

Albumin Is a Component of the Esterase Status of Human Blood Plasma
 , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
2.1. The Effect of Sequential Addition of Inhibitors on the Esterase Status of Plasma of Healthy Humans
- The use of BW284c51, a specific inhibitor of AChE, is not required for measuring the activity of BChE towards ATCh and BTCh in plasma after centrifugation at high rotations;
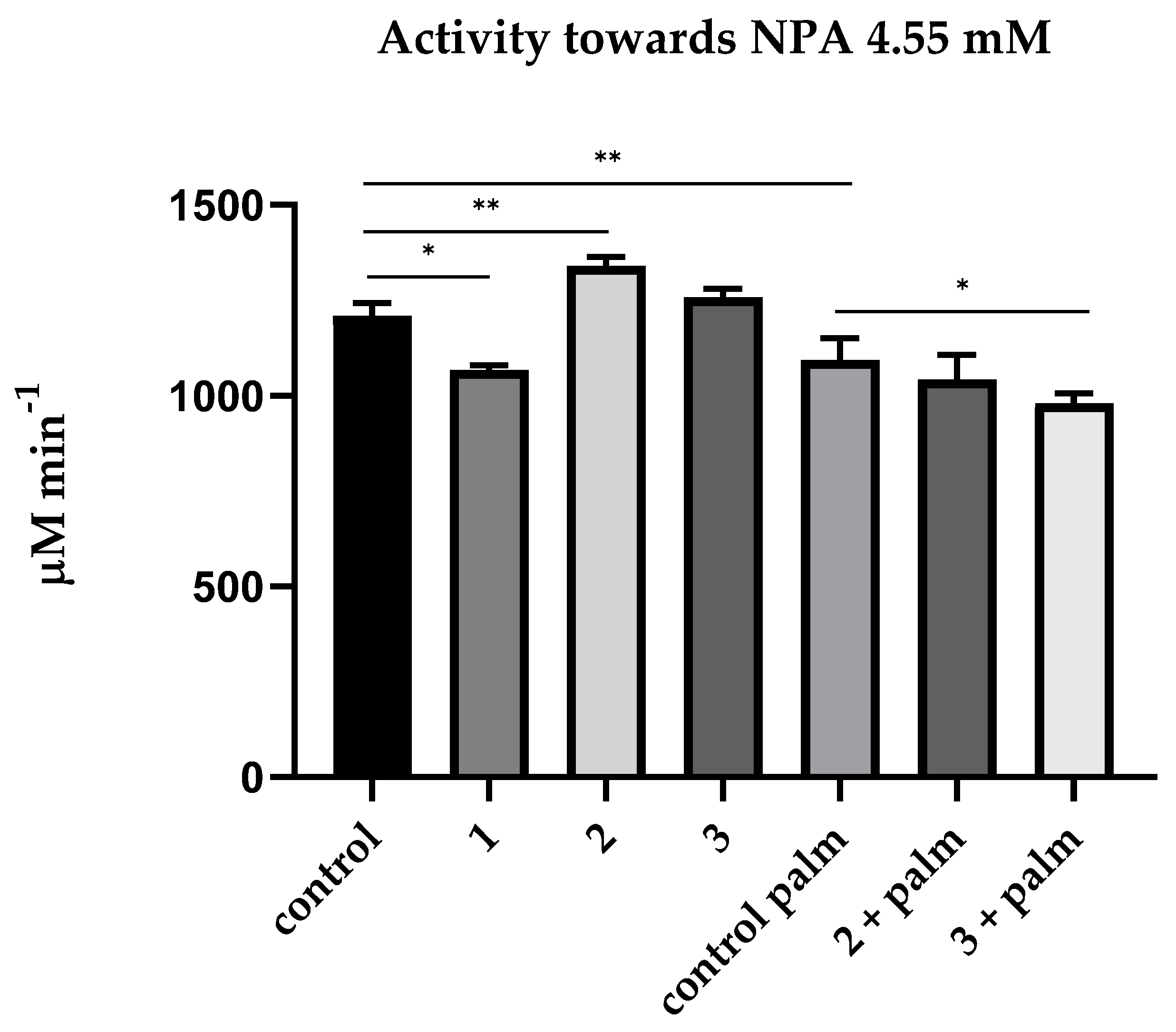
- To determine the activity of albumin towards NPA, it is necessary to add neostigmine (50 μM) and EDTA, and calculate the activity as the difference between the mean values with and without palmitate;
- Measurements of PON1 activity should be carried out ideally with two substrates (POX and PhA), but to spare the time and to avoid the risk of dealing with toxic POX in clinical laboratories, applying PhA alone with the addition of neostigmine 50 μM is quite adequate.
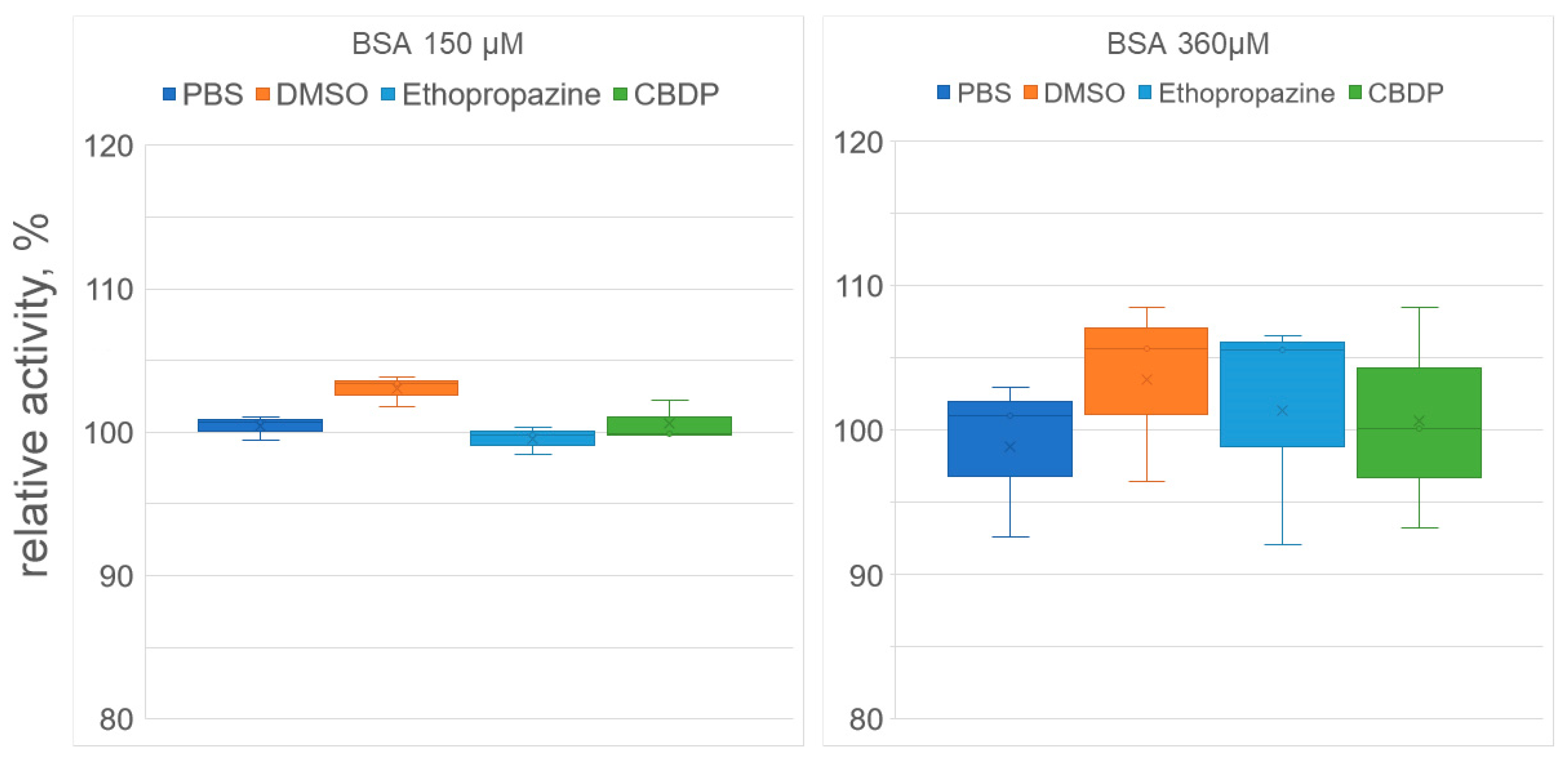
2.2. Effect of Ethopropazine and CBDP on the Esterase Activity of Albumin Preparation According to NMR Data
2.3. Effect of Ethopropazine and CBDP on the (Pseudo)esterase Activity of Albumin Preparation According to Spectrophotometric Data
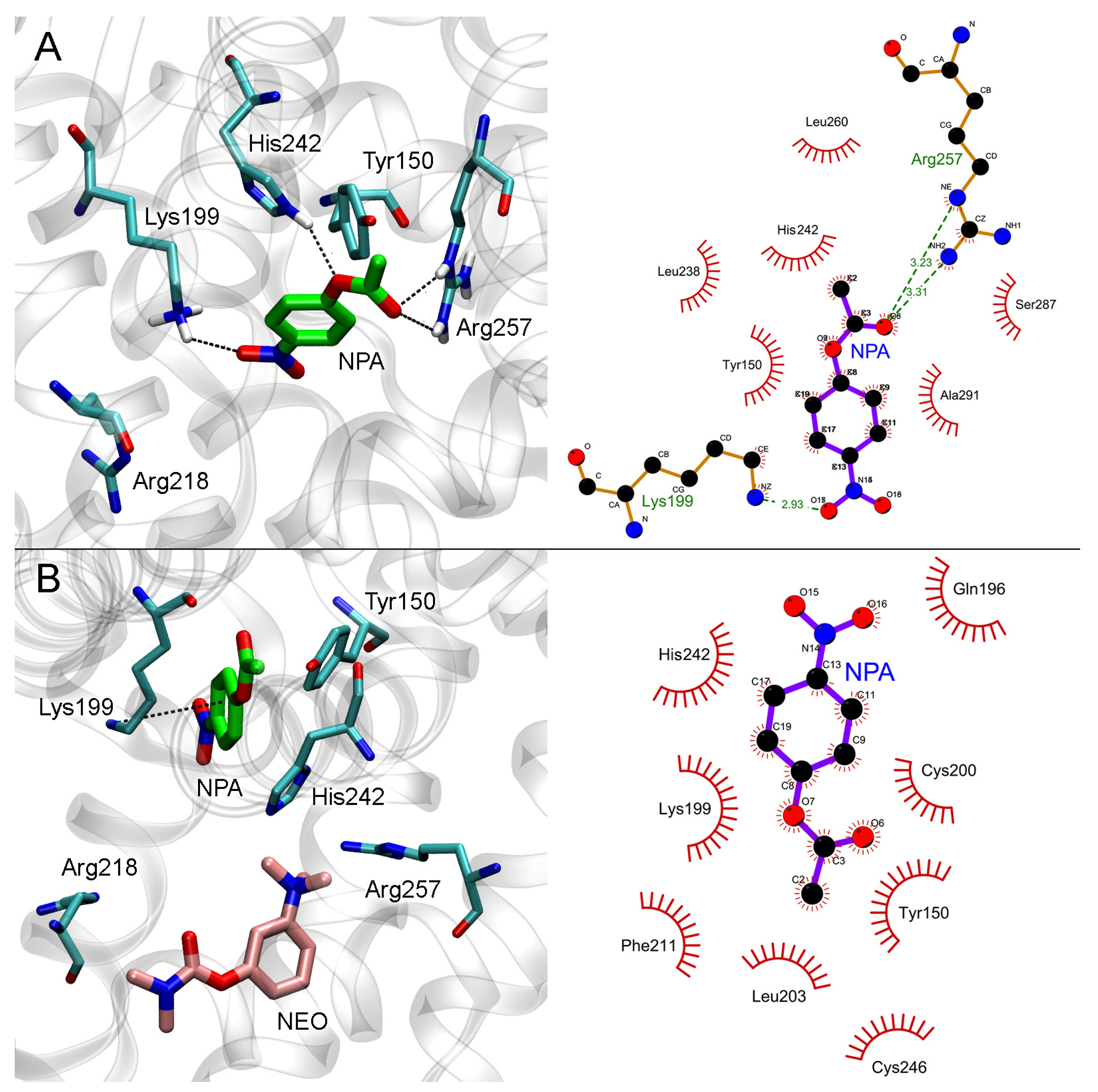
2.4. Interaction of Neostigmine with Human Serum Albumin According to Molecular Modeling Data
2.4.1. Interaction of Neostigmine with Sudlow Sites I and II of HSA
2.4.2. Effect of Neostigmine on the Interaction of NPA with Sudlow Site I of HSA
2.4.3. Effect of Neostigmine on the Interaction of NPA with Sudlow Site II of HSA
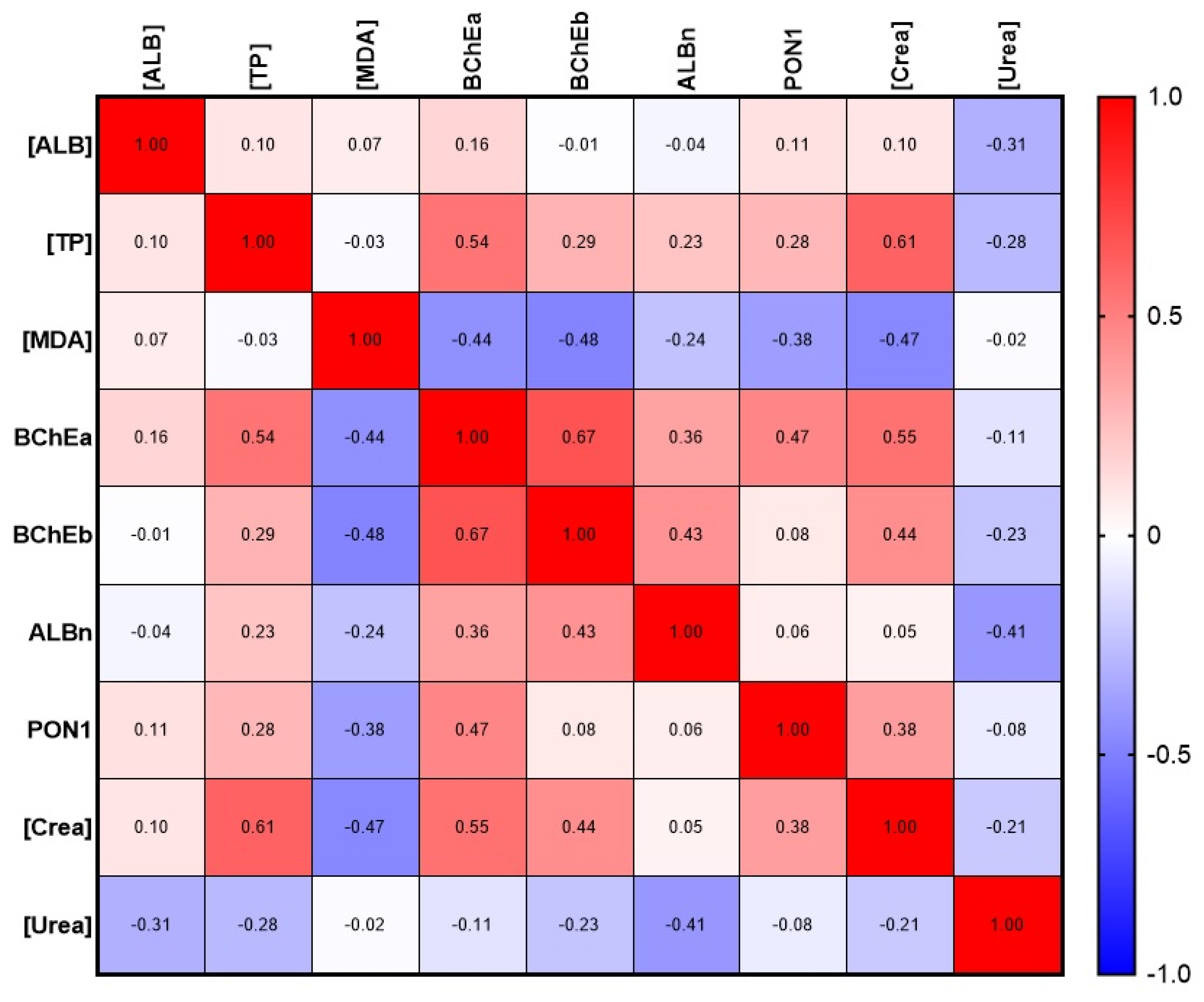
2.5. Esterase Status of Human Plasma in Normal and Pathological Conditions
2.5.1. Esterase Status of Healthy Subjects: Selection of the Control Group
2.5.2. Comparative Analysis of Esterase Status of Survivors and Deceased Patients with COVID-19
2.5.3. Comparative Analysis of Esterase Status of Healthy Subjects and Patients with COVID-19
3. Methods
3.1. Chemicals
3.2. Patients
3.3. Assay Methods
3.4. Albumin Electrophoresis in Polyacrylamide Gel
3.5. Effect of Inhibitors on the Esterase Activity of Serum Albumin In Vitro According to Spectrophotometry Data
3.6. Effect of Inhibitors on the Esterase Activity of Serum Albumin In Vitro According to NMR Data
3.7. Molecular Modeling Methods
3.7.1. Three-Dimensional Models Preparation
3.7.2. Molecular Docking
3.7.3. Molecular Dynamics
3.8. Statistics
4. Conclusions
5. Limitations of the Research
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AChE | acetylcholinesterase |
| ALB | concentration of albumin |
| ALBn | activity of albumin towards p-nitrophenyl acetate |
| ATCh | acetylthiocholine |
| BChE | butyrylcholinesterase |
| BChEa | activity of BChE towards acetylthiocholine |
| BChEb | activity of BChE towards butyrylthiocholine |
| BMI | body mass index |
| BSA | bovine serum albumin |
| BTCh | butyrylthiocholine |
| BW284C51 | 4-(5-{4-[Dimethyl(prop-2-enyl)ammonio]phenyl}-3-oxopentyl)-N,N-dimethyl-N-prop-2-enylbenzenaminium |
| CBDP | 2-(2-cresyl)-4H-1-3-2- benzodioxaphosphorin-2-oxide |
| CE | carboxylesterase |
| CK_MB | creatine kinase of myocardial band |
| CK_NAC | NAC-activated creatine kinase |
| Crea | creatinine |
| CRP | C-reactive protein |
| DMSO | dimethyl sulfoxide |
| DTNB | 5,5′-dithiobis-(2-nitrobenzoic acid) |
| DTT | dithiothreitol |
| EDTA | ethylenediaminetetraacetate |
| ESD | esterase D |
| FA1-7 | fatty-acid binding sites 1-7 in albumin |
| FRUC | fructosamine |
| GHSR1 | growth hormone secretagogue receptor 1 |
| HSA | human serum albumin |
| LDL | low-density lipoproteins |
| LINCS | linear constraint solver |
| MD | molecular dynamics |
| MDA | malondialdehyde |
| NEFA | non-esterified fatty acids |
| NEO | neostigmine |
| NMR | nuclear magnetic resonance |
| NPA | p-nitrophenyl acetate |
| NTE | neuropathy target esterase |
| NVT and NPT ensembles | constant volume and pressure ensembles |
| OPs | organophosphates |
| PDB | Protein Data Bank |
| PhA | phenyl acetate |
| PON1 | paraoxonase 1 |
| POX | paraoxon |
| SDS | sodium dodecyl sulfate |
| SFA | subcutaneous fat area |
| TP | total protein |
| TRIGS | triglycerides |
| Urea | urea |
| VFA | visceral fat area |
References
- Sokolovskaya, L.G.; Sigolaeva, L.V.; Eremenko, A.V.; Kurochkin, I.N.; Makhaeva, G.F.; Malygin, V.V.; Zykova, I.E.; Kholstov, V.I.; Zavyalova, N.V. Family of biosensor analyzers for assessment of “esterase status” of organism. Chem. Biol. Def. 2004, 1–2, 21–31. (In Russian) [Google Scholar]
- Makhaeva, G.; Rudakova, E.; Richardson, R. Investigation of the Esterase Status as a Complex Biomarker of Exposure to Organophosphorus Compounds. Biomed. Chem. Res. Methods 2018, 1, e00028. [Google Scholar] [CrossRef]
- Hajiasgharzadeh, K.; Khabbazi, A.; Mokhtarzadeh, A.; Baghbanzadeh, A.; Asadzadeh, Z.; Adlravan, E.; Baradaran, B. Cholinergic anti-inflammatory pathway and connective tissue diseases. Inflammopharmacology 2021, 29, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Piovesana, R.; Salazar Intriago, M.S.; Dini, L.; Tata, A.M. Cholinergic Modulation of Neuroinflammation: Focus on α7 Nicotinic Receptor. Int. J. Mol. Sci. 2021, 22, 4912. [Google Scholar] [CrossRef] [PubMed]
- Simões, J.L.B.; Sobierai, L.D.; Leal, I.F.; Dos Santos, M.V.R.; Victor Coiado, J.; Bagatini, M.D. Action of the purinergic and cholinergic anti-inflammatory pathways on oxidative stress in patients with Alzheimer’s disease in the context of the COVID-19 pandemic. Neuroscience 2022, 512, 110–132. [Google Scholar] [CrossRef]
- Li, B.; Sedlacek, M.; Manoharan, I.; Boopathy, R.; Duysen, E.G.; Masson, P.; Lockridge, O. Butyrylcholinesterase, paraoxonase, and albumin esterase, but not carboxylesterase, are present in human plasma. Biochem. Pharmacol. 2005, 70, 1673–1684. [Google Scholar] [CrossRef]
- Kou, Y.; Zou, L.; Liu, R.; Zhao, X.; Wang, Y.; Zhang, C.; Dong, Z.; Kou, J.; Bi, Y.; Fu, L.; et al. Intravascular cells and circulating microparticles induce procoagulant activity via phosphatidylserine exposure in heart failure. J. Thromb. Thrombolysis 2019, 48, 187–194. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Goncharov, N.V. Theoretical and practical aspects of albumin esterase activity. Russ. J. Bioorg. Chem. 2020, 46, 287–298. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Voronina, P.A.; Vovk, M.A.; Shmurak, V.I.; Batalova, A.A.; Jenkins, R.O.; Goncharov, N.V. Esterase Activity of Serum Albumin Studied by 1H NMR Spectroscopy and Molecular Modelling. Int. J. Mol. Sci. 2021, 22, 10593. [Google Scholar] [CrossRef]
- Grigoryan, H.; Li, H.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Cys34 adducts of reactive oxygen species in human serum albumin. Chem. Res. Toxicol. 2012, 25, 1633–1642. [Google Scholar] [CrossRef]
- Di Simplicio, P.; Frosali, S.; Priora, R.; Summa, D.; Cherubini Di Simplicio, F.; Di Giuseppe, D.; Di Stefano, A. Biochemical and biological aspects of protein thiolation in cells and plasma. Antioxid. Redox. Signal. 2005, 7, 951–963. [Google Scholar] [CrossRef]
- Nagumo, K.; Tanaka, M.; Chuang, V.T.; Setoyama, H.; Watanabe, H.; Yamada, N.; Kubota, K.; Tanaka, M.; Matsushita, K.; Yoshida, A.; et al. Cys34-Cysteinylated Human Serum Albumin Is a Sensitive Plasma Marker in Oxidative Stress-Related Chronic Diseases. PLoS ONE 2014, 9, e85216. [Google Scholar] [CrossRef] [PubMed]
- Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Vovk, M.A.; Batalova, A.A.; Jenkins, R.O.; Goncharov, N.V. The Universal Soldier: Enzymatic and Non-Enzymatic Antioxidant Functions of Serum Albumin. Antioxidants 2020, 9, 966. [Google Scholar] [CrossRef] [PubMed]
- Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Jenkins, R.O.; Goncharov, N.V. Serum Albumin in Health and Disease: Esterase, Antioxidant, Transporting and Signaling Properties. Int. J. Mol. Sci. 2021, 22, 10318. [Google Scholar] [CrossRef] [PubMed]
- Rahmani-Kukia, N.; Abbasi, A.; Pakravan, N.; Hassan, Z.M. Measurement of oxidized albumin: An opportunity for diagnoses or treatment of COVID-19. Bioorg. Chem. 2020, 105, 104429. [Google Scholar] [CrossRef]
- Tabata, F.; Wada, Y.; Kawakami, S.; Miyaji, K. Serum Albumin Redox States: More Than Oxidative Stress Biomarker. Antioxidants 2021, 10, 503. [Google Scholar] [CrossRef]
- Kovalic, A.J.; Huang, G.; Thuluvath, P.J.; Satapathy, S.K. Elevated Liver Biochemistries in Hospitalized Chinese Patients With Severe COVID-19: Systematic Review and Meta-analysis. Hepatology 2021, 73, 1521–1530. [Google Scholar] [CrossRef]
- Wybranowski, T.; Napiórkowska, M.; Bosek, M.; Pyskir, J.; Ziomkowska, B.; Cyrankiewicz, M.; Pyskir, M.; Pilaczyńska-Cemel, M.; Rogańska, M.; Kruszewski, S.; et al. Study of Albumin Oxidation in COVID-19 Pneumonia Patients: Possible Mechanisms and Consequences. Int. J. Mol. Sci. 2022, 23, 10103. [Google Scholar] [CrossRef]
- Lu, W.; Yu, S.; Liu, H.; Suo, L.; Tang, K.; Hu, J.; Shi, Y.; Hu, K. Survival Analysis and Risk Factors in COVID-19 Patients. Disaster Med. Public Health Prep. 2022, 16, 1916–1921. [Google Scholar] [CrossRef]
- Deng, X.; Li, H.; Liao, X.; Qin, Z.; Xu, F.; Friedman, S.; Ma, G.; Ye, K.; Lin, S. Building a predictive model to identify clinical indicators for COVID-19 using machine learning method. Med. Biol. Eng. Comput. 2022, 60, 1763–1774. [Google Scholar] [CrossRef]
- Saini, R.K.; Saini, N.; Ram, S.; Soni, S.L.; Suri, V.; Malhotra, P.; Kaur, J.; Verma, I.; Sharma, S.; Zohmangaihi, D. COVID-19 associated variations in liver function parameters: A retrospective study. Postgrad. Med. J. 2022, 98, 91–97. [Google Scholar] [CrossRef]
- Al-Shajlawi, M.; Alsayed, A.R.; Abazid, H.; Awajan, D.; Al-Imam, A.; Basheti, I. Using laboratory parameters as predictors for the severity and mortality of COVID-19 in hospitalized patients. Pharm. Pract. 2022, 20, 2721. [Google Scholar] [CrossRef]
- Ascenzi, P.; Fasano, M. Allostery in a monomeric protein: The case of human serum albumin. Biophys. Chem. 2010, 48, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Sogorb, M.A.; García-Argüelles, S.; Carrera, V.; Vilanova, E. Serum albumin is as efficient as paraxonase in the detoxication of paraoxon at toxicologically relevant concentrations. Chem. Res. Toxicol. 2008, 21, 1524–1529. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.; Xie, W.; Froment, M.T.; Lockridge, O. Effects of mutations of active site residues and amino acids interacting with the Omega loop on substrate activation of butyrylcholinesterase. Biochim. Biophys. Acta 2001, 1544, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Chiou, S.Y.; Wu, Y.G.; Lin, Y.F.; Lin, L.Y.; Lin, G. Substrate activation of butyrylcholinesterase and substrate inhibition of acetylcholinesterase by 3,3-dimethylbutyl-N-n-butylcarbamate and 2-trimethylsilyl-ethyl-N-n-butylcarbamate. J. Biochem. Mol. Toxicol. 2007, 21, 24–31. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Voronina, P.A.; Batalova, A.A.; Goncharov, N.V. Serum Albumin. Encyclopedia 2021, 1, 65–75. [Google Scholar] [CrossRef]
- Lockridge, O.; Quinn, D.M. Esterases. In Comprehensive Toxicology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 4, pp. 243–273. [Google Scholar]
- Pang, H.; Ding, Y.; Li, Y.; Wang, L.; Tian, X.; Wang, B.; Ding, M. Mismatched multiplex PCR amplification and subsequent RFLP analysis to simultaneously identify polymorphisms of erythrocytic ESD, GLO1, and GPT genes. J. Forensic. Sci. 2011, 56 (Suppl. S1), S176–S178. [Google Scholar] [CrossRef]
- Draganov, D.I.; Teiber, J.F.; Speelman, A.; Osawa, Y.; Sunahara, R.; La Du, B.N. Human paraoxonases (PON1, PON2, and PON3) are lactonases with overlapping and distinct substrate specificities. J. Lipid. Res. 2005, 46, 1239–1247. [Google Scholar] [CrossRef]
- Lockridge, O.; Xue, W.; Gaydess, A.; Grigoryan, H.; Ding, S.J.; Schopfer, L.M.; Hinrichs, S.H.; Masson, P. Pseudo-esterase activity of human albumin: Slow turnover on tyrosine 411 and stable acetylation of 82 residues including 59 lysines. J. Biol. Chem. 2008, 283, 22582–22590. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Terpilovskii, M.A.; Belinskaya, D.A.; Shmurak, V.I.; Avdonin, P.V. Comparative analysis of esterase and paraoxonase activities of different serum albumin species. J. Evol. Biochem. Physiol. 2017, 53, 271–281. [Google Scholar] [CrossRef]
- Blomberg, R.; Kries, H.; Pinkas, D.M.; Mittl, P.R.; Grütter, M.G.; Privett, H.K.; Mayo, S.L.; Hilvert, D. Precision is essential for efficient catalysis in an evolved Kemp eliminase. Nature 2013, 503, 418–421. [Google Scholar] [CrossRef]
- Richard, J.P.; Amyes, T.L.; Goryanova, B.; Zhai, X. Enzyme architecture: On the importance of being in a protein cage. Curr. Opin. Chem. Biol. 2014, 21, 1–10. [Google Scholar] [CrossRef]
- Moroz, Y.S.; Dunston, T.T.; Makhlynets, O.V.; Moroz, O.V.; Wu, Y.; Yoon, J.H.; Olsen, A.B.; McLaughlin, J.M.; Mack, K.L.; Gosavi, P.M.; et al. New Tricks for Old Proteins: Single Mutations in a Nonenzymatic Protein Give Rise to Various Enzymatic Activities. J. Am. Chem. Soc. 2015, 137, 14905–14911. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.L.; Møller, A.M.; Wildgaard, K. Butyrylcholinesterase deficiency and its clinical importance in anaesthesia: A systematic review. Anaesthesia 2019, 74, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Brimijoin, S.; Chen, V.P.; Pang, Y.P.; Geng, L.; Gao, Y. Physiological roles for butyrylcholinesterase: A BChE-ghrelin axis. Chem. Biol. Interact. 2016, 259 Pt B, 271–275. [Google Scholar] [CrossRef]
- Iwasaki, T.; Yoneda, M.; Nakajima, A.; Terauchi, Y. Serum butyrylcholinesterase is strongly associated with adiposity, the serum lipid profile and insulin resistance. Intern. Med. 2007, 46, 1633–1639. [Google Scholar] [CrossRef]
- Cucuianu, M.; Opincaru, A.; Tapalagă, D. Similar behaviour of lecithin:cholesterol acyltransferase and pseudocholinesterase in liver disease and hyperlipoproteinemia. Clin. Chim. Acta 1978, 85, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Abou-Hatab, K.; O’Mahony, M.S.; Patel, S.; Woodhouse, K. Relationship between age and plasma esterases. Age Ageing 2001, 30, 41–45. [Google Scholar] [CrossRef]
- Delacour, H.; Dedome, E.; Courcelle, S.; Hary, B.; Ceppa, F. Butyrylcholinesterase deficiency. Ann. Biol. Clin. 2016, 74, 279–285. [Google Scholar] [CrossRef]
- Sipahioglu, H.; Onuk, S. Lactate dehydrogenase/albumin ratio as a prognostic factor in severe acute respiratory distress syndrome cases associated with COVID-19. Medicine 2022, 101, 30759. [Google Scholar] [CrossRef]
- Sai, I.N.; Prasad, R.; Varsha, T.V. Assessing the Prognostic Value of Crp/Albumin Ratio and Lactate/Albumin Ratio in Critically Ill Patients. J. Assoc. Physicians India. 2022, 70, 11–12. [Google Scholar]
- Anzo, F.M.; Buan-Mayo, M. Nutritional biomarkers as predictors of clinical outcomes between COVID-19 severity groups in a tertiary government hospital. Clin. Nutr. ESPEN 2023, 53, 134–143. [Google Scholar] [CrossRef]
- Torun, A.; Çakırca, T.D.; Çakırca, G.; Portakal, R.D. The value of C-reactive protein/albumin, fibrinogen/albumin, and neutrophil/lymphocyte ratios in predicting the severity of COVID-19. Rev. Assoc. Med. Bras. (1992) 2021, 67, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Makkar, K.; Sharma, Y.P.; Batta, A.; Hatwal, J.; Panda, P.K. Role of fibrinogen, albumin and fibrinogen to albumin ratio in determining angiographic severity and outcomes in acute coronary syndrome. World J. Cardiol. 2023, 15, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Tosun, F.; Comert, E.; Duran, M.; Tuna, V.D. The Relationship of CRP/Albumin ratio level and prognosis in pregnant COVID-19 patients. Niger J. Clin. Pract. 2022, 25, 1745–1750. [Google Scholar] [PubMed]
- Kalyon, S.; Gültop, F.; Şimşek, F.; Adaş, M. Relationships of the neutrophil-lymphocyte and CRP-albumin ratios with the duration of hospitalization and fatality in geriatric patients with COVID-19. J. Int. Med. Res. 2021, 49, 3000605211046112. [Google Scholar] [CrossRef] [PubMed]
- Detsika, M.G.; Grigoriou, E.; Psarra, K.; Jahaj, E.; Tsipilis, S.; Athanassiou, N.; Zacharis, A.; Dimopoulou, I.; Orfanos, S.E.; Tsirogianni, A.; et al. Combination of the CD8+:B-cell and Neutrophil-to-Lymphocyte Ratio as a Novel Prediction Model for Intubation Need and Disease Severity in COVID-19 Patients. Vivo 2021, 35, 3305–3313. [Google Scholar] [CrossRef]
- Tocoglu, A.; Dheir, H.; Bektas, M.; Acikgoz, S.B.; Karabay, O.; Sipahi, S. Predictors of Mortality in Patients with COVID-19 Infection-associated Acute Kidney Injury. J. Coll. Physicians Surg. Pak. 2021, 30, S60–S65. [Google Scholar] [CrossRef]
- Feketea, G.M.; Vlacha, V. The Diagnostic Significance of Usual Biochemical Parameters in Coronavirus Disease 19 (COVID-19): Albumin to Globulin Ratio and CRP to Albumin Ratio. Front. Med. 2020, 7, 566591. [Google Scholar] [CrossRef]
- Afsin, D.E.; Kerget, B. Evaluation of the Relationship between CRP/Albumin Ratio and Pulmonary Function Parameters in Patients with Post-Acute COVID-19. Clin. Lab. 2022, 1, 68. [Google Scholar] [CrossRef]
- Ducastel, M.; Chenevier-Gobeaux, C.; Ballaa, Y.; Meritet, J.F.; Brack, M.; Chapuis, N.; Pene, F.; Carlier, N.; Szwebel, T.A.; Roche, N.; et al. Oxidative Stress and Inflammatory Biomarkers for the Prediction of Severity and ICU Admission in Unselected Patients Hospitalized with COVID-19. Int. J. Mol. Sci. 2021, 22, 7462. [Google Scholar] [CrossRef] [PubMed]
- Semiz, S. COVID19 biomarkers: What did we learn from systematic reviews? Front. Cell Infect. Microbiol. 2022, 12, 1038908. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Lucas, R.; Fulton, D. The subcellular compartmentalization of arginine metabolizing enzymes and their role in endothelial dysfunction. Front. Immunol. 2013, 4, 184. [Google Scholar] [CrossRef]
- Durante, W. Role of arginase in vessel wall remodeling. Front. Immunol. 2013, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Elms, S.; Chen, F.; Wang, Y.; Qian, J.; Askari, B.; Yu, Y.; Pandey, D.; Iddings, J.; Caldwell, R.B.; Fulton, D.J. Insights into the arginine paradox: Evidence against the importance of subcellular location of arginase and eNOS. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H651–H666. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, L.; Ren, Y.; Huang, Y.; Liu, W.; Lv, Z.; Qian, L.; Yu, Y.; Xiong, Y. Arginase: Shedding light on the mechanisms and opportunities in cardiovascular diseases. Cell Death Discov. 2022, 8, 413. [Google Scholar] [CrossRef]
- Durante, W. Targeting Arginine in COVID-19-Induced Immunopathology and Vasculopathy. Metabolites 2022, 12, 240. [Google Scholar] [CrossRef]
- Salazar, J.; Martínez, M.S.; Chávez-Castillo, M.; Núñez, V.; Añez, R.; Torres, Y.; Toledo, A.; Chacín, M.; Silva, C.; Pacheco, E.; et al. C-Reactive Protein: An In-Depth Look into Structure, Function, and Regulation. Int. Sch. Res. Not. 2014, 2014, 653045. [Google Scholar] [CrossRef]
- Barnes, P.W.; McFadden, S.L.; Machin, S.J.; Simson, E.; International Consensus Group for Hematology. The international consensus group for hematology review: Suggested criteria for action following automated CBC and WBC differential analysis. Lab. Hematol. 2005, 11, 83–90. [Google Scholar] [CrossRef]
- Collier, H.B. Letter: A note on the molar absorptivity of reduced Ellman’s reagent, 3-carboxylato-4-nitrothiophenolate. J. Anal. Biochem. 1973, 56, 310–311. [Google Scholar] [CrossRef]
- Aguilar, J.A.; Nilsson, M.; Bodenhausen, G.; Morris, G.A. Spin echo NMR spectra without J modulation. Chem. Commun. 2012, 48, 811–813. [Google Scholar] [CrossRef]
- Froimowitz, M. HyperChem: A software package for computational chemistry and molecular modeling. Biotechniques 1993, 14, 1010–1013. [Google Scholar]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural basis of the drug-binding specificity of human serum albumin. J. Mol. Biol. 2005, 353, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Mackerell, A.D., Jr.; Feig, M.; Brooks, C.L. 3rd. Extending the treatment of backbone energetics in protein force fields: Limitations of gas-phase quantum mechanics in reproducing protein conformational distributions in molecular dynamics simulations. J. Comput. Chem. 2004, 25, 1400–1415. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L. Quantum and statistical mechanical studies of liquids. 10. Transferable intermolecular potential functions for water, alcohols, and ethers. Application to liquid water. J. Am. Chem. Soc. 1981, 103, 335–340. [Google Scholar] [CrossRef]
- Bussi, G.; Zykova-Timan, T.; Parrinello, M. Isothermal-isobaric molecular dynamics using stochastic velocity rescaling. J. Chem. Phys. 2009, 130, 074101. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 3, 10089–10092. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comp. Chem. 1997, 18, 1463–1473. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group 1 “Elderlies” | Control Group 2 “Swimmers” | |
|---|---|---|
| Number of samples | 9 | 12 |
| [ALB], g L−1 | 45.2 (43.7; 48.9) | 45.7 (42.5; 51.4) |
| [Urea], mM | 5.8 (4.4; 9.7) | 4.5 (2.9; 7.8) |
| [Creatinine], µM | 57.8 (49.4; 62.3) | 82.5 (68.0; 104.0) **** |
| [TP], g L−1 | 73.5 (67.9; 80.2) | 79.7 (73.2; 86.7) ** |
| [MDA], µM | 1.10 (0.60; 1.30) | 0.80 (0.33; 2.30) * |
| BChEa, µM min−1 | 1272 (1015; 1602) | 1572 (1189; 2051) * |
| BChEb, µM min−1 | 2182 (1270; 3358) | 3060 (2274; 3953) ** |
| ALBn, µM min−1 | 37.2 (21.7; 70.9) | 47.8 (6.0; 186.2) |
| PON1, mM min−1 | 40.0 (24.6; 51.0) | 44.2 (28.9; 55.3) |
| Indices | ||
| [ALB]/[TP] | 0.62 (0.59; 0.66) | 0.58 (0.50; 0.67) * |
| [Urea] × 10/[ALB] | 1.34 (0.93; 2.14) | 0.95 (0.58; 1.71) |
| [MDA]/[ALB] | 2.53 (1.41; 2.94) | 1.72 (0.73; 5.27) |
| BChEa/[ALB] | 28.2 (22.5; 32.7) | 34.6 (27.1; 46.6) ** |
| BChEb/[ALB] | 48.8 (26.0; 74.5) | 66.7 (52.0; 87.9) ** |
| [Creatinine] × [MDA]/[ALB] | 1.29 (0.81; 1.81) | 1.40 (0.76; 3.58) |
| [Urea] × [MDA]/[ALB] | 0.15 (0.09; 0.21) | 0.08 (0.03; 0.23) * |
| BChEa/[MDA] | 1144 (870; 1999) | 1987 (516; 4024) ** |
| BChEb/[MDA] | 1908 (1255; 4282) | 3789 (987; 7265) * |
| BChEa/ALBn | 28.5 (17.9; 61.6) | 23.8 (10.3; 110.7) |
| BChEb/ALBn | 45.2 (35.2; 154.5) | 45.3 (18.8; 199.8) |
| [Urea] × [MDA] × 1000/(BChEb × [ALB]) | 0.06 (0.04; 0.12) | 0.02 (0.01; 0.10) ** |
| Parameter | Survivors | Non-Survivors |
|---|---|---|
| Number of samples, n | 19 | 10 |
| [ALB], g L−1 | 37.0 (29.0; 43.0) | 32.7 (25.0; 38.9) * |
| [TP], g L−1 | 67.7 (58.8; 82.1) | 65.1 (48.9; 82.2) |
| [ALB]/[TP] | 0.54 (0.42; 0.66) | 0.52 (0.40; 0.58) |
| [MDA], μM | 3.59 (0.74; 7.87) | 5.72 (2.12; 8.37) |
| BChEa, μM min−1 | 1177 (798; 1528) | 832 (350; 1535) * |
| BChEb, μM min−1 | 2435 (1515; 2890) | 1471 (850; 2372) **** |
| ALBn, μM min−1 | 48.3 (6.0; 113.3) | 43.0 (6.0; 223.5) |
| PON1, mM min−1 | 25.8 (1.3; 48.6) | 25.9 (11.8; 32.9) |
| Indices | ||
| [Urea] × 10/[ALB] | 1.29 (0.91; 3.44) | 5.40 (1.62; 20.20) **** |
| [MDA]/[ALB] | 10.98 (2.00; 25.56) | 18.78 (7.29; 27.14) * |
| BChEa/[ALB] | 30.73 (22.61; 46.85) | 24.90 (12.38; 39.47) |
| BChEb/[ALB] | 64.01 (40.94; 89.35) | 46.85 (25.51; 60.97) *** |
| [Creatinine] × [MDA]/[ALB] | 11.06 (1.36; 26.25) | 19.37 (2.14; 53.08) |
| [Urea] × [MDA]/[ALB] | 0.42 (0.07; 1.92) | 2.48 (0.54; 13.71) *** |
| BChEa/[MDA] | 293 (101; 1741) | 161 (66; 374) * |
| BChEb/[MDA] | 571 (251; 3295) | 261 (195; 590) *** |
| [Urea] × [MDA] × 1000/(BChEb × [ALB]) | 0.18 (0.03; 0.96) | 1.60 (0.56; 10.36) **** |
| Control Group | Survivors | Non-Survivors | |
|---|---|---|---|
| Number of samples | 9 | 19 | 10 |
| [ALB], g L−1 | 45.2 (43.7; 48.9) | 37.0 (29.0; 43.0) *** | 32.7 (25.0; 38.9) **** |
| [TP], g L−1 | 73.5 (67.9; 80.2) | 67.7 (58.8; 82.1) | 65.1 (48.9; 82.2) * |
| [MDA], μM | 1.1 (0.6; 1.3) | 3.59 (0.74; 7.87) * | 5.72 (2.12; 8.37) *** |
| BChEa, μM min−1 | 1272 (1015; 1602) | 1177 (798; 1528) | 832 (350; 1535) * # |
| BChEb, μM min−1 | 2182 (1270; 3358) | 2435 (1515; 2890) | 1471 (850; 2372) * ### |
| ALBn, μM min−1 | 37.2 (21.7; 70.9) | 48.3 (6.0; 113.3) | 43.0 (6.0; 223.5) |
| PON1, mM min−1 | 40.0 (24.6; 51.0) | 25.8 (1.28; 48.6) ** | 25.9 (11.8; 32.6) * |
| [Urea] × 10/[ALB] | 1.34 (0.93; 2.14) | 1.29 (0.91; 3.44) | 5.4 (1.62; 20.2) **, ### |
| [Urea] × [MDA] × 1000/(BChEb × [ALB]) | 0.06 (0.04; 0.12) | 0.19 (0.03; 0.96) | 1.60 (0.56; 10.36) ****, ## |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belinskaia, D.A.; Voronina, P.A.; Popova, P.I.; Voitenko, N.G.; Shmurak, V.I.; Vovk, M.A.; Baranova, T.I.; Batalova, A.A.; Korf, E.A.; Avdonin, P.V.; et al. Albumin Is a Component of the Esterase Status of Human Blood Plasma. Int. J. Mol. Sci. 2023, 24, 10383. https://doi.org/10.3390/ijms241210383
Belinskaia DA, Voronina PA, Popova PI, Voitenko NG, Shmurak VI, Vovk MA, Baranova TI, Batalova AA, Korf EA, Avdonin PV, et al. Albumin Is a Component of the Esterase Status of Human Blood Plasma. International Journal of Molecular Sciences. 2023; 24(12):10383. https://doi.org/10.3390/ijms241210383
Chicago/Turabian StyleBelinskaia, Daria A., Polina A. Voronina, Polina I. Popova, Natalia G. Voitenko, Vladimir I. Shmurak, Mikhail A. Vovk, Tatiana I. Baranova, Anastasia A. Batalova, Ekaterina A. Korf, Pavel V. Avdonin, and et al. 2023. "Albumin Is a Component of the Esterase Status of Human Blood Plasma" International Journal of Molecular Sciences 24, no. 12: 10383. https://doi.org/10.3390/ijms241210383
APA StyleBelinskaia, D. A., Voronina, P. A., Popova, P. I., Voitenko, N. G., Shmurak, V. I., Vovk, M. A., Baranova, T. I., Batalova, A. A., Korf, E. A., Avdonin, P. V., Jenkins, R. O., & Goncharov, N. V. (2023). Albumin Is a Component of the Esterase Status of Human Blood Plasma. International Journal of Molecular Sciences, 24(12), 10383. https://doi.org/10.3390/ijms241210383