
The Potentiality of Plant-Derived Nanovesicles in Human Health—A Comparison with Human Exosomes and Artificial Nanoparticles



Abstract
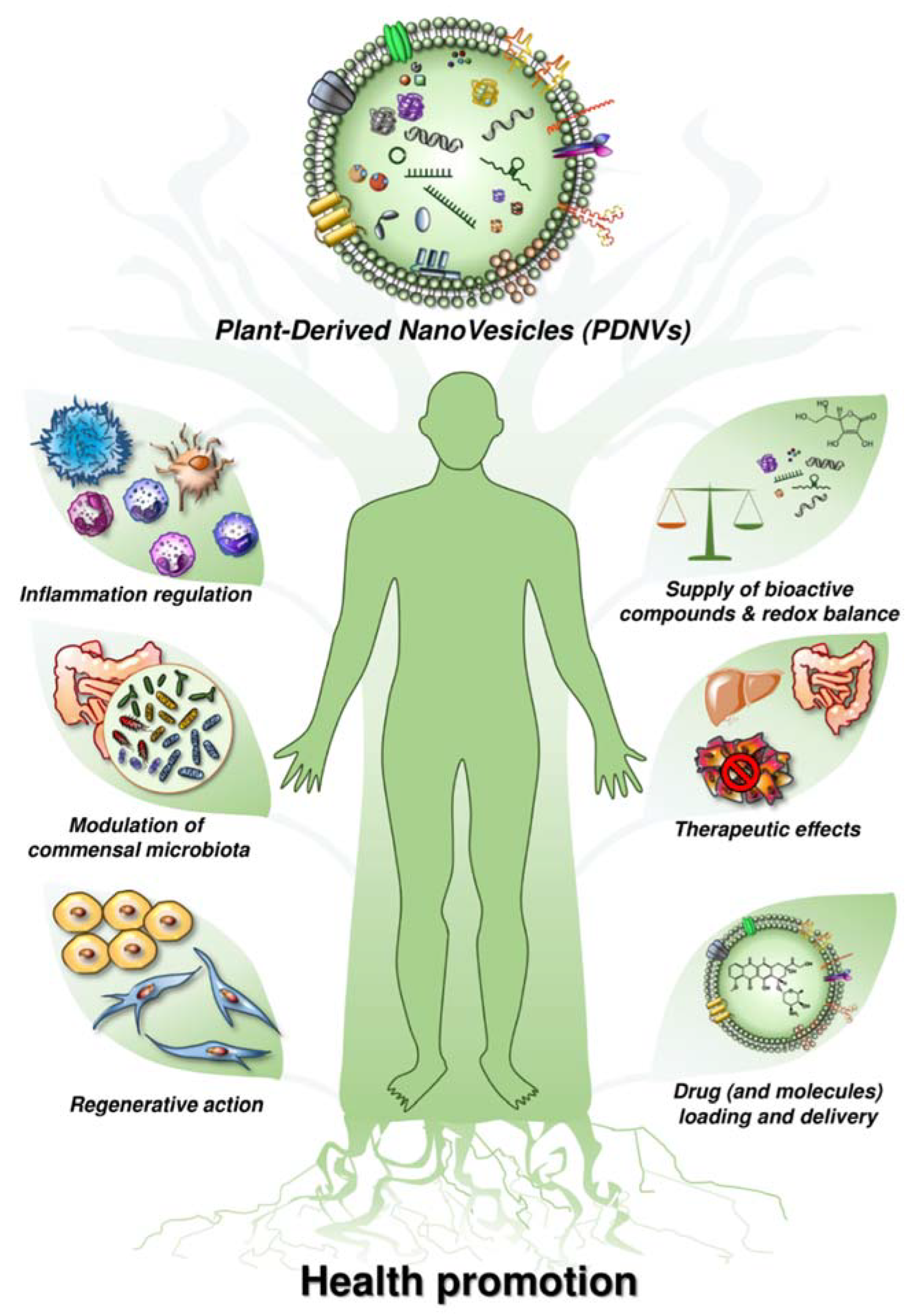
:1. Introduction
2. Comparing Human Exosomes to Plant Nanovesicles
3. The Role of Plant-Derived Nanovesicles in Supplementation and Drug Delivery
3.1. The Role of Plant-Derived Nanovesicles in Future Approach of Supplementation
3.2. The Role of Plant-Derived Nanovesicles in Drug Delivery

{kind=link}
| Source | Derivation | Application | Reference |
|---|---|---|---|
| Aloe vera (Aloe vera barbadensis) | Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] |
| Apple (Malus domestica) | Non-specified | Apple nanovesicles exert a potent anti-inflammatory effect in vitro | [64] |
| Arabidopsis (Arabidopsis thaliana) | Non-specified | EVs secretion is enhanced during biotic stress and EVs proteome changes in response to P. syringae infection | [32] |
| Blueberry (Vaccinium myrtillus, Vaccinium corymbosum) | Non-specified | Characterization of small RNAs in nanovesicles; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Broccoli (Brassica oleracea) | Non-specified | Broccoli-derived nanoparticles inhibit colitis in treated mice | [8] |
| Cactus (Cactus) | Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] |
| Carrot (Daucus carota) | Non-specified | Isolated vesicles are taken up by intestinal macrophages; vesicles induce Nrf2 expression | [76] |
| Non-specified | miRNAs of plant-derived nanovesicles influence microbiota composition | [77] | |
| Non-specified | Nanovesicles isolated from carrots exert potent anti-oxidative and apoptotic effects in in vitro cardiomyoblasts and neuroblastoma cell lines | [19] | |
| Cilantro (Coriandrum sativum) | Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] |
| Clementine (Citrus clementina) | Non-specified | Characterization of membrane transporters in nanovesicles derived from clementine juice | [36] |
| Coconut (Cocos nucifera) | Non-specified | Characterization of small RNAs in nanovesicles; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Non-specified | Characterization of miRNAs in extracellular vesicles isolated from immature coconut water and mature coconut water | [78] | |
| Cucumber (Cucumis sativus) | Non-specified | Characterization of cucumber-derived exosome-like vesicles | [79] |
| Dandelion (Taraxacum) | Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] |
| Garlic (Allium sativum) | Non-specified | miRNAs of plant-derived nanovesicles influence microbiota composition | [77] |
| Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] | |
| Ginger (Zingiber officinale) | Non-specified | Isolated vesicles are taken up by intestinal macrophages; ginger-derived vesicles induce heme oxygenase-1 and IL-10 expression | [76] |
| Non-specified | Ginger-derived vesicles protect treated mouse from alcohol-induced liver damage | [80] | |
| Non-specified | Lipid re-assembled ginger nanovesicles are loaded with doxorubicin; loaded nanovesicles exert apoptotic effects in vitro and reduce tumor growth in vivo | [16] | |
| Non-specified | Nanovesicles from ginger are efficiently internalized in colon cells after oral administration in treated mice; vesicles prevent and treat chronic colitis and colitis-associated cancer | [47] | |
| Non-specified | Ginger-derived lipid vehicles loaded with siRNA-CD98 are taken up by colon cells and efficiently decrease CD98 expression in vitro and in vivo | [67] | |
| Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] | |
| Non-specified | miRNAs of plant-derived nanovesicles influence microbiota composition | [77] | |
| Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] | |
| Non-specified | Ginger-derived nanovesicles prevent and treat periodontitis in vitro and in vivo | [27] | |
| Ginseng (Panax ginseng) | Non-specified | Ginseng-derived nanovesicles improved the replicative senescent or senescence-associated pigmented phenotypes of human dermal fibroblasts or ultraviolet B radiation-treated human melanocyte | [81] |
| Grape (Vitis vinifera) | Non-specified | Grape exosome-like nanoparticles penetrate intestinal mucus barrier and protect mice from dextran sulfate sodium (DSS)-induced colitis | [17] |
| Non-specified | Isolated vesicles are taken up by intestinal macrophages; vesicles induce Nrf2 expression | [76] | |
| Non-specified | Characterization of grape-derived vesicles | [82] | |
| Grapefruit (Citrus paradisi) | Non-specified | Grapefruit-derived nanovector inhibit tumor growth in vivo | [42] |
| Non-specified | Isolated vesicles are taken up by intestinal macrophages; vesicles induce Nrf2 expression | [76] | |
| Non-specified | Grapefruit-derived nanovesicles are taken up by intestinal macrophages and ameliorate dextran sulfate sodium (DSS)-induced mouse colitis | [40] | |
| Non-specified | Intranasally administrated grapefruit nanovesicles deliver miR17 to mice brain tumors | [48] | |
| Non-specified | Grapefruit-derived lipids carrying miR18 inhibit liver metastasis through induction of M1 macrophages | [37] | |
| Non-specified | Grapefruit-derived nanovector coated with inflammatory-related receptor enriched membranes of activated leukocytes (IGNVs) are enhanced for homing in on inflammatory tumor tissues | [41] | |
| Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] | |
| Non-specified | miRNAs of plant-derived nanovesicles influence microbiota composition | [77] | |
| Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophage and inhibit activation of NLRP3 inflammasome | [74] | |
| Organic agriculture | Nanovesicles from organic agriculture show a higher anti-oxidant level compared to nanovesicles from conventional agriculture | [30] | |
| Kiwi (Actinidia chinensis) | Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Organic agriculture | Nanovesicles from organic agriculture show a higher anti-oxidant level compared to nanovesicles from conventional agriculture | [30] | |
| Lavender (Lavandula) | Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] |
| Lemon (Citrus limon) | Non-specified | Lemon-derived nanovesicles inhibits tumor cell proliferation in vitro and tumor growth in vivo | [21] |
| Non-specified | Nanovesicles isolated from lemon exert a significant protective effect against oxidative stress | [29] | |
| Organic agriculture | Nanovesicles from organic agriculture show a higher anti-oxidant level compared to nanovesicles from conventional agriculture | [30] | |
| Mistletoe (Viscum album) | Non-specified | Vesicles are highly stable and overcome the drying process of plant material | [83] |
| Orange (Citrus sinensis) | Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Orange (Citrus sinensis ‘Blood Orange’) | Organic agriculture | Nanovesicles from organic agriculture show a higher anti-oxidant level compared to nanovesicles from conventional agriculture | [72] |
| Pear (Pyrus communis) | Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Peas (Pisum sativum) | Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Periwinkle (Vinca minor) | Non-specified | Vesicles are highly stable and overcome the drying process of plant material | [83] |
| Soybean (Glycine soja) | Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Strawberry (Fragaria ananassa) | Non-specified | Nanovesicles from strawberry are internalized by human mesenchymal stromal cells and prevent oxidative stress | [84] |
| Sunflower (Helianthus annuus) | Non-specified | Characterization of Rab proteins in isolated vesicles | [5] |
| Tobacco (Nicotiana tabacum) | Non-specified | Vesicles are highly stable and overcome the drying process of plant material | [83] |
| Tomato (Lycopersicon esculentum) | Non-specified | Characterization of small RNAs; miRNAs regulate the expression of inflammatory cytokines and cancer-related genes in vitro | [75] |
| Tomato (Lycopersicon esculentum ‘Piccadilly’) | Non-specified | Characterization of an improved isolation method | [85] |
| Turmeric (Curcuma longa) | Non-specified | miRNAs of plant-derived nanovesicles influence microbiota composition | [77] |
| Non-specified | Vesicles are efficiently taken up by bone marrow-derived macrophages and inhibit activation of NLRP3 inflammasome | [74] | |
| Watermelon (Citrullus lanatus) | Non-specified | Characterization of watermelon EVs | [86] |
4. Comments and Discussion
- Micro/nanovesicles can be obtained from a wide range of fruits and vegetables, and are better if from organic agriculture. Citrus fruits have been by far the more investigated in this area, and we have data that include Citrus limon, Citrus paradisi, Citrus medica, Citrus reticulata, Citrus sinensis (both yellow and red oranges), Citrus bergamia, Citrus clementina, and Citrus monstruosa.
- It is possible to perform quality control of any preparation by using standard methods established from the international community in the field (e.g., nanoparticle tracking analysis and immunocapture-based technology).
- FVNVs contain measurable levels of a series of anti-oxidants, including ascorbic acid (Vit. C), glutathione, superoxide dismutase-1 (SOD-1) and catalase.
- The anti-oxidants are complexed in a nanostructure that can be called the nano-phyto complex.
- Due to their structure, FVNVs are stable and contain fully active molecules, showing a clear anti-oxidant effect against both normal and tumor cells.
- FVNVs may deliver drugs of various origins to target cells, showing significantly higher effects than free molecules.
- FVNVs can be well used instead of artificial nanoparticles (i.e., liposomes). Compared to the currently available drug delivery systems, plant-derived nanovesicles have multiple advantages, such as low immunogenicity and stability in the gastrointestinal tract (Yang et al., 2018).
- FVNVs possess high biocompatibility and promises large-scale production.
- More and more evidence suggests that FVNVs can enter mammalian cells and mediate plant–animal cross-kingdom gene regulation. This evidence show that plant small RNAs packed by plant-derived nanovesicles could survive in their active forms in animals and exogenously modulate the host cellular processes via genetic crosstalk [63]. Moreover, plant-derived nanovesicles have exhibited promising activities in the homeostatic regulation of the immune system, development of tissue engineering and reconstruction, delivery of chemotherapeutic drugs and nucleic acids, etc. [17,43,47,63,66].
- The above evidence indicates a potential medical application of plant-derived nanovesicles in the regulation of the fundamental biological processes in the human body.
Author Contributions
Funding
Conflicts of Interest
References
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fais, S.; O’Driscoll, L.; Borras, F.E.; Buzas, E.; Camussi, G.; Cappello, F.; Carvalho, J.; Cordeiro da Silva, A.; Del Portillo, H.; El Andaloussi, S.; et al. Evidence-Based Clinical Use of Nanoscale Extracellular Vesicles in Nanomedicine. ACS Nano 2016, 10, 3886–3899. [Google Scholar] [CrossRef] [PubMed]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying extracellular vesicles based therapeutics in clinical trials—An ISEV position paper. J. Extracell. Vesicles 2015, 4, 30087. [Google Scholar] [CrossRef] [PubMed]
- Karamanidou, T.; Tsouknidas, A. Plant-Derived Extracellular Vesicles as Therapeutic Nanocarriers. Int. J. Mol. Sci. 2021, 23, 191. [Google Scholar] [CrossRef]
- Regente, M.; Corti-Monzón, G.; Maldonado, A.M.; Pinedo, M.; Jorrín, J.; de la Canal, L. Vesicular fractions of sunflower apoplastic fluids are associated with potential exosome marker proteins. FEBS Lett. 2009, 583, 3363–3366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfieri, M.; Leone, A.; Ambrosone, A. Plant-Derived Nano and Microvesicles for Human Health and Therapeutic Potential in Nanomedicine. Pharmaceutics 2021, 13, 498. [Google Scholar] [CrossRef] [PubMed]
- Dad, H.A.; Gu, T.-W.; Zhu, A.-Q.; Huang, L.-Q.; Peng, L.-H. Plant Exosome-like Nanovesicles: Emerging Therapeutics and Drug Delivery Nanoplatforms. Mol. Ther. 2021, 29, 13–31. [Google Scholar] [CrossRef]
- Deng, Z.; Rong, Y.; Teng, Y.; Mu, J.; Zhuang, X.; Tseng, M.; Samykutty, A.; Zhang, L.; Yan, J.; Miller, D.; et al. Broccoli-Derived Nanoparticle Inhibits Mouse Colitis by Activating Dendritic Cell AMP-Activated Protein Kinase. Mol. Ther. 2017, 25, 1641–1654. [Google Scholar] [CrossRef] [Green Version]
- Iravani, S.; Varma, R.S. Plant-Derived Edible Nanoparticles and miRNAs: Emerging Frontier for Therapeutics and Targeted Drug-Delivery. ACS Sustain. Chem. Eng. 2019, 7, 8055–8069. [Google Scholar] [CrossRef]
- Zhang, M.; Viennois, E.; Xu, C.; Merlin, D. Plant derived edible nanoparticles as a new therapeutic approach against diseases. Tissue Barriers 2016, 4, e1134415. [Google Scholar] [CrossRef]
- Berger, E.; Colosetti, P.; Jalabert, A.; Meugnier, E.; Wiklander, O.P.B.; Jouhet, J.; Errazurig-Cerda, E.; Chanon, S.; Gupta, D.; Rautureau, G.J.P.; et al. Use of Nanovesicles from Orange Juice to Reverse Diet-Induced Gut Modifications in Diet-Induced Obese Mice. Mol. Ther.-Methods Clin. Dev. 2020, 18, 880–892. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, B.; Li, X.; An, T.T.; Zhou, Y.; Li, G.; Wu-Smart, J.; Alvarez, S.; Naldrett, M.J.; Eudy, J.; et al. Identification of anti-inflammatory vesicle-like nanoparticles in honey. J. Extracell. Vesicles 2021, 10, e12069. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Lin, E.-Y.; Chiou, T.-W.; Harn, H.-J. Exosomes in clinical trial and their production in compliance with good manufacturing practice. Tzu-Chi Med. J. 2020, 32, 113–120. [Google Scholar] [CrossRef]
- Sarwa, K.K.; Das, P.J.; Mazumder, B. A nanovesicle topical formulation of Bhut Jolokia (hottest capsicum): A potential anti-arthritic medicine. Expert Opin. Drug Deliv. 2014, 11, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Sriwastva, M.K.; Deng, Z.-B.; Wang, B.; Teng, Y.; Kumar, A.; Sundaram, K.; Mu, J.; Lei, C.; Dryden, G.W.; Xu, F.; et al. Exosome-like nanoparticles from Mulberry bark prevent DSS-induced colitis via the AhR/COPS8 pathway. EMBO Rep. 2022, 23, e53365. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S.; et al. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, S.; Mu, J.; Dokland, T.; Zhuang, X.; Wang, Q.; Jiang, H.; Xiang, X.; Deng, Z.-B.; Wang, B.; Zhang, L.; et al. Grape exosome-like nanoparticles induce intestinal stem cells and protect mice from DSS-induced colitis. Mol. Ther. 2013, 21, 1345–1357. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.K.; Rhee, W.J. Antioxidative Effects of Carrot-Derived Nanovesicles in Cardiomyoblast and Neuroblastoma Cells. Pharmaceutics 2021, 13, 1203. [Google Scholar] [CrossRef]
- Kim, K.; Jung, J.-H.; Yoo, H.J.; Hyun, J.-K.; Park, J.-H.; Na, D.; Yeon, J.H. Anti-Metastatic Effects of Plant Sap-Derived Extracellular Vesicles in a 3D Microfluidic Cancer Metastasis Model. JFB 2020, 11, 49. [Google Scholar] [CrossRef]
- Raimondo, S.; Naselli, F.; Fontana, S.; Monteleone, F.; Lo Dico, A.; Saieva, L.; Zito, G.; Flugy, A.; Manno, M.; Di Bella, M.A.; et al. Citrus limon-derived nanovesicles inhibit cancer cell proliferation and suppress CML xenograft growth by inducing TRAIL-mediated cell death. Oncotarget 2015, 6, 19514–19527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Liu, X.; Luo, Q.; Xu, L.; Chen, F. An efficient method to isolate lemon derived extracellular vesicles for gastric cancer therapy. J. Nanobiotechnol. 2020, 18, 100. [Google Scholar] [CrossRef] [PubMed]
- Stanly, C.; Alfieri, M.; Ambrosone, A.; Leone, A.; Fiume, I.; Pocsfalvi, G. Grapefruit-Derived Micro and Nanovesicles Show Distinct Metabolome Profiles and Anticancer Activities in the A375 Human Melanoma Cell Line. Cells 2020, 9, 2722. [Google Scholar] [CrossRef] [PubMed]
- Pinna, R.; Filigheddu, E.; Juliano, C.; Palmieri, A.; Manconi, M.; D’hallewin, G.; Petretto, G.; Maioli, M.; Caddeo, C.; Manca, M.L.; et al. Antimicrobial Effect of Thymus capitatus and Citrus limon var. pompia as Raw Extracts and Nanovesicles. Pharmaceutics 2019, 11, 234. [Google Scholar] [CrossRef] [Green Version]
- Regente, M.; Pinedo, M.; San Clemente, H.; Balliau, T.; Jamet, E.; de la Canal, L. Plant extracellular vesicles are incorporated by a fungal pathogen and inhibit its growth. J. Exp. Bot. 2017, 68, 5485–5495. [Google Scholar] [CrossRef]
- Schuh, C.M.A.P.; Aguayo, S.; Zavala, G.; Khoury, M. Exosome-like vesicles in Apis mellifera bee pollen, honey and royal jelly contribute to their antibacterial and pro-regenerative activity. J. Exp. Biol. 2019, 222, jeb208702. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, K.; Miller, D.P.; Kumar, A.; Teng, Y.; Sayed, M.; Mu, J.; Lei, C.; Sriwastva, M.K.; Zhang, L.; Yan, J.; et al. Plant-Derived Exosomal Nanoparticles Inhibit Pathogenicity of Porphyromonas gingivalis. iScience 2019, 21, 308–327. [Google Scholar] [CrossRef] [Green Version]
- Benavente-García, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef]
- Baldini, N.; Torreggiani, E.; Roncuzzi, L.; Perut, F.; Zini, N.; Avnet, S. Exosome-like Nanovesicles Isolated from Citrus limon L. Exert Antioxidative Effect. Curr. Pharm. Biotechnol. 2018, 19, 877–885. [Google Scholar] [CrossRef]
- Logozzi, M.; Di Raimo, R.; Mizzoni, D.; Fais, S. Nanovesicles from Organic Agriculture-Derived Fruits and Vegetables: Characterization and Functional Antioxidant Content. IJMS 2021, 22, 8170. [Google Scholar] [CrossRef]
- Garaeva, L.; Kamyshinsky, R.; Kil, Y.; Varfolomeeva, E.; Verlov, N.; Komarova, E.; Garmay, Y.; Landa, S.; Burdakov, V.; Myasnikov, A.; et al. Delivery of functional exogenous proteins by plant-derived vesicles to human cells in vitro. Sci. Rep. 2021, 11, 6489. [Google Scholar] [CrossRef]
- Rutter, B.D.; Innes, R.W. Extracellular Vesicles Isolated from the Leaf Apoplast Carry Stress-Response Proteins. Plant. Physiol. 2017, 173, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Sagini, K.; Urbanelli, L.; Buratta, S.; Leonardi, L.; Emiliani, C. Nanovesicles from plants as edible carriers of bioactive compounds. AgroLife Sci. J. 2017, 6, 167–171. [Google Scholar]
- Sarvarian, P.; Samadi, P.; Gholipour, E.; Shams Asenjan, K.; Hojjat-Farsangi, M.; Motavalli, R.; Motavalli Khiavi, F.; Yousefi, M. Application of Emerging Plant-Derived Nanoparticles as a Novel Approach for Nano-Drug Delivery Systems. Immunol. Investig. 2021, 1–21. [Google Scholar] [CrossRef]
- Stanly, C.; Kim, H.; Antonucci, G.; Fiume, I.; Guescini, M.; Kim, K.P.; Ciardiello, M.A.; Giangrieco, I.; Mari, A.; Pocsfalvi, G. Crosstalk Between the Immune System and Plant-Derived Nanovesicles: A Study of Allergen Transporting. Front. Bioeng. Biotechnol. 2021, 9, 760730. [Google Scholar] [CrossRef]
- Stanly, C.; Moubarak, M.; Fiume, I.; Turiák, L.; Pocsfalvi, G. Membrane Transporters in Citrus clementina Fruit Juice-Derived Nanovesicles. Int. J. Mol. Sci. 2019, 20, 6205. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Mu, J.; Hu, X.; Samykutty, A.; Zhuang, X.; Deng, Z.; Zhang, L.; Cao, P.; Yan, J.; Miller, D.; et al. Grapefruit-derived nanovectors deliver miR-18a for treatment of liver metastasis of colon cancer by induction of M1 macrophages. Oncotarget 2016, 7, 25683–25697. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Zhang, H.-X.; He, C.-P.; Fan, S.; Zhu, Y.-L.; Qi, C.; Huang, N.-P.; Xiao, Z.-D.; Lu, Z.-H.; Tannous, B.A.; et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials 2018, 150, 137–149. [Google Scholar] [CrossRef]
- Umezu, T.; Takanashi, M.; Murakami, Y.; Ohno, S.-I.; Kanekura, K.; Sudo, K.; Nagamine, K.; Takeuchi, S.; Ochiya, T.; Kuroda, M. Acerola exosome-like nanovesicles to systemically deliver nucleic acid medicine via oral administration. Mol. Ther.—Methods Clin. Dev. 2021, 21, 199–208. [Google Scholar] [CrossRef]
- Wang, B.; Zhuang, X.; Deng, Z.-B.; Jiang, H.; Mu, J.; Wang, Q.; Xiang, X.; Guo, H.; Zhang, L.; Dryden, G.; et al. Targeted Drug Delivery to Intestinal Macrophages by Bioactive Nanovesicles Released from Grapefruit. Mol. Ther. 2014, 22, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ren, Y.; Mu, J.; Egilmez, N.K.; Zhuang, X.; Deng, Z.; Zhang, L.; Yan, J.; Miller, D.; Zhang, H.-G. Grapefruit-Derived Nanovectors Use an Activated Leukocyte Trafficking Pathway to Deliver Therapeutic Agents to Inflammatory Tumor Sites. Cancer Res. 2015, 75, 2520–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zhuang, X.; Mu, J.; Deng, Z.-B.; Jiang, H.; Zhang, L.; Xiang, X.; Wang, B.; Yan, J.; Miller, D.; et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat. Commun. 2013, 4, 1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Zhang, M.; Merlin, D. Advances in Plant-derived Edible Nanoparticle-based lipid Nano-drug Delivery Systems as Therapeutic Nanomedicines. J. Mater. Chem. B 2018, 6, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Deng, Z.; Liu, L.; Zhang, W.; Wang, C. Plant-Derived Nanovesicles: A Novel Form of Nanomedicine. Front. Bioeng. Biotechnol. 2020, 8, 584391. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wang, H.; Shi, W.; Chen, L.; Chen, T.; Chen, G.; Wang, W.; Lan, J.; Huang, Z.; Zhang, J.; et al. Aloe derived nanovesicle as a functional carrier for indocyanine green encapsulation and phototherapy. J. Nanobiotechnol. 2021, 19, 439. [Google Scholar] [CrossRef]
- Zhang, L.; He, F.; Gao, L.; Cong, M.; Sun, J.; Xu, J.; Wang, Y.; Hu, Y.; Asghar, S.; Hu, L.; et al. Engineering Exosome-Like Nanovesicles Derived from Asparagus cochinchinensis Can Inhibit the Proliferation of Hepatocellular Carcinoma Cells with Better Safety Profile. Int. J. Nanomed. 2021, 16, 1575–1586. [Google Scholar] [CrossRef]
- Zhang, M.; Collins, J.F.; Merlin, D. Do ginger-derived nanoparticles represent an attractive treatment strategy for inflammatory bowel diseases? Nanomedicine 2016, 11, 3035–3037. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Teng, Y.; Samykutty, A.; Mu, J.; Deng, Z.; Zhang, L.; Cao, P.; Rong, Y.; Yan, J.; Miller, D.; et al. Grapefruit-derived Nanovectors Delivering Therapeutic miR17 Through an Intranasal Route Inhibit Brain Tumor Progression. Mol. Ther. 2016, 24, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Federici, C.; Petrucci, F.; Caimi, S.; Cesolini, A.; Logozzi, M.; Borghi, M.; D’Ilio, S.; Lugini, L.; Violante, N.; Azzarito, T.; et al. Exosome Release and Low pH Belong to a Framework of Resistance of Human Melanoma Cells to Cisplatin. PLoS ONE 2014, 9, e88193. [Google Scholar] [CrossRef] [Green Version]
- Iessi, E.; Logozzi, M.; Lugini, L.; Azzarito, T.; Federici, C.; Spugnini, E.P.; Mizzoni, D.; Di Raimo, R.; Angelini, D.F.; Battistini, L.; et al. Acridine Orange/exosomes increase the delivery and the effectiveness of Acridine Orange in human melanoma cells: A new prototype for theranostics of tumors. J. Enzym. Inhib. Med. Chem. 2017, 32, 648–657. [Google Scholar] [CrossRef]
- Logozzi, M.; Mizzoni, D.; Bocca, B.; Di Raimo, R.; Petrucci, F.; Caimi, S.; Alimonti, A.; Falchi, M.; Cappello, F.; Campanella, C.; et al. Human primary macrophages scavenge AuNPs and eliminate it through exosomes. A natural shuttling for nanomaterials. Eur. J. Pharm. Biopharm. 2019, 137, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Logozzi, M.; De Milito, A.; Lugini, L.; Borghi, M.; Calabrò, L.; Spada, M.; Perdicchio, M.; Marino, M.L.; Federici, C.; Iessi, E.; et al. High levels of exosomes expressing CD63 and caveolin-1 in plasma of melanoma patients. PLoS ONE 2009, 4, e5219. [Google Scholar] [CrossRef] [Green Version]
- Logozzi, M.; Spugnini, E.; Mizzoni, D.; Di Raimo, R.; Fais, S. Extracellular acidity and increased exosome release as key phenotypes of malignant tumors. Cancer Metastasis Rev. 2019, 38, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Zorrilla, S.; Pérez-Sayans, M.; Fais, S.; Logozzi, M.; Gallas Torreira, M.; García García, A. A Pilot Clinical Study on the Prognostic Relevance of Plasmatic Exosomes Levels in Oral Squamous Cell Carcinoma Patients. Cancers 2019, 11, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osti, D.; Del Bene, M.; Rappa, G.; Santos, M.; Matafora, V.; Richichi, C.; Faletti, S.; Beznoussenko, G.V.; Mironov, A.; Bachi, A.; et al. Clinical Significance of Extracellular Vesicles in Plasma from Glioblastoma Patients. Clin. Cancer Res. 2019, 25, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Logozzi, M.; Mizzoni, D.; Di Raimo, R.; Giuliani, A.; Maggi, M.; Sciarra, A.; Fais, S. Plasmatic Exosome Number and Size Distinguish Prostate Cancer Patients From Healthy Individuals: A Prospective Clinical Study. Front. Oncol. 2021, 11, 727317. [Google Scholar] [CrossRef]
- Logozzi, M.; Di Raimo, R.; Mizzoni, D.; Fais, S. Immunocapture-based ELISA to characterize and quantify exosomes in both cell culture supernatants and body fluids. Methods Enzym. 2020, 645, 155–180. [Google Scholar] [CrossRef]
- Foglio, E.; Puddighinu, G.; Fasanaro, P.; D’Arcangelo, D.; Perrone, G.A.; Mocini, D.; Campanella, C.; Coppola, L.; Logozzi, M.; Azzarito, T.; et al. Exosomal clusterin, identified in the pericardial fluid, improves myocardial performance following MI through epicardial activation, enhanced arteriogenesis and reduced apoptosis. Int. J. Cardiol. 2015, 197, 333–347. [Google Scholar] [CrossRef]
- Properzi, F.; Logozzi, M.; Abdel-Haq, H.; Federici, C.; Lugini, L.; Azzarito, T.; Cristofaro, I.; di Sevo, D.; Ferroni, E.; Cardone, F.; et al. Detection of exosomal prions in blood by immunochemistry techniques. J. Gen. Virol. 2015, 96, 1969–1974. [Google Scholar] [CrossRef]
- Canitano, A.; Venturi, G.; Borghi, M.; Ammendolia, M.G.; Fais, S. Exosomes released in vitro from Epstein–Barr virus (EBV)-infected cells contain EBV-encoded latent phase mRNAs. Cancer Lett. 2013, 337, 193–199. [Google Scholar] [CrossRef]
- Lugini, L.; Valtieri, M.; Federici, C.; Cecchetti, S.; Meschini, S.; Condello, M.; Signore, M.; Fais, S. Exosomes from human colorectal cancer induce a tumor-like behavior in colonic mesenchymal stromal cells. Oncotarget 2016, 7, 50086–50098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossetti, C.; Lugini, L.; Astrologo, L.; Saggio, I.; Fais, S.; Spadafora, C. Soma-to-germline transmission of RNA in mice xenografted with human tumour cells: Possible transport by exosomes. PLoS ONE 2014, 9, e101629. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Yin, H.; Bennett, C.; Zhang, H.; Guo, P. Arrowtail RNA for Ligand Display on Ginger Exosome-like Nanovesicles to Systemic Deliver siRNA for Cancer Suppression. Sci. Rep. 2018, 8, 14644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trentini, M.; Zanotti, F.; Tiengo, E.; Camponogara, F.; Degasperi, M.; Licastro, D.; Lovatti, L.; Zavan, B. An Apple a Day Keeps the Doctor Away: Potential Role of miRNA 146 on Macrophages Treated with Exosomes Derived from Apples. Biomedicines 2022, 10, 415. [Google Scholar] [CrossRef]
- Şahin, F.; Koçak, P.; Güneş, M.Y.; Özkan, İ.; Yıldırım, E.; Kala, E.Y. In Vitro Wound Healing Activity of Wheat-Derived Nanovesicles. Appl. Biochem. Biotechnol. 2019, 188, 381–394. [Google Scholar] [CrossRef]
- Quesenberry, P.J.; Aliotta, J.; Camussi, G.; Abdel-Mageed, A.B.; Wen, S.; Goldberg, L.; Zhang, H.-G.; Tetta, C.; Franklin, J.; Coffey, R.J.; et al. Potential functional applications of extracellular vesicles: A report by the NIH Common Fund Extracellular RNA Communication Consortium. J. Extracell. Vesicles 2015, 4, 27575. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, X.; Han, M.K.; Collins, J.F.; Merlin, D. Oral administration of ginger-derived nanolipids loaded with siRNA as a novel approach for efficient siRNA drug delivery to treat ulcerative colitis. Nanomedicine 2017, 12, 1927–1943. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, Q. Development of liposomal formulations: From concept to clinical investigations. Asian J. Pharm. Sci. 2013, 8, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Ishida, O.; Maruyama, K.; Sasaki, K.; Iwatsuru, M. Size-dependent extravasation and interstitial localization of polyethyleneglycol liposomes in solid tumor-bearing mice. Int. J. Pharm. 1999, 190, 49–56. [Google Scholar] [CrossRef]
- Theek, B.; Baues, M.; Ojha, T.; Möckel, D.; Veettil, S.K.; Steitz, J.; van Bloois, L.; Storm, G.; Kiessling, F.; Lammers, T. Sonoporation enhances liposome accumulation and penetration in tumors with low EPR. J. Control. Release 2016, 231, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Logozzi, M.; Mizzoni, D.; Capasso, C.; Del Prete, S.; Di Raimo, R.; Falchi, M.; Angelini, D.F.; Sciarra, A.; Maggi, M.; Supuran, C.T.; et al. Plasmatic exosomes from prostate cancer patients show increased carbonic anhydrase IX expression and activity and low pH. J. Enzym. Inhib. Med. Chem. 2020, 35, 280–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logozzi, M.; Di Raimo, R.; Properzi, F.; Barca, S.; Angelini, D.F.; Mizzoni, D.; Falchi, M.; Battistini, L.; Fais, S. Nanovesicles released by OKT3 hybridoma express fully active antibodies. J. Enzym. Inhib. Med. Chem. 2021, 36, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, S.; Hossain, M.N.; Conese, M. Biological properties and therapeutic effects of plant-derived nanovesicles. Open Med. 2020, 15, 1096–1122. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, Y.; Yu, J. Exosome-like Nanoparticles from Ginger Rhizomes Inhibited NLRP3 Inflammasome Activation. Mol. Pharm. 2019, 16, 2690–2699. [Google Scholar] [CrossRef]
- Xiao, J.; Feng, S.; Wang, X.; Long, K.; Luo, Y.; Wang, Y.; Ma, J.; Tang, Q.; Jin, L.; Li, X.; et al. Identification of exosome-like nanoparticle-derived microRNAs from 11 edible fruits and vegetables. PeerJ 2018, 6, e5186. [Google Scholar] [CrossRef]
- Mu, J.; Zhuang, X.; Wang, Q.; Jiang, H.; Deng, Z.-B.; Wang, B.; Zhang, L.; Kakar, S.; Jun, Y.; Miller, D.; et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. Mol. Nutr. Food Res. 2014, 58, 1561–1573. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Yu, S.; Li, M.; Gui, X.; Li, P. Isolation of Exosome-Like Nanoparticles and Analysis of MicroRNAs Derived from Coconut Water Based on Small RNA High-Throughput Sequencing. J. Agric. Food Chem. 2018, 66, 2749–2757. [Google Scholar] [CrossRef]
- Abraham, A.M.; Wiemann, S.; Ambreen, G.; Zhou, J.; Engelhardt, K.; Brüßler, J.; Bakowsky, U.; Li, S.-M.; Mandic, R.; Pocsfalvi, G.; et al. Cucumber-Derived Exosome-like Vesicles and PlantCrystals for Improved Dermal Drug Delivery. Pharmaceutics 2022, 14, 476. [Google Scholar] [CrossRef]
- Zhuang, X.; Deng, Z.-B.; Mu, J.; Zhang, L.; Yan, J.; Miller, D.; Feng, W.; McClain, C.J.; Zhang, H.-G. Ginger-derived nanoparticles protect against alcohol-induced liver damage. J. Extracell. Vesicles 2015, 4, 28713. [Google Scholar] [CrossRef]
- Cho, E.-G.; Choi, S.-Y.; Kim, H.; Choi, E.-J.; Lee, E.-J.; Park, P.-J.; Ko, J.; Kim, K.P.; Baek, H.S. Panax ginseng-Derived Extracellular Vesicles Facilitate Anti-Senescence Effects in Human Skin Cells: An Eco-Friendly and Sustainable Way to Use Ginseng Substances. Cells 2021, 10, 486. [Google Scholar] [CrossRef]
- Pérez-Bermúdez, P.; Blesa, J.; Soriano, J.M.; Marcilla, A. Extracellular vesicles in food: Experimental evidence of their secretion in grape fruits. Eur. J. Pharm. Sci. 2017, 98, 40–50. [Google Scholar] [CrossRef]
- Woith, E.; Melzig, M.F. Extracellular Vesicles from Fresh and Dried Plants-Simultaneous Purification and Visualization Using Gel Electrophoresis. Int. J. Mol. Sci. 2019, 20, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perut, F.; Roncuzzi, L.; Avnet, S.; Massa, A.; Zini, N.; Sabbadini, S.; Giampieri, F.; Mezzetti, B.; Baldini, N. Strawberry-Derived Exosome-Like Nanoparticles Prevent Oxidative Stress in Human Mesenchymal Stromal Cells. Biomolecules 2021, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Bokka, R.; Ramos, A.P.; Fiume, I.; Manno, M.; Raccosta, S.; Turiák, L.; Sugár, S.; Adamo, G.; Csizmadia, T.; Pocsfalvi, G. Biomanufacturing of Tomato-Derived Nanovesicles. Foods 2020, 9, 1852. [Google Scholar] [CrossRef] [PubMed]
- Timms, K.; Holder, B.; Day, A.; McLaughlin, J.; Westwood, M.; Forbes, K. Isolation and Characterisation of Watermelon (Citrullus lanatus) Extracellular Vesicles and their Cargo. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Logozzi, M.; Di Raimo, R.; Mizzoni, D.; Fais, S. The Potentiality of Plant-Derived Nanovesicles in Human Health—A Comparison with Human Exosomes and Artificial Nanoparticles. Int. J. Mol. Sci. 2022, 23, 4919. https://doi.org/10.3390/ijms23094919
Logozzi M, Di Raimo R, Mizzoni D, Fais S. The Potentiality of Plant-Derived Nanovesicles in Human Health—A Comparison with Human Exosomes and Artificial Nanoparticles. International Journal of Molecular Sciences. 2022; 23(9):4919. https://doi.org/10.3390/ijms23094919
Chicago/Turabian StyleLogozzi, Mariantonia, Rossella Di Raimo, Davide Mizzoni, and Stefano Fais. 2022. "The Potentiality of Plant-Derived Nanovesicles in Human Health—A Comparison with Human Exosomes and Artificial Nanoparticles" International Journal of Molecular Sciences 23, no. 9: 4919. https://doi.org/10.3390/ijms23094919
APA StyleLogozzi, M., Di Raimo, R., Mizzoni, D., & Fais, S. (2022). The Potentiality of Plant-Derived Nanovesicles in Human Health—A Comparison with Human Exosomes and Artificial Nanoparticles. International Journal of Molecular Sciences, 23(9), 4919. https://doi.org/10.3390/ijms23094919