
Cannabinoids and Their Receptors in Skin Diseases


Abstract
:1. Introduction
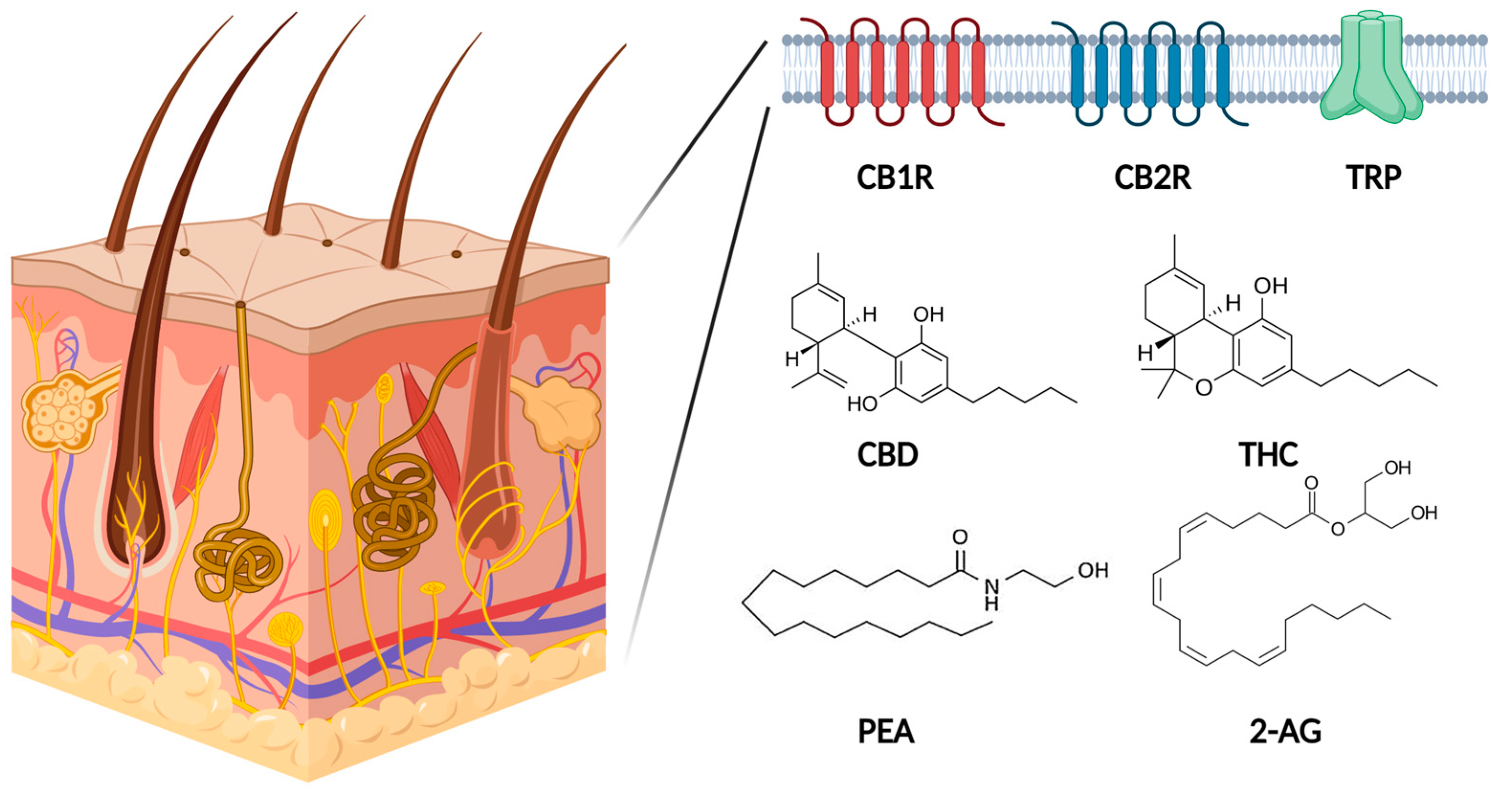
1.1. Types of Cannabinoids
1.2. Endocannabinoid System in Human Skin

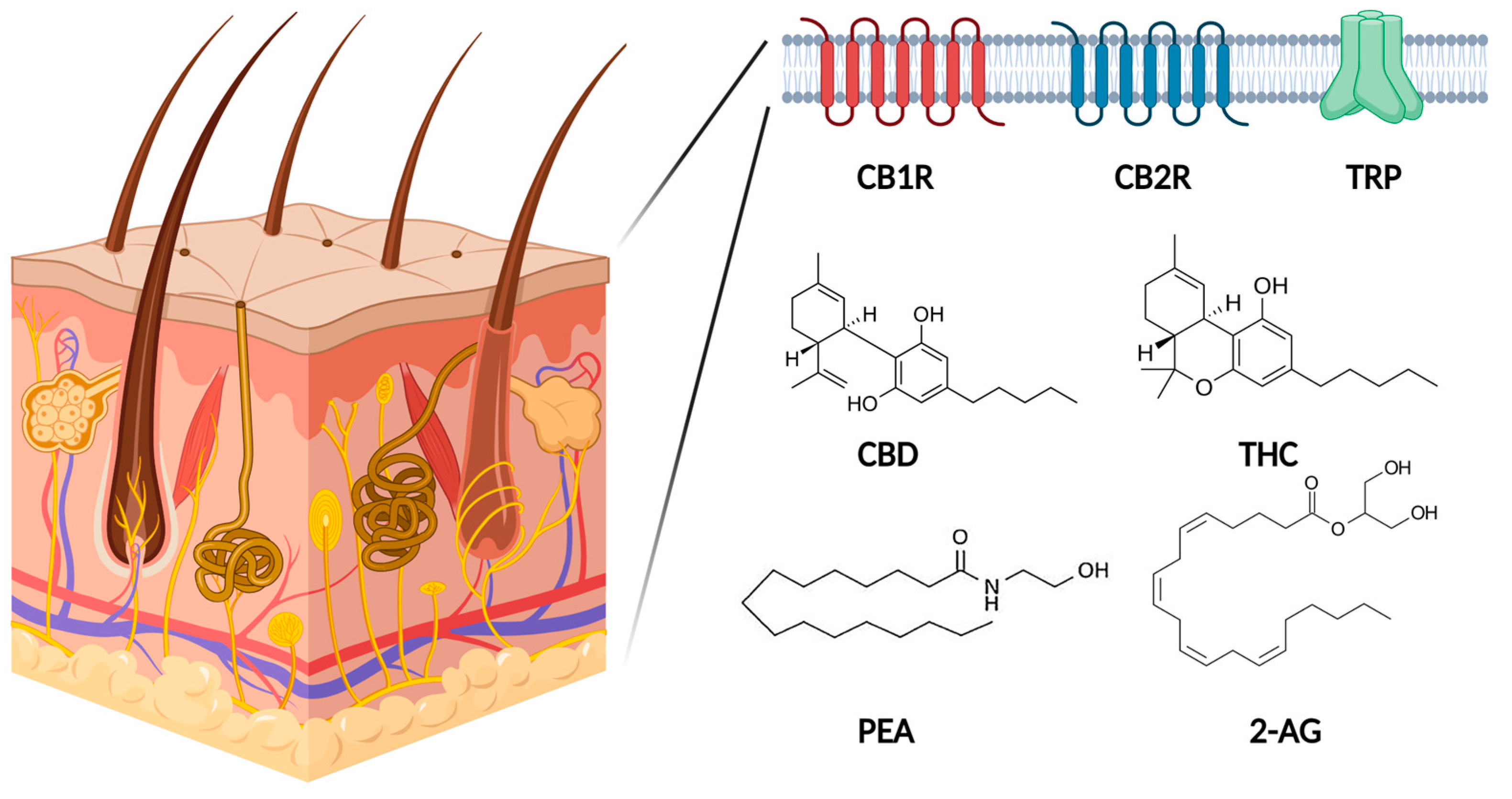{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Cannabinoids | Constituents |
|---|---|
| Endocannabinoids (ECB) | Palmitoylethanolamide (PEA) |
| Anandamide (AEA) or N-arachidonoylethanolamine | |
| 2-arachidonoyl-glycerol (2-AG) | |
| Phytocannabinoids | Δ(9)-Tetrahydrocannabinol (THC) |
| Cannabidiol (CBD) | |
| Cannabigerol (CBG) | |
| Cannabichromene (CBC) | |
| Cannabigerovarin (CBGV) | |
| Cannabinol (CBN) | |
| Cannabigerolic acid (CBGA) | |
| Synthetic cannabinoids | WIN55,212-2 |
| JTE-907 |
1.3. Other Receptors Activated by CANNABINOIDS
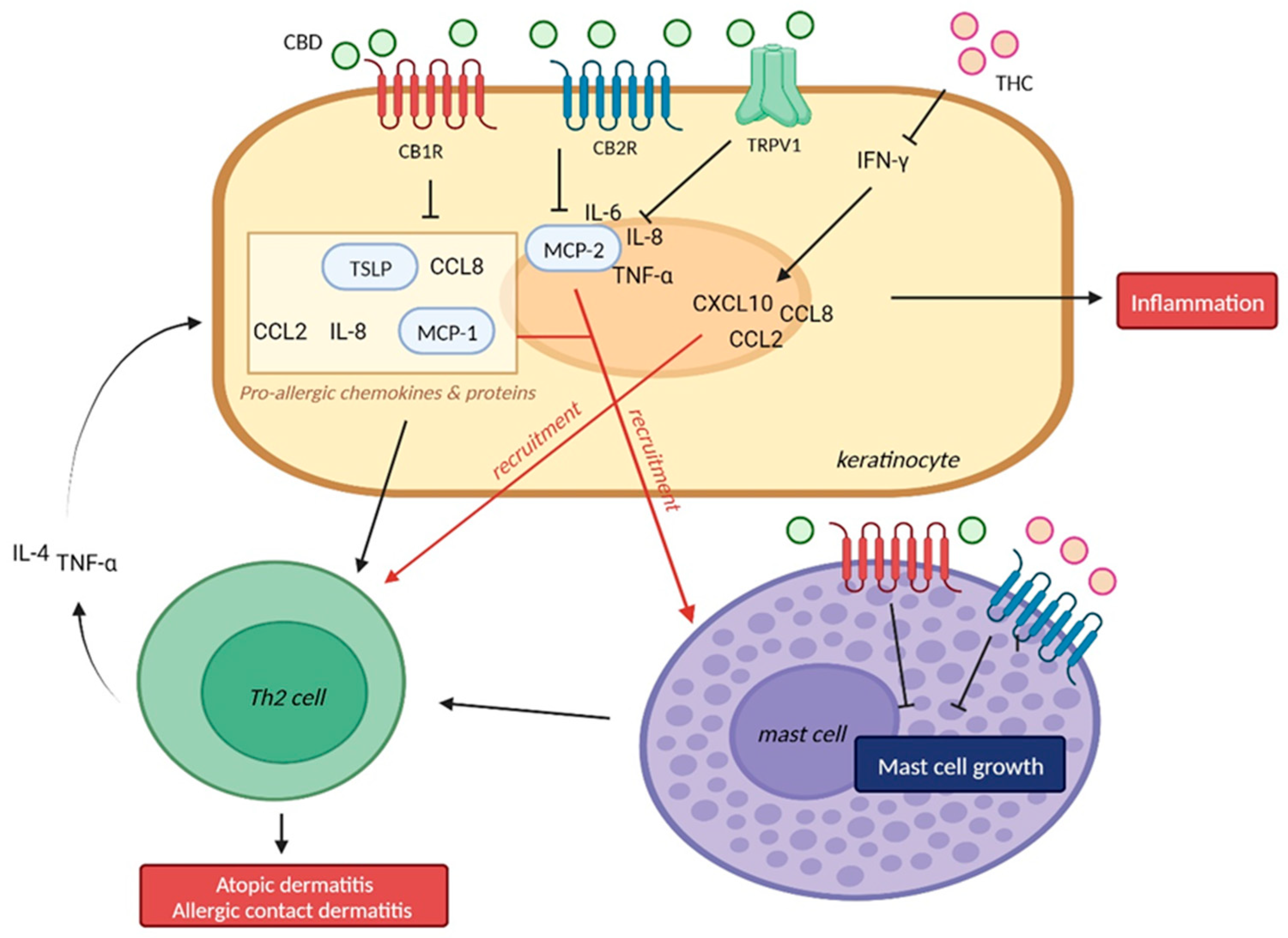
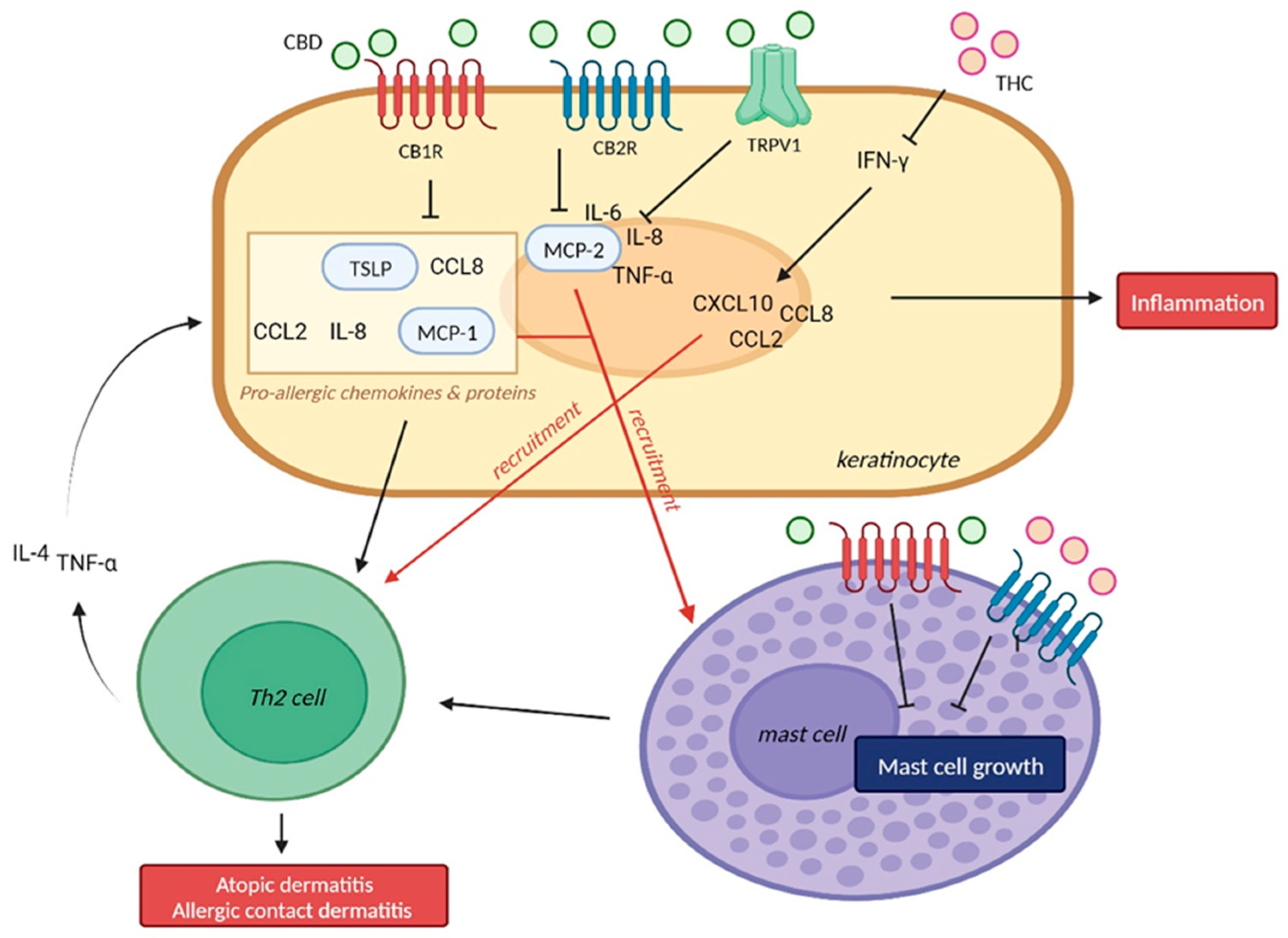
2. Eczema, Atopic Dermatitis, Allergic Contact Dermatitis
2.1. Allergic Contact Dermatitis
2.1.1. Cannabidiol (CBD) and Allergic Contact Dermatitis
2.1.2. Δ-9-Tetrahydrocannabinol (THC) and Allergic Contact Dermatitis
2.1.3. CB1R/CB2R and Allergic Contact Dermatitis
2.2. Atopic Dermatitis
2.2.1. CB1R/CB2R and Atopic Dermatitis
2.2.2. Palmitoylethanolamide (PEA) and Atopic Dermatitis
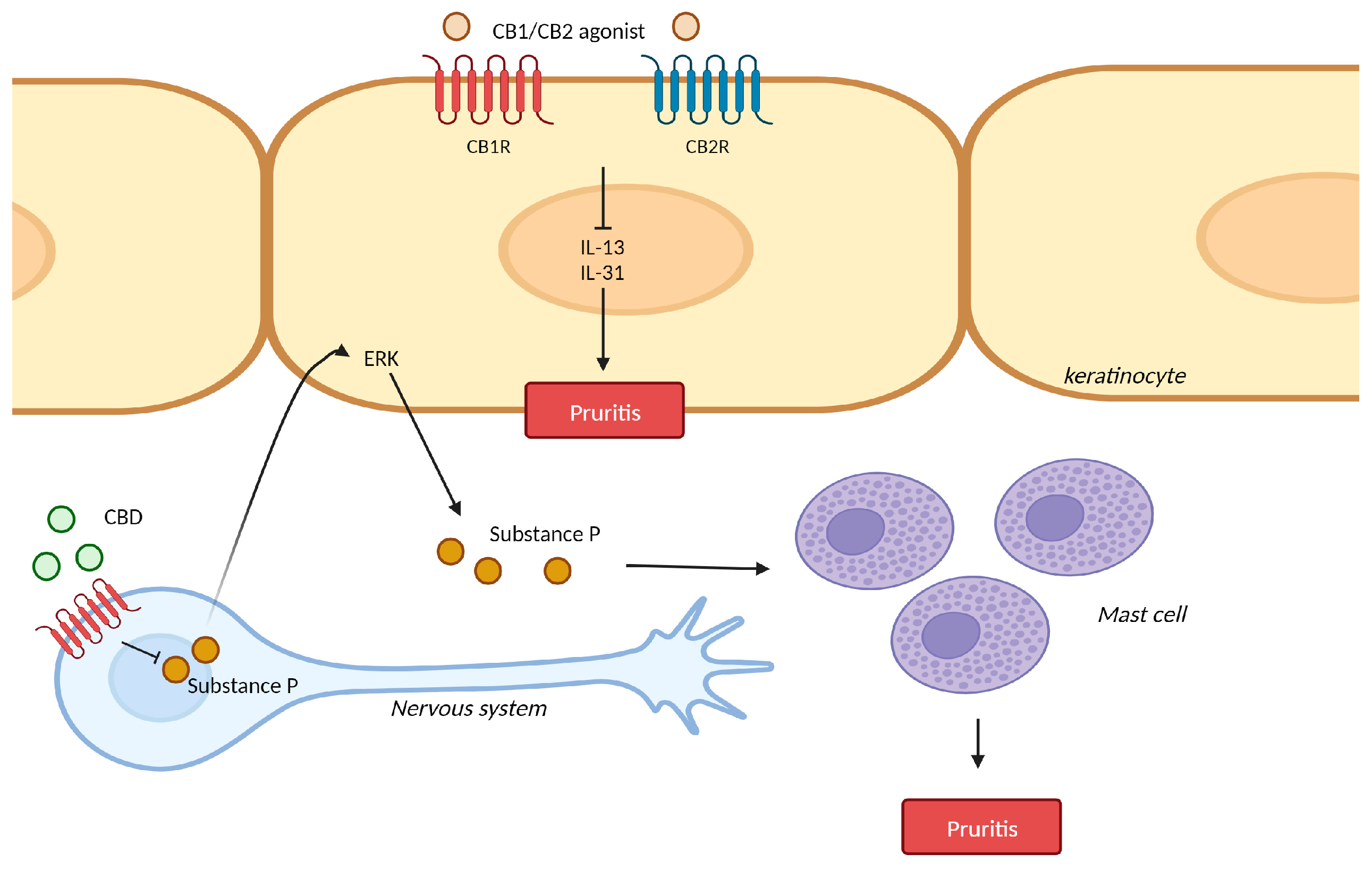
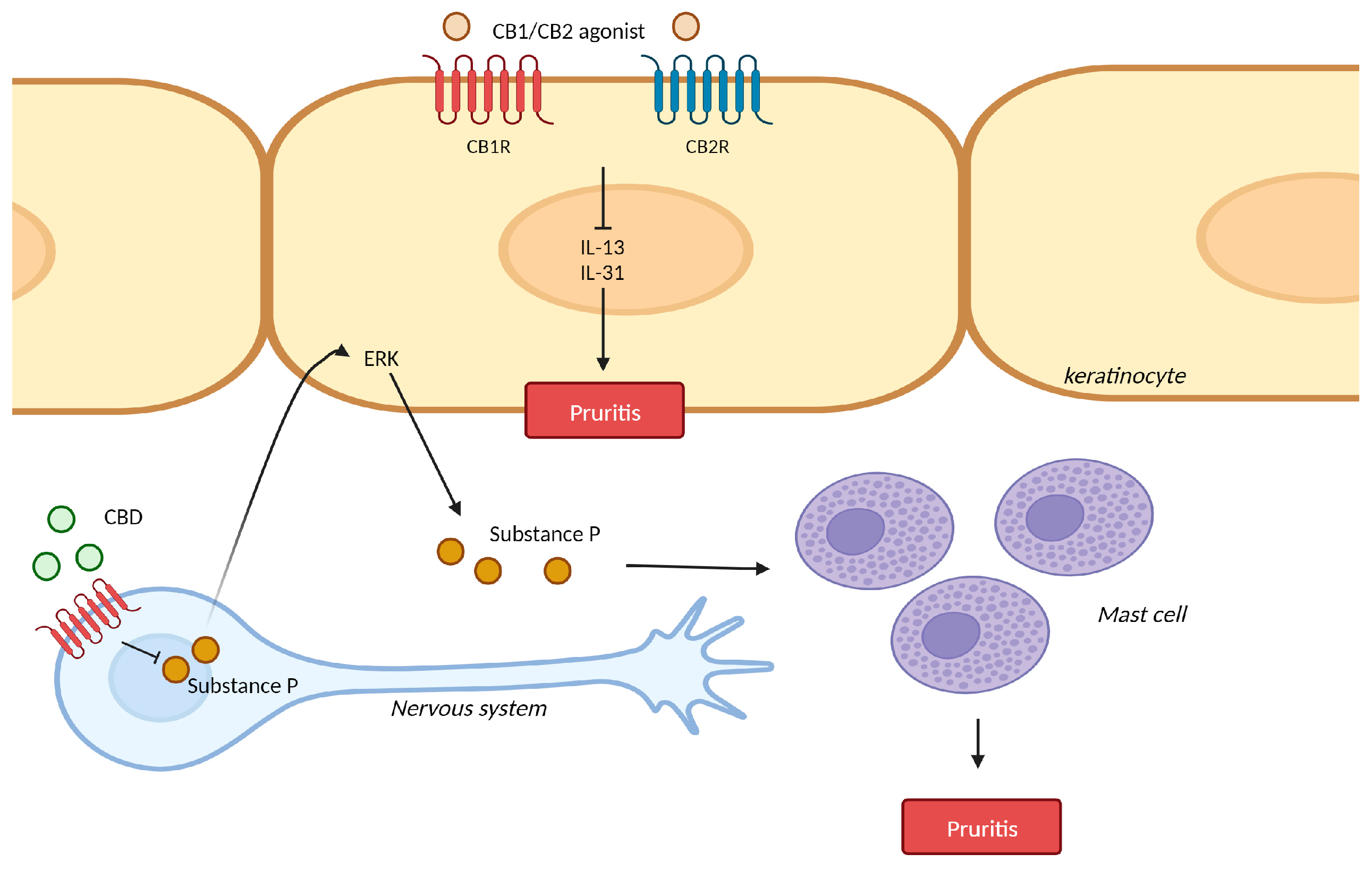
3. Pruritis
3.1. Pruritis and Cannabinoid Receptors
3.2. Pruritis and Cannabinoid Receptor Agonist
3.3. Pruritis and Transient Receptor Potential (TRP) Ion Channels
3.4. Pruritis and Δ-9-tetrahydrocannabinol (THC) Ion Channels
3.5. Pruritis and Fatty Acid Amide Hydrolase (FAAH)
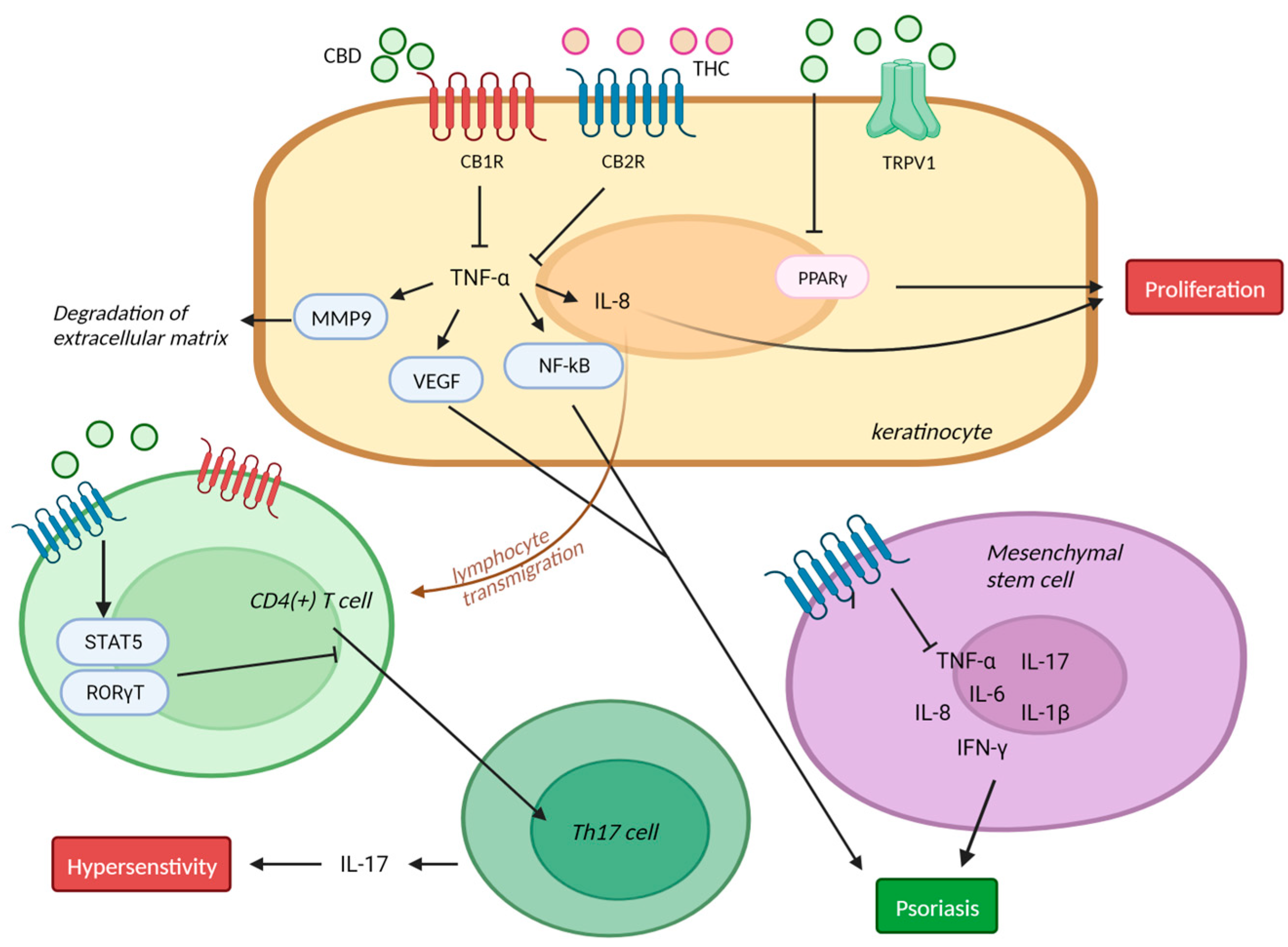
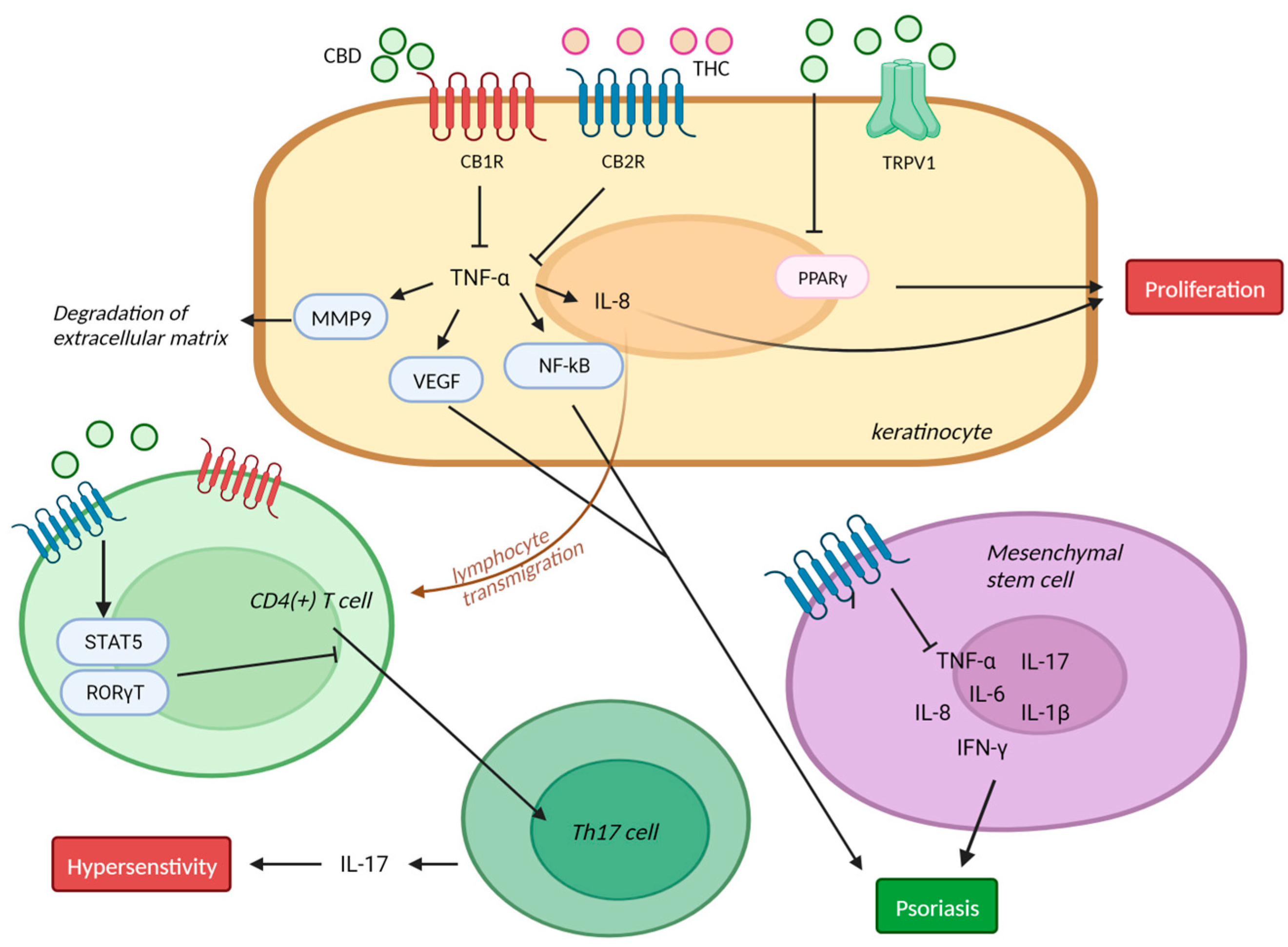
4. Psoriasis
4.1. Psoriasis and Endocannbinoids
4.2. Psoriasis and Cannabinoid Receptors
4.3. Psoriasis and CBD in Clinical Trials
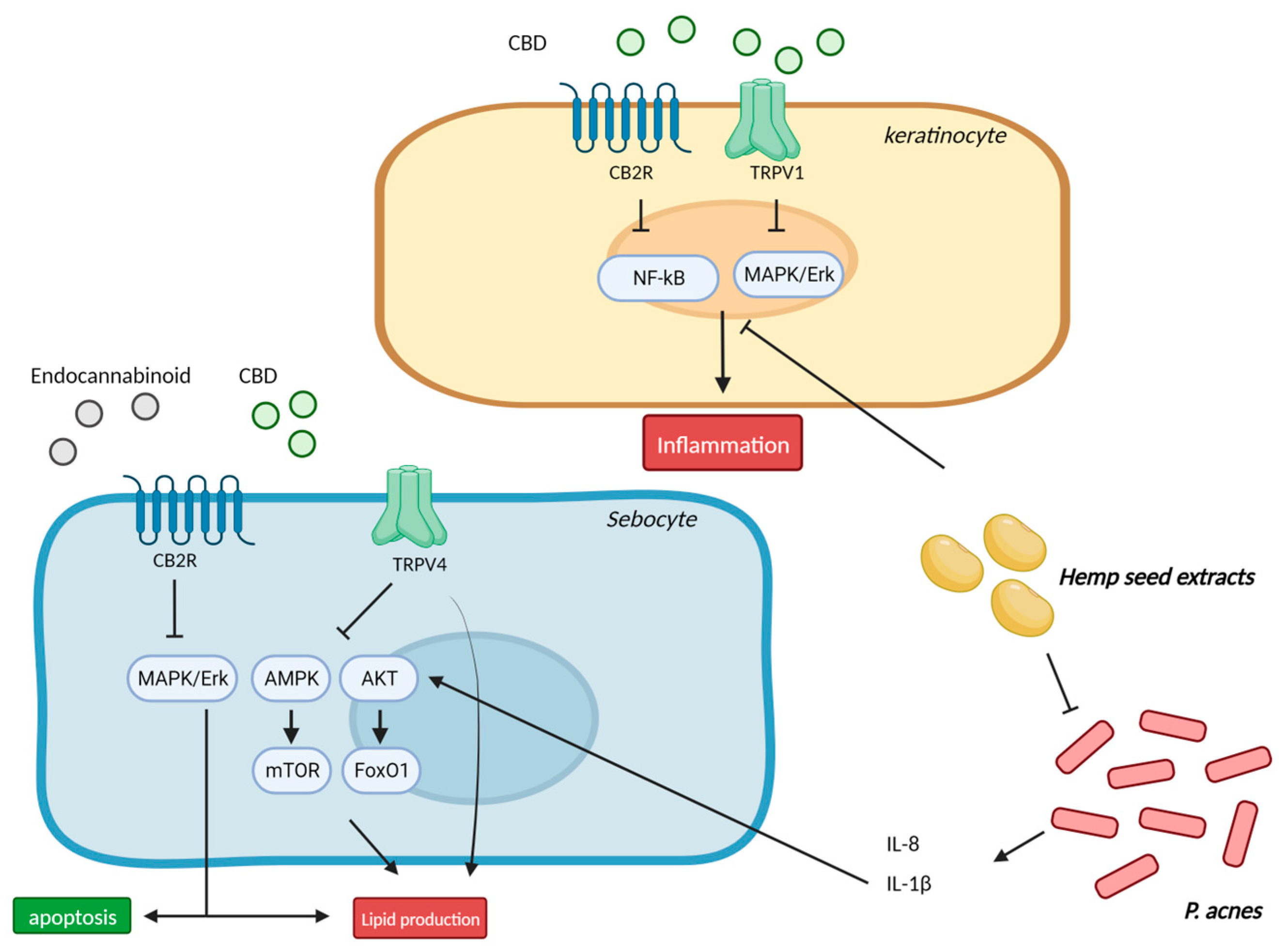
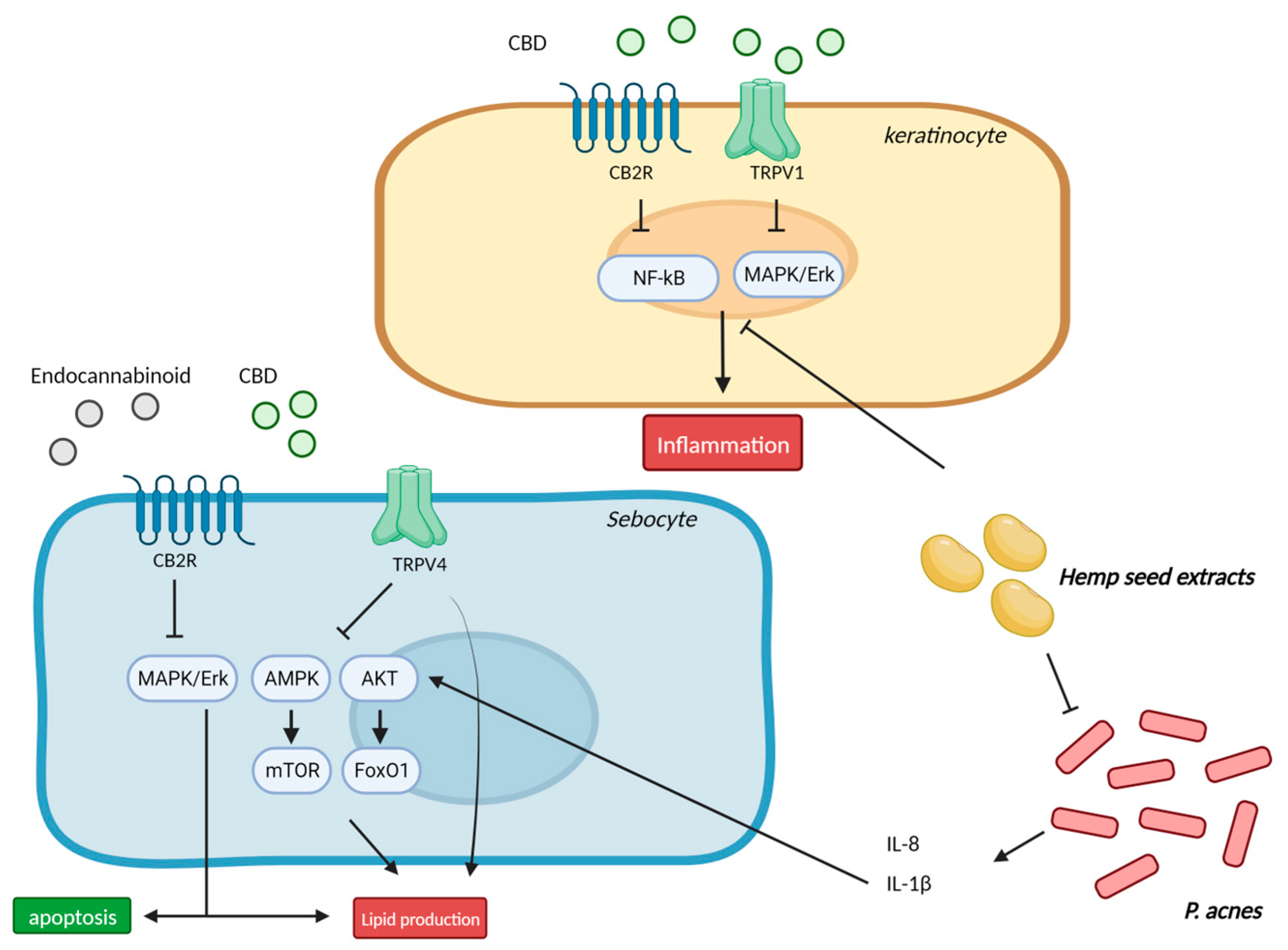
5. Acne and Seborrhea
5.1. Cannabinoids and Inflammation in Sebocytes
5.2. Cannabinoids in Acne and Seborrhea—Clinical Studies
6. Hair Growth
6.1. Cannabinoids Receptor 1 (CB1R) and Hair Growth
6.2. Cannabidiol (CBD) and Hair Growth
6.3. Bimatoprost and Hair Growth
7. Skin Cancer
7.1. Cannabinoids and Melanoma
7.2. Cannabinoids and Kaposi Sarcoma
8. Skin Aging
8.1. Cannabinoids Receptor (CB1R) and Anti-Aging
8.2. CBD and Anti-Aging
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dhadwal, G.; Kirchhof, M.G. The Risks and Benefits of Cannabis in the Dermatology Clinic. J. Cutan. Med. Surg. 2018, 22, 194–199. [Google Scholar] [CrossRef]
- Sheriff, T.; Lin, M.J.; Dubin, D.; Khorasani, H. The potential role of cannabinoids in dermatology. J. Dermatol. Treat. 2020, 31, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Eagleston, L.R.M.; Kalani, N.K.; Patel, R.R.; Flaten, H.K.; Dunnick, C.A.; Dellavalle, R.P. Cannabinoids in dermatology: A scoping review. Dermatol. Online J. 2018, 24, 1. [Google Scholar] [CrossRef]
- Gupta, A.K.; Talukder, M. Cannabinoids for skin diseases and hair regrowth. J. Cosmet. Dermatol. 2021, 20, 2703–2711. [Google Scholar] [CrossRef]
- Tóth, K.F.; Ádám, D.; Bíró, T.; Oláh, A. Cannabinoid Signaling in the Skin: Therapeutic Potential of the “C(ut)annabinoid” System. Molecules 2019, 24, 918. [Google Scholar] [CrossRef] [PubMed]
- Nikan, M.; Nabavi, S.M.; Manayi, A. Ligands for cannabinoid receptors, promising anticancer agents. Life Sci. 2016, 146, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Di Rienzo, M.; Battista, N.; Gasperi, V.; Guerrieri, P.; Rossi, A.; Finazzi-Agrò, A. The endocannabinoid system in human keratinocytes. Evidence that anandamide inhibits epidermal differentiation through CB1 receptor-dependent inhibition of protein kinase C, activation protein-1, and transglutaminase. J. Biol. Chem. 2003, 278, 33896–33903. [Google Scholar] [CrossRef]
- Ständer, S.; Schmelz, M.; Metze, D.; Luger, T.; Rukwied, R. Distribution of cannabinoid receptor 1 (CB1) and 2 (CB2) on sensory nerve fibers and adnexal structures in human skin. J. Dermatol. Sci. 2005, 38, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Samson, M.T.; Small-Howard, A.; Shimoda, L.M.; Koblan-Huberson, M.; Stokes, A.J.; Turner, H. Differential roles of CB1 and CB2 cannabinoid receptors in mast cells. J. Immunol. 2003, 170, 4953–4962. [Google Scholar] [CrossRef]
- Río, C.D.; Millán, E.; García, V.; Appendino, G.; DeMesa, J.; Muñoz, E. The endocannabinoid system of the skin. A potential approach for the treatment of skin disorders. Biochem. Pharmacol. 2018, 157, 122–133. [Google Scholar] [CrossRef]
- Perálvarez-Marín, A.; Doñate-Macian, P.; Gaudet, R. What do we know about the transient receptor potential vanilloid 2 (TRPV2) ion channel? FEBS J. 2013, 280, 5471–5487. [Google Scholar] [CrossRef] [PubMed]
- Baswan, S.M.; Klosner, A.E.; Glynn, K.; Rajgopal, A.; Malik, K.; Yim, S.; Stern, N. Therapeutic Potential of Cannabidiol (CBD) for Skin Health and Disorders. Clin. Cosmet. Investig. Dermatol. 2020, 13, 927–942. [Google Scholar] [CrossRef]
- Petrosino, S.; Verde, R.; Vaia, M.; Allarà, M.; Iuvone, T.; Di Marzo, V. Anti-inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis. J. Pharmacol. Exp. Ther. 2018, 365, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Maghfour, J.; Rietcheck, H.R.; Rundle, C.W.; Runion, T.M.; Jafri, Z.A.; Dercon, S.; Lio, P.; Fernandez, J.; Fujita, M.; Dellavalle, R.P.; et al. An Observational Study of the Application of a Topical Cannabinoid Gel on Sensitive Dry Skin. J. Drugs Dermatol. 2020, 19, 1204–1208. [Google Scholar] [CrossRef]
- Palmieri, B.; Laurino, C.; Vadalà, M. A therapeutic effect of cbd-enriched ointment in inflammatory skin diseases and cutaneous scars. Clin. Ter. 2019, 170, e93–e99. [Google Scholar]
- Callaway, J.; Schwab, U.; Harvima, I.; Halonen, P.; Mykkänen, O.; Hyvönen, P.; Järvinen, T. Efficacy of dietary hempseed oil in patients with atopic dermatitis. J. Dermatol. Treat. 2005, 16, 87–94. [Google Scholar] [CrossRef]
- Gaffal, E.; Cron, M.; Glodde, N.; Tüting, T. Anti-inflammatory activity of topical THC in DNFB-mediated mouse allergic contact dermatitis independent of CB1 and CB2 receptors. Allergy 2013, 68, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Karsak, M.; Gaffal, E.; Date, R.; Wang-Eckhardt, L.; Rehnelt, J.; Petrosino, S.; Starowicz, K.; Steuder, R.; Schlicker, E.; Cravatt, B.; et al. Attenuation of allergic contact dermatitis through the endocannabinoid system. Science 2007, 316, 1494–1497. [Google Scholar] [CrossRef] [PubMed]
- Leonti, M.; Casu, L.; Raduner, S.; Cottiglia, F.; Floris, C.; Altmann, K.H.; Gertsch, J. Falcarinol is a covalent cannabinoid CB1 receptor antagonist and induces pro-allergic effects in skin. Biochem. Pharmacol. 2010, 79, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Wakui, J.; Gokoh, M.; Kishimoto, S.; Sugiura, T. Suppression by WIN55212-2, a cannabinoid receptor agonist, of inflammatory reactions in mouse ear: Interference with the actions of an endogenous ligand, 2-arachidonoylglycerol. Eur. J. Pharmacol. 2006, 538, 154–162. [Google Scholar] [CrossRef]
- Ueda, Y.; Miyagawa, N.; Matsui, T.; Kaya, T.; Iwamura, H. Involvement of cannabinoid CB(2) receptor-mediated response and efficacy of cannabinoid CB(2) receptor inverse agonist, JTE-907, in cutaneous inflammation in mice. Eur. J. Pharmacol. 2005, 520, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, B.; Park, B.M.; Jeon, J.E.; Lee, S.H.; Mann, S.; Ahn, S.K.; Hong, S.P.; Jeong, S.K. Topical cannabinoid receptor 1 agonist attenuates the cutaneous inflammatory responses in oxazolone-induced atopic dermatitis model. Int. J. Dermatol. 2015, 54, e401–e408. [Google Scholar] [CrossRef]
- Gaffal, E.; Glodde, N.; Jakobs, M.; Bald, T.; Tüting, T. Cannabinoid 1 receptors in keratinocytes attenuate fluorescein isothiocyanate-induced mouse atopic-like dermatitis. Exp. Dermatol. 2014, 23, 401–406. [Google Scholar] [CrossRef]
- Nam, G.; Jeong, S.K.; Park, B.M.; Lee, S.H.; Kim, H.J.; Hong, S.P.; Kim, B.; Kim, B.W. Selective Cannabinoid Receptor-1 Agonists Regulate Mast Cell Activation in an Oxazolone-Induced Atopic Dermatitis Model. Ann. Dermatol. 2016, 28, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Nojima, H.; Kuraishi, Y.; Aisaka, K. The cannabinoid CB2 receptor inverse agonist JTE-907 suppresses spontaneous itch-associated responses of NC mice, a model of atopic dermatitis. Eur. J. Pharmacol. 2006, 542, 179–183. [Google Scholar] [CrossRef]
- Roelandt, T.; Heughebaert, C.; Bredif, S.; Giddelo, C.; Baudouin, C.; Msika, P.; Roseeuw, D.; Uchida, Y.; Elias, P.M.; Hachem, J.P. Cannabinoid receptors 1 and 2 oppositely regulate epidermal permeability barrier status and differentiation. Exp. Dermatol. 2012, 21, 688–693. [Google Scholar] [CrossRef]
- Clayton, P.; Subah, S.; Venkatesh, R.; Hill, M.; Bogoda, N. Palmitoylethanolamide: A Potential Alternative to Cannabidiol. J. Diet. Suppl. 2023, 20, 505–530. [Google Scholar] [CrossRef] [PubMed]
- Kircik, L. A nonsteroidal lamellar matrix cream containing palmitoylethanolamide for the treatment of atopic dermatitis. J. Drugs Dermatol. 2010, 9, 334–338. [Google Scholar]
- Yuan, C.; Wang, X.M.; Guichard, A.; Tan, Y.M.; Qian, C.Y.; Yang, L.J.; Humbert, P. N-palmitoylethanolamine and N-acetylethanolamine are effective in asteatotic eczema: Results of a randomized, double-blind, controlled study in 60 patients. Clin. Interv. Aging 2014, 9, 1163–1169. [Google Scholar] [CrossRef]
- Del Rosso, J.Q. Use of a palmitoylethanolamide-containing nonsteroidal cream for treating atopic dermatitis: Impact on the duration of response and time between flares. Cosmet. Dermatol. 2007, 20, 4. [Google Scholar]
- Eberlein, B.; Eicke, C.; Reinhardt, H.W.; Ring, J. Adjuvant treatment of atopic eczema: Assessment of an emollient containing N-palmitoylethanolamine (ATOPA study). J. Eur. Acad. Dermatol. Venereol. 2008, 22, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Pulvirenti, N.; Nasca, M.R.; Micali, G. Topical adelmidrol 2% emulsion, a novel aliamide, in the treatment of mild atopic dermatitis in pediatric subjects: A pilot study. Acta Dermatovenerol. Croat. 2007, 15, 80–83. [Google Scholar] [PubMed]
- Liu, X.; Li, L.; Jiang, J.; Ge, W.; Huang, Y.; Jin, Z.; Liu, X.; Kong, Y.; Zhanmu, O.; Zeng, X.; et al. Role of Type I Cannabinoid Receptor in Sensory Neurons in Psoriasiform Skin Inflammation and Pruritus. J. Investig. Dermatol. 2023, 143, 812–821.e3. [Google Scholar] [CrossRef]
- Li, L.; Liu, X.; Ge, W.; Chen, C.; Huang, Y.; Jin, Z.; Zhan, M.; Duan, X.; Liu, X.; Kong, Y.; et al. CB2R Deficiency Exacerbates Imiquimod-Induced Psoriasiform Dermatitis and Itch through the Neuro-Immune Pathway. Front. Pharmacol. 2022, 13, 790712. [Google Scholar] [CrossRef] [PubMed]
- Todurga Seven, Z.G.; Çakır Gündoğdu, A.; Ozyurt, R.; Özyazgan, S. The Effects of Cannabinoid Agonist, Heat Shock Protein 90 and Nitric Oxide Synthase Inhibitors on Increasing IL-13 and IL-31 Levels in Chronic Pruritus. Immunol. Investig. 2022, 51, 1938–1949. [Google Scholar] [CrossRef] [PubMed]
- Bilir, K.A.; Anli, G.; Ozkan, E.; Gunduz, O.; Ulugol, A. Involvement of spinal cannabinoid receptors in the antipruritic effects of WIN 55,212-2, a cannabinoid receptor agonist. Clin. Exp. Dermatol. 2018, 43, 553–558. [Google Scholar] [CrossRef]
- Odan, M.; Ishizuka, N.; Hiramatsu, Y.; Inagaki, M.; Hashizume, H.; Fujii, Y.; Mitsumori, S.; Morioka, Y.; Soga, M.; Deguchi, M.; et al. Discovery of S-777469: An orally available CB2 agonist as an antipruritic agent. Bioorg. Med. Chem. Lett. 2012, 22, 2803–2806. [Google Scholar] [CrossRef]
- Dvorak, M.; Watkinson, A.; McGlone, F.; Rukwied, R. Histamine induced responses are attenuated by a cannabinoid receptor agonist in human skin. Inflamm. Res. 2003, 52, 238–245. [Google Scholar] [CrossRef]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 2018, 11, 487. [Google Scholar] [CrossRef]
- Yun, J.W.; Seo, J.A.; Jang, W.H.; Koh, H.J.; Bae, I.H.; Park, Y.H.; Lim, K.M. Antipruritic effects of TRPV1 antagonist in murine atopic dermatitis and itching models. J. Investig. Dermatol. 2011, 131, 1576–1579. [Google Scholar] [CrossRef]
- Imamachi, N.; Park, G.H.; Lee, H.; Anderson, D.J.; Simon, M.I.; Basbaum, A.I.; Han, S.K. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11330–11335. [Google Scholar] [CrossRef] [PubMed]
- Ambrosino, P.; Soldovieri, M.V.; Russo, C.; Taglialatela, M. Activation and desensitization of TRPV1 channels in sensory neurons by the PPARα agonist palmitoylethanolamide. Br. J. Pharmacol. 2013, 168, 1430–1444. [Google Scholar] [CrossRef] [PubMed]
- Szepietowski, J.C.; Reich, A.; Szepietowski, T. Emollients with endocannabinoids in the treatment of uremic pruritus: Discussion of the therapeutic options. Ther. Apher. Dial. 2005, 9, 277–279. [Google Scholar] [CrossRef]
- Szepietowski, J.C.; Szepietowski, T.; Reich, A. Efficacy and tolerance of the cream containing structured physiological lipids with endocannabinoids in the treatment of uremic pruritus: A preliminary study. Acta Dermatovenerol. Croat. 2005, 13, 97–103. [Google Scholar] [PubMed]
- Ständer, S.; Reinhardt, H.W.; Luger, T.A. Topical cannabinoid agonists. An effective new possibility for treating chronic pruritus. Hautarzt 2006, 57, 801–807. [Google Scholar] [CrossRef]
- Visse, K.; Blome, C.; Phan, N.Q.; Augustin, M.; Ständer, S. Efficacy of Body Lotion Containing N-palmitoylethanolamine in Subjects with Chronic Pruritus due to Dry Skin: A Dermatocosmetic Study. Acta Derm.-Venereol. 2017, 97, 639–641. [Google Scholar] [CrossRef]
- Schräder, N.H.B.; Duipmans, J.C.; Molenbuur, B.; Wolff, A.P.; Jonkman, M.F. Combined tetrahydrocannabinol and cannabidiol to treat pain in epidermolysis bullosa: A report of three cases. Br. J. Dermatol. 2019, 180, 922–924. [Google Scholar] [CrossRef]
- Neff, G.W.; O’Brien, C.B.; Reddy, K.R.; Bergasa, N.V.; Regev, A.; Molina, E.; Amaro, R.; Rodriguez, M.J.; Chase, V.; Jeffers, L.; et al. Preliminary observation with dronabinol in patients with intractable pruritus secondary to cholestatic liver disease. Am. J. Gastroenterol. 2002, 97, 2117–2119. [Google Scholar] [CrossRef]
- Schlosburg, J.E.; Boger, D.L.; Cravatt, B.F.; Lichtman, A.H. Endocannabinoid modulation of scratching response in an acute allergenic model: A new prospective neural therapeutic target for pruritus. J. Pharmacol. Exp. Ther. 2009, 329, 314–323. [Google Scholar] [CrossRef]
- Yesilyurt, O.; Cayirli, M.; Sakin, Y.S.; Seyrek, M.; Akar, A.; Dogrul, A. Systemic and spinal administration of FAAH, MAGL inhibitors and dual FAAH/MAGL inhibitors produce antipruritic effect in mice. Arch. Dermatol. Res. 2016, 308, 335–345. [Google Scholar] [CrossRef]
- Tosun, N.C.; Gunduz, O.; Ulugol, A. Attenuation of serotonin-induced itch responses by inhibition of endocannabinoid degradative enzymes, fatty acid amide hydrolase and monoacylglycerol lipase. J. Neural Transm. 2015, 122, 363–367. [Google Scholar] [CrossRef]
- Namazi, M.R. Cannabinoids, loratadine and allopurinol as novel additions to the antipsoriatic ammunition. J. Eur. Acad. Dermatol. Venereol. 2005, 19, 319–322. [Google Scholar] [CrossRef]
- Wilkinson, J.D.; Williamson, E.M. Cannabinoids inhibit human keratinocyte proliferation through a non-CB1/CB2 mechanism and have a potential therapeutic value in the treatment of psoriasis. J. Dermatol. Sci. 2007, 45, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Tóth, B.I.; Dobrosi, N.; Dajnoki, A.; Czifra, G.; Oláh, A.; Szöllosi, A.G.; Juhász, I.; Sugawara, K.; Paus, R.; Bíró, T. Endocannabinoids modulate human epidermal keratinocyte proliferation and survival via the sequential engagement of cannabinoid receptor-1 and transient receptor potential vanilloid-1. J. Investig. Dermatol. 2011, 131, 1095–1104. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Fumagalli, M.; Pacchetti, B.; Piazza, S.; Magnavacca, A.; Khalilpour, S.; Melzi, G.; Martinelli, G.; Dell’Agli, M. Cannabis sativa L. extract and cannabidiol inhibit in vitro mediators of skin inflammation and wound injury. Phytother. Res. 2019, 33, 2083–2093. [Google Scholar] [CrossRef] [PubMed]
- Körver, J.E.; van Duijnhoven, M.W.; Pasch, M.C.; van Erp, P.E.; van de Kerkhof, P.C. Assessment of epidermal subpopulations and proliferation in healthy skin, symptomless and lesional skin of spreading psoriasis. Br. J. Dermatol. 2006, 155, 688–694. [Google Scholar] [CrossRef]
- Ramot, Y.; Sugawara, K.; Zákány, N.; Tóth, B.I.; Bíró, T.; Paus, R. A novel control of human keratin expression: Cannabinoid receptor 1-mediated signaling down-regulates the expression of keratins K6 and K16 in human keratinocytes in vitro and in situ. PeerJ 2013, 1, e40. [Google Scholar] [CrossRef]
- Guillot, A.; Hamdaoui, N.; Bizy, A.; Zoltani, K.; Souktani, R.; Zafrani, E.S.; Mallat, A.; Lotersztajn, S.; Lafdil, F. Cannabinoid receptor 2 counteracts interleukin-17-induced immune and fibrogenic responses in mouse liver. Hepatology 2014, 59, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Bernardo, M.E.; Bellini, G.; Luongo, L.; Conforti, A.; Manzo, I.; Guida, F.; Cristino, L.; Imperatore, R.; Petrosino, S.; et al. The cannabinoid receptor type 2 as mediator of mesenchymal stromal cell immunosuppressive properties. PLoS ONE 2013, 8, e80022. [Google Scholar] [CrossRef]
- Friedman, A.J.; Momeni, K.; Kogan, M. Topical Cannabinoids for the Management of Psoriasis Vulgaris: Report of a Case and Review of the Literature. J. Drugs Dermatol. 2020, 19, 795. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, C.; Tosti, A. Efficacy and Tolerability of a Shampoo Containing Broad-Spectrum Cannabidiol in the Treatment of Scalp Inflammation in Patients with Mild to Moderate Scalp Psoriasis or Seborrheic Dermatitis. Skin Appendage Disord. 2020, 6, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Puaratanaarunkon, T.; Sittisaksomjai, S.; Sivapornpan, N.; Pongcharoen, P.; Chakkavittumrong, P.; Ingkaninan, K.; Temkitthawon, P.; Promgool, T.; Waranuch, N.; Asawanonda, P. Topical cannabidiol-based treatment for psoriasis: A dual-centre randomized placebo-controlled study. J. Eur. Acad. Dermatol. Venereol. 2022, 36, e718–e720. [Google Scholar] [CrossRef]
- Dobrosi, N.; Tóth, B.I.; Nagy, G.; Dózsa, A.; Géczy, T.; Nagy, L.; Zouboulis, C.C.; Paus, R.; Kovács, L.; Bíró, T. Endocannabinoids enhance lipid synthesis and apoptosis of human sebocytes via cannabinoid receptor-2-mediated signaling. FASEB J. 2008, 22, 3685–3695. [Google Scholar] [CrossRef] [PubMed]
- Oláh, A.; Markovics, A.; Szabó-Papp, J.; Szabó, P.T.; Stott, C.; Zouboulis, C.C.; Bíró, T. Differential effectiveness of selected non-psychotropic phytocannabinoids on human sebocyte functions implicates their introduction in dry/seborrhoeic skin and acne treatment. Exp. Dermatol. 2016, 25, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Oláh, A.; Tóth, B.I.; Borbíró, I.; Sugawara, K.; Szöllõsi, A.G.; Czifra, G.; Pál, B.; Ambrus, L.; Kloepper, J.; Camera, E.; et al. Cannabidiol exerts sebostatic and antiinflammatory effects on human sebocytes. J. Clin. Investig. 2014, 124, 3713–3724. [Google Scholar] [CrossRef]
- Jin, S.; Lee, M.Y. The ameliorative effect of hemp seed hexane extracts on the Propionibacterium acnes-induced inflammation and lipogenesis in sebocytes. PLoS ONE 2018, 13, e0202933. [Google Scholar] [CrossRef]
- Jiang, Z.; Jin, S.; Fan, X.; Cao, K.; Liu, Y.; Wang, X.; Ma, Y.; Xiang, L. Cannabidiol Inhibits Inflammation Induced by Cutibacterium acnes-Derived Extracellular Vesicles via Activation of CB2 Receptor in Keratinocytes. J. Inflamm. Res. 2022, 15, 4573–4583. [Google Scholar] [CrossRef]
- Ali, A.; Akhtar, N. The safety and efficacy of 3% Cannabis seeds extract cream for reduction of human cheek skin sebum and erythema content. Pak. J. Pharm. Sci. 2015, 28, 1389–1395. [Google Scholar] [PubMed]
- Shao, K.; Stewart, C.; Grant-Kels, J.M. Cannabis and the skin. Clin. Dermatol. 2021, 39, 784–795. [Google Scholar] [CrossRef]
- Krause, K.; Foitzik, K. Biology of the hair follicle: The basics. Semin. Cutan. Med. Surg. 2006, 25, 2–10. [Google Scholar] [CrossRef]
- Telek, A.; Bíró, T.; Bodó, E.; Tóth, B.I.; Borbíró, I.; Kunos, G.; Paus, R. Inhibition of human hair follicle growth by endo- and exocannabinoids. FASEB J. 2007, 21, 3534–3541. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, B.K.; Soni, R.; Patel, J.Z.; Joharapurkar, A.; Sadhwani, N.; Kshirsagar, S.; Mishra, B.; Takale, V.; Gupta, S.; Pandya, P.; et al. Hair growth stimulator property of thienyl substituted pyrazole carboxamide derivatives as a CB1 receptor antagonist with in vivo antiobesity effect. Bioorg. Med. Chem. Lett. 2009, 19, 2546–2550. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.L.; Lisztes, E.; Béke, G.; Tóth, K.F.; Paus, R.; Oláh, A.; Bíró, T. The Phytocannabinoid (−)-Cannabidiol Operates as a Complex, Differential Modulator of Human Hair Growth: Anti-Inflammatory Submicromolar versus Hair Growth Inhibitory Micromolar Effects. J. Investig. Dermatol. 2020, 140, 484–488.e5. [Google Scholar] [CrossRef]
- Smith, G.L.; Satino, J. Hair Regrowth with Cannabidiol (CBD)-rich Hemp Extract—A Case Series. Cannabis 2021, 4, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Chanasumon, N.; Sriphojanart, T.; Suchonwanit, P. Therapeutic potential of bimatoprost for the treatment of eyebrow hypotrichosis. Drug Des. Dev. Ther. 2018, 12, 365–372. [Google Scholar] [CrossRef]
- Khidhir, K.G.; Woodward, D.F.; Farjo, N.P.; Farjo, B.K.; Tang, E.S.; Wang, J.W.; Picksley, S.M.; Randall, V.A. The prostamide-related glaucoma therapy, bimatoprost, offers a novel approach for treating scalp alopecias. FASEB J. 2013, 27, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Zaher, H.; Gawdat, H.I.; Hegazy, R.A.; Hassan, M. Bimatoprost versus Mometasone Furoate in the Treatment of Scalp Alopecia Areata: A Pilot Study. Dermatology 2015, 230, 308–313. [Google Scholar] [CrossRef]
- Gupta, A.K.; Talukder, M. A cannabinoid Hairy-Tale: Hair loss or hair gain? J. Cosmet. Dermatol. 2022, 21, 6653–6660. [Google Scholar] [CrossRef]
- Pucci, M.; Pasquariello, N.; Battista, N.; Di Tommaso, M.; Rapino, C.; Fezza, F.; Zuccolo, M.; Jourdain, R.; Finazzi Agrò, A.; Breton, L.; et al. Endocannabinoids stimulate human melanogenesis via type-1 cannabinoid receptor. J. Biol. Chem. 2012, 287, 15466–15478. [Google Scholar] [CrossRef]
- Blázquez, C.; Carracedo, A.; Barrado, L.; Real, P.J.; Fernández-Luna, J.L.; Velasco, G.; Malumbres, M.; Guzmán, M. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006, 20, 2633–2635. [Google Scholar] [CrossRef]
- Casanova, M.L.; Blázquez, C.; Martínez-Palacio, J.; Villanueva, C.; Fernández-Aceñero, M.J.; Huffman, J.W.; Jorcano, J.L.; Guzmán, M. Inhibition of skin tumor growth and angiogenesis in vivo by activation of cannabinoid receptors. J. Clin. Investig. 2003, 111, 43–50. [Google Scholar] [CrossRef]
- Simmerman, E.; Qin, X.; Yu, J.C.; Baban, B. Cannabinoids as a Potential New and Novel Treatment for Melanoma: A Pilot Study in a Murine Model. J. Surg. Res. 2019, 235, 210–215. [Google Scholar] [CrossRef]
- Armstrong, J.L.; Hill, D.S.; McKee, C.S.; Hernandez-Tiedra, S.; Lorente, M.; Lopez-Valero, I.; Eleni Anagnostou, M.; Babatunde, F.; Corazzari, M.; Redfern, C.P.F.; et al. Exploiting cannabinoid-induced cytotoxic autophagy to drive melanoma cell death. J. Investig. Dermatol. 2015, 135, 1629–1637. [Google Scholar] [CrossRef]
- Luca, T.; Di Benedetto, G.; Scuderi, M.R.; Palumbo, M.; Clementi, S.; Bernardini, R.; Cantarella, G. The CB1/CB2 receptor agonist WIN-55,212-2 reduces viability of human Kaposi’s sarcoma cells in vitro. Eur. J. Pharmacol. 2009, 616, 16–21. [Google Scholar] [CrossRef]
- Maor, Y.; Yu, J.; Kuzontkoski, P.M.; Dezube, B.J.; Zhang, X.; Groopman, J.E. Cannabidiol inhibits growth and induces programmed cell death in kaposi sarcoma-associated herpesvirus-infected endothelium. Genes Cancer 2012, 3, 512–520. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.F.; Kunos, G.; Groopman, J.E. Cannabinoid modulation of Kaposi’s sarcoma-associated herpesvirus infection and transformation. Cancer Res. 2007, 67, 7230–7237. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Drews, E.; Albayram, Ö.; Piyanova, A.; Gaffal, E.; Tueting, T.; Michel, K.; Mauer, D.; Maier, W.; Zimmer, A. Early onset of aging-like changes is restricted to cognitive abilities and skin structure in Cnr1⁻/⁻ mice. Neurobiol. Aging 2012, 33, e11–e22. [Google Scholar] [CrossRef]
- Leal, E.C.; Moura, L.I.; Pirzgalska, R.M.; Marques-da-Silva, D.; Ledent, C.; Köfalvi, A.; Carvalho, E. Diabetes and Cannabinoid CB1 receptor deficiency promote similar early onset aging-like changes in the skin. Exp. Gerontol. 2021, 154, 111528. [Google Scholar] [CrossRef] [PubMed]
- Gerasymchuk, M.; Robinson, G.I.; Groves, A.; Haselhorst, L.; Nandakumar, S.; Stahl, C.; Kovalchuk, O.; Kovalchuk, I. Phytocannabinoids Stimulate Rejuvenation and Prevent Cellular Senescence in Human Dermal Fibroblasts. Cells 2022, 11, 3939. [Google Scholar] [CrossRef] [PubMed]
- Styrczewska, M.; Kostyn, A.; Kulma, A.; Majkowska-Skrobek, G.; Augustyniak, D.; Prescha, A.; Czuj, T.; Szopa, J. Flax Fiber Hydrophobic Extract Inhibits Human Skin Cells Inflammation and Causes Remodeling of Extracellular Matrix and Wound Closure Activation. BioMed Res. Int. 2015, 2015, 862391. [Google Scholar] [CrossRef] [PubMed]
- Casares, L.; García, V.; Garrido-Rodríguez, M.; Millán, E.; Collado, J.A.; García-Martín, A.; Peñarando, J.; Calzado, M.A.; de la Vega, L.; Muñoz, E. Cannabidiol induces antioxidant pathways in keratinocytes by targeting BACH1. Redox Biol. 2020, 28, 101321. [Google Scholar] [CrossRef] [PubMed]
| Study Type | Patients (Sample Size) | Treatment | Outcome | Study Results |
|---|---|---|---|---|
| Observational study (Maghfour, J. et al.) [14] | Atopic dermatitis (n = 20) | Topical application of CBD gel | Patient-oriented eczema measure score (POEM) and Quality of Life Hand Eczema Questionnaire (QOLHEQ). | Significant reduction of POEM (p < 0.0007) and QOLHEQ (p < 0.004) |
| Retrospective study (Palmieri, B. et al.) [15] | Psoriasis (n = 5) Atopic dermatitis (n = 5) Scars (n = 10) | Topical CBD-enriched ointment twice daily for three months | Skin hydration (TEWL) assessed using a DermaLab® device Elasticity assessed using an ElastiMeter® device Photographic assessment | Significant reduction in TEWL (p < 0.001) Significant improvement of elasticity (p < 0.001) Significant improvement in symptoms |
| Randomized, single-blinded, crossover study (Callaway, J. et al.) [16] | Atopic dermatitis (n = 20) | Dietary hemp oil and olive oil for 20 weeks | Fatty acid profiles Skin dryness, itchiness, and use of dermal medications, assessed by patient questionnaire Skin transepidermal water loss (TEWL) | Increased levels of essential fatty acids after hemp oil Improvement in clinical symptoms (p < 0.05) Intra-group TEWL values decreased (p = 0.074) |
| Randomized, double-blind, controlled trial (Yuan, C. et al.) [29] | Asteatotic eczema (n = 60) | Emollient creams containing PEA (N-acylethanolamine) or NEA (N-acetyl ethanolamine) for 28 days | Clinical assessment using Eczema Area and Severity Index Skin surface hydration assessed using Corneometer CM820® | Decreased skin erythema, scaling, dryness, and itching (p < 0.05) Skin surface hydration was increased (p < 0.05) |
| Split-body clinical study (Del Rosso, J.Q. et al.) [30] | Atopic dermatitis (n = 43) | Combination of topical steroid and PEA compared with the sides treated with only topical steroid | Clearance rate | Significant improvement in clearance rate of symptoms |
| Observational, prospective cohort study (Eberlein, B. et al.) [31] | Atopic dermatitis (n = 2456) | Topical application of PEA for 4–6 weeks | Objective symptoms such as dryness, excoriation, lichenification, scaling, erythema, pruritis | Significant improvement with combined score reduction of 58.6% in the entire population (p < 0.001) |
| Observational study (Pulvirenti, N. et al.) [32] | Atopic dermatitis (n = 20) | Topical emulsion containing adelmidrol 2% twice daily for 4 weeks | Clinical symptoms | Complete resolution with no side effects in 80% of patients |
| Observational, non-blinded, prospective cohort study (Szepietowski, J.C. et al.) [43] | Uremic patients (n = 21) | Topical treatment of PEA and anandamide twice daily for 3 weeks | Pruritis and scales assessed by questionnaire | Significant reduction in both pruritis and scales (p < 0.0001) |
| Observational study (Schräder, N.H.B. et al.) [47] | Patients with pruritis (n = 22) | Emollient cream containing PEA | Pruritis | Reduction in itch (86.4%) |
| Randomized, single-blinded study (Visse, K. et al.) [46] | Patients with pruritic dry skin (n = 100) | PEA-containing lotion | Pruritis intensity assessed by visual analogue scale | No significant difference between the PEA-containing lotion and control groups |
| Randomized, placebo-controlled study (Puaratanaarunkon, T. et al.) [62] | Plaque-type psoriasis (n = 51) | 2.5% CBD ointment twice daily for 12 weeks | Psoriasis severity index score (PASI) | Significant reduction of PASI (p = 0.026) |
| Single-blinded study (Ali, A. et al.) [68] | Healthy patients (n = 11) | 3% cannabis-seed-extract cream twice daily for 12 weeks | Sebum and erythema content assessed using Sebumeter and Mexameter | Significant reduction of sebum production and erythema (p < 0.05) |
| Randomized controlled trial (Zaher, H. et al.) [77] | Alopecia areata (n = 30) | Bimatoprost 0.03% solution twice daily for 3 months compared with mometasone furoate 0.1% cream | Severity of Alopecia Tool | Significant improvement of hair-regrowth (p = 0.001) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, E.H.; Lee, J.H. Cannabinoids and Their Receptors in Skin Diseases. Int. J. Mol. Sci. 2023, 24, 16523. https://doi.org/10.3390/ijms242216523
Yoo EH, Lee JH. Cannabinoids and Their Receptors in Skin Diseases. International Journal of Molecular Sciences. 2023; 24(22):16523. https://doi.org/10.3390/ijms242216523
Chicago/Turabian StyleYoo, Eun Hee, and Ji Hyun Lee. 2023. "Cannabinoids and Their Receptors in Skin Diseases" International Journal of Molecular Sciences 24, no. 22: 16523. https://doi.org/10.3390/ijms242216523
APA StyleYoo, E. H., & Lee, J. H. (2023). Cannabinoids and Their Receptors in Skin Diseases. International Journal of Molecular Sciences, 24(22), 16523. https://doi.org/10.3390/ijms242216523