
APOA1 Is a Novel Marker for Preeclampsia

 ,
,
Abstract
:1. Introduction
2. Results
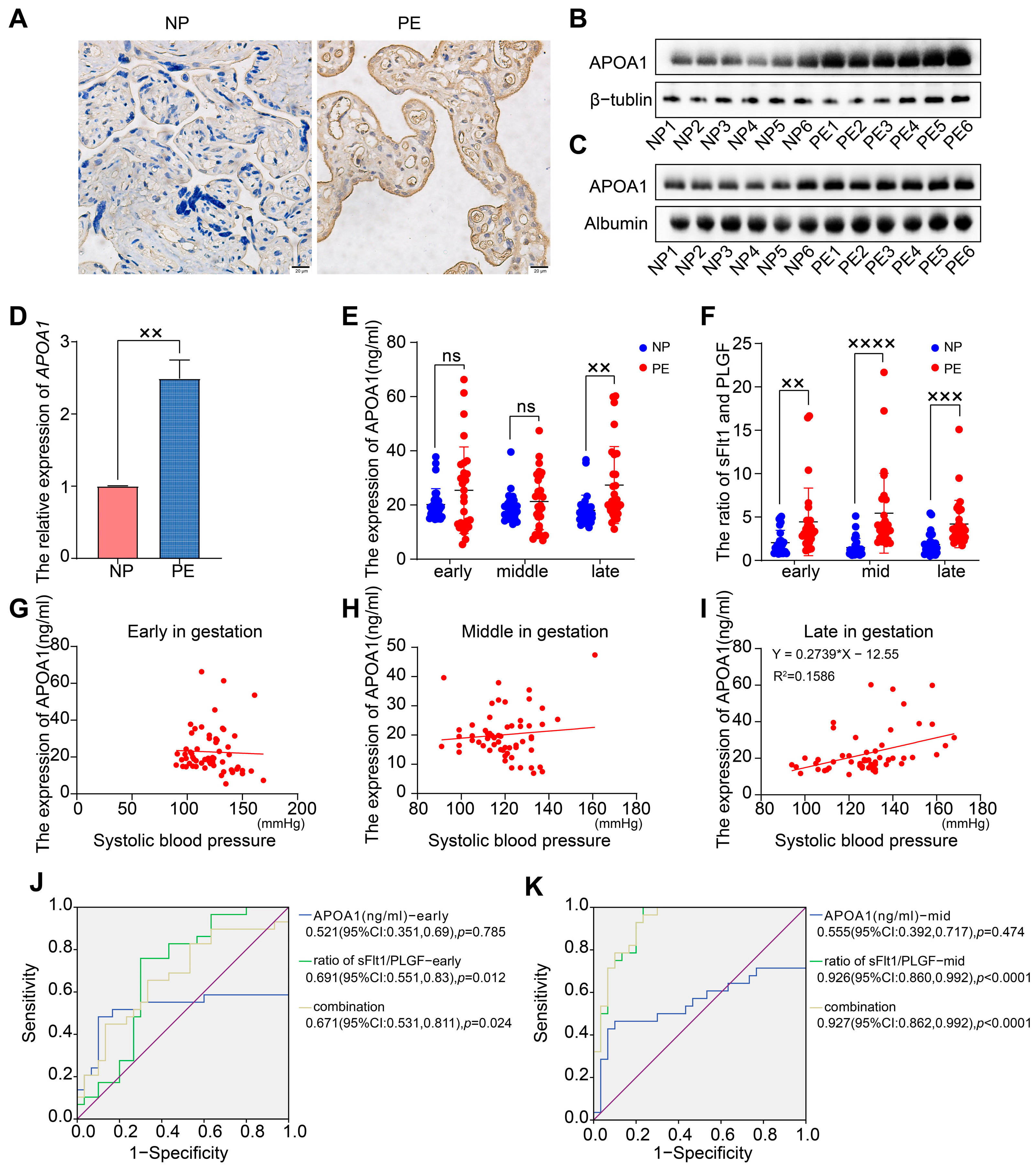
2.1. APOA1 Was Significantly Elevated in the Placental and Plasma Samples from Women with PE
2.2. APOA1 Cannot Predict the Risk of Preeclampsia
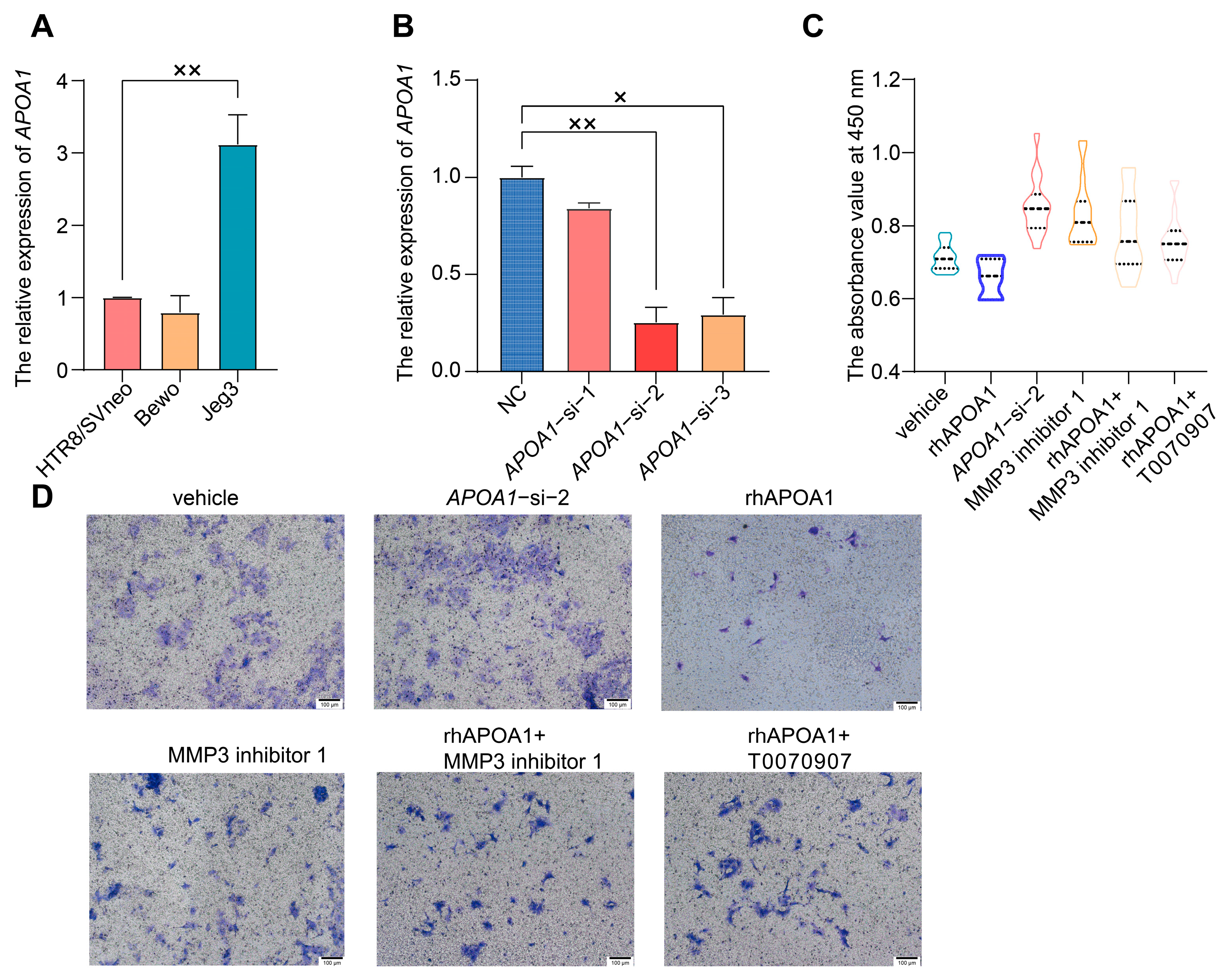
2.3. APOA1 Can Inhibit the Proliferation and Invasion of Trophoblast Cells
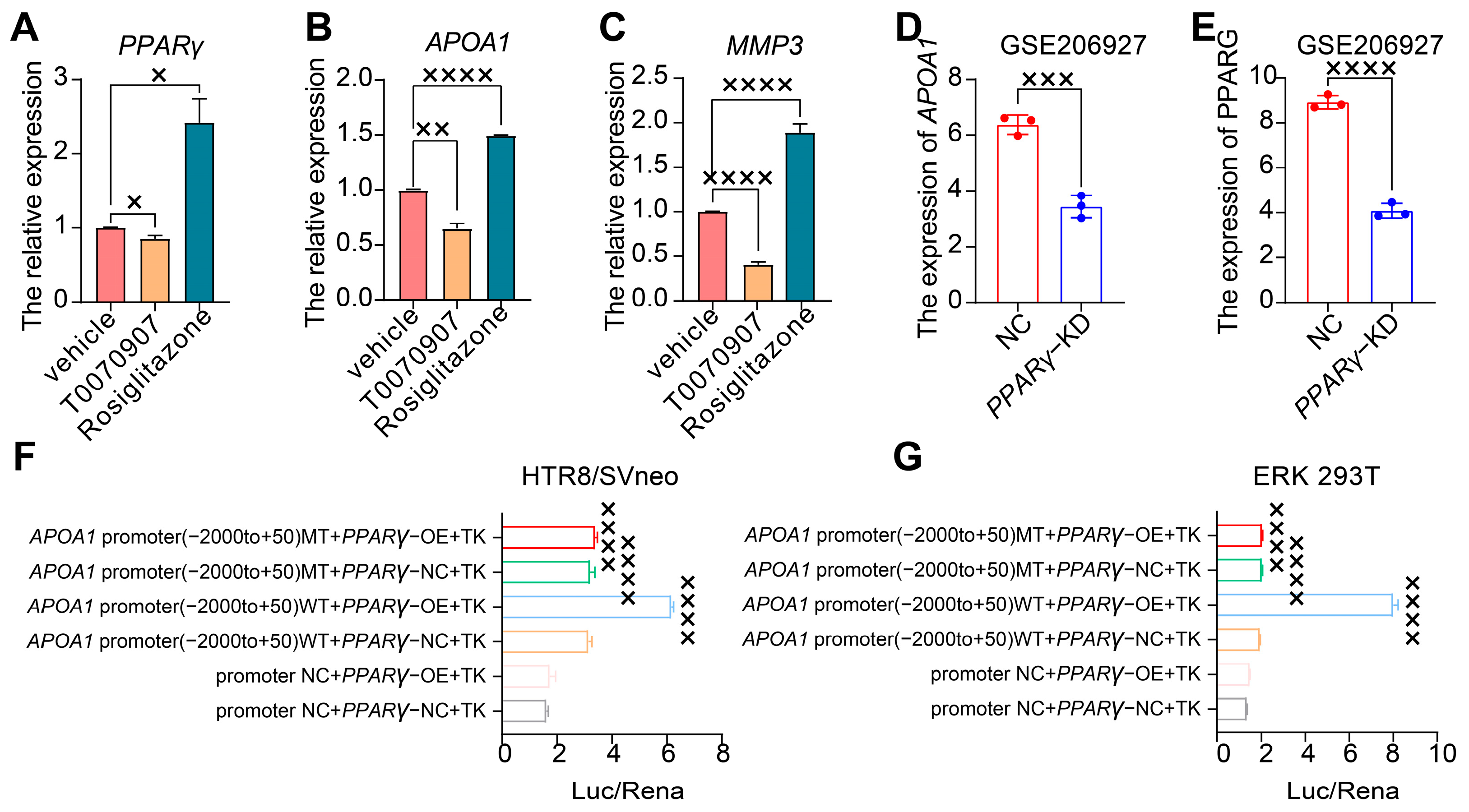
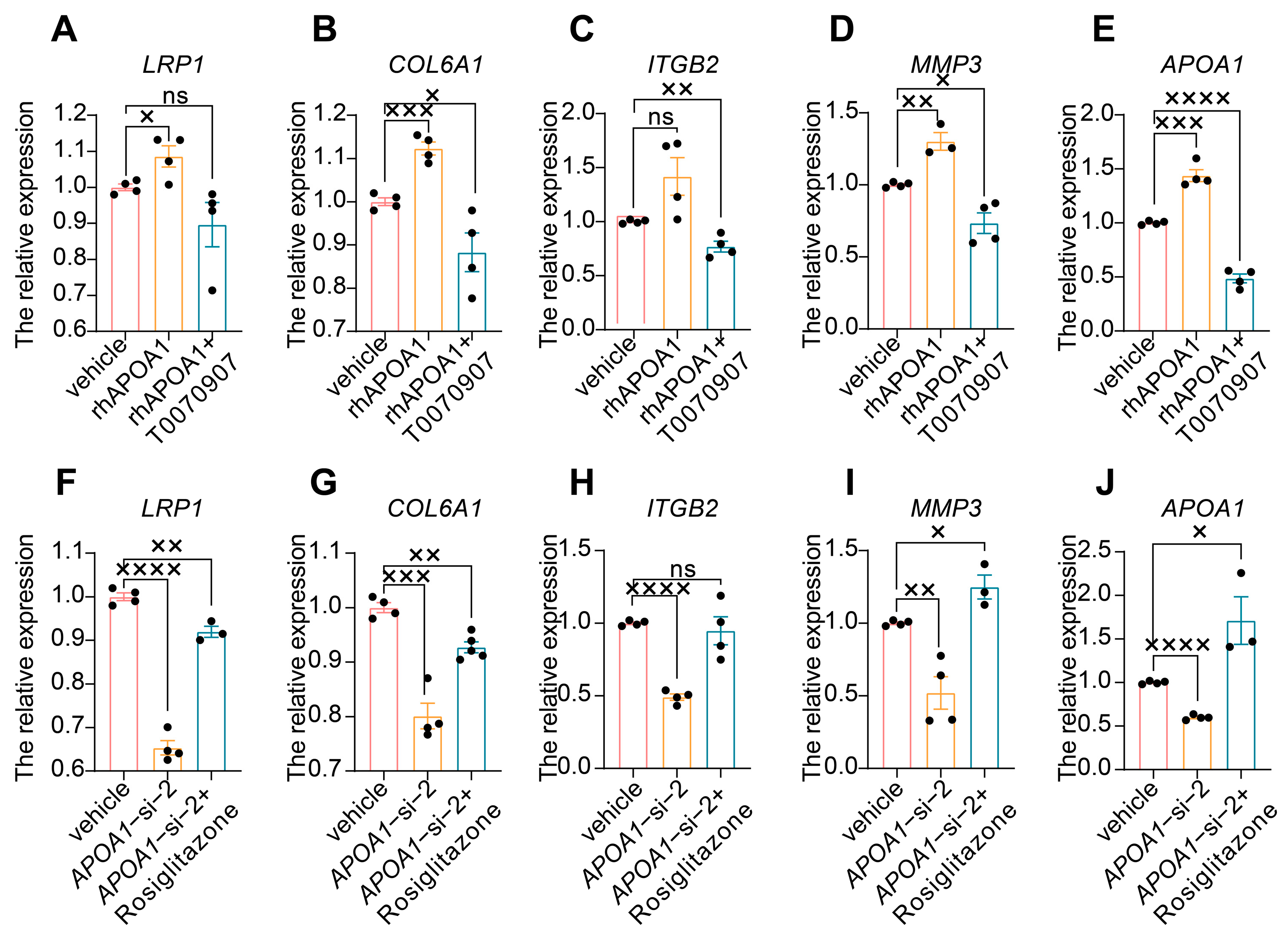
2.4. APOA1 Can Be Transcriptionally Regulated by PPARγ
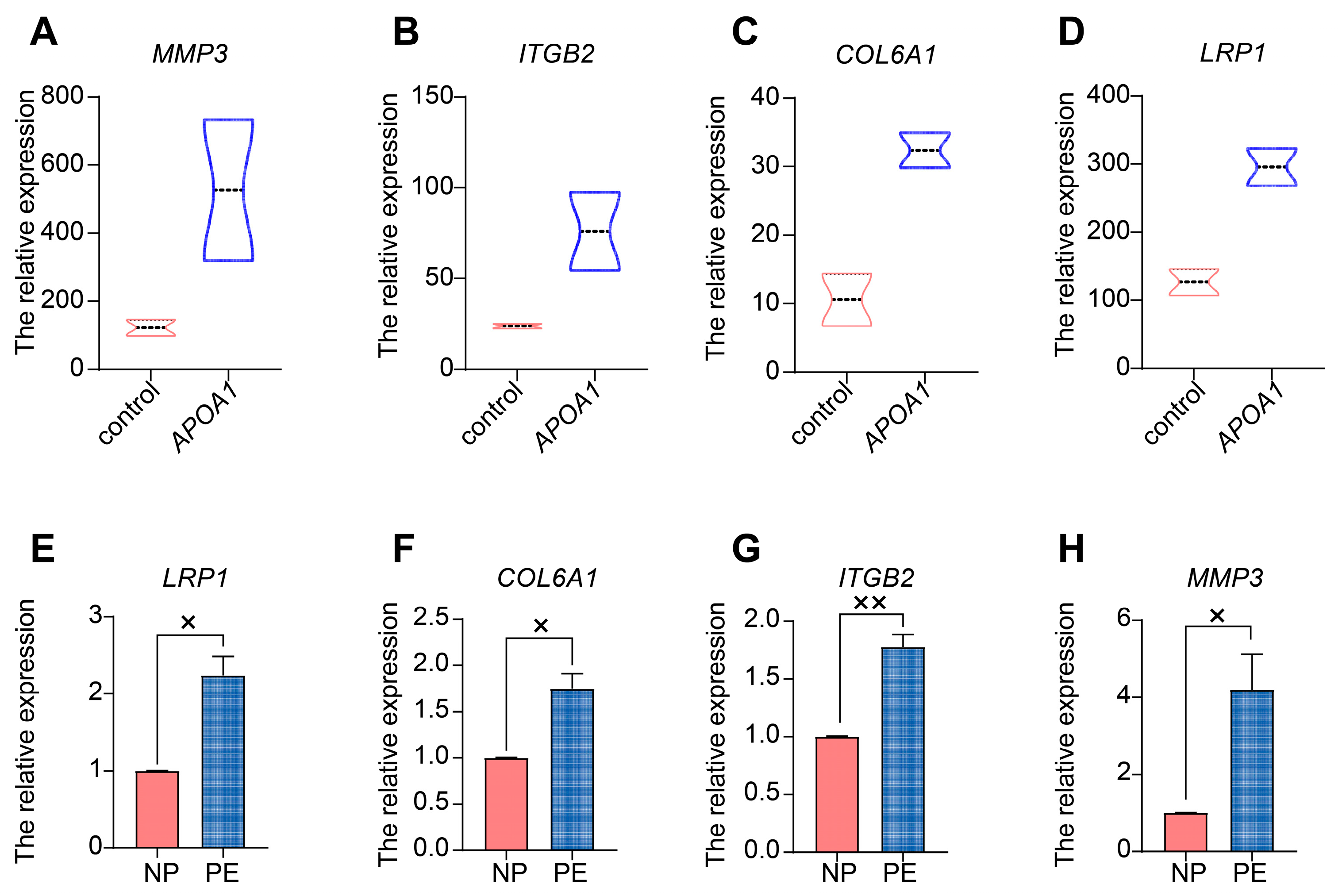
2.5. APOA1 Functions by Regulating Several Downstream Targets
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.1.1. Subjects of the Study
4.1.2. Inclusion and Exclusion Criteria
4.1.3. Samples Collection and Testing
4.2. Expression Profile Datasets Selection and Analysis
4.3. Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
4.4. Cell Experiment
4.4.1. Cell Culture
4.4.2. Small Interfering RNA (siRNA) Transfection
4.4.3. Cell Treatment
4.4.4. Transwell Assay
4.4.5. Cell Proliferation Assay
4.5. Western Blotting
4.6. Immunohistochemistry (IHC)
4.7. Luciferase Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.N.; Oparil, S. Preeclampsia-Pathophysiology and Clinical Presentations: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 76, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, K.R.; Blumenstein, M.; Black, M.A.; Wu, S.H.; Kasabov, N.; Taylor, R.S.; Taylor, R.S.; Cooper, G.J.; North, R.A.; SCOPE Consortium. An altered pattern of circulating apolipoprotein E3 isoforms is implicated in preeclampsia. J. Lipid Res. 2009, 50, 71–80. [Google Scholar] [CrossRef]
- Bendix, E.J.; Ravn, J.D.; Sperling, L.; Overgaard, M. First trimester serum apolipoproteins in the prediction of late-onset preeclampsia. Scand. J. Clin. Lab. Investig. 2023, 83, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Serrano, N.C.; Guio-Mahecha, E.; Quintero-Lesmes, D.C.; Becerra-Bayona, S.; Paez, M.C.; Beltran, M.; Herrera, V.M.; Leon, L.J.; Williams, D.; Casas, J.P. Lipid profile, plasma apolipoproteins, and pre-eclampsia risk in the GenPE case-control study. Atherosclerosis 2018, 276, 189–194. [Google Scholar] [CrossRef]
- Stadler, J.T.; Scharnagl, H.; Wadsack, C.; Marsche, G. Preeclampsia Affects Lipid Metabolism and HDL Function in Mothers and Their Offspring. Antioxidants 2023, 12, 795. [Google Scholar] [CrossRef]
- Ding, X.; Yang, Z.; Han, Y.; Yu, H. Adverse factors increase preeclampsia-like changes in pregnant mice with abnormal lipid metabolism. Chin. Med. J. 2014, 127, 2814–2818. [Google Scholar]
- Ramanjaneya, M.; Butler, A.E.; Bashir, M.; Bettahi, I.; Moin, A.S.M.; Ahmed, L.; Elrayess, M.A.; Hunt, S.C.; Atkin, S.L.; Abou-Samra, A.B. apoA2 correlates to gestational age with decreased apolipoproteins A2, C1, C3 and E in gestational diabetes. BMJ Open Diabetes Res. Care 2021, 9, e001925. [Google Scholar] [CrossRef]
- Mahley, R.W.; Innerarity, T.L.; Rall, S.C., Jr.; Weisgraber, K.H. Plasma lipoproteins: Apolipoprotein structure and function. J. Lipid Res. 1984, 25, 1277–1294. [Google Scholar] [CrossRef]
- Breslow, J.L.; Ross, D.; McPherson, J.; Williams, H.; Kurnit, D.; Nussbaum, A.L.; Karathanasis, S.K.; Zannis, V.I. Isolation and characterization of cDNA clones for human apolipoprotein A-I. Proc. Natl. Acad. Sci. USA 1982, 79, 6861–6865. [Google Scholar] [CrossRef]
- Segrest, J.P.; Li, L.; Anantharamaiah, G.M.; Harvey, S.C.; Liadaki, K.N.; Zannis, V. Structure and function of apolipoprotein A-I and high-density lipoprotein. Curr. Opin. Lipidol. 2000, 11, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Jiang, L. The involvement of long noncoding RNA APOA1-AS in the pathogenesis of preeclampsia. Hum. Exp. Toxicol. 2022, 41, 9603271211066586. [Google Scholar] [CrossRef]
- Charet, J.C.; Watine, J.; Marre, A.; Charet, P. Prognostic value of serum levels of cholesterol and apolipoprotein A1 in pulmonary cancer. Ann. Biol. Clin. 1997, 55, 52. [Google Scholar]
- Clarke, C.H.; Yip, C.; Badgwell, D.; Fung, E.T.; Coombes, K.R.; Zhang, Z.; Lu, K.H.; Bast, R.C., Jr. Proteomic biomarkers apolipoprotein A1, truncated transthyretin and connective tissue activating protein III enhance the sensitivity of CA125 for detecting early stage epithelial ovarian cancer. Gynecol. Oncol. 2011, 122, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.E.; Fung, E.T.; McGuire, M.; Rabkin, C.C.; Molinaro, A.; Wang, Z.; Zhang, F.; Wang, J.; Yip, C.; Meng, X.Y.; et al. Evaluation of apolipoprotein A1 and posttranslationally modified forms of transthyretin as biomarkers for ovarian cancer detection in an independent study population. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1641–1646. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, X.; Wu, F.; Peng, H. Hsa_circ_0088212-mediated miR-520 h/APOA1 axis inhibits osteosarcoma progression. Transl. Oncol. 2021, 14, 101219. [Google Scholar] [CrossRef]
- Verma, P.; Nair, R.R.; Singh, S.; Rajender, S.; Khanna, A.; Jha, R.K.; Singh, K. High Level of APOA1 in Blood and Maternal Fetal Interface Is Associated with Early Miscarriage. Reprod. Sci. 2019, 26, 649–656. [Google Scholar] [CrossRef]
- Graham, C.H.; Hawley, T.S.; Hawley, R.G.; MacDougall, J.R.; Kerbel, R.S.; Khoo, N.; Lala, P.K. Establishment and characterization of first trimester human trophoblast cells with extended lifespan. Exp. Cell Res. 1993, 206, 204–211. [Google Scholar] [CrossRef]
- Lee, S.M.; Moon, J.Y.; Lim, B.Y.; Kim, S.M.; Park, C.W.; Kim, B.J.; Jun, J.K.; Norwitz, E.R.; Choi, M.H.; Park, J.S. Increased biosynthesis and accumulation of cholesterol in maternal plasma, but not amniotic fluid in pre-eclampsia. Sci. Rep. 2019, 9, 1550. [Google Scholar] [CrossRef]
- Anand, S.; Young, S.; Esplin, M.S.; Peaden, B.; Tolley, H.D.; Porter, T.F.; Varner, M.W.; D’Alton, M.E.; Jackson, B.J.; Graves, S.W. Detection and confirmation of serum lipid biomarkers for preeclampsia using direct infusion mass spectrometry. J. Lipid Res. 2016, 57, 687–696. [Google Scholar] [CrossRef]
- Timur, H.; Daglar, H.K.; Kara, O.; Kirbas, A.; Inal, H.A.; Turkmen, G.G.; Yilmaz, Z.; Elmas, B.; Uygur, D. A study of serum Apo A-1 and Apo B-100 levels in women with preeclampsia. Pregnancy Hypertens. 2016, 6, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xiao, Y.; Tang, L.; Jiang, L.; Wang, Y.; Zhang, R.; Wei, Q.; Lu, Y. Apolipoprotein A1 −75 G/A and +83 C/T polymorphisms and renal cancer risk. Lipids Health Dis. 2015, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, M. Altered Maternal Serum Matrix Metalloproteinases MMP-2, MMP-3, MMP-9, and MMP-13 in Severe Early- and Late-Onset Preeclampsia. Biomed. Res. Int. 2017, 2017, 6432426. [Google Scholar] [CrossRef]
- Shoaito, H.; Chauveau, S.; Gosseaume, C.; Bourguet, W.; Vigouroux, C.; Vatier, C.; Pienkowski, C.; Fournier, T.; Degrelle, S.A. Peroxisome proliferator-activated receptor gamma-ligand-binding domain mutations associated with familial partial lipodystrophy type 3 disrupt human trophoblast fusion and fibroblast migration. J. Cell Mol. Med. 2020, 24, 7660–7669. [Google Scholar] [CrossRef] [PubMed]
- Fruchart, J.C.; Staels, B.; Duriez, P. PPARS, metabolic disease and atherosclerosis. Pharmacol. Res. 2001, 44, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstet. Gynecol. 2020, 135, e237–e260. [CrossRef] [PubMed]
- Bosco, C.B.; Diaz, E.G.; Gutierrez, R.R.; Gonzalez, J.M.; Parra-Cordero, M.; Rodrigo, R.S.; Barja, P.Y. Placental Hypoxia Developed During Preeclampsia Induces Telocytes Apoptosis in Chorionic Villi Affecting The Maternal-Fetus Metabolic Exchange. Curr. Stem Cell Res. Ther. 2016, 11, 420–425. [Google Scholar] [CrossRef]
- Ji, J.; Xue, T.F.; Guo, X.D.; Yang, J.; Guo, R.B.; Wang, J.; Huang, J.Y.; Zhao, X.J.; Sun, X.L. Antagonizing peroxisome proliferator-activated receptor gamma facilitates M1-to-M2 shift of microglia by enhancing autophagy via the LKB1-AMPK signaling pathway. Aging Cell 2018, 17, e12774. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Normal Pregnancy (n = 30) | Preeclampsia (n = 30) | p-Value |
|---|---|---|---|
| Maternal age (years) | 36.55 ± 2.587 | 37.07 ± 2.433 | 0.3435 |
| Gestational age (weeks) | 38.69 ± 1.161 | 36.25 ± 3.152 | <0.0001 **** |
| Pregnancy BMI (kg/m2) | 21.66 ± 2.880 | 24.53 ± 3.920 | 0.0003 *** |
| Systolic BP (mmHg) | 110.1 ± 10.46 | 130.8 ± 12.91 | <0.0001 **** |
| Diastolic BP (mmHg) | 63.38 ± 8.999 | 81.12 ± 10.83 | <0.0001 **** |
| Proteinuria (g/24 h) | - | 0.8990 ± 1.433 | - |
| Neonatal weight (g) | 3500 ± 461.3 | 2785 ± 703.5 | <0.0001 **** |
| Gene | Primers | Sequence (5′-3′) |
|---|---|---|
| GAPDH | FORWARD | TTCGACAGTCAGCCGCATCTT |
| REVERSE | CCCAATACGACCAAATCCGTT | |
| APOA1 | FORWARD | CTAAAGCTCCTTGACAACTGGG |
| REVERSE | TTTCCAGGTTATCCCAGAACTC | |
| PPARγ | FORWARD | AGATCATTTACACAATGCTGGC |
| REVERSE | TAAAGTCACCAAAAGGCTTTCG | |
| LRP1 | FORWARD | AGTCTGCTTCGTGTGCCTATCC |
| REVERSE | AGTCATTGTCATTGTCGCATCTCC | |
| COL6A1 | FORWARD | AGCACCTGGGCGTCAAAGTC |
| REVERSE | TGTGGTCCGTGGCGATGATG | |
| ITGB2 | FORWARD | GGAGCAGCAGGACGGGATG |
| REVERSE | GACGATGGCGGCGATGTTG | |
| MMP3 | FORWARD | CTTTCCTGGCATCCCGAAGTG |
| REVERSE | CTCAACAGCAGAATCAACAGCATC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Pei, J.; Zhang, X.; Wang, C.; Tang, Y.; Liu, H.; Yu, Y.; Luo, S.; Gu, W. APOA1 Is a Novel Marker for Preeclampsia. Int. J. Mol. Sci. 2023, 24, 16363. https://doi.org/10.3390/ijms242216363
Liu Z, Pei J, Zhang X, Wang C, Tang Y, Liu H, Yu Y, Luo S, Gu W. APOA1 Is a Novel Marker for Preeclampsia. International Journal of Molecular Sciences. 2023; 24(22):16363. https://doi.org/10.3390/ijms242216363
Chicago/Turabian StyleLiu, Zhenzhen, Jiangnan Pei, Xiaoyue Zhang, Chengjie Wang, Yao Tang, Haiyan Liu, Yi Yu, Shouling Luo, and Weirong Gu. 2023. "APOA1 Is a Novel Marker for Preeclampsia" International Journal of Molecular Sciences 24, no. 22: 16363. https://doi.org/10.3390/ijms242216363
APA StyleLiu, Z., Pei, J., Zhang, X., Wang, C., Tang, Y., Liu, H., Yu, Y., Luo, S., & Gu, W. (2023). APOA1 Is a Novel Marker for Preeclampsia. International Journal of Molecular Sciences, 24(22), 16363. https://doi.org/10.3390/ijms242216363