
Next-Generation Probiotics for Inflammatory Bowel Disease

 ,
,  , , , and
, , , and
Abstract
1. Introduction
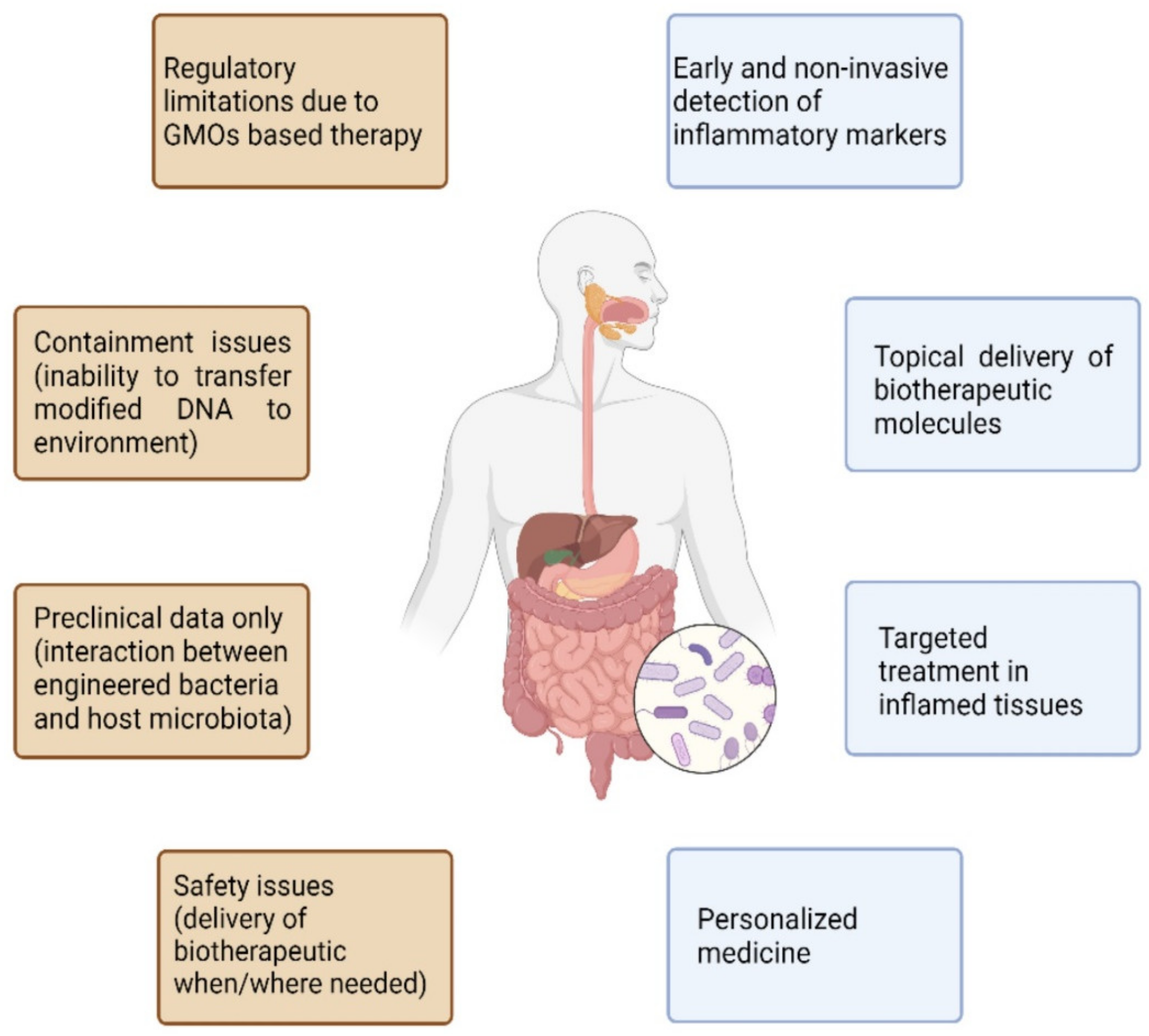
2. Microbiota Manipulation in IBD: Rationale and Current Limitations
3. Engineered Probiotics in IBD
3.1. Engineered Escherichia coli
3.2. Engineered Lactococcus lactis
3.3. Engineered Lactobacillus paracasei
3.4. Engineered Bifidobacterium longum
3.5. Engineered Bacteroides ovatus
4. Bacterial Outer Membrane Vesicles: Potential New Tools in Probiotic Engineering
5. Conclusions and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, K.; Kanai, T. The gut microbiota and inflammatory bowel disease. Semin. Immunopathol. 2015, 37, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- De Conno, B.; Pesce, M.; Chiurazzi, M.; Andreozzi, M.; Rurgo, S.; Corpetti, C.; Seguella, L.; Del Re, A.; Palenca, I.; Esposito, G.; et al. Nutraceuticals and Diet Supplements in Crohn’s Disease: A General Overview of the Most Promising Approaches in the Clinic. Foods 2022, 11, 1044. [Google Scholar] [CrossRef]
- Martini, E.; Krug, S.; Siegmund, B.; Neurath, M.F.; Becker, C. Mend Your Fences: The Epithelial Barrier and its Relationship with Mucosal Immunity in Inflammatory Bowel Disease. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 33–46. [Google Scholar] [CrossRef]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Frank, D.N.; Robertson, C.; Hamm, C.M.; Kpadeh, Z.; Zhang, T.; Chen, H.; Zhu, W.; Sartor, R.B.; Boedeker, E.C.; Harpaz, N.; et al. Disease phenotype and genotype are associated with shifts in intestinal-associated microbiota in inflammatory bowel diseases. Inflamm. Bowel Dis. 2011, 17, 179–184. [Google Scholar] [CrossRef]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humaran, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 558–572. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD—What role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Koren, O.; Goodrich, J.K.; Johansson, M.E.V.; Nalbantoglu, I.; Aitken, J.D.; Su, Y.; Chassaing, B.; Walters, W.A.; González, A.; et al. Transient Inability to Manage Proteobacteria Promotes Chronic Gut Inflammation in TLR5-Deficient Mice. Cell Host Microbe 2012, 12, 139–152. [Google Scholar] [CrossRef]
- Sartor, R. Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: Antibiotics, probiotics, and prebiotics. Gastroenterology 2004, 126, 1620–1633. [Google Scholar] [CrossRef]
- Aryayev, M.L.; Senkivska, L.I.; Bredeleva, N.K.; Talashova, I.V. Prophylaxis of acute respiratory infections via improving the immune system in late preterm newborns with E. coli strain Nissle 1917: A controlled pilot trial. Pilot Feasibility Stud. 2018, 4, 79. [Google Scholar] [CrossRef]
- Trebichavsky, I.; Splichal, I.; Rada, V.; Splichalova, A. Modulation of natural immunity in the gut by Escherichia coli strain Nissle 1917. Nutr. Rev. 2010, 68, 459–464. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Shao, L.; Kandasamy, S.; Fischer, D.D.; Rauf, A.; Langel, S.N.; Chattha, K.S.; Kumar, A.; Huang, H.-C.; Rajashekara, G.; et al. Escherichia coli Nissle 1917 protects gnotobiotic pigs against human rotavirus by modulating pDC and NK-cell responses. Eur. J. Immunol. 2016, 46, 2426–2437. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, J.; Wang, X.; Jin, Z.; Zhang, P.; Su, H.; Sun, X. Effect of Probiotics on Respiratory Tract Allergic Disease and Gut Microbiota. Front. Nutr. 2022, 9, 821900. [Google Scholar] [CrossRef]
- Dhanani, A.S.; Bagchi, T. Lactobacillus plantarum CS24.2 prevents Escherichia coli adhesion to HT-29 cells and also down-regulates enteropathogen-induced tumor necrosis factor-α and interleukin-8 expression. Microbiol. Immunol. 2013, 57, 309–315. [Google Scholar] [CrossRef]
- Kaur, L.; Gordon, M.; Baines, P.A.; Iheozor-Ejiofor, Z.; Sinopoulou, V.; Akobeng, A.K. Probiotics for induction of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2020, 3, CD005573. [Google Scholar] [CrossRef]
- Mimee, M.; Nadeau, P.; Hayward, A.; Carim, S.; Flanagan, S.; Jerger, L.; Collins, J.; McDonnell, S.; Swartwout, R.; Citorik, R.J.; et al. An ingestible bacterial-electronic system to monitor gastrointestinal health. Science 2018, 360, 915–918. [Google Scholar] [CrossRef]
- Cui, M.; Pang, G.; Zhang, T.; Sun, T.; Zhang, L.; Kang, R.; Xue, X.; Pan, H.; Yang, C.; Zhang, X.; et al. Optotheranostic Nanosystem with Phone Visual Diagnosis and Optogenetic Microbial Therapy for Ulcerative Colitis At-Home Care. ACS Nano 2021, 15, 7040–7052. [Google Scholar] [CrossRef]
- Riglar, D.; Giessen, T.; Baym, M.; Kerns, S.J.; Niederhuber, M.J.; Bronson, R.T.; Kotula, J.W.; Gerber, G.K.; Way, J.C.; Silver, P.A. Engineered bacteria can function in the mammalian gut long-term as live diagnostics of inflammation. Nat. Biotechnol. 2017, 35, 653–658. [Google Scholar] [CrossRef]
- Wang, L.; Liao, Y.; Yang, R.; Zhu, Z.; Zhang, L.; Wu, Z.; Sun, X. An engineered probiotic secreting Sj16 ameliorates colitis via Ruminococcaceae/butyrate/retinoic acid axis. Bioeng. Transl. Med. 2021, 6, e10219. [Google Scholar] [CrossRef]
- Esposito, G.; Pesce, M.; Seguella, L.; Lu, J.; Corpetti, C.; Del Re, A.; De Palma, F.; Esposito, G.; Sanseverino, W.; Sarnelli, G. Engineered Lactobacillus paracasei Producing Palmitoylethanolamide (PEA) Prevents Colitis in Mice. Int. J. Mol. Sci. 2021, 22, 2945. [Google Scholar] [CrossRef]
- Hamady, Z.Z.R.; Scott, N.; Farrar, M.D.; A Lodge, J.P.; Holland, K.T.; Whitehead, T.; Carding, S.R. Xylan-regulated delivery of human keratinocyte growth factor-2 to the inflamed colon by the human anaerobic commensal bacterium Bacteroides ovatus. Gut 2010, 59, 461–469. [Google Scholar] [CrossRef]
- Barra, M.; Danino, T.; Garrido, D. Engineered Probiotics for Detection and Treatment of Inflammatory Intestinal Diseases. Front. Bioeng. Biotechnol. 2020, 8, 265. [Google Scholar] [CrossRef]
- Naydich, A.D.; Nangle, S.N.; Bues, J.J.; Trivedi, D.; Nissar, N.; Inniss, M.C.; Niederhuber, M.J.; Way, J.C.; Silver, P.A.; Riglar, D.T. Synthetic Gene Circuits Enable Systems-Level Biosensor Trigger Discovery at the Host-Microbe Interface. mSystems 2019, 4, e00125-19. [Google Scholar] [CrossRef]
- Blum-Oehler, G.; Oswald, S.; Eiteljörge, K.; Sonnenborn, U.; Schulze, J.; Kruis, W.; Hacker, J. Development of strain-specific PCR reactions for the detection of the probiotic Escherichia coli strain Nissle 1917 in fecal samples. Res. Microbiol. 2003, 154, 59–66. [Google Scholar] [CrossRef]
- Sonnenborn, U.; Schulze, J. The non-pathogenic Escherichia coli strain Nissle 1917—Features of a versatile probiotic. Microb. Ecol. Health Dis. 2009, 21, 122–158. [Google Scholar] [CrossRef]
- Yan, X.; Liu, X.-Y.; Zhang, D.; Zhang, Y.-D.; Li, Z.-H.; Liu, X.; Wu, F.; Chen, G.-Q. Construction of a sustainable 3-hydroxybutyrate-producing probiotic Escherichia coli for treatment of colitis. Cell. Mol. Immunol. 2021, 18, 2344–2357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, Y.; Lan, X.; Xu, X.; Zhang, X.; Li, X.; Zhao, Y.; Li, G.; Du, C.; Lu, S.; et al. Oral Escherichia coli expressing IL-35 meliorates experimental colitis in mice. J. Transl. Med. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed]
- Spisni, E.; Valerii, M.C.; De Fazio, L.; Cavazza, E.; Borsetti, F.; Sgromo, A.; Candela, M.; Centanni, M.; Rizello, F.; Strillacci, A. Cyclooxygenase-2 Silencing for the Treatment of Colitis: A Combined In Vivo Strategy Based on RNA Interference and Engineered Escherichia Coli. Mol. Ther. 2015, 23, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, U.; O’Connell-Motherway, M.; Zomer, A.; Buist, G.; Shearman, C.; Canchaya, C.; Ventura, M.; Goesmann, A.; Gasson, M.J.; Kuipers, O.P.; et al. Complete Genome Sequence of the Prototype Lactic Acid Bacterium Lactococcus lactis subsp. cremoris MG1363. J. Bacteriol. 2007, 189, 3256–3270. [Google Scholar] [CrossRef] [PubMed]
- Morello, E.; Bermúdez-Humarán, L.; Llull, D.; Solé, V.; Miraglio, N.; Langella, P.; Poquet, I. Lactococcus lactis, an Efficient Cell Factory for Recombinant Protein Production and Secretion. J. Mol. Microbiol. Biotechnol. 2007, 14, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Berlec, A.; Škrlec, K.; Kocjan, J.; Olenic, M.; Štrukelj, B. Single plasmid systems for inducible dual protein expression and for CRISPR-Cas9/CRISPRi gene regulation in lactic acid bacterium Lactococcus lactis. Sci. Rep. 2018, 8, 1009. [Google Scholar] [CrossRef] [PubMed]
- Benbouziane, B.; Ribelles, P.; Aubry, C.; Martin, R.; Kharrat, P.; Riazi, A.; Langella, P.; Humaran, L.G.B. Development of a Stress-Inducible Controlled Expression (SICE) system in Lactococcus lactis for the production and delivery of therapeutic molecules at mucosal surfaces. J. Biotechnol. 2013, 168, 120–129. [Google Scholar] [CrossRef]
- Vandenbroucke, K.; Hans, W.; Van Huysse, J.; Neirynck, S.; Demetter, P.; Remaut, E.; Rottiers, P.; Steidler, L. Active delivery of trefoil factors by genetically modified Lactococcus lactis prevents and heals acute colitis in mice. Gastroenterology 2004, 127, 502–513. [Google Scholar] [CrossRef]
- Vandenbroucke, K.; De Haard, H.; Beirnaert, E.; Dreier, T.; Lauwereys, M.; Huyck, L.; Van Huysse, J.; Demetter, P.; Steidler, L.; Remaut, E.; et al. Orally administered L. lactis secreting an anti-TNF Nanobody demonstrate efficacy in chronic colitis. Mucosal Immunol. 2010, 3, 49–56. [Google Scholar] [CrossRef]
- Breyner, N.; Michon, C.; Sousa, C.; Boas, P.C.B.V.; Chain, F.; Azevedo, V.A.; Langella, P.; Chatel, J.-M. Microbial Anti-Inflammatory Molecule (MAM) from Faecalibacterium prausnitzii Shows a Protective Effect on DNBS and DSS-Induced Colitis Model in Mice through Inhibition of NF-κB Pathway. Front. Microbiol. 2017, 8, 114. [Google Scholar] [CrossRef]
- Hanson, M.L.; Hixon, J.A.; Li, W.; Felber, B.K.; Anver, M.R.; Stewart, C.A.; Janelsins, B.M.; Datta, S.; Shen, W.; McLean, M.; et al. Oral Delivery of IL-27 Recombinant Bacteria Attenuates Immune Colitis in Mice. Gastroenterology 2014, 146, 210–221.e13. [Google Scholar] [CrossRef]
- Actogenix, O. ActoGeniX Completes Phase 2A Clinical Trial of AG011. 2009. Available online: http://www.aescap.com/pdf/actogenix_press_release_october_9.pdf (accessed on 4 May 2022).
- Esposito, G.; Corpetti, C.; Pesce, M.; Seguella, L.; Annunziata, G.; Del Re, A.; Vincenzi, M.; Lattanzi, R.; Lu, J.; Sanseverino, W.; et al. A Palmitoylethanolamide Producing Lactobacillus paracasei Improves Clostridium difficile Toxin A-Induced Colitis. Front. Pharmacol. 2021, 12, 639728. [Google Scholar] [CrossRef]
- Pesce, M.; Esposito, G.; Sarnelli, G. Endocannabinoids in the treatment of gastrointestinal inflammation and symptoms. Curr. Opin. Pharmacol. 2018, 43, 81–86. [Google Scholar] [CrossRef]
- Pesce, M.; D’Alessandro, A.; Borrelli, O.; Gigli, S.; Seguella, L.; Cuomo, R.; Esposito, G.; Sarnelli, G. Endocannabinoid-related compounds in gastrointestinal diseases. J. Cell. Mol. Med. 2018, 22, 706–715. [Google Scholar] [CrossRef]
- Couch, D.G.; Cook, H.; Ortori, C.; Barrett, D.; Lund, J.N.; O’Sullivan, S.E. Palmitoylethanolamide and Cannabidiol Prevent Inflammation-induced Hyperpermeability of the Human Gut In Vitro and In Vivo—A Randomized, Placebo-controlled, Double-blind Controlled Trial. Inflamm. Bowel Dis. 2019, 25, 1006–1018. [Google Scholar] [CrossRef]
- Esposito, G.; Capoccia, E.; Turco, F.; Palumbo, I.; Lu, J.; Steardo, A.; Cuomo, R.; Sarnelli, G.; Steardo, L. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-α activation. Gut 2014, 63, 1300–1312. [Google Scholar] [CrossRef]
- Kriek, R. Palmitoylethanolamide: Problems regarding micronization, ultra-micronization and additives. Inflammopharmacology 2014, 22, 195–198. [Google Scholar] [CrossRef]
- Schell, M.A.; Karmirantzou, M.; Snel, B.; Vilanova, D.; Berger, B.; Pessi, G.; Zwahlen, M.-C.; Desiere, F.; Bork, P.; Delley, M.; et al. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2002, 99, 14422–14427. [Google Scholar] [CrossRef]
- Yazawa, K.; Fujimori, M.; Amano, J.; Kano, Y.; Taniguchi, S. Bifidobacterium longum as a delivery system for cancer gene therapy: Selective localization and growth in hypoxic tumors. Cancer Gene Ther. 2000, 7, 269–274. [Google Scholar] [CrossRef]
- Liu, M.; Li, S.; Zhang, Q.; Xu, Z.; Wang, J.; Sun, H. Oral engineered Bifidobacterium longum expressing rhMnSOD to suppress experimental colitis. Int. Immunopharmacol. 2018, 57, 25–32. [Google Scholar] [CrossRef]
- Wei, P.; Yang, Y.; Ding, Q.; Li, X.; Sun, H.; Liu, Z.; Huang, J.; Gong, Y. Oral delivery of Bifidobacterium longum expressing α-melanocyte-stimulating hormone to combat ulcerative colitis. J. Med. Microbiol. 2016, 65, 160–168. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brzoska, T.; Böhm, M.; Lügering, A.; Loser, K.; Luger, T.A. Terminal Signal: Anti-Inflammatory Effects of α-Melanocyte-Stimulating Hormone Related Peptides Beyond the Pharmacophore. Adv. Exp. Med. Biol. 2010, 681, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Hamady, Z.Z.; Scott, N.; Farrar, M.D.; Wadhwa, M.; Dilger, P.; Whitehead, T.R.; Thorpe, R.; Holland, K.T.; Lodge, P.J.; Carding, S.R. Treatment of colitis with a commensal gut bacterium engineered to secrete human tgf-β1 under the control of dietary xylan. Inflamm. Bowel Dis. 2011, 17, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- Barnard, J.A.; Beauchamp, R.D.; Coffey, R.J.; Moses, H.L. Regulation of intestinal epithelial cell growth by transforming growth factor type beta. Proc. Natl. Acad. Sci. USA 1989, 86, 1578–1582. [Google Scholar] [CrossRef]
- Hespell, R.; Whitehead, T. Physiology and Genetics of Xylan Degradation by Gastrointestinal Tract Bacteria. J. Dairy Sci. 1990, 73, 3013–3022. [Google Scholar] [CrossRef]
- Steidler, L.; Hans, W.; Schotte, L.; Neirynck, S.; Obermeier, F.; Falk, W.; Fiers, W.; Remaut, E. Treatment of Murine Colitis by Lactococcus lactis Secreting Interleukin-10. Science 2000, 289, 1352–1355. [Google Scholar] [CrossRef]
- Del Carmen, S.; LeBlanc, A.D.M.D.; Perdigon, G.; Pereira, V.B.; Miyoshi, A.; Azevedo, V.; LeBlanc, J.G. Evaluation of the Anti-Inflammatory Effect of Milk Fermented by a Strain of IL-10-Producing Lactococcus lactis Using a Murine Model of Crohn’s Disease. J. Mol. Microbiol. Biotechnol. 2011, 21, 138–146. [Google Scholar] [CrossRef]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; Araiza-Villanueva, M.G.; Cancino-Diaz, J.C.; López-Villegas, E.O.; Sriranganathan, N.; Boyle, S.M.; Contreras-Rodríguez, A. Roles of bacterial membrane vesicles. Arch. Microbiol. 2015, 197, 1–10. [Google Scholar] [CrossRef]
- Fazal, S.; Lee, R. Biomimetic Bacterial Membrane Vesicles for Drug Delivery Applications. Pharmaceutics 2021, 13, 1430. [Google Scholar] [CrossRef]
- Zhang, T.; Li, P.; Wu, X.; Lu, G.; Marcella, C.; Ji, X.; Ji, G.; Zhang, F. Alterations of Akkermansia muciniphila in the inflammatory bowel disease patients with washed microbiota transplantation. Appl. Microbiol. Biotechnol. 2020, 104, 10203–10215. [Google Scholar] [CrossRef]
- Kang, C.-S.; Ban, M.; Choi, E.-J.; Moon, H.-G.; Jeon, J.-S.; Kim, D.-K.; Park, S.-K.; Jeon, S.G.; Roh, T.-Y.; Myung, S.-J.; et al. Extracellular Vesicles Derived from Gut Microbiota, Especially Akkermansia muciniphila, Protect the Progression of Dextran Sulfate Sodium-Induced Colitis. PLoS ONE 2013, 8, e76520. [Google Scholar] [CrossRef]
- Ashrafian, F.; Behrouzi, A.; Shahriary, A.; Badi, S.A.; Davari, M.; Khatami, S.; Jamnani, F.R.; Fateh, A.; Vaziri, F.; Siadat, S.D. Comparative study of effect of Akkermansia muciniphila and its extracellular vesicles on toll-like receptors and tight junction. Gastroenterol. Hepatol. Bed Bench 2019, 12, 163–168. [Google Scholar]
- Yoon, H.S.; Cho, C.H.; Yun, M.S.; Jang, S.J.; You, H.J.; Kim, J.-H.; Han, D.; Cha, K.H.; Moon, S.H.; Lee, K.; et al. Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat. Microbiol. 2021, 6, 563–573. [Google Scholar] [CrossRef]
- Fabrega, M.J.; Rodríguez-Nogales, A.; Mesa, J.G.; Algieri, F.; Badía, J.; Giménez, R.; Galvez, J.; Baldomà, L. Intestinal Anti-inflammatory Effects of Outer Membrane Vesicles from Escherichia coli Nissle 1917 in DSS-Experimental Colitis in Mice. Front. Microbiol. 2017, 8, 1274. [Google Scholar] [CrossRef]
- Shen, Y.; Torchia, M.L.G.; Lawson, G.W.; Karp, C.; Ashwell, J.D.; Mazmanian, S.K. Outer Membrane Vesicles of a Human Commensal Mediate Immune Regulation and Disease Protection. Cell Host Microbe 2012, 12, 509–520. [Google Scholar] [CrossRef]
- Irene, C.; Fantappiè, L.; Caproni, E.; Zerbini, F.; Anesi, A.; Tomasi, M.; Zanella, I.; Stupia, S.; Prete, S.; Valensin, S.; et al. Bacterial outer membrane vesicles engineered with lipidated antigens as a platform for Staphylococcus aureus vaccine. Proc. Natl. Acad. Sci. USA 2019, 116, 21780–21788. [Google Scholar] [CrossRef]
- Carvalho, A.L.; Fonseca, S.; Miquel-Clopés, A.; Cross, K.; Kok, K.-S.; Wegmann, U.; Gil Cardoso, K.; Bentley, E.G.; Al Katy, S.H.; Coombes, J.L.; et al. Bioengineering commensal bacteria-derived outer membrane vesicles for delivery of biologics to the gastrointestinal and respiratory tract. J. Extracell. Vesicles 2019, 8, 1632100. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Models Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef]
- Corb Aron, R.A.; Abid, A.; Vesa, C.M.; Nechifor, A.C.; Behl, T.; Ghitea, T.C.; Munteanu, M.A.; Fratila, O.; Andronie-Cioara, F.L.; Toma, M.M.; et al. Recognizing the Benefits of Pre-/Probiotics in Metabolic Syndrome and Type 2 Diabetes Mellitus Considering the Influence of Akkermansia muciniphila as a Key Gut Bacterium. Microorganisms 2021, 9, 618. [Google Scholar] [CrossRef]
- Pavel, F.; Vesa, C.; Gheorghe, G.; Diaconu, C.; Stoicescu, M.; Munteanu, M.; Babes, E.; Tit, D.; Toma, M.; Bungau, S. Highlighting the Relevance of Gut Microbiota Manipulation in Inflammatory Bowel Disease. Diagnostics 2021, 11, 1090. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Therapeutic Factor | Effect | Target/Model | Reference |
|---|---|---|---|---|
| Escherichia coli Nissle | (R)-3-hydroxybutyrate | SCFA levels ↑ Akkermansia spp. ↑ Microscopical/macroscopical damage and DAI scores ↓ ILs levels ↓ | Colitis/DSS induced Colitis | [31] |
| Schistosome immunoregulatory protein | Ruminococcaceae↑ butyrate and retinoic acid production ↑ Treg cells ↑ Th17 cells ↓ Microscopical/macroscopical damage score ↓ | Colitis/DSS induced Colitis | [24] | |
| IL-35 | CD4+ IL-17A+ Th17 cells ↓ CD4 + CD25+Foxp3+ Tregs cells in spleen and mesenteric lymph nodes ↑ Colonic and serum IL-10 and IL-35 ↑ Colonic and serum IL-6 ↓ Microscopical/macroscopical damage score ↓ DAI score ↓ | Colitis/DSS induced Colitis | [32] | |
| IL-10 | Microscopical/macroscopical damage score ↓ IL-1, IL-12, MPO, and TNF-α levels ↓ | Colitis/DSS induced Colitis | [22] | |
| InvColi anti-COX-2 RNAi | DAI score ↓ Microscopical/macroscopical damage score ↓ ILs levels ↓ colitis-associated shift of gut microbiota ↓ COX-2 expression ↓ | Colitis/DSS induced Colitis | [33] | |
| Lactococcus lactis | IL-27 | DAI score ↓ Microscopical/macroscopical damage score ↓ ILs levels ↓ CD4(+) and IL-17(+) T cells in gut-associated lymphoid tissue ↓ | Colitis/transfer of CD4(+) CD45RB(hi) T cells into Rag(−/−) mice | [41] |
| TNFα-neutralizing Nanobodies | Microscopical/macroscopical damage score ↓ MPO levels ↓ Do not interfere with systemic Salmonella infection in colitic IL10−/− mice. | Colitis/DSS induced Colitis and IL10−/− mice | [39] | |
| Trefoil factors | Ptgs2 expression Microscopical/macroscopical damage score ↓ MPO levels ↓ | Colitis/DSS induced Colitis and IL10−/− mice | [38] | |
| IL-10 | Microscopical/macroscopical damage score ↓ | Colitis/DSS induced Colitis | [57] | |
| Microscopical/macroscopical damage score ↓ IFN-γ levels ↓ IFN-γ, IL-12, IL-17 positive cells ↓ | Crohn’s Disease/TNBS induced gut inflammation | [58] | ||
| pILMAM (cDNA coding for Faecalibacterium-prausnitzii-derived Microbial Anti-inflammatory Molecule) | IL-17, IL-6, and IL-5 levels ↓ IL10, TGFβ levels ↑ DAI score ↓ | Colitis/DSS and DNBS induced Colitis | [40] | |
| Bifidobacterium longum | MnSOD | Microscopical/macroscopical damage score ↓ MPO levels ↓ TNF-α, IL-1β, IL-6 and IL-8 levels ↓ | Colitis/DSS induced Colitis | [51] |
| α-MSH | Microscopical damage score ↓ NF-kB activation, TNFα, NO, and IL-6 levels ↓ MPO levels ↓ IL10 levels ↑ | Colitis/DSS induced Colitis | [52] | |
| Lactobacillus paracasei subsp. Paracasei F19 | NAPE-PLD/Palmitoylethanolamide | DAI score ↓ Microscopical/macroscopical damage score ↓ MPO levels ↓ iNOS and COX-2 expression, IL-1β, PGE-2, TNFα, and NO levels ↓ tight-junction protein expression in the gut epithelium ↑ | Colitis/DSS induced Colitis | [25] |
| Microscopical/macroscopical damage score and MAC387+ cells ↓ MPO levels ↓ TLR4, RhoA GTP, p-p38 MAPK, NF-kB, HIF1-α expression ↓ tight-junction proteins expression in the gut epithelium ↑ | Colitis/TCdA induced Colitis | [43] | ||
| Bacteroides ovatus | TGF-β | DAI score ↓ Microscopical/macroscopical damage score ↓ IL-1β and TNFα levels, and COX-2 expression ↓ | Colitis/DSS induced Colitis | [54] |
| Strain | Therapeutic Factor | Effect | Target/Model | Reference |
|---|---|---|---|---|
| Akkermansia muciniphila | OMVs | DAI score ↓ Microscopical/macroscopical damage score ↓ | Colitis/DSS induced Colitis | [63] |
| OMVs | Microscopical/macroscopical damage score ↓ epithelial permeability in vitro and in vivo ↓ tight-junction proteins expression in the epithelium in vitro and in vivo ↑ | low-grade inflammation and leaky gut/HFD-induced leaky gut in vitro leaky gut model | [65] | |
| OMVs | tight-junction proteins expression ↑ TLR4 and TLR2 expression ↓ | IBDs and metabolic syndrome/Caco-2 culture | [64] | |
| Bacteroides fragilis | OMVs | Microscopical/macroscopical damage score ↓ Foxp3 and IL-10 from CD4+ T cells ↑ | Colitis/DSS induced Colitis | [67] |
| Escherichia coli Nissle 1917 | OMVs | DAI score ↓ Microscopical/macroscopical damage score ↓ ILs, iNOS, and TNFα levels ↓ IL10 levels ↑ tight-junction proteins expression in the gut epithelium ↑ | Colitis/DSS induced Colitis | [66] |
| Bacteroides thetaiotaomicron | engineered OMVs containing KGF-2 | DAI score ↓ Microscopical/macroscopical damage score ↓ Protection and restoring of globet cells↑ | Colitis/DSS induced Colitis | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pesce, M.; Seguella, L.; Del Re, A.; Lu, J.; Palenca, I.; Corpetti, C.; Rurgo, S.; Sanseverino, W.; Sarnelli, G.; Esposito, G. Next-Generation Probiotics for Inflammatory Bowel Disease. Int. J. Mol. Sci. 2022, 23, 5466. https://doi.org/10.3390/ijms23105466
Pesce M, Seguella L, Del Re A, Lu J, Palenca I, Corpetti C, Rurgo S, Sanseverino W, Sarnelli G, Esposito G. Next-Generation Probiotics for Inflammatory Bowel Disease. International Journal of Molecular Sciences. 2022; 23(10):5466. https://doi.org/10.3390/ijms23105466
Chicago/Turabian StylePesce, Marcella, Luisa Seguella, Alessandro Del Re, Jie Lu, Irene Palenca, Chiara Corpetti, Sara Rurgo, Walter Sanseverino, Giovanni Sarnelli, and Giuseppe Esposito. 2022. "Next-Generation Probiotics for Inflammatory Bowel Disease" International Journal of Molecular Sciences 23, no. 10: 5466. https://doi.org/10.3390/ijms23105466
APA StylePesce, M., Seguella, L., Del Re, A., Lu, J., Palenca, I., Corpetti, C., Rurgo, S., Sanseverino, W., Sarnelli, G., & Esposito, G. (2022). Next-Generation Probiotics for Inflammatory Bowel Disease. International Journal of Molecular Sciences, 23(10), 5466. https://doi.org/10.3390/ijms23105466