
An Exploration of the Coherent Effects between METTL3 and NDUFA10 on Alzheimer’s Disease
 ,
,
Abstract
1. Introduction
2. Results
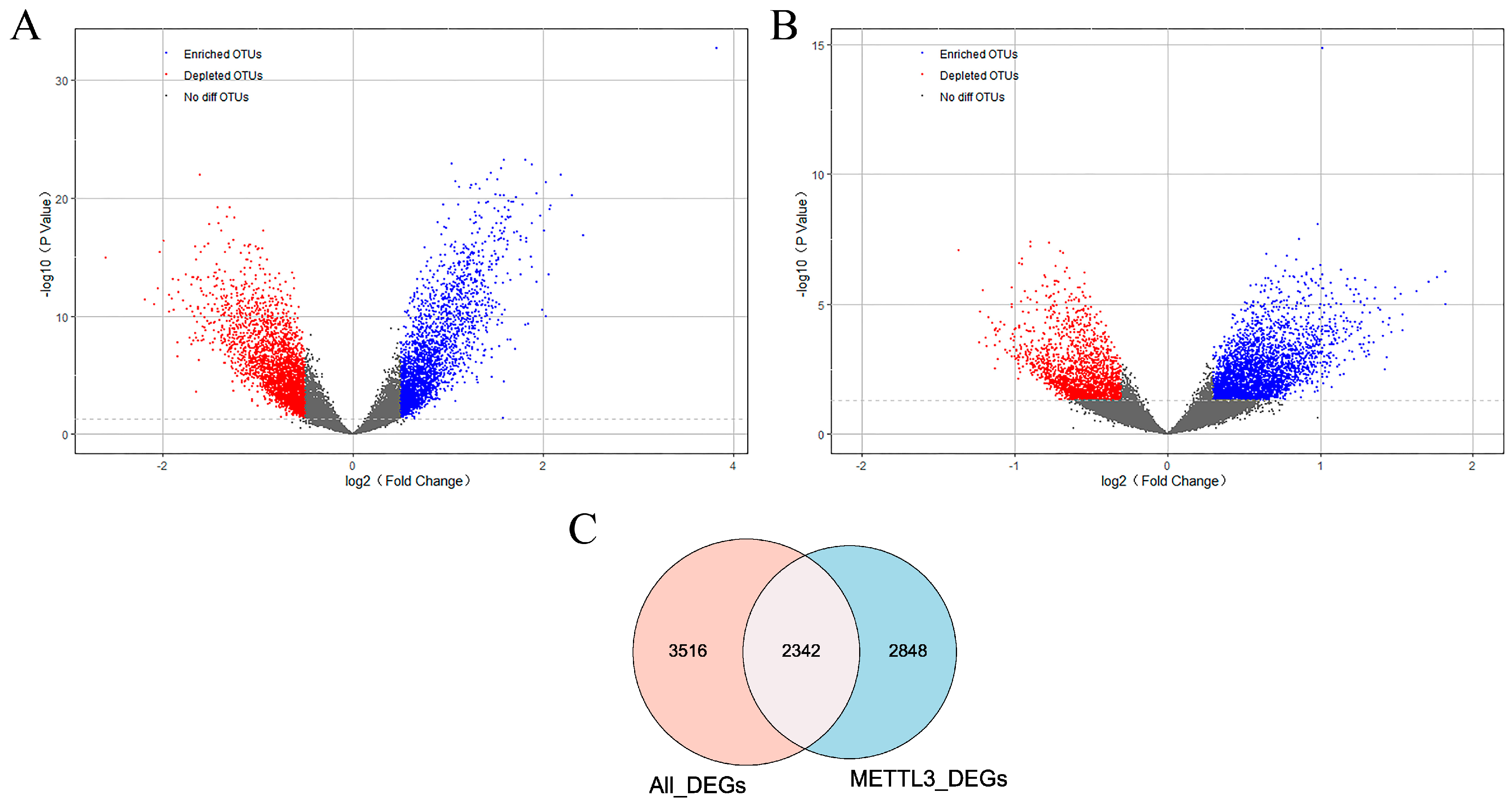
2.1. m6A Writer Gene METTL3 Is Downregulated in AD
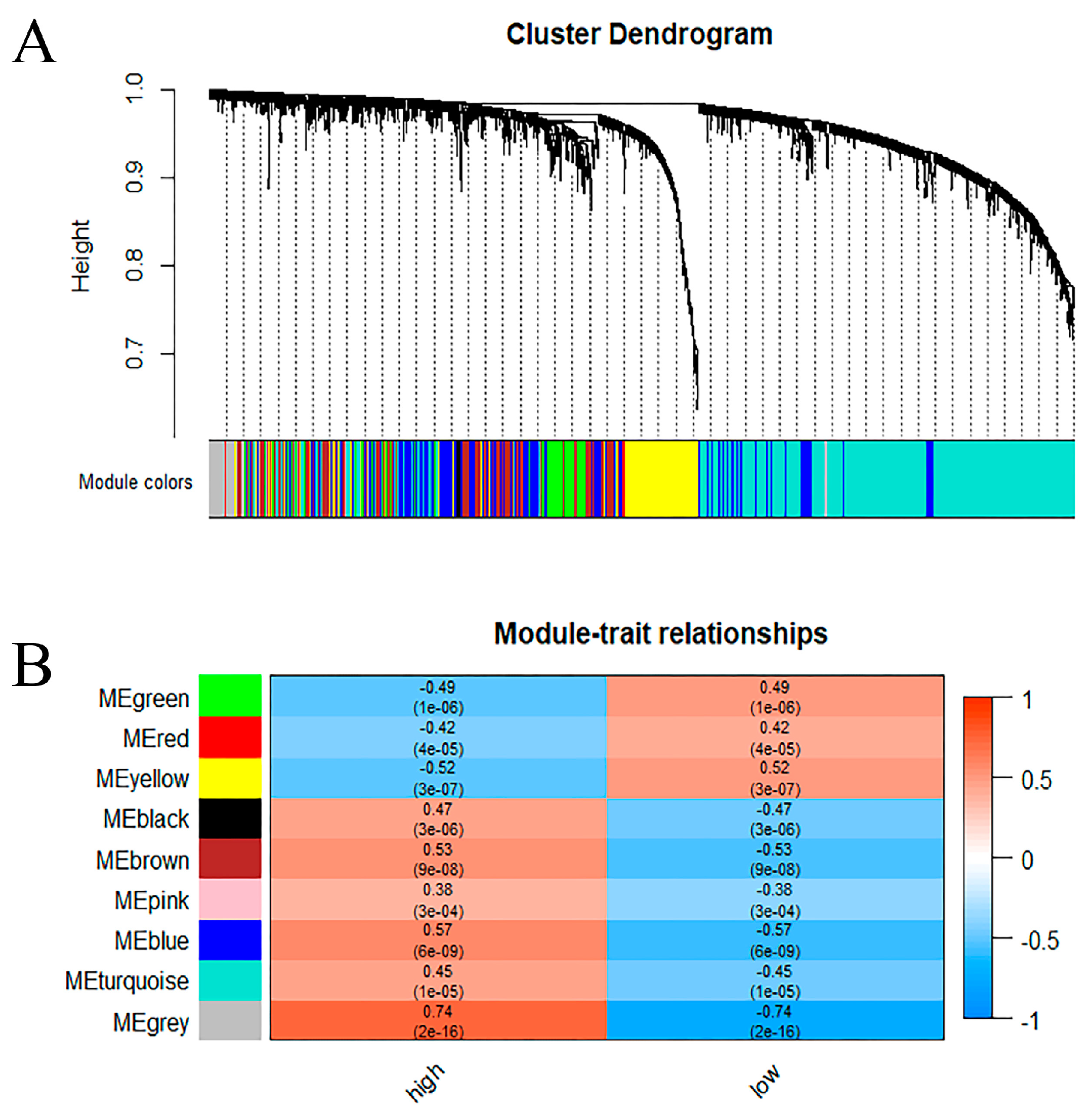
2.2. The Set of Candidate AD Genes Positively Correlated with METTL3
2.3. Functional Enrichment Analysis for the Genes That Positively Related to METTL3
2.4. The Effect of METTL3 on Energy Metabolism
2.5. Co-Effects of METTL3 and NDUFA10 on AD
3. Discussion
4. Materials and Methods
4.1. Data Collection and Preprocessing
4.2. Screening of METTl3-Related AD Genes
4.3. Identification of Key Module Based on the METTl3-Related AD Genes
4.4. PPI Network Construction
4.5. Functional Enrichment Analysis of Key Module Genes and Key AD Genes
4.6. Identification of Candidate Gene
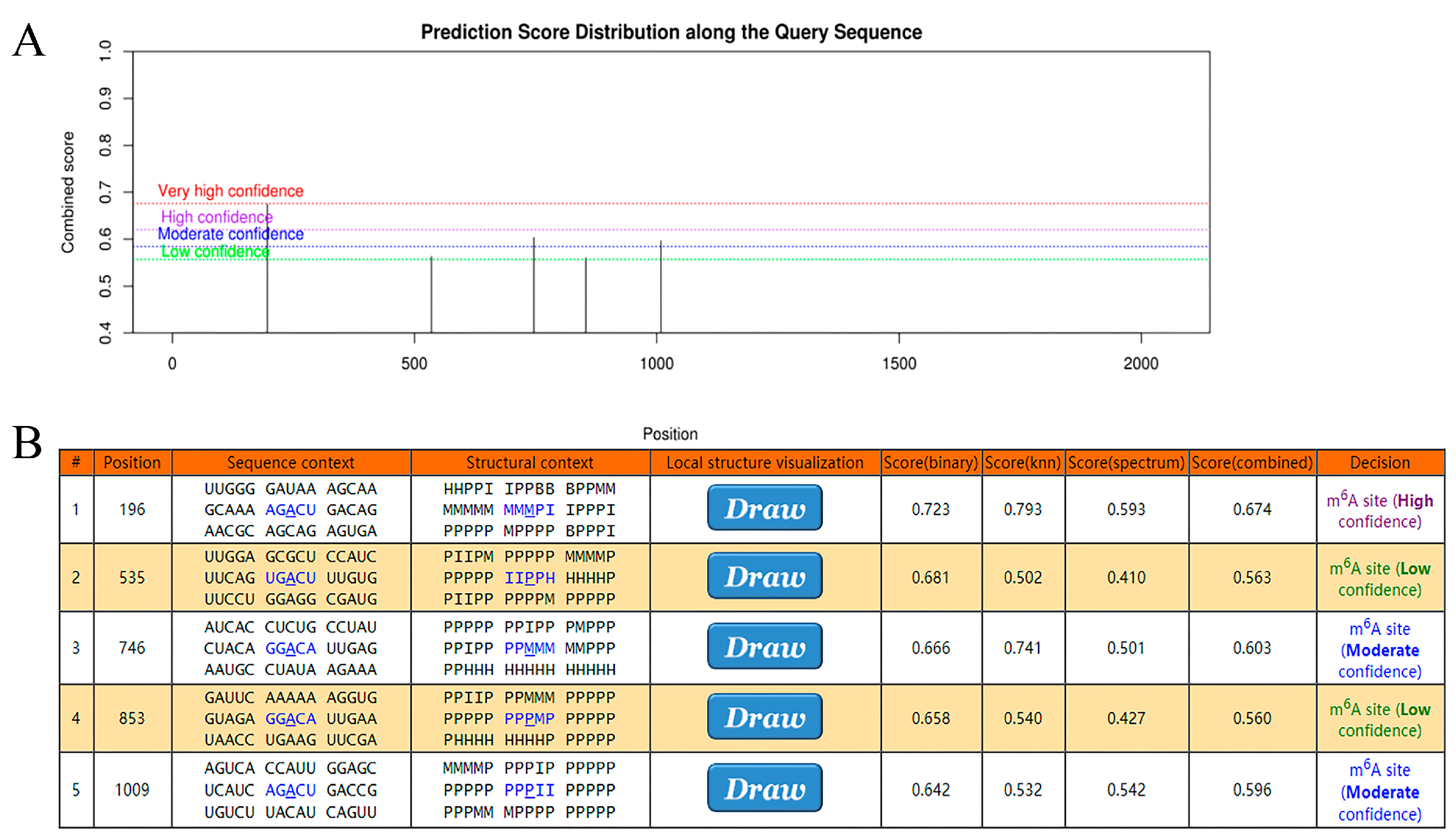
4.7. The m6A Mechanism between METTL3 and Candidate Gene
4.8. Construction and Validation of the Diagnostic Model for AD
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s Disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Kim, S.; Park, K.W.; Park, K.H.; Youn, Y.C.; Lee, D.W.; Lee, J.-Y.; Lee, J.H.; Jeong, J.H.; Choi, S.H.; et al. A Comparative Evaluation of the KDSQ-C, AD8, and SMCQ as a Cognitive Screening Test to Be Used in National Medical Check-Ups in Korea. J. Korean Med. Sci. 2019, 34, e111. [Google Scholar] [CrossRef] [PubMed]
- Sperling, R.A.; Aisen, P.S.; Beckett, L.A.; Bennett, D.A.; Craft, S.; Fagan, A.M.; Iwatsubo, T.; Jack, C.R., Jr.; Kaye, J.; Montine, T.J.; et al. Toward Defining the Preclinical Stages of Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Alzheimer’s Dement. 2011, 7, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, M.; Yang, Q.; Xu, X.; Zhang, R.; Chen, X.; Le, W. The Electrophysiological and Neuropathological Profiles of Cerebellum in APPswe/PS1ΔE9 Mice: A Hypothesis on the Role of Cerebellum in Alzheimer’s Disease. Alzheimer’s Dement. 2022, 1–11. [Google Scholar] [CrossRef]
- Bateman, R.J.; Xiong, C.; Benzinger, T.L.S.; Fagan, A.M.; Goate, A.; Fox, N.C.; Marcus, D.S.; Cairns, N.J.; Xie, X.; Blazey, T.M.; et al. Clinical and Biomarker Changes in Dominantly Inherited Alzheimer’s Disease. N. Engl. J. Med. 2012, 367, 795–804. [Google Scholar] [CrossRef]
- Frye, M.; Harada, B.T.; Behm, M.; He, C. RNA Modifications Modulate Gene Expression during Development. Science 2018, 361, 1346–1349. [Google Scholar] [CrossRef]
- Neueder, A. RNA-Mediated Disease Mechanisms in Neurodegenerative Disorders. J. Mol. Biol. 2019, 431, 1780–1791. [Google Scholar] [CrossRef]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, Writing and Erasing MRNA Methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef]
- Shi, H.; Wei, J.; He, C. Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers. Mol. Cell 2019, 74, 640–650. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive Analysis of MRNA Methylation Reveals Enrichment in 3′ UTRs and near Stop Codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef]
- Widagdo, J.; Zhao, Q.-Y.; Kempen, M.-J.; Tan, M.C.; Ratnu, V.S.; Wei, W.; Leighton, L.; Spadaro, P.A.; Edson, J.; Anggono, V.; et al. Experience-Dependent Accumulation of N6-Methyladenosine in the Prefrontal Cortex Is Associated with Memory Processes in Mice. J. Neurosci. 2016, 36, 6771–6777. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ren, Y.; Mao, K.; Hua, F.; Yang, Y.; Wei, N.; Yue, C.; Li, D.; Zhang, H. FTO Is Involved in Alzheimer’s Disease by Targeting TSC1-MTOR-Tau Signaling. Biochem. Biophys. Res. Commun. 2018, 498, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Röh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of M6A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403.e9. [Google Scholar] [CrossRef] [PubMed]
- Merkurjev, D.; Hong, W.-T.; Iida, K.; Oomoto, I.; Goldie, B.J.; Yamaguti, H.; Ohara, T.; Kawaguchi, S.; Hirano, T.; Martin, K.C.; et al. Synaptic N6-Methyladenosine (M6A) Epitranscriptome Reveals Functional Partitioning of Localized Transcripts. Nat. Neurosci. 2018, 21, 1004–1014. [Google Scholar] [CrossRef]
- Walters, B.J.; Mercaldo, V.; Gillon, C.J.; Yip, M.; Neve, R.L.; Boyce, F.M.; Frankland, P.W.; Josselyn, S.A. The Role of The RNA Demethylase FTO (Fat Mass and Obesity-Associated) and MRNA Methylation in Hippocampal Memory Formation. Neuropsychopharmacology 2017, 42, 1502–1510. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, X.; Weng, Y.-L.; Lu, Z.; Liu, Y.; Lu, Z.; Li, J.; Hao, P.; Zhang, Y.; Zhang, F.; et al. M6A Facilitates Hippocampus-Dependent Learning and Memory through YTHDF1. Nature 2018, 563, 249–253. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, M.; Xie, D.; Huang, Z.; Zhang, L.; Yang, Y.; Ma, D.; Li, W.; Zhou, Q.; Yang, Y.-G.; et al. METTL3-Mediated N6-Methyladenosine MRNA Modification Enhances Long-Term Memory Consolidation. Cell Res. 2018, 28, 1050–1061. [Google Scholar] [CrossRef]
- Zhao, F.; Xu, Y.; Gao, S.; Qin, L.; Austria, Q.; Siedlak, S.L.; Pajdzik, K.; Dai, Q.; He, C.; Wang, W.; et al. METTL3-Dependent RNA M6A Dysregulation Contributes to Neurodegeneration in Alzheimer’s Disease through Aberrant Cell Cycle Events. Mol. Neurodegener. 2021, 16, 70. [Google Scholar] [CrossRef]
- Molina-Granada, D.; González-Vioque, E.; Dibley, M.G.; Cabrera-Pérez, R.; Vallbona-Garcia, A.; Torres-Torronteras, J.; Sazanov, L.A.; Ryan, M.T.; Cámara, Y.; Martí, R. Most Mitochondrial DGTP Is Tightly Bound to Respiratory Complex I through the NDUFA10 Subunit. Commun. Biol. 2022, 5, 620. [Google Scholar] [CrossRef]
- Xie, J.; Zhao, Y.; Dong, N.; Tian, X.; Feng, J.; Liu, P.; Li, M.; Wang, M.; Ying, X.; Yuan, J.; et al. Proteomics and Transcriptomics Jointly Identify the Key Role of Oxidative Phosphorylation in Fluoride-Induced Myocardial Mitochondrial Dysfunction in Rats. Ecotoxicol. Environ. Saf. 2021, 218, 112271. [Google Scholar] [CrossRef]
- Oruganty-Das, A.; Ng, T.; Udagawa, T.; Goh, E.L.K.; Richter, J.D. Translational Control of Mitochondrial Energy Production Mediates Neuron Morphogenesis. Cell Metab. 2012, 16, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Manjeri, G.R.; Rodenburg, R.J.; Blanchet, L.; Roelofs, S.; Nijtmans, L.G.; Smeitink, J.A.; Driessen, J.J.; Koopman, W.J.H.; Willems, P.H. Increased Mitochondrial ATP Production Capacity in Brain of Healthy Mice and a Mouse Model of Isolated Complex I Deficiency after Isoflurane Anesthesia. J. Inherit. Metab. Dis. 2016, 39, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lv, D.; Yang, X.; Li, M.; Zhang, H. M6A RNA Methylation Regulators Could Contribute to the Occurrence of Chronic Obstructive Pulmonary Disease. J. Cell. Mol. Med. 2020, 24, 12706–12715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, Y.; Guo, F.; Li, S.; Cui, H. RNA N6-Methyladenosine Modifications and Its Roles in Alzheimer’s Disease. Front. Cell. Neurosci. 2022, 16, 820378. [Google Scholar] [CrossRef] [PubMed]
- Koshi-Mano, K.; Mano, T.; Morishima, M.; Murayama, S.; Tamaoka, A.; Tsuji, S.; Toda, T.; Iwata, A. Neuron-Specific Analysis of Histone Modifications with Post-Mortem Brains. Sci. Rep. 2020, 10, 3767. [Google Scholar] [CrossRef]
- Du, B.; Zhang, Y.; Liang, M.; Du, Z.; Li, H.; Fan, C.; Zhang, H.; Jiang, Y.; Bi, X. N6-Methyladenosine (M6A) Modification and Its Clinical Relevance in Cognitive Dysfunctions. Aging 2021, 13, 20716–20737. [Google Scholar] [CrossRef]
- Shafik, A.M.; Zhang, F.; Guo, Z.; Dai, Q.; Pajdzik, K.; Li, Y.; Kang, Y.; Yao, B.; Wu, H.; He, C.; et al. N6-Methyladenosine Dynamics in Neurodevelopment and Aging, and Its Potential Role in Alzheimer’s Disease. Genome Biol. 2021, 22, 17. [Google Scholar] [CrossRef]
- Adamopoulos, P.G.; Athanasopoulou, K.; Daneva, G.N.; Scorilas, A. The Repertoire of RNA Modifications Orchestrates a Plethora of Cellular Responses. Int. J. Mol. Sci. 2023, 24, 2387. [Google Scholar] [CrossRef]
- Phillips, S.; Mishra, T.; Khadka, S.; Bohan, D.; Espada, C.E.; Maury, W.; Wu, L. Epitranscriptomic N6-Methyladenosine Profile of SARS-CoV-2-Infected Human Lung Epithelial Cells. Microbiol. Spectr. 2023, 11, e03943-22. [Google Scholar] [CrossRef]
- Huang, H.; Camats-Perna, J.; Medeiros, R.; Anggono, V.; Widagdo, J. Altered Expression of the M6A Methyltransferase METTL3 in Alzheimer’s Disease. eNeuro 2020, 7, ENEURO.0125-20.2020. [Google Scholar] [CrossRef]
- Guo, Y.; Li, S.; Zeng, L.-H.; Tan, J. Tau-Targeting Therapy in Alzheimer’s Disease: Critical Advances and Future Opportunities. Ageing Neurodegener. Dis. 2022, 2, 11. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, M.; Zhu, F.; Li, X.; Yang, Y.; Yan, Q.; Jia, L.; Xie, L.; Chen, Z. METTL3 Is Essential for Postnatal Development of Brown Adipose Tissue and Energy Expenditure in Mice. Nat. Commun. 2020, 11, 1648. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.-Q.; Shuai, P.; Shan, Z.; Pang, C.-Y. Define a Function on Gene Order of DNA Microarray Data and Use It to Identify Genes Associated with Alzheimer’s Disease. In Proceedings of the 2011 IEEE International Conference on Granular Computing, Kaohsiung, Taiwan, 8–10 November 2011; pp. 836–839. [Google Scholar]
- Pang, C.; Jiang, G.; Wang, S.; Hu, B.; Liu, Q.; Deng, Y.; Huang, X. Gene Order Computation Using Alzheimer’s DNA Microarray Gene Expression Data and the Ant Colony Optimisation Algorithm. Int. J. Data Min. Bioinform. 2012, 6, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Fassone, E.; Rahman, S. Complex I Deficiency: Clinical Features, Biochemistry and Molecular Genetics. J. Med. Genet. 2012, 49, 578–590. [Google Scholar] [CrossRef]
- Rodenburg, R.J. Mitochondrial Complex I-Linked Disease. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2016, 1857, 938–945. [Google Scholar] [CrossRef]
- Abramov, A.Y.; Angelova, P.R. Cellular Mechanisms of Complex I–Associated Pathology. Biochem. Soc. Trans. 2019, 47, 1963–1969. [Google Scholar] [CrossRef]
- Hoefs, S.J.G.; van Spronsen, F.J.; Lenssen, E.W.H.; Nijtmans, L.G.; Rodenburg, R.J.; Smeitink, J.A.M.; van den Heuvel, L.P. NDUFA10 Mutations Cause Complex I Deficiency in a Patient with Leigh Disease. Eur. J. Hum. Genet. 2011, 19, 270–274. [Google Scholar] [CrossRef]
- Agip, A.-N.A.; Blaza, J.N.; Bridges, H.R.; Viscomi, C.; Rawson, S.; Muench, S.P.; Hirst, J. Cryo-EM Structures of Complex I from Mouse Heart Mitochondria in Two Biochemically Defined States. Nat. Struct. Mol. Biol. 2018, 25, 548–556. [Google Scholar] [CrossRef]
- Murayama, C.; Kawaguchi, A.T.; Kamijo, A.; Naito, K.; Kanazawa, M.; Tsukada, H. Monitoring Mitochondrial Complex-I Activity Using Novel PET Probe 18F-BCPP-EF Allows Early Detection of Radiotherapy Effect in Murine Squamous Cell Carcinoma. PLoS ONE 2017, 12, e0170911. [Google Scholar] [CrossRef]
- Terada, T.; Therriault, J.; Kang, M.S.P.; Savard, M.; Pascoal, T.A.; Lussier, F.; Tissot, C.; Wang, Y.-T.; Benedet, A.; Matsudaira, T.; et al. Mitochondrial Complex I Abnormalities Is Associated with Tau and Clinical Symptoms in Mild Alzheimer’s Disease. Mol. Neurodegener. 2021, 16, 28. [Google Scholar] [CrossRef]
- Terada, T.; Yokokura, M.; Obi, T.; Bunai, T.; Yoshikawa, E.; Ando, I.; Shimada, H.; Suhara, T.; Higuchi, M.; Ouchi, Y. In Vivo Direct Relation of Tau Pathology with Neuroinflammation in Early Alzheimer’s Disease. J. Neurol. 2019, 266, 2186–2196. [Google Scholar] [CrossRef] [PubMed]
- Terada, T.; Obi, T.; Bunai, T.; Matsudaira, T.; Yoshikawa, E.; Ando, I.; Futatsubashi, M.; Tsukada, H.; Ouchi, Y. In Vivo Mitochondrial and Glycolytic Impairments in Patients with Alzheimer Disease. Neurology 2020, 94, e1592–e1604. [Google Scholar] [CrossRef] [PubMed]
- Perez Ortiz, J.M.; Swerdlow, R.H. Mitochondrial Dysfunction in Alzheimer’s Disease: Role in Pathogenesis and Novel Therapeutic Opportunities. Br. J. Pharmacol. 2019, 176, 3489–3507. [Google Scholar] [CrossRef] [PubMed]
- Adav, S.S.; Park, J.E.; Sze, S.K. Quantitative Profiling Brain Proteomes Revealed Mitochondrial Dysfunction in Alzheimer’s Disease. Mol. Brain 2019, 12, 8. [Google Scholar] [CrossRef]
- Rhein, V.; Baysang, G.; Rao, S.; Meier, F.; Bonert, A.; Müller-Spahn, F.; Eckert, A. Amyloid-Beta Leads to Impaired Cellular Respiration, Energy Production and Mitochondrial Electron Chain Complex Activities in Human Neuroblastoma Cells. Cell Mol. Neurobiol. 2009, 29, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, L.; de Leon, M.; Murray, J.; E, L.; Lu, J.; Javier, E.; McHugh, P.; Swerdlow, R.H. Reduced Mitochondria Cytochrome Oxidase Activity in Adult Children of Mothers with Alzheimer’s Disease. J. Alzheimer’s Dis. 2011, 27, 483–490. [Google Scholar] [CrossRef]
- Hauptmann, S.; Scherping, I.; Dröse, S.; Brandt, U.; Schulz, K.L.; Jendrach, M.; Leuner, K.; Eckert, A.; Müller, W.E. Mitochondrial Dysfunction: An Early Event in Alzheimer Pathology Accumulates with Age in AD Transgenic Mice. Neurobiol. Aging 2009, 30, 1574–1586. [Google Scholar] [CrossRef]
- Appel, S.H. Oxidative Stress-Mediated Inflammation Promotes the Pathogenesis of Amyotrophic Lateral Sclerosis. Ageing Neurodegener. Dis. 2022, 2, 18. [Google Scholar] [CrossRef]
- Wang, X.; Feng, J.; Xue, Y.; Guan, Z.; Zhang, D.; Liu, Z.; Gong, Z.; Wang, Q.; Huang, J.; Tang, C.; et al. Structural Basis of N6-Adenosine Methylation by the METTL3–METTL14 Complex. Nature 2016, 534, 575–578. [Google Scholar] [CrossRef]
- Wang, P.; Doxtader, K.A.; Nam, Y. Structural Basis for Cooperative Function of Mettl3 and Mettl14 Methyltransferases. Mol. Cell 2016, 63, 306–317. [Google Scholar] [CrossRef]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 SnRNA M6A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell 2017, 169, 824–835.e14. [Google Scholar] [CrossRef] [PubMed]
- Ianniello, Z.; Paiardini, A.; Fatica, A. N6-Methyladenosine (M6A): A Promising New Molecular Target in Acute Myeloid Leukemia. Front. Oncol. 2019, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Lesbirel, S.; Wilson, S.A. The M6A-methylase Complex and MRNA Export. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2019, 1862, 319–328. [Google Scholar] [CrossRef]
- Ping, X.-L.; Sun, B.-F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP Is a Regulatory Subunit of the RNA N6-Methyladenosine Methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef]
- Yue, Y.; Liu, J.; Cui, X.; Cao, J.; Luo, G.; Zhang, Z.; Cheng, T.; Gao, M.; Shu, X.; Ma, H.; et al. VIRMA Mediates Preferential M6A MRNA Methylation in 3′UTR and near Stop Codon and Associates with Alternative Polyadenylation. Cell Discov. 2018, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Knuckles, P.; Lence, T.; Haussmann, I.U.; Jacob, D.; Kreim, N.; Carl, S.H.; Masiello, I.; Hares, T.; Villaseñor, R.; Hess, D.; et al. Zc3h13/Flacc Is Required for Adenosine Methylation by Bridging the MRNA-Binding Factor Rbm15/Spenito to the M6A Machinery Component Wtap/Fl(2)d. Genes Dev. 2018, 32, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Yu, G.; Zhu, X.; Peng, T.; Lv, Y. Critical Roles of FTO-Mediated MRNA M6A Demethylation in Regulating Adipogenesis and Lipid Metabolism: Implications in Lipid Metabolic Disorders. Genes Dis. 2022, 9, 51–61. [Google Scholar] [CrossRef]
- Hu, Y.; Gong, C.; Li, Z.; Liu, J.; Chen, Y.; Huang, Y.; Luo, Q.; Wang, S.; Hou, Y.; Yang, S.; et al. Demethylase ALKBH5 Suppresses Invasion of Gastric Cancer via PKMYT1 M6A Modification. Mol. Cancer 2022, 21, 34. [Google Scholar] [CrossRef]
- Huang, X.-T.; Li, J.-H.; Zhu, X.-X.; Huang, C.-S.; Gao, Z.-X.; Xu, Q.-C.; Zhao, W.; Yin, X.-Y. HNRNPC Impedes M6A-Dependent Anti-Metastatic Alternative Splicing Events in Pancreatic Ductal Adenocarcinoma. Cancer Lett. 2021, 518, 196–206. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of M6A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-Methyladenosine Modulates Messenger RNA Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-Methyladenosine-Dependent Regulation of Messenger RNA Stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 Facilitates Translation and Decay of N6-Methyladenosine-Modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.-S.; Hao, Y.-J.; Sun, B.-F.; Sun, H.-Y.; Li, A.; Ping, X.-L.; Lai, W.-Y.; et al. Nuclear M6A Reader YTHDC1 Regulates MRNA Splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef]
- Hsu, P.J.; Zhu, Y.; Ma, H.; Guo, Y.; Shi, X.; Liu, Y.; Qi, M.; Lu, Z.; Shi, H.; Wang, J.; et al. Ythdc2 Is an N6-Methyladenosine Binding Protein That Regulates Mammalian Spermatogenesis. Cell Res. 2017, 27, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-Methyladenosine by IGF2BP Proteins Enhances MRNA Stability and Translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Frequency |
|---|---|
| SDHA | 10 |
| NDUFA10 | 10 |
| GPI | 8 |
| YTHDC1 | 6 |
| HNRNPC | 6 |
| YTHDF2 | 5 |
| NDUFC2 | 5 |
| m6A Member | Gene Symbol | Mechanisms | References |
|---|---|---|---|
| Writers | METTL3 | Enables S-adenosyl-L-methionine binding | [50] |
| METTL14 | Contributes to mRNA-methyltransferase activity | [51] | |
| METTL16 | Enables METTL3/14 activity | [52,53] | |
| RBM15/15B | Recruits m6A complex to target sites of mRNA precursor | [54] | |
| WTAP | Regulates members and handles m6A complex formation | [55] | |
| VIRMA | Recruits METTL3/METTL14/WTAP to guide methylations | [56] | |
| ZC3H13 | Bridges RBM15/15B and WTAP | [57] | |
| Erasers | FTO | Removes m6A modification | [58] |
| ALKBH5 | Removes m6A modification | [59] | |
| Readers | HNRNPC | Mediates mRNA splicing | [60] |
| HNRNPA2B1 | Promotes primary miRNA processing | [61] | |
| YTHDF1 | Promotes mRNA translation | [62] | |
| YTHDF2 | Regulates mRNA stability | [63] | |
| YTHDF3 | Regulates mRNA translation and splicing | [64] | |
| YTHDC1 | Regulates mRNA Splicing | [65] | |
| YTHDC2 | Enhances mRNA translation and decreases mRNA abundance | [66] | |
| IGF2BP1/2/3 | Enhances mRNA stability | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Pang, X.; Guo, W.; Zhu, C.; Yu, L.; Song, X.; Wang, K.; Pang, C. An Exploration of the Coherent Effects between METTL3 and NDUFA10 on Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 10111. https://doi.org/10.3390/ijms241210111
Yang L, Pang X, Guo W, Zhu C, Yu L, Song X, Wang K, Pang C. An Exploration of the Coherent Effects between METTL3 and NDUFA10 on Alzheimer’s Disease. International Journal of Molecular Sciences. 2023; 24(12):10111. https://doi.org/10.3390/ijms241210111
Chicago/Turabian StyleYang, Lin, Xinping Pang, Wenbo Guo, Chengjiang Zhu, Lei Yu, Xianghu Song, Kui Wang, and Chaoyang Pang. 2023. "An Exploration of the Coherent Effects between METTL3 and NDUFA10 on Alzheimer’s Disease" International Journal of Molecular Sciences 24, no. 12: 10111. https://doi.org/10.3390/ijms241210111
APA StyleYang, L., Pang, X., Guo, W., Zhu, C., Yu, L., Song, X., Wang, K., & Pang, C. (2023). An Exploration of the Coherent Effects between METTL3 and NDUFA10 on Alzheimer’s Disease. International Journal of Molecular Sciences, 24(12), 10111. https://doi.org/10.3390/ijms241210111