
The Role of TGF-β Signaling in Saphenous Vein Graft Failure after Peripheral Arterial Disease Bypass Surgery

Abstract
1. Introduction
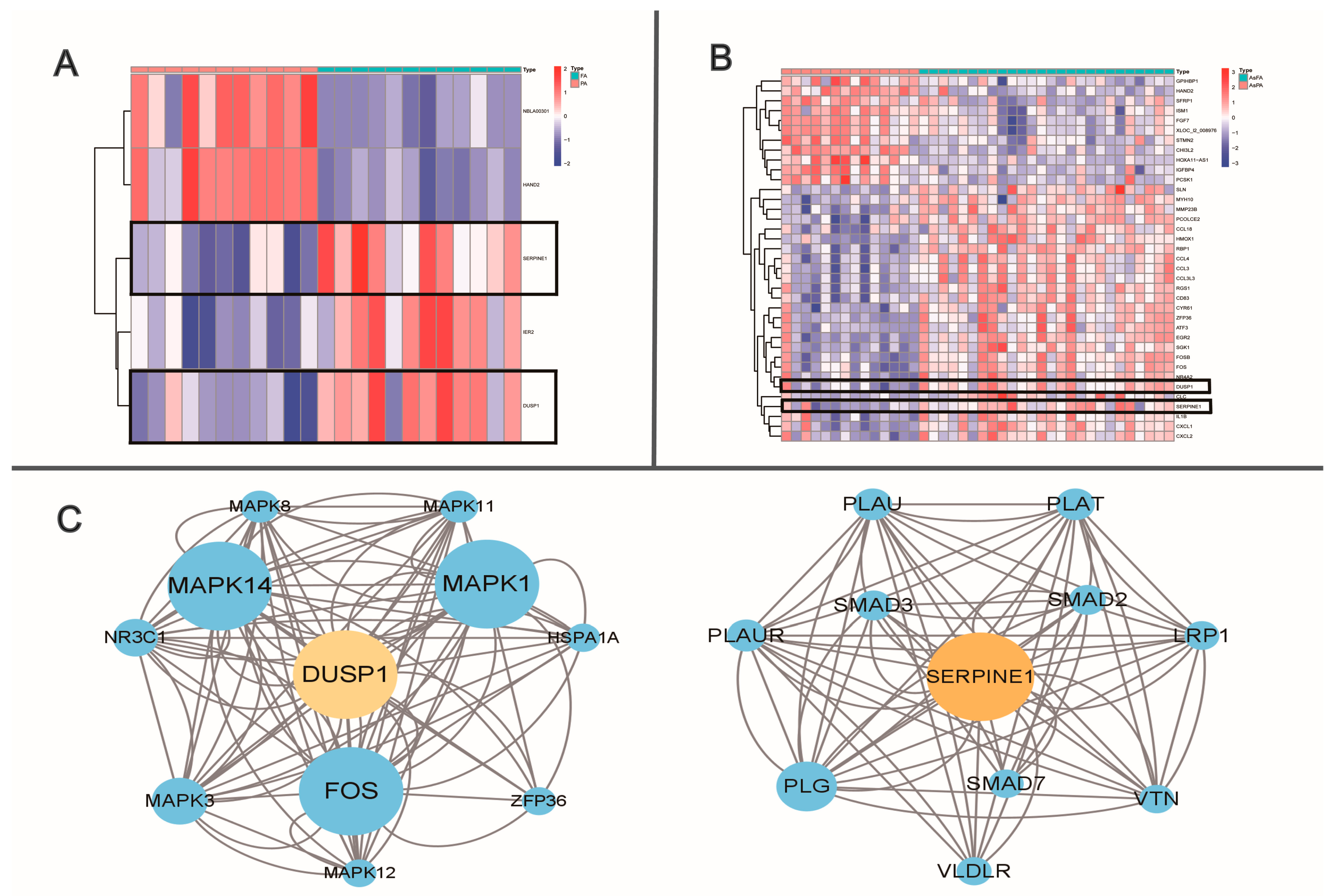
2. Identification and Analysis of Special Pathways
3. TGF-β Family and Signaling Pathway
4. TGF-β Signaling Mediates Endothelial–Mesenchymal Transition (EMT) during Vein Graft Remodeling
5. Extracellular Matrix
6. Fibrosis
7. TGF-β Signaling and Intimal Hyperplasia
8. Risk Factors of Bypass Graft Failure
9. Case Presentation
10. Clinical Treatment Strategies
11. Prospects
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stevens, L.M.; Lymn, C.; Glass, R.M. Peripheral Arterial Disease. JAMA 2006, 295, 584. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Rudan, D.; Wang, M.; Chang, X.; Rudan, I. National and subnational estimation of the prevalence of peripheral artery disease (PAD) in China: A systematic review and meta-analysis. J. Glob. Health 2019, 9, 010601. [Google Scholar] [CrossRef]
- Gerhard-Herman, M.D.; Gornik, H.L.; Barrett, C.; Barshes, N.R.; Corriere, M.A.; Drachman, D.E.; Fleisher, L.A.; Fowkes, F.G.; Hamburg, N.M.; Kinlay, S.; et al. 2016 AHA/ACC Guideline on the Management of Patients With Lower Extremity Peripheral Artery Disease: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2017, 135, e686–e725. [Google Scholar] [CrossRef] [PubMed]
- Allaqaband, S.; Solis, J.; Kazemi, S.; Bajwa, T. Endovascular treatment of peripheral vascular disease. Curr. Probl. Cardiol. 2006, 31, 711–760. [Google Scholar] [CrossRef] [PubMed]
- Owens, C.D. Adaptive changes in autogenous vein grafts for arterial reconstruction: Clinical implications. J. Vasc. Surg. 2010, 51, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.; Zadina, K.; Moritz, T.; Ovitt, T.; Sethi, G.; Copeland, J.G.; Thottapurathu, L.; Krasnicka, B.; Ellis, N.; Anderson, R.J.; et al. Long-term patency of saphenous vein and left internal mammary artery grafts after coronary artery bypass surgery: Results from a Department of Veterans Affairs Cooperative Study. J. Am. Coll. Cardiol. 2004, 44, 2149–2156. [Google Scholar] [CrossRef]
- de Vries, M.R.; Simons, K.H.; Jukema, J.W.; Braun, J.; Quax, P.H. Vein graft failure: From pathophysiology to clinical outcomes. Nat. Rev. Cardiol. 2016, 13, 451–470. [Google Scholar] [CrossRef]
- Goteri, G.; Altobelli, E.; Tossetta, G.; Zizzi, A.; Avellini, C.; Licini, C.; Lorenzi, T.; Castellucci, M.; Ciavattini, A.; Marzioni, D. High temperature requirement A1, transforming growth factor beta1, phosphoSmad2 and Ki67 in eutopic and ectopic endometrium of women with endometriosis. Eur. J. Histochem. EJH 2015, 59, 2570. [Google Scholar] [CrossRef]
- Nikol, S.; Weir, L.; Sullivan, A.; Sharaf, B.; White, C.J.; Zemel, G.; Hartzler, G.; Stack, R.; Leclerc, G.; Isner, J.M. Persistently increased expression of the transforming growth factor-β1 gene in human vascular restenosis: Analysis of 62 patients with one or more episode of restenosis. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 1994, 3, 57–64. [Google Scholar] [CrossRef]
- Friedl, R.; Li, J.; Schumacher, B.; Hanke, H.; Waltenberger, J.; Hannekum, A.; Stracke, S. Intimal hyperplasia and expression of transforming growth factor-beta1 in saphenous veins and internal mammary arteries before coronary artery surgery. Ann. Thorac. Surg. 2004, 78, 1312–1318. [Google Scholar] [CrossRef]
- Dalager, S.; Paaske, W.P.; Kristensen, I.B.; Laurberg, J.M.; Falk, E. Artery-related differences in atherosclerosis expression: Implications for atherogenesis and dynamics in intima-media thickness. Stroke 2007, 38, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Marinelli Busilacchi, E.; Costantini, A.; Mancini, G.; Tossetta, G.; Olivieri, J.; Poloni, A.; Viola, N.; Butini, L.; Campanati, A.; Goteri, G.; et al. Nilotinib Treatment of Patients Affected by Chronic Graft-versus-Host Disease Reduces Collagen Production and Skin Fibrosis by Downmodulating the TGF-β and p-SMAD Pathway. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2020, 26, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Nigro, P.; Abe, J.; Berk, B.C. Flow shear stress and atherosclerosis: A matter of site specificity. Antioxid. Redox Signal. 2011, 15, 1405–1414. [Google Scholar] [CrossRef]
- Liao, J.; Liu, Q.; Chen, J.; Lu, Z.; Mo, H.; Jia, J. A risk score model based on TGF-β pathway-related genes predicts survival, tumor microenvironment and immunotherapy for liver hepatocellular carcinoma. Proteome Sci. 2022, 20, 11. [Google Scholar] [CrossRef]
- Agarwal, I.; Arnold, A.; Glazer, N.L.; Barasch, E.; Djousse, L.; Fitzpatrick, A.L.; Gottdiener, J.S.; Ix, J.H.; Jensen, R.A.; Kizer, J.R.; et al. Fibrosis-related biomarkers and large and small vessel disease: The Cardiovascular Health Study. Atherosclerosis 2015, 239, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, Y.; Liang, C.; Li, J.; Li, Y.; Wu, Q.; Liu, Z.; Pang, X.; Chang, Y.X. Buyang Huanwu Decoction ameliorates atherosclerosis by regulating TGF-β/Smad2 pathway to promote the differentiation of regulatory T cells. J. Ethnopharmacol. 2021, 269, 113724. [Google Scholar] [CrossRef]
- Schuppan, D.; Ashfaq-Khan, M.; Yang, A.T.; Kim, Y.O. Liver fibrosis: Direct antifibrotic agents and targeted therapies. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 68–69, 435–451. [Google Scholar] [CrossRef]
- Zhao, M.; Mishra, L.; Deng, C.X. The role of TGF-β/SMAD4 signaling in cancer. Int. J. Biol. Sci. 2018, 14, 111–123. [Google Scholar] [CrossRef]
- Lodyga, M.; Hinz, B. TGF-β1—A truly transforming growth factor in fibrosis and immunity. Semin. Cell Dev. Biol. 2020, 101, 123–139. [Google Scholar] [CrossRef]
- Takahashi, H.; Alves, C.R.R.; Stanford, K.I.; Middelbeek, R.J.W.; Nigro, P.; Ryan, R.E.; Xue, R.; Sakaguchi, M.; Lynes, M.D.; So, K.; et al. TGF-β2 is an exercise-induced adipokine that regulates glucose and fatty acid metabolism. Nat. Metab. 2019, 1, 291–303. [Google Scholar] [CrossRef]
- Komai, T.; Okamura, T.; Inoue, M.; Yamamoto, K.; Fujio, K. Reevaluation of Pluripotent Cytokine TGF-β3 in Immunity. Int. J. Mol. Sci. 2018, 19, 2261. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Tzavlaki, K.; Moustakas, A. TGF-β Signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Humeres, C.; Venugopal, H.; Frangogiannis, N.G. Smad-dependent pathways in the infarcted and failing heart. Curr. Opin. Pharmacol. 2022, 64, 102207. [Google Scholar] [CrossRef]
- Pascual, G.; Mendieta, C.; García-Honduvilla, N.; Corrales, C.; Bellón, J.M.; Buján, J. TGF-beta1 upregulation in the aging varicose vein. J. Vasc. Res. 2007, 44, 192–201. [Google Scholar] [CrossRef]
- Wang, J.C.; Gu, J.; Li, Y.; Ma, Q.; Feng, J.; Lu, S. Transforming growth factor-β1 and inducible nitric oxide synthase signaling were involved in effects of prostaglandin E2 on progression of lower limb varicose veins. J. Vasc. Surg. Venous Lymphat. Disord. 2021, 9, 1535–1544. [Google Scholar] [CrossRef]
- Buján, J.; Gimeno, M.J.; Jiménez, J.A.; Kielty, C.M.; Mecham, R.P.; Bellón, J.M. Expression of elastic components in healthy and varicose veins. World J. Surg. 2003, 27, 901–905. [Google Scholar] [CrossRef]
- Jacob, T.; Hingorani, A.; Ascher, E. Overexpression of transforming growth factor-beta1 correlates with increased synthesis of nitric oxide synthase in varicose veins. J. Vasc. Surg. 2005, 41, 523–530. [Google Scholar] [CrossRef]
- Kowalewski, R.; Malkowski, A.; Sobolewski, K.; Gacko, M. Evaluation of transforming growth factor-beta signaling pathway in the wall of normal and varicose veins. Pathobiology 2010, 77, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Serralheiro, P.; Cairrão, E.; Maia, C.J.; João, M.; Almeida, C.M.C.; Verde, I. Effect of TGF-beta1 on MMP/TIMP and TGF-beta1 receptors in great saphenous veins and its significance on chronic venous insufficiency. Phlebology 2017, 32, 334–341. [Google Scholar] [CrossRef] [PubMed]
- George, S.J.; Johnson, J.L.; Smith, M.A.; Angelini, G.D.; Jackson, C.L. Transforming growth factor-beta is activated by plasmin and inhibits smooth muscle cell death in human saphenous vein. J. Vasc. Res. 2005, 42, 247–254. [Google Scholar] [CrossRef]
- Serralheiro, P.; Novais, A.; Cairrão, E.; Maia, C.; Costa Almeida, C.M.; Verde, I. Variability of MMP/TIMP and TGF-β1 Receptors throughout the Clinical Progression of Chronic Venous Disease. Int. J. Mol. Sci. 2017, 19, 6. [Google Scholar] [CrossRef]
- Mii, S.; Ware, J.A.; Kent, K.C. Transforming growth factor-beta inhibits human vascular smooth muscle cell growth and migration. Surgery 1993, 114, 464–470. [Google Scholar] [PubMed]
- Jackson, A.O.; Zhang, J.; Jiang, Z.; Yin, K. Endothelial-to-mesenchymal transition: A novel therapeutic target for cardiovascular diseases. Trends Cardiovasc. Med. 2017, 27, 383–393. [Google Scholar] [CrossRef]
- Cai, C.; Zhao, C.; Kilari, S.; Sharma, A.; Singh, A.K.; Simeon, M.L.; Misra, A.; Li, Y.; Misra, S. Effect of sex differences in treatment response to angioplasty in a murine arteriovenous fistula model. Am. J. Physiol. Ren. Physiol. 2020, 318, F565–F575. [Google Scholar] [CrossRef]
- Moonen, J.R.; Lee, E.S.; Schmidt, M.; Maleszewska, M.; Koerts, J.A.; Brouwer, L.A.; van Kooten, T.G.; van Luyn, M.J.; Zeebregts, C.J.; Krenning, G.; et al. Endothelial-to-mesenchymal transition contributes to fibro-proliferative vascular disease and is modulated by fluid shear stress. Cardiovasc. Res. 2015, 108, 377–386. [Google Scholar] [CrossRef]
- Vanchin, B.; Offringa, E.; Friedrich, J.; Brinker, M.G.; Kiers, B.; Pereira, A.C.; Harmsen, M.C.; Moonen, J.A.; Krenning, G. MicroRNA-374b induces endothelial-to-mesenchymal transition and early lesion formation through the inhibition of MAPK7 signaling. J. Pathol. 2019, 247, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.M.; Li, S.; Wang, X.W.; Chen, D.; Jiang, Z.L.; Zhang, C.; He, X.J.; Huang, C.; Jiang, Y.J.; Wu, Q.C. MicroRNA-92a -mediated endothelial to mesenchymal transition controls vein graft neointimal lesion formation. Exp. Cell Res. 2021, 398, 112402. [Google Scholar] [CrossRef]
- Hong, L.; Du, X.; You, T.; Sun, L.; Li, W.; Xiao, L.; Lu, H.; Wang, W.; Li, X. Reciprocal enhancement of thrombosis by endothelial-to-mesenchymal transition induced by iliac vein compression. Life Sci. 2019, 233, 116659. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Fu, Y.Y.; Zhu, J.; Wang, L.; Aafaqi, S.; Rahkonen, O.; Slorach, C.; Traister, A.; Leung, C.H.; Chiasson, D.; et al. Pulmonary vein stenosis and the pathophysiology of “upstream” pulmonary veins. J. Thorac. Cardiovasc. Surg. 2014, 148, 245–253. [Google Scholar] [CrossRef]
- Waller, J.R.; Nicholson, M.L. Molecular mechanisms of renal allograft fibrosis. Br. J. Surg. 2001, 88, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Waller, J.R.; Brook, N.R.; Bicknell, G.R.; Nicholson, M.L. Differential effects of modern immunosuppressive agents on the development of intimal hyperplasia. Transpl. Int. Off. J. Eur. Soc. Organ Transplant. 2004, 17, 9–14. [Google Scholar] [CrossRef]
- Cai, C.; Kilari, S.; Singh, A.K.; Zhao, C.; Simeon, M.L.; Misra, A.; Li, Y.; Misra, S. Differences in Transforming Growth Factor-β1/BMP7 Signaling and Venous Fibrosis Contribute to Female Sex Differences in Arteriovenous Fistulas. J. Am. Heart Assoc. 2020, 9, e017420. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Barron, A.O.; Gindina, S.; Kumar, S.; Chintala, S.; Nayyar, A.; Danias, J. Investigations on the Role of the Fibrinolytic Pathway on Outflow Facility Regulation. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1571–1580. [Google Scholar] [CrossRef]
- Logan, R.; Jeffers, A.; Qin, W.; Owens, S.; Chauhan, P.; Komatsu, S.; Ikebe, M.; Idell, S.; Tucker, T.A. TGF-β regulation of the uPA/uPAR axis modulates mesothelial-mesenchymal transition (MesoMT). Sci. Rep. 2021, 11, 21210. [Google Scholar] [CrossRef] [PubMed]
- Plekhanova, O.S.; Parfyonova, Y.V.; Bibilashvily, R.; Stepanova, V.V.; Erne, P.; Bobik, A.; Tkachuk, V.A. Urokinase plasminogen activator enhances neointima growth and reduces lumen size in injured carotid arteries. J. Hypertens. 2000, 18, 1065–1069. [Google Scholar] [CrossRef]
- Guzzoni, V.; Marqueti, R.C.; Durigan, J.L.Q.; Faustino de Carvalho, H.; Lino, R.L.B.; Mekaro, M.S.; Costa Santos, T.O.; Mecawi, A.S.; Rodrigues, J.A.; Hord, J.M.; et al. Reduced collagen accumulation and augmented MMP-2 activity in left ventricle of old rats submitted to high-intensity resistance training. J. Appl. Physiol. 2017, 123, 655–663. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, X.; Chen, W.; Wang, Z.; Xu, G.; Zeng, Y.; Ma, Y. TGF-β1 prevents blood-brain barrier damage and hemorrhagic transformation after thrombolysis in rats. Exp. Neurol. 2015, 266, 120–126. [Google Scholar] [CrossRef]
- Guimarães, D.A.; Rizzi, E.; Ceron, C.S.; Martins-Oliveira, A.; Gerlach, R.F.; Shiva, S.; Tanus-Santos, J.E. Atorvastatin and sildenafil decrease vascular TGF-β levels and MMP-2 activity and ameliorate arterial remodeling in a model of renovascular hypertension. Redox Biol. 2015, 6, 386–395. [Google Scholar] [CrossRef]
- Müller, K.M.; Dasbach, G. The pathology of vascular grafts. Curr. Top. Pathol. Ergeb. Pathol. 1994, 86, 273–306. [Google Scholar] [CrossRef]
- Sun, D.X.; Liu, Z.; Tan, X.D.; Cui, D.X.; Wang, B.S.; Dai, X.W. Nanoparticle-mediated local delivery of an antisense TGF-β1 construct inhibits intimal hyperplasia in autogenous vein grafts in rats. PLoS ONE 2012, 7, e41857. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.H.; Huang, X.Q.; Tan, H.M. Vascular fibrosis in atherosclerosis. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2013, 22, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Ostriker, A.C.; Jin, Y.; Hu, H.; Sizer, A.J.; Peng, G.; Morris, A.H.; Ryu, C.; Herzog, E.L.; Kyriakides, T.; et al. LMO7 Is a Negative Feedback Regulator of Transforming Growth Factor β Signaling and Fibrosis. Circulation 2019, 139, 679–693. [Google Scholar] [CrossRef]
- Latko, M.; Czyrek, A.; Porębska, N.; Kucińska, M.; Otlewski, J.; Zakrzewska, M.; Opaliński, Ł. Cross-Talk between Fibroblast Growth Factor Receptors and Other Cell Surface Proteins. Cells 2019, 8, 455. [Google Scholar] [CrossRef]
- Chen, M.; Bao, L.; Zhao, M.; Cao, J.; Zheng, H. Progress in Research on the Role of FGF in the Formation and Treatment of Corneal Neovascularization. Front. Pharmacol. 2020, 11, 111. [Google Scholar] [CrossRef]
- Tran, P.K.; Tran-Lundmark, K.; Soininen, R.; Tryggvason, K.; Thyberg, J.; Hedin, U. Increased intimal hyperplasia and smooth muscle cell proliferation in transgenic mice with heparan sulfate-deficient perlecan. Circ. Res. 2004, 94, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Nabel, E.G.; Yang, Z.Y.; Plautz, G.; Forough, R.; Zhan, X.; Haudenschild, C.C.; Maciag, T.; Nabel, G.J. Recombinant fibroblast growth factor-1 promotes intimal hyperplasia and angiogenesis in arteries in vivo. Nature 1993, 362, 844–846. [Google Scholar] [CrossRef] [PubMed]
- Edelman, E.R.; Nugent, M.A.; Smith, L.T.; Karnovsky, M.J. Basic fibroblast growth factor enhances the coupling of intimal hyperplasia and proliferation of vasa vasorum in injured rat arteries. J. Clin. Investig. 1992, 89, 465–473. [Google Scholar] [CrossRef]
- Neschis, D.G.; Safford, S.D.; Hanna, A.K.; Fox, J.C.; Golden, M.A. Antisense basic fibroblast growth factor gene transfer reduces early intimal thickening in a rabbit femoral artery balloon injury model. J. Vasc. Surg. 1998, 27, 126–134. [Google Scholar] [CrossRef]
- Zhao, X.M.; Frist, W.H.; Yeoh, T.K.; Miller, G.G. Modification of alternative messenger RNA splicing of fibroblast growth factor receptors in human cardiac allografts during rejection. J. Clin. Investig. 1994, 94, 992–1003. [Google Scholar] [CrossRef]
- Akatsu, Y.; Takahashi, N.; Yoshimatsu, Y.; Kimuro, S.; Muramatsu, T.; Katsura, A.; Maishi, N.; Suzuki, H.I.; Inazawa, J.; Hida, K.; et al. Fibroblast growth factor signals regulate transforming growth factor-β-induced endothelial-to-myofibroblast transition of tumor endothelial cells via Elk1. Mol. Oncol. 2019, 13, 1706–1724. [Google Scholar] [CrossRef]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into TGF-β/Smad signaling in tissue fibrosis. Chem.-Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef]
- Ma, Z.G.; Yuan, Y.P.; Wu, H.M.; Zhang, X.; Tang, Q.Z. Cardiac fibrosis: New insights into the pathogenesis. Int. J. Biol. Sci. 2018, 14, 1645–1657. [Google Scholar] [CrossRef]
- Smith, J.D.; Bryant, S.R.; Couper, L.L.; Vary, C.P.; Gotwals, P.J.; Koteliansky, V.E.; Lindner, V. Soluble transforming growth factor-beta type II receptor inhibits negative remodeling, fibroblast transdifferentiation, and intimal lesion formation but not endothelial growth. Circ. Res. 1999, 84, 1212–1222. [Google Scholar] [CrossRef]
- Wang, X.M.; Liu, X.M.; Wang, Y.; Chen, Z.Y. Activating transcription factor 3 (ATF3) regulates cell growth, apoptosis, invasion and collagen synthesis in keloid fibroblast through transforming growth factor beta (TGF-beta)/SMAD signaling pathway. Bioengineered 2021, 12, 117–126. [Google Scholar] [CrossRef]
- Kramer, E.L.; Clancy, J.P. TGFβ as a therapeutic target in cystic fibrosis. Expert Opin. Ther. Targets 2018, 22, 177–189. [Google Scholar] [CrossRef]
- Li, M.; Krishnaveni, M.S.; Li, C.; Zhou, B.; Xing, Y.; Banfalvi, A.; Li, A.; Lombardi, V.; Akbari, O.; Borok, Z.; et al. Epithelium-specific deletion of TGF-β receptor type II protects mice from bleomycin-induced pulmonary fibrosis. J. Clin. Investig. 2011, 121, 277–287. [Google Scholar] [CrossRef]
- Baganha, F.; de Jong, A.; Jukema, J.W.; Quax, P.H.A.; de Vries, M.R. The Role of Immunomodulation in Vein Graft Remodeling and Failure. J. Cardiovasc. Transl. Res. 2021, 14, 100–109. [Google Scholar] [CrossRef]
- Davies, M.G.; Hagen, P.O. Pathophysiology of vein graft failure: A review. Eur. J. Vasc. Endovasc. Surg. Off. J. Eur. Soc. Vasc. Surg. 1995, 9, 7–18. [Google Scholar] [CrossRef]
- Zeng, Z.; Xia, L.; Fan, X.; Ostriker, A.C.; Yarovinsky, T.; Su, M.; Zhang, Y.; Peng, X.; Xie, Y.; Pi, L.; et al. Platelet-derived miR-223 promotes a phenotypic switch in arterial injury repair. J. Clin. Investig. 2019, 129, 1372–1386. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.O.; Caputo, M.; Angelini, G.D.; George, S.J.; Zakkar, M. Activation and inflammation of the venous endothelium in vein graft disease. Atherosclerosis 2017, 265, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Xenogiannis, I.; Zenati, M.; Bhatt, D.L.; Rao, S.V.; Rodés-Cabau, J.; Goldman, S.; Shunk, K.A.; Mavromatis, K.; Banerjee, S.; Alaswad, K.; et al. Saphenous Vein Graft Failure: From Pathophysiology to Prevention and Treatment Strategies. Circulation 2021, 144, 728–745. [Google Scholar] [CrossRef]
- Gao, L.; Wang, L.Y.; Liu, Z.Q.; Jiang, D.; Wu, S.Y.; Guo, Y.Q.; Tao, H.M.; Sun, M.; You, L.N.; Qin, S.; et al. TNAP inhibition attenuates cardiac fibrosis induced by myocardial infarction through deactivating TGF-β1/Smads and activating P53 signaling pathways. Cell Death Dis. 2020, 11, 44. [Google Scholar] [CrossRef]
- Li, Y.Q.; Wang, J.Y.; Qian, Z.Q.; Li, Y.L.; Li, W.N.; Gao, Y.; Yang, D.L. Osthole inhibits intimal hyperplasia by regulating the NF-κB and TGF-β1/Smad2 signalling pathways in the rat carotid artery after balloon injury. Eur. J. Pharmacol. 2017, 811, 232–239. [Google Scholar] [CrossRef]
- Suwanabol, P.A.; Seedial, S.M.; Shi, X.; Zhang, F.; Yamanouchi, D.; Roenneburg, D.; Liu, B.; Kent, K.C. Transforming growth factor-β increases vascular smooth muscle cell proliferation through the Smad3 and extracellular signal-regulated kinase mitogen-activated protein kinases pathways. J. Vasc. Surg. 2012, 56, 446–454. [Google Scholar] [CrossRef]
- Zhu, Y.; Takayama, T.; Wang, B.; Kent, A.; Zhang, M.; Binder, B.Y.; Urabe, G.; Shi, Y.; DiRenzo, D.; Goel, S.A.; et al. Restenosis Inhibition and Re-differentiation of TGFβ/Smad3-activated Smooth Muscle Cells by Resveratrol. Sci. Rep. 2017, 7, 41916. [Google Scholar] [CrossRef]
- Lan, W.; Yang, F.; Li, Z.; Liu, L.; Sang, H.; Jiang, Y.; Xiong, Y.; Zhang, R. Human urine kininogenase attenuates balloon-induced intimal hyperplasia in rabbit carotid artery through transforming growth factor β1/Smad2/3 signaling pathway. J. Vasc. Surg. 2016, 64, 1074–1083. [Google Scholar] [CrossRef]
- Lu, P.; Wang, S.; Cai, W.; Sheng, J. Role of TGF-β1/Smad3 signaling pathway in secretion of type I and III collagen by vascular smooth muscle cells of rats undergoing balloon injury. J. Biomed. Biotechnol. 2012, 2012, 965953. [Google Scholar] [CrossRef]
- Wu, J.K.; Yang, Q.; Li, Y.Y.; Xie, D.J. [Effect of leech on VSMCs in early atherosclerosis rats via p38MAPK signaling pathway]. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2017, 42, 3191–3197. [Google Scholar] [CrossRef]
- Shi, X.; Guo, L.W.; Seedial, S.; Takayama, T.; Wang, B.; Zhang, M.; Franco, S.R.; Si, Y.; Chaudhary, M.A.; Liu, B.; et al. Local CXCR4 Upregulation in the Injured Arterial Wall Contributes to Intimal Hyperplasia. Stem Cells 2016, 34, 2744–2757. [Google Scholar] [CrossRef]
- Wei, X.; Fang, Z.; Sheng, J.; Wang, Y.; Lu, P. Honokiol-mesoporous Silica Nanoparticles Inhibit Vascular Restenosis via the Suppression of TGF-β Signaling Pathway. Int. J. Nanomed. 2020, 15, 5239–5252. [Google Scholar] [CrossRef]
- Otsuka, G.; Agah, R.; Frutkin, A.D.; Wight, T.N.; Dichek, D.A. Transforming growth factor beta 1 induces neointima formation through plasminogen activator inhibitor-1-dependent pathways. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 737–743. [Google Scholar] [CrossRef]
- Sun, Q.; Chen, L.; Zhou, D.; Yang, Q.; Song, W.; Chen, D.; Wang, W. Mfn2 inhibits chronic rejection of the rat abdominal aorta by regulating TGF-β1 levels. Transpl. Immunol. 2019, 55, 101211. [Google Scholar] [CrossRef] [PubMed]
- Sluijter, J.P.; Verloop, R.E.; Pulskens, W.P.; Velema, E.; Grimbergen, J.M.; Quax, P.H.; Goumans, M.J.; Pasterkamp, G.; de Kleijn, D.P. Involvement of furin-like proprotein convertases in the arterial response to injury. Cardiovasc. Res. 2005, 68, 136–143. [Google Scholar] [CrossRef]
- Weitz, J.I.; Byrne, J.; Clagett, G.P.; Farkouh, M.E.; Porter, J.M.; Sackett, D.L.; Strandness, D.E., Jr.; Taylor, L.M. Diagnosis and treatment of chronic arterial insufficiency of the lower extremities: A critical review. Circulation 1996, 94, 3026–3049. [Google Scholar] [CrossRef] [PubMed]
- Russo, I.; Frangogiannis, N.G. Diabetes-associated cardiac fibrosis: Cellular effectors, molecular mechanisms and therapeutic opportunities. J. Mol. Cell. Cardiol. 2016, 90, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Son, M.; Park, C.H.; Jang, J.T.; Son, K.H.; Byun, K. Pyrogallol-Phloroglucinol-6,6-Bieckolon Attenuates Vascular Smooth Muscle Cell Proliferation and Phenotype Switching in Hyperlipidemia through Modulation of Chemokine Receptor 5. Mar. Drugs 2020, 18, 393. [Google Scholar] [CrossRef]
- Davies, M.G.; Dalen, H.; Kim, J.H.; Barber, L.; Svendsen, E.; Hagen, P.O. The influence of the combined presence of diabetes mellitus and hypercholesterolaemia on the function and morphology of experimental vein grafts. Eur. J. Vasc. Endovasc. Surg. Off. J. Eur. Soc. Vasc. Surg. 1995, 10, 142–155. [Google Scholar] [CrossRef]
- Brooks, M.M.; Jones, R.H.; Bach, R.G.; Chaitman, B.R.; Kern, M.J.; Orszulak, T.A.; Follmann, D.; Sopko, G.; Blackstone, E.H.; Califf, R.M. Predictors of mortality and mortality from cardiac causes in the bypass angioplasty revascularization investigation (BARI) randomized trial and registry. For the BARI Investigators. Circulation 2000, 101, 2682–2689. [Google Scholar] [CrossRef]
- Kornowski, R.; Mintz, G.S.; Kent, K.M.; Pichard, A.D.; Satler, L.F.; Bucher, T.A.; Hong, M.K.; Popma, J.J.; Leon, M.B. Increased restenosis in diabetes mellitus after coronary interventions is due to exaggerated intimal hyperplasia. A serial intravascular ultrasound study. Circulation 1997, 95, 1366–1369. [Google Scholar] [CrossRef]
- Li, Q.; Fu, J.; Xia, Y.; Qi, W.; Ishikado, A.; Park, K.; Yokomizo, H.; Huang, Q.; Cai, W.; Rask-Madsen, C.; et al. Homozygous receptors for insulin and not IGF-1 accelerate intimal hyperplasia in insulin resistance and diabetes. Nat. Commun. 2019, 10, 4427. [Google Scholar] [CrossRef]
- Westerweel, P.E.; van Velthoven, C.T.; Nguyen, T.Q.; den Ouden, K.; de Kleijn, D.P.; Goumans, M.J.; Goldschmeding, R.; Verhaar, M.C. Modulation of TGF-β/BMP-6 expression and increased levels of circulating smooth muscle progenitor cells in a type I diabetes mouse model. Cardiovasc. Diabetol. 2010, 9, 55. [Google Scholar] [CrossRef]
- Liu, X.; Luo, F.; Pan, K.; Wu, W.; Chen, H. High glucose upregulates connective tissue growth factor expression in human vascular smooth muscle cells. BMC Cell Biol. 2007, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.Q.; Yu, J.; Guo, Q.X.; Zhang, X.M. [Analysis of relative factors of restenosis after artificially grafting bypasses on chronic ischemia of lower extremities]. Zhonghua Wai Ke Za Zhi [Chin. J. Surg.] 2009, 47, 1698–1701. [Google Scholar] [PubMed]
- Aronson, D. Potential role of advanced glycosylation end products in promoting restenosis in diabetes and renal failure. Med. Hypotheses 2002, 59, 297–301. [Google Scholar] [CrossRef]
- Wu, F.; Yang, J.; Liu, J.; Wang, Y.; Mu, J.; Zeng, Q.; Deng, S.; Zhou, H. Signaling pathways in cancer-associated fibroblasts and targeted therapy for cancer. Signal Transduct. Target. Ther. 2021, 6, 218. [Google Scholar] [CrossRef]
- Liu, S.; Ren, J.; Ten Dijke, P. Targeting TGFβ signal transduction for cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, T.C.; Johnson, J.M.; Auster, M.; Huynh, C.; Muralidharan, S.; Contreras, M.; LoGerfo, F.W.; Pradhan-Nabzdyk, L. Intraluminal delivery of thrombospondin-2 small interfering RNA inhibits the vascular response to injury in a rat carotid balloon angioplasty model. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 109–119. [Google Scholar] [CrossRef]
- Kapur, N.K.; Bian, C.; Lin, E.; Deming, C.B.; Sperry, J.L.; Hansen, B.S.; Kakouros, N.; Rade, J.J. Inhibition of transforming growth factor-β restores endothelial thromboresistance in vein grafts. J. Vasc. Surg. 2011, 54, 1117–1123.e1111. [Google Scholar] [CrossRef]
- Pethig, K.; Heublein, B.; Wahlers, T.; Dannenberg, O.; Oppelt, P.; Haverich, A. Mycophenolate mofetil for secondary prevention of cardiac allograft vasculopathy: Influence on inflammation and progression of intimal hyperplasia. J. Heart Lung Transplant. Off. Publ. Int. Soc. Heart Transplant. 2004, 23, 61–66. [Google Scholar] [CrossRef]
- Jin, D.; Ueda, H.; Takai, S.; Muramatsu, M.; Furubayashi, K.; Ibaraki, T.; Kishi, K.; Katsuoka, Y.; Miyazaki, M. Roles of chymase in stenosis occurring after polytetrafluoroethylene graft implantations. Life Sci. 2007, 81, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.; Lim, C.S.; Brookes, J.; Tsui, J.; Hamilton, G. Emerging importance of molecular pathogenesis of vascular malformations in clinical practice and classifications. Vasc. Med. 2020, 25, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Grootaert, M.O.J.; Moulis, M.; Roth, L.; Martinet, W.; Vindis, C.; Bennett, M.R.; De Meyer, G.R.Y. Vascular smooth muscle cell death, autophagy and senescence in atherosclerosis. Cardiovasc. Res. 2018, 114, 622–634. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Xu, J.; Zhang, B.; Liu, J.; Liang, C.; Hua, J.; Meng, Q.; Yu, X.; Shi, S. Ferroptosis, necroptosis, and pyroptosis in anticancer immunity. J. Hematol. Oncol. 2020, 13, 110. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Function | Super Pathway |
|---|---|---|
| Dusp1 | Dual specificity phosphatase that dephosphorylates MAP kinase MAPK1/ERK2 on both ‘Thr-183’ and ‘Tyr-185’ | RAF/MAP kinase cascade |
| MAP kinase signaling | ||
| Neuropathic pain signaling in dorsal horn neurons | ||
| ERK signaling | ||
| TGF-β pathway | ||
| Serpine1 | Primary inhibitor of tissue-type plasminogen activator (PLAT) and urokinase-type plasminogen activator (PLAU) | Response to elevated platelet cytosolic Ca2+ |
| Defects of contact activation system (CAS) and kallikrein/kinin system (KKS) | ||
| Gene expression (transcription) | ||
| Circadian clock | ||
| Signaling by TGF-β family members |
| No. | Author | Source | Research Content | Conclusion |
|---|---|---|---|---|
| 1 | Pedro Serralheiro [30] | Healthy saphenous vein sample | TGF-β1 modulates MMP, TIMP | Involvement of TGF-β1 in the vein wall pathology. |
| 2 | Sarah J George [31] | Segments of human saphenous vein | TGF-β1 modulates smooth muscle cell proliferation or apoptosis | Plasmin, generated by uPA, appears to be an important activator of endogenous latent TGF-β. |
| 3 | Pedro Serralheiro [32] | Saphenous vein samples | TGF-β1 modulates MMP, TIMP | Decreased expression of TGFβRs might suggest a reduction in TGF-β1 participation in the MMP/TIMP imbalance throughout CVeD progression. |
| 4 | Gemma Pascual [25] | Saphenous veins in patients with varicose veins and normal subjects | TGF-β1 modulates chronic venous insufficiency | Aging and varicose pathology induce dysregulation of TGF-β1 that could play an important role in the fibrous process. |
| 5 | Ji-Chang Wang [26] | Saphenous veins in patients with varicose veins and normal subjects | PGE2 modulates MMP, TGF-β | PGE2 can affect the remodeling of the extracellular matrix and reduce the elasticity of the vascular walls by promoting the synthesis of TGF-β1 and MMP-1. |
| 6 | Julia Buján [27] | Saphenous veins in patients with varicose veins and normal subjects | Varicose veins specimens showed greater LTBP-2 and TGF expression | Varicose veins specimens showed greater LTBP-2 and TGF expression. |
| 7 | Theresa Jacob [28] | Saphenous veins in patients with varicose veins and normal subjects | TGF-β1 modulates iNOS | The increased expression of TGF-β1 and presence of macrophages, correlating with overproduction of iNOS, may be associated with varicosity development and deserves further study. |
| 8 | Radoslaw Kowalewski [29] | Saphenous veins in patients with varicose veins and normal subjects | TGF-β RII modulates TGF-β | Increased TGF-β RII expression and activation in the wall of varicose veins may be involved in extracellular matrix remodeling related to TGF-β1 and supports its role in the disease pathogenesis. |
| 9 | S Mii [33] | SMC derived from human saphenous vein | TGF-β modulates PDGF, b-FGF | TGF-β inhibits both migration and proliferation of human SMC. |
| Animal Model | Intervention | Result | Reference |
|---|---|---|---|
| Rats: myocardial infarction | TNAP ↓ → TGF-β1 ↓, α-SMA ↓, Vimentin ↓, and fibronectin ↓ | Provides direct evidence that inhibition of TNAP is a novel regulator in cardiac fibrosis and exerts antifibrotic effect mainly through AMPK-TGF-β1/Smads and p53 signals. | Gao et al., 2020 [73] |
| Rats: carotid artery after balloon injury | Osthole → TGF-β1 ↓, p-Smad2 ↓, TNF-α ↓, IL-1β ↓, NF-κB ↓ | Osthole inhibited neointimal hyperplasia in balloon-induced rat carotid artery injury, and the mechanism may involve NF-κB, IL-1β, and TNF-α downregulation, and TGF-β1/Smad2 signaling pathway inhibition. | Li et al., 2017 [74] |
| Mice and human vasculopathy samples | LMO7 ↓ → TGF-β ↑, SMAD3 ↑ | LMO7 induced by TGF-β and limits vascular fibrotic responses through negative feedback regulation of TGF-β pathway. | Xie et al., 2019 [53] |
| Rats: carotid balloon injury | ERK MAPK ↓ → TGF-β/Smad3 ↓ → VSMC proliferation ↓ | TGF-β enhances VSMC proliferation through a Smad3/ERK MAPK signaling pathway. | Suwanabol et al., 2012 [75] |
| Rats: carotid injury | Resveratrol-(Akt-mTOR pathway) → TGF-β/Smad3 ↓ | Resveratrol produces durable inhibition of all three prorestenotic pathologies; a rare feat among existing antirestenotic methods. | Zhu et al., 2017 [76] |
| Rabbits: balloon-injured carotid arteries | HUK→ TGF-β1/Smad2/3 ↓ → eNOS ↑ | HUK attenuates atherosclerosis formation and inhibits intimal hyperplasia by downregulating TGF-β1 expression and Smad2/3 phosphorylation, upregulating eNOS activity. | Lan et al., 2016 [77] |
| Rats: balloon-injured carotid arteries | Antisense Smad3 → collagen(1, 3) | Transfection of VSMCs with antisense Smad3 can reduce the secretion of type I and III collagen, which then inhibits intimal hyperplasia. | Lu et al., 2012 [78] |
| Rats: atherosclerosis | Leech → p38MAPK ↓ → TGF-β ↓, blood lipid ↓ | Leech may affect the p38MAPK signaling pathway to inhibit proliferation and promote apoptosis of VSMCs via reducing blood lipid levels and suppressing TGF-β. | Wu et al., 2017 [79] |
| Rats: injured arterial wall | TGF-β/Smad3 ↑ → SMC migration, CXCR4 ↑ | Upregulation of TGFβ/Smad3 in injured arteries induces local SMC CXCR4 expression and cell migration, and, consequently, IH. | Shi et al., 2016 [80] |
| Rats: carotid artery injury | HNK-MSNPs → Smad3 ↓ | Honokiol (HNK), a small-molecule polyphenol, can inhibit neointimal hyperplasia after balloon injury. | Wei et al., 2020 [81] |
| Mice: carotid artery injury | PAI-1 ↓ → TGF-β ↓ | PAI-1 is both a critical mediator of TGF-beta1-induced intimal growth and a key negative regulator of TGF-beta1 expression in the artery wall | Otsuka et al., 2006 [82] |
| Rats: aortals | Mfn2 ↑ → TGF-β ↓ | Mfn2 influences TGF-β/Smad pathway and may function as potential chronic rejection inhibitor. | Sun et al., 2019 [83] |
| Rabbit: femoral and iliac arteries | Furin-like PCs → TGF-β ↑ | Furin-like PCs are involved in arterial response to injury possibly through activation of TGF-beta–Smad signaling pathway. | Sluijter et al., 2005 [84] |
| NCT Number | Conditions | Brief Title | Start Date | Phase |
|---|---|---|---|---|
| NCT02842424 | Peripheral arterial disease | Ramipril Treatment of Claudication | July 2020 | Phase 4 |
| NCT00382213 | Atherosclerosis | A Randomized, Double-blind Study To Compare The Effects Of Olmesartan Medoxomil Versus Placebo In Patients With Established Atherosclerosis | June 2000 | Phase 3 |
| NCT02632877 | Peripheral arterial disease | Efficacy of Pirfenidone Plus MODD in Diabetic Foot Ulcers | January 2014 | Phase 1, Phase 2 |
| NCT04489251 | Pulmonary arterial hypertension | Assessment of the TGF-beta Pathway and Micro-RNA in Pediatric Pulmonary Arterial Hypertension | July 2020 | - |
| NCT05269849 | Hereditary hemorrhagic telangiectasia | Sirolimus for Nosebleeds in HHT | March 2022 | Phase 2 |
| NCT04998227 | Acute coronary syndrome | Latent TGF-β-binding Proteins Affect the Fibrotic Process in Renal Impairment and Cardiac Dysfunction | August 2021 | - |
| NCT01847716 | Pulmonary arterial hypertension | Transforming Growth Factor Beta Signalling in the Development of Muscle Weakness in Pulmonary Arterial Hypertension | October 2013 | - |
| NCT01893710 | Peritoneal dialysis | International (Pediatric) Peritoneal Biobank | February 2011 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, C.; Ye, P.; Zhang, X.; Esmaeili, E.; Li, Y.; Lü, P.; Cai, C. The Role of TGF-β Signaling in Saphenous Vein Graft Failure after Peripheral Arterial Disease Bypass Surgery. Int. J. Mol. Sci. 2023, 24, 10381. https://doi.org/10.3390/ijms241210381
He C, Ye P, Zhang X, Esmaeili E, Li Y, Lü P, Cai C. The Role of TGF-β Signaling in Saphenous Vein Graft Failure after Peripheral Arterial Disease Bypass Surgery. International Journal of Molecular Sciences. 2023; 24(12):10381. https://doi.org/10.3390/ijms241210381
Chicago/Turabian StyleHe, Changhuai, Pin Ye, Xuecheng Zhang, Elham Esmaeili, Yiqing Li, Ping Lü, and Chuanqi Cai. 2023. "The Role of TGF-β Signaling in Saphenous Vein Graft Failure after Peripheral Arterial Disease Bypass Surgery" International Journal of Molecular Sciences 24, no. 12: 10381. https://doi.org/10.3390/ijms241210381
APA StyleHe, C., Ye, P., Zhang, X., Esmaeili, E., Li, Y., Lü, P., & Cai, C. (2023). The Role of TGF-β Signaling in Saphenous Vein Graft Failure after Peripheral Arterial Disease Bypass Surgery. International Journal of Molecular Sciences, 24(12), 10381. https://doi.org/10.3390/ijms241210381