Investig. 2015, 125, 2690–2701. [Google Scholar] [CrossRef]
- Lorbach, O.; Baums, M.H.; Kostuj, T.; Pauly, S.; Scheibel, M.; Carr, A.; Zargar, N.; Saccomanno, M.F.; Milano, G. Advances in biology and mechanics of rotator cuff repair. Knee Surg. Sport. Traumatol. Arthrosc. 2015, 23, 530–541. [Google Scholar] [CrossRef]
- Marx, R.E. Platelet-rich plasma: Evidence to support its use. J. Oral Maxillofac. Surg. 2004, 62, 489–496. [Google Scholar] [CrossRef]
- Pauly, S.; Klatte-Schulz, F.; Stahnke, K.; Scheibel, M.; Wildemann, B. The effect of autologous platelet rich plasma on tenocytes of the human rotator cuff. BMC Musculoskelet. Disord. 2018, 19, 422. [Google Scholar] [CrossRef]
- Riboh, J.C.; Saltzman, B.M.; Yanke, A.B.; Cole, B.J. Human Amniotic Membrane-Derived Products in Sports Medicine: Basic Science, Early Results, and Potential Clinical Applications. Am. J. Sport. Med. 2016, 44, 2425–2434. [Google Scholar] [CrossRef]
- McQuilling, J.P.; Kimmerling, K.A.; Staples, M.C.; Mowry, K.C. Evaluation of two distinct placental-derived membranes and their effect on tenocyte responses in vitro. J. Tissue Eng. Regen. Med. 2019, 13, 1316–1330. [Google Scholar] [CrossRef]
- Usuelli, F.G.; Grassi, M.; Maccario, C.; Vigano, M.; Lanfranchi, L.; Alfieri Montrasio, U.; de Girolamo, L. Intratendinous adipose-derived stromal vascular fraction (SVF) injection provides a safe, efficacious treatment for Achilles tendinopathy: Results of a randomized controlled clinical trial at a 6-month follow-up. Knee Surg. Sport. Traumatol. Arthrosc. 2018, 26, 2000–2010. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.H.; Chai, J.W.; Jeong, E.C.; Oh, S.; Kim, P.S.; Yoon, J.Y.; Yoon, K.S. Intratendinous Injection of Autologous Adipose Tissue-Derived Mesenchymal Stem Cells for the Treatment of Rotator Cuff Disease: A First-In-Human Trial. Stem Cells 2018, 36, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Barrachina, L.; Ranera, B.; Remacha, A.R.; Moreno, B.; de Blas, I.; Sanz, A.; Vázquez, F.J.; Vitoria, A.; Junquera, C.; et al. Comparison of autologous bone marrow and adipose tissue derived mesenchymal stem cells, and platelet rich plasma, for treating surgically induced lesions of the equine superficial digital flexor tendon. Vet. J. 2017, 224, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Nixon, A.J.; Dahlgren, L.A.; Haupt, J.L.; Yeager, A.E.; Ward, D.L. Effect of adipose-derived nucleated cell fractions on tendon repair in horses with collagenase-induced tendinitis. Am. J. Vet. Res. 2008, 69, 928–937. [Google Scholar] [CrossRef]
- Polly, S.S.; Nichols, A.E.C.; Donnini, E.; Inman, D.J.; Scott, T.J.; Apple, S.M.; Werre, S.R.; Dahlgren, L.A. Adipose-Derived Stromal Vascular Fraction and Cultured Stromal Cells as Trophic Mediators for Tendon Healing. J. Orthop. Res. 2019, 37, 1429–1439. [Google Scholar] [CrossRef]
- Savaris, M.; dos Santos, V.; Brandalise, R.N. Influence of different sterilization processes on the properties of commercial poly(lactic acid). Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 69, 661–667. [Google Scholar] [CrossRef]
- Mainil-Varlet, P.; Curtis, R.; Gogolewski, S. Effect of in vivo and in vitro degradation on molecular and mechanical properties of various low-molecular-weight polylactides. J. Biomed. Mater. Res. 1997, 36, 360–380. [Google Scholar] [CrossRef]
- Moioli, E.K.; Hong, L.I.U.; Guardado, J.; Clark, P.A.; Mao, J.J. Sustained Release of TGFβ3 from PLGA Microspheres and Its Effect on Early Osteogenic Differentiation of Human Mesenchymal Stem Cells. Tissue Eng. 2006, 12, 537–546. [Google Scholar] [CrossRef]
- Dahlgren, L.A.; van der Meulen, M.C.; Bertram, J.E.; Starrak, G.S.; Nixon, A.J. Insulin-like growth factor-I improves cellular and molecular aspects of healing in a collagenase-induced model of flexor tendinitis. J. Orthop. Res. 2002, 20, 910–919. [Google Scholar] [CrossRef]
- Lyras, D.N.; Kazakos, K.; Verettas, D.; Chronopoulos, E.; Folaranmi, S.; Agrogiannis, G. Effect of combined administration of Transforming Growth Factor-b1 and Insulin-like Growth Factor I on the mechanical properties of a patellar tendon defect model in rabbits. Acta Orthop. Belg. 2010, 76, 380–386. [Google Scholar]
- Andersson, T.; Eliasson, P.; Aspenberg, P. Growth hormone does not stimulate early healing in rat tendons. Int J. Sport. Med. 2012, 33, 240–243. [Google Scholar] [CrossRef]
- Hansen, M.; Boesen, A.; Holm, L.; Flyvbjerg, A.; Langberg, H.; Kjaer, M. Local administration of insulin-like growth factor-I (IGF-I) stimulates tendon collagen synthesis in humans. Scand. J. Med. Sci. Sport. 2013, 23, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Doessing, S.; Heinemeier, K.M.; Holm, L.; Mackey, A.L.; Schjerling, P.; Rennie, M.; Smith, K.; Reitelseder, S.; Kappelgaard, A.M.; Rasmussen, M.H.; et al. Growth hormone stimulates the collagen synthesis in human tendon and skeletal muscle without affecting myofibrillar protein synthesis. J. Physiol. 2010, 588(Pt. 2), 341–351. [Google Scholar] [CrossRef]
- Li, L.; Ni, R.; Shao, Y.; Mao, S. Carrageenan and its applications in drug delivery. Carbohydr. Polym. 2014, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.L.; Hansen, M.; Turtumoygard, I.F.; Hoffner, R.; Schjerling, P.; Christensen, J.; Mendias, C.L.; Magnusson, P.S.; Kjaer, M. No Treatment Benefits of Local Administration of Insulin-like Growth Factor-1 in Addition to Heavy Slow Resistance Training in Tendinopathic Human Patellar Tendons: A Randomized, Double-Blind, Placebo-Controlled Trial With 1-Year Follow-up. Am. J. Sport. Med. 2021, 49, 2361–2370. [Google Scholar] [CrossRef] [PubMed]
- Voleti, P.B.; Buckley, M.R.; Soslowsky, L.J. Tendon Healing: Repair and Regeneration. Annu. Rev. Biomed. Eng. 2012, 14, 47–71. [Google Scholar] [CrossRef]
- Notermans, T.; Hammerman, H.; Eliasson, P.; Isaksson, H. Tendon mechanobiology in small-animal experiments during post-transection healing. Eur. Cells Mater. 2021, 42, 375–391. [Google Scholar] [CrossRef]
- Kominsky, D.J.; Campbell, E.L.; Colgan, S.P. Metabolic shifts in immunity and inflammation. J. Immunol. 2010, 184, 4062–4068. [Google Scholar] [CrossRef]
- Vannella, K.M.; Wynn, T.A. Mechanisms of Organ Injury and Repair by Macrophages. Annu. Rev. Physiol. 2017, 79, 593–617. [Google Scholar] [CrossRef]
- Li, Y.; Fessel, G.; Georgiadis, M.; Snedeker, J.G. Advanced glycation end-products diminish tendon collagen fiber sliding. Matrix Biol. 2013, 32, 169–177. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of advanced glycation end products in cellular signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, M.; Aston, C.; Hintz, R.; Vlassara, H. Receptor-specific induction of insulin-like growth factor I in human monocytes by advanced glycosylation end product-modified proteins. J. Clin. Investig. 1992, 90, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J. Insulin-like growth factor-I regulation of immune function: A potential therapeutic target in autoimmune diseases? Pharmacol. Rev. 2010, 62, 199–236. [Google Scholar] [CrossRef] [PubMed]
- Lyras, D.N.; Kazakos, K.; Agrogiannis, G.; Verettas, D.; Kokka, A.; Kiziridis, G.; Chronopoulos, E.; Tryfonidis, M. Experimental study of tendon healing early phase: Is IGF-1 expression influenced by platelet rich plasma gel? Orthop. Traumatol. Surg. Res. 2010, 96, 381–387. [Google Scholar] [CrossRef]
- Wong, C.C.; Huang, Y.M.; Chen, C.H.; Lin, F.H.; Yeh, Y.Y.; Bai, M.Y. Cytokine and Growth Factor Delivery from Implanted Platelet-Rich Fibrin Enhances Rabbit Achilles Tendon Healing. Int. J. Mol. Sci. 2020, 21, 3221. [Google Scholar] [CrossRef] [PubMed]
- Khiste, S.V.; Naik Tari, R. Platelet-rich fibrin as a biofuel for tissue regeneration. Int. Sch. Res. Not. 2013, 2013, 627367. [Google Scholar] [CrossRef]
- Prabhath, A.; Vernekar, V.N.; Esdaille, C.J.; Eisenberg, E.; Lebaschi, A.; Badon, M.; Seyedsalehi, A.; Dzidotor, G.; Tang, X.; Dyment, N. Pegylated insulin-like growth factor-1 biotherapeutic delivery promotes rotator cuff regeneration in a rat model. J. Biomed. Mater. Res. Part A 2022, 110, 1356–1371. [Google Scholar] [CrossRef]
- Vaysman, M.; Alben, M.; Todd, M.; Ruotolo, C. Pharmacologic Enhancement of Rotator Cuff Repair: A Narrative Review. Orthop. Rev. 2022, 14, 37782. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miescher, I.; Rieber, J.; Calcagni, M.; Buschmann, J. In Vitro and In Vivo Effects of IGF-1 Delivery Strategies on Tendon Healing: A Review. Int. J. Mol. Sci. 2023, 24, 2370. https://doi.org/10.3390/ijms24032370
Miescher I, Rieber J, Calcagni M, Buschmann J. In Vitro and In Vivo Effects of IGF-1 Delivery Strategies on Tendon Healing: A Review. International Journal of Molecular Sciences. 2023; 24(3):2370. https://doi.org/10.3390/ijms24032370
Chicago/Turabian StyleMiescher, Iris, Julia Rieber, Maurizio Calcagni, and Johanna Buschmann. 2023. "In Vitro and In Vivo Effects of IGF-1 Delivery Strategies on Tendon Healing: A Review" International Journal of Molecular Sciences 24, no. 3: 2370. https://doi.org/10.3390/ijms24032370
APA StyleMiescher, I., Rieber, J., Calcagni, M., & Buschmann, J. (2023). In Vitro and In Vivo Effects of IGF-1 Delivery Strategies on Tendon Healing: A Review. International Journal of Molecular Sciences, 24(3), 2370. https://doi.org/10.3390/ijms24032370