
Proteomic Characterization of Virulence Factors and Related Proteins in Enterococcus Strains from Dairy and Fermented Food Products

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results
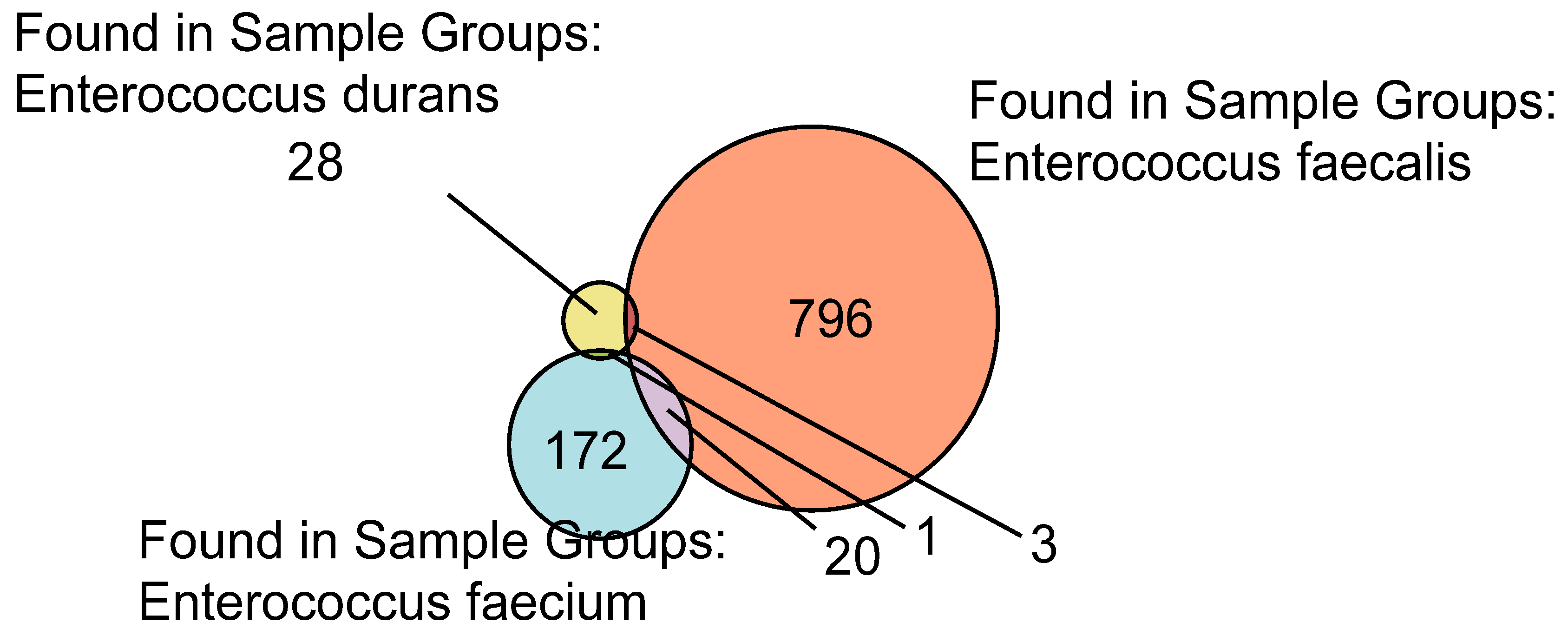
2.1. Enterococcus spp. Proteomics Data Repository
2.2. Label-Free Quantification (LFQ) of Enterococcus Species
2.3. Proteins Involved in Bacterial Resistance to Antibiotics and Other Toxic Substances
2.4. Antibacterial Compounds and Proteins Involved in Antibacterial Production
2.5. Proteins Involved in Bacterial Toxicity
2.6. Proteins Involved in Host Colonization and Immune Evasion
2.7. Transporters Associated with Virulence Factors
2.8. Other Bacterial Virulence Factors
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Protein Extraction
4.3. Peptide Sample Preparation
4.4. Shotgun LC–ESI–MS/MS Analysis
4.5. LC-ESI-MS/MS Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed]
- Stagliano, D.R.; Susi, A.; Adams, D.J.; Nylund, C.M. Epidemiology and Outcomes of Vancomycin-Resistant Enterococcus Infections in the U.S. Military Health System. Mil. Med. 2021, 186, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Devoe, C.; Segal, M.R.; Wang, L.; Stanley, K.; Madera, S.; Fan, J.; Schouest, J.; Graham-Ojo, R.; Nichols, A.; Prasad, P.A.; et al. Increased rates of secondary bacterial infections, including Enterococcus bacteremia, in patients hospitalized with coronavirus disease 2019 (COVID-19). Infect. Control Hosp. Epidemiol. 2021, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Goh, H.M.S.; Yong, M.H.A.; Chong, K.K.L.; Kline, K.A. Model systems for the study of Enterococcal colonization and infection. Virulence 2017, 8, 1525–1562. [Google Scholar] [CrossRef] [PubMed]
- Garsin, D.A.; Frank, K.L.; Silanpää, J.; Ausubel, F.M.; Hartke, A.; Shankar, N.; Murray, B.E. Pathogenesis and Models of Enterococcal Infection. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Banla, L.I.; Salzman, N.H.; Kristich, C.J. Colonization of the mammalian intestinal tract by enterococci. Curr. Opin. Microbiol. 2019, 47, 26–31. [Google Scholar] [CrossRef]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal gene transfer and the genomics of enterococcal antibiotic resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef]
- Hamzah, A.M.; Kadim, H.K. Isolation and identification of Enterococcus fecalis from cow milk samples and vaginal swab from human. J. Entomol. Zool. Stud. 2018, 6, 218–222. [Google Scholar]
- Sorescu, I.; Dumitru, M.; Ciurescu, G. Lactobacillus spp. and Enterococcus faecium strains isolation, identification, preservation and quantitative determinations from turkey gut content. Rom. Biotechnol. Lett. 2019, 24, 41–49. [Google Scholar] [CrossRef]
- Hayat, Z.; Shahzad, K.; Ali, H.; Casini, R.; Naveed, K.; Hafeez, A.; El-Ansary, D.O.; Elansary, H.O.; Fiaz, S.; Abaid-Ullah, M.; et al. 16S rRNA gene flow in Enterococcus spp. and SNP analysis: A reliable approach for specie level identification. Biochem. Syst. Ecol. 2022, 103, 104445. [Google Scholar] [CrossRef]
- Rogers, L.A.; Strong, K.; Cork, S.C.; McAllister, T.A.; Liljebjelke, K.; Zaheer, R.; Checkley, S.L. The Role of Whole Genome Sequencing in the Surveillance of Antimicrobial Resistant Enterococcus spp.: A Scoping Review. Front. Public Health 2021, 9, 656. [Google Scholar] [CrossRef]
- Waters, S.M.; Doyle, S.; Murphy, R.A.; Power, R.F.G. Development of solution phase hybridisation PCR-ELISA for the detection and quantification of Enterococcus faecalis and Pediococcus pentosaceus in Nurmi-type cultures. J. Microbiol. Methods 2005, 63, 264–275. [Google Scholar] [CrossRef] [PubMed][Green Version]
- La Rosa, S.L.; Solheim, M.; Diep, D.B.; Nes, I.F.; Brede, D.A. Bioluminescence based biosensors for quantitative detection of Enterococcal peptide–pheromone activity reveal inter-strain telesensing in vivo during polymicrobial systemic infection. Sci. Rep. 2015, 5, 8339. [Google Scholar] [CrossRef] [PubMed]
- Suriyanarayanan, T.; Qingsong, L.; Kwang, L.T.; Mun, L.Y.; Seneviratne, C.J. Quantitative proteomics of strong and weak biofilm formers of Enterococcus faecalis reveals novel regulators of biofilm formation. Mol. Cell. Proteom. 2018, 17, 643–654. [Google Scholar] [CrossRef]
- Giard, J.-C.; Auffray, Y.; Benachour, A.; Hartke, A.; Laplace, J.-M.; Rince, A.; Verneuil, N.; Pichereau, V. Proteomics Analysis: A Powerful Tool to Identify Proteome Phenotype and Proteome Signature in Enterococcus faecalis. Curr. Proteom. 2012, 1, 273–282. [Google Scholar] [CrossRef]
- Brackmann, M.; Leib, S.L.; Tonolla, M.; Schürch, N.; Wittwer, M. Antimicrobial resistance classification using MALDI-TOF-MS is not that easy: Lessons from vancomycin-resistant Enterococcus faecium. Clin. Microbiol. Infect. 2020, 26, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Holzknecht, B.J.; Dargis, R.; Pedersen, M.; Pinholt, M.; Christensen, J.J.; Hammerum, A.M.; Littauer, P.; Worning, P.; Westh, H.; Moser, C.; et al. Typing of vancomycin-resistant Enterococci with MALDI-TOF mass spectrometry in a nosocomial outbreak setting. Clin. Microbiol. Infect. 2018, 24, 1104.e1–1104.e4. [Google Scholar] [CrossRef]
- Cirrincione, S.; Neumann, B.; Zühlke, D.; Riedel, K.; Pessione, E. Detailed soluble proteome analyses of a dairy-isolated Enterococcus faecalis: A possible approach to assess food safety and potential probiotic value. Front. Nutr. 2019, 6, 71. [Google Scholar] [CrossRef]
- Carrera, M.; Böhme, K.; Gallardo, J.M.; Barros-Velázquez, J.; Cañas, B.; Calo-Mata, P. Characterization of foodborne strains of Staphylococcus aureus by shotgun proteomics: Functional networks, virulence factors and species-specific peptide biomarkers. Front. Microbiol. 2017, 8, 1–15. [Google Scholar]
- Pfrunder, S.; Grossmann, J.; Hunziker, P.; Brunisholz, R.; Gekenidis, M.T.; Drissner, D. Bacillus cereus Group-Type Strain-Specific Diagnostic Peptides. J. Proteome Res. 2016, 15, 3098–3107. [Google Scholar] [CrossRef]
- Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Calo-Mata, P.; Sánchez-Pérez, A.; Villa, T.G. Proteomic Characterization of Antibiotic Resistance in Listeria and Production of Antimicrobial and Virulence Factors. Int. J. Mol. Sci. 2021, 22, 8141. [Google Scholar] [CrossRef]
- Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Rama, J.L.R.; Calo-Mata, P.; Sánchez-Pérez, A.; Villa, T.G. Proteomic Characterization of Antibiotic Resistance, and Production of Antimicrobial and Virulence Factors in Streptococcus Species Associated with Bovine Mastitis. Could Enzybiotics Represent Novel Therapeutic Agents Against These Pathogens? Antibiotics 2020, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Brown, L.; Maryam, M.; Vij, R.; Smith, D.F.Q.; Burnet, M.C.; Kyle, J.E.; Heyman, H.M.; Ramirez, J.; Prados-Rosales, R.; et al. Listeria monocytogenes virulence factors, including listeriolysin O, are secreted in biologically active extracellular vesicles. J. Biol. Chem. 2019, 294, 1202–1217. [Google Scholar] [CrossRef] [PubMed]
- Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Cañas, B.; Rama, J.L.R.; Villa, T.G.; Calo-Mata, P. Proteomic Characterization of Bacteriophage Peptides from the Mastitis Producer Staphylococcus aureus by LC-ESI-MS/MS and the Bacteriophage Phylogenomic Analysis. Foods 2021, 10, 799. [Google Scholar] [CrossRef] [PubMed]
- Abril, A.G.; Villa, T.G.; Calo-Mata, P.; Barros-Velázquez, J.; Carrera, M. Application of proteomics to the identification of foodborne pathogens. In Food Proteom: Technological Advances, Current Applications and Future Perspectives; Academic Press: New York, NY, USA, 2022; pp. 337–362. [Google Scholar]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Dhasmana, N.; Dubey, N.; Kumar, N.; Gangwal, A.; Gupta, M.; Singh, Y. Bacterial Virulence Factors: Secreted for Survival. Indian J. Microbiol. 2017, 57, 1–10. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Łaniewska-Trokenheim, Ł. Virulence factors of Enterococcus spp. presented in food. LWT 2017, 75, 670–676. [Google Scholar] [CrossRef]
- Wagner, T.; Joshi, B.; Janice, J.; Askarian, F.; Škalko-Basnet, N.; Hagestad, O.C.; Mekhlif, A.; Wai, S.N.; Hegstad, K.; Johannessen, M. Enterococcus faecium produces membrane vesicles containing virulence factors and antimicrobial resistance related proteins. J. Proteom. 2018, 187, 28–38. [Google Scholar] [CrossRef]
- Ghaziasgar, F.S.; Poursina, F.; Hassanzadeh, A. Virulence factors, biofilm formation and antibiotic resistance pattern in Enterococcus faecalis and Enterococcus faecium isolated from clinical and commensal human samples in Isfahan, Iran. Ann. Ig. 2019, 31, 156–164. [Google Scholar]
- Najafi, K.; Ganbarov, K.; Gholizadeh, P.; Tanomand, A.; Rezaee, M.A.; Mahmood, S.S.; Asgharzadeh, M.; Kafil, H.S. Oral cavity infection by Enterococcus faecalis: Virulence factors and pathogenesis. Rev. Res. Med. Microbiol. 2020, 31, 51–60. [Google Scholar] [CrossRef]
- Dowson, C.G.; Hutchison, A.; Brannigan, J.A.; George, R.C.; Hansman, D.; Linares, J.; Tomasz, A.; Smith, J.M.; Spratt, B.G. Horizontal transfer of penicillin-binding protein genes in penicillin-resistant clinical isolates of Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 1989, 86, 8842–8846. [Google Scholar] [CrossRef]
- Grove, A. MarR family transcription factors. Curr. Biol. 2013, 23, R142–R143. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, L.; Nodwell, J.R. The TetR Family of Regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [PubMed]
- Yeats, C.; Finn, R.D.; Bateman, A. The PASTA domain: A β-lactam-binding domain. Trends Biochem. Sci. 2002, 27, 438–440. [Google Scholar] [CrossRef]
- Alabdali, Y.A.J.; Oatley, P.; Kirk, J.A.; Fagan, R.P. A cortex-specific penicillin-binding protein contributes to heat resistance in Clostridioides difficile spores. Anaerobe 2021, 70, 102379. [Google Scholar] [CrossRef] [PubMed]
- Kristich, C.J.; Wells, C.L.; Dunny, G.M. A eukaryotic-type Ser/Thr kinase in Enterococcus faecalis mediates antimicrobial resistance and intestinal persistence. Proc. Natl. Acad. Sci. USA 2007, 104, 3508–3513. [Google Scholar] [CrossRef] [PubMed]
- Vetting, M.W.; Luiz, L.P.; Yu, M.; Hegde, S.S.; Magnet, S.; Roderick, S.L.; Blanchard, J.S. Structure and functions of the GNAT superfamily of acetyltransferases. Arch. Biochem. Biophys. 2005, 433, 212–226. [Google Scholar] [CrossRef]
- Arthur, M.; Molinas, C.; Courvalin, P. Sequence of the vanY gene required for production of a vancomycin-inducible D,D-carboxypeptidase in Enterococcus faecium BM4147. Gene 1992, 120, 111–114. [Google Scholar] [CrossRef]
- Kim, H.S.; Hahn, H.; Kim, J.; Jang, D.M.; Lee, J.Y.; Back, J.M.; Im, H.N.; Kim, H.; Han, B.W.; Suh, S.W. Structural basis for the substrate recognition of peptidoglycan pentapeptides by Enterococcus faecalis VanYB. Int. J. Biol. Macromol. 2018, 119, 335–344. [Google Scholar] [CrossRef]
- Erlandson, A.; Gade, P.; Menikpurage, I.P.; Kim, C.Y.; Mera, P.E. The UvrA-like protein Ecm16 requires ATPase activity to render resistance against echinomycin. Mol. Microbiol. 2022, 117, 1434–1446. [Google Scholar] [CrossRef]
- Viswanath, L.S.; Sugumar, M.; Peela, S.C.M.; Walia, K.; Sistla, S. Detection of vancomycin variable enterococci (VVE) among clinical isolates of Enterococcus faecium collected across India-first report from the subcontinent. Indian J. Med. Microbiol. 2022, 40, 285–288. [Google Scholar] [CrossRef]
- Park, S.C.; Kwak, Y.M.; Song, W.S.; Hong, M.; Yoon, S. Il Structural basis of effector and operator recognition by the phenolic acid-responsive transcriptional regulator PadR. Nucleic Acids Res. 2017, 45, 13080–13093. [Google Scholar] [CrossRef] [PubMed]
- Luque-Sastre, L.; Arroyo, C.; Fox, E.M.; McMahon, B.J.; Bai, L.; Li, F.; Fanning, S. Antimicrobial Resistance in Listeria Species. In Antimicrobial Resistance in Bacteria from Livestock and Companion Animals; ASM Press: Washington, DC, USA, 2018; pp. 237–259. [Google Scholar]
- Suprenant, K.A.; Bloom, N.; Fang, J.; Lushington, G. The major vault protein is related to the toxic anion resistance protein (TelA) family. J. Exp. Biol. 2007, 210, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, C.N.; Harrop, S.J.; Boucher, Y.; Hassan, K.A.; Leo, R.D.; Xu, X.; Cui, H.; Savchenko, A.; Chang, C.; Labbate, M.; et al. Crystal Structure of an Integron Gene Cassette-Associated Protein from Vibrio cholerae Identifies a Cationic Drug-Binding Module. PLoS ONE 2011, 6, e16934. [Google Scholar] [CrossRef] [PubMed]
- Hobman, J.L.; Wilkie, J.; Brown, N.L. A design for life: Prokaryotic metal-binding MerR family regulators. BioMetals 2005, 18, 429–436. [Google Scholar] [CrossRef]
- Son, M.S.; Del Castilho, C.; Duncalf, K.A.; Carney, D.; Weiner, J.H.; Turner, R.J. Mutagenesis of SugE, a small multidrug resistance protein. Biochem. Biophys. Res. Commun. 2003, 312, 914–921. [Google Scholar] [CrossRef]
- Ermolenko, D.N.; Makhatadze, G.I. Bacterial cold-shock proteins. Cell. Mol. Life Sci. C 2002, 59, 1902–1913. [Google Scholar] [CrossRef]
- Barcarolo, M.V.; Gottig, N.; Ottado, J.; Garavaglia, B.S. Participation of two general stress response proteins from Xanthomonas citri subsp. citri in environmental stress adaptation and virulence. FEMS Microbiol. Ecol. 2020, 96, 138. [Google Scholar] [CrossRef]
- Reuß, D.R.; Altenbuchner, J.; Mäder, U.; Rath, H.; Ischebeck, T.; Sappa, P.K.; Thürmer, A.; Guérin, C.; Nicolas, P.; Steil, L.; et al. Large-scale reduction of the Bacillus subtilis genome: Consequences for the transcriptional network, resource allocation, and metabolism. Genome Res. 2017, 27, 289–299. [Google Scholar] [CrossRef]
- Begley, M.; Cotter, P.D.; Hill, C.; Ross, R.P. Identification of a novel two-peptide lantibiotic, lichenicidin, following rational genome mining for LanM proteins. Appl. Environ. Microbiol. 2009, 75, 5451–5460. [Google Scholar] [CrossRef]
- Montalbán-López, M.; Scott, T.A.; Ramesh, S.; Rahman, I.R.; Van Heel, A.J.; Viel, J.H.; Bandarian, V.; Dittmann, E.; Genilloud, O.; Goto, Y.; et al. New developments in RiPP discovery, enzymology and engineering. Nat. Prod. Rep. 2021, 38, 130–239. [Google Scholar] [CrossRef]
- Abril, A.G.; Villa, T.G.; Barros-Velázquez, J.; Cañas, B.; Sánchez-Pérez, A.; Calo-Mata, P.; Carrera, M. Staphylococcus aureus Exotoxins and Their Detection in the Dairy Industry and Mastitis. Toxins 2020, 12, 537. [Google Scholar] [CrossRef] [PubMed]
- Brielle, R.; Pinel-Marie, M.L.; Felden, B. Linking bacterial type I toxins with their actions. Curr. Opin. Microbiol. 2016, 30, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Lima-Mendez, G.; Alvarenga, D.O.; Ross, K.; Hallet, B.; Van Melderen, L.; Varani, A.M.; Chandler, M. Toxin-Antitoxin Gene Pairs Found in Tn3 Family Transposons Appear To Be an Integral Part of the Transposition Module. MBio 2020, 11, e00452-20. [Google Scholar] [CrossRef] [PubMed]
- Szczepanowski, R.; Braun, S.; Riedel, V.; Schneiker, S.; Krahn, I.; Pühler, A.; Schlüter, A. The 120 592 bp IncF plasmid pRSB107 isolated from a sewage-treatment plant encodes nine different antiobiotic-resistance determinants, two iron-acquisition systems and other putative virulence-associated functions. Microbiology 2005, 151, 1095–1111. [Google Scholar] [CrossRef]
- Arcus, V.L.; Mckenzie, J.L.; Robson, J.; Cook, G.M. The PIN-domain ribonucleases and the prokaryotic VapBC toxin–antitoxin array. Protein Eng. Des. Sel. 2011, 24, 33–40. [Google Scholar] [CrossRef]
- Nes, I.F.; Diep, D.B.; Holo, H. Bacteriocin diversity in Streptococcus and Enterococcus. J. Bacteriol. 2007, 189, 1189–1198. [Google Scholar] [CrossRef]
- Dramsi, S.; Bierne, H. Spatial organization of cell wall-anchored proteins at the surface of gram-positive bacteria. Curr. Top. Microbiol. Immunol. 2017, 404, 177–201. [Google Scholar]
- Hendrickx, A.P.A.; Van Luit-Asbroek, M.; Schapendonk, C.M.E.; Van Wamel, W.J.B.; Braat, J.C.; Wijnands, L.M.; Bonten, M.J.M.; Willems, R.J.L. SgrA, a nidogen-binding LPXTG surface adhesin implicated in biofilm formation, and EcbA, a collagen binding MSCRAMM, are two novel adhesins of hospital-acquired Enterococcus faecium. Infect. Immun. 2009, 77, 5097–5106. [Google Scholar] [CrossRef]
- Jacobitz, A.W.; Kattke, M.D.; Wereszczynski, J.; Clubb, R.T. Sortase Transpeptidases: Structural Biology and Catalytic Mechanism. Adv. Protein Chem. Struct. Biol. 2017, 109, 223–264. [Google Scholar]
- Guiton, P.S.; Hung, C.S.; Kline, K.A.; Roth, R.; Kau, A.L.; Hayes, E.; Heuser, J.; Dodson, K.W.; Caparon, M.G.; Hultgren, S.J. Contribution of autolysin and sortase A during Enterococcus faecalis DNA-dependent biofilm development. Infect. Immun. 2009, 77, 3626–3638. [Google Scholar] [CrossRef]
- Kristich, C.J.; Nguyen, V.T.; Le, T.; Barnes, A.M.T.; Grindle, S.; Dunny, G.M. Development and use of an efficient system for random mariner transposon mutagenesis to identify novel genetic determinants of biofilm formation in the core Enterococcus faecalis genome. Appl. Environ. Microbiol. 2008, 74, 3377–3386. [Google Scholar] [CrossRef] [PubMed]
- Rego, S.; Heal, T.J.; Pidwill, G.R.; Till, M.; Robson, A.; Lamont, R.J.; Sessions, R.B.; Jenkinson, H.F.; Race, P.R.; Nobbs, A.H. Structural and Functional Analysis of Cell Wall-anchored Polypeptide Adhesin BspA in Streptococcus agalactiae. J. Biol. Chem. 2016, 291, 15985–16000. [Google Scholar] [CrossRef]
- Epler Barbercheck, C.R.; Bullitt, E.; Andersson, M. Bacterial adhesion pili. In Subcellular Biochemistry; Springer: New York, NY, USA, 2018; Volume 87, pp. 1–18. [Google Scholar]
- Spraggon, G.; Koesema, E.; Scarselli, M.; Malito, E.; Biagini, M.; Norais, N.; Emolo, C.; Barocchi, M.A.; Giusti, F.; Hilleringmann, M.; et al. Supramolecular Organization of the Repetitive Backbone Unit of the Streptococcus pneumoniae Pilus. PLoS ONE 2010, 5, e10919. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. The Pseudomonas aeruginosa flagellar cap protein, FliD, is responsible for mucin adhesion. Infect. Immun. 1998, 66, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Pozzo, T.; Pasten, J.L.; Karlsson, E.N.; Logan, D.T. Structural and Functional Analyses of β-Glucosidase 3B from Thermotoga neapolitana: A Thermostable Three-Domain Representative of Glycoside Hydrolase 3. J. Mol. Biol. 2010, 397, 724–739. [Google Scholar] [CrossRef]
- Campellone, K.G. Cytoskeleton-modulating effectors of enteropathogenic and enterohaemorrhagic Escherichia coli: Tir, EspFU and actin pedestal assembly. FEBS J. 2010, 277, 2390–2402. [Google Scholar] [CrossRef]
- Ben Said, L.; Jouini, A.; Fliss, I.; Torres, C.; Klibi, N. Antimicrobial resistance genes and virulence gene encoding intimin in Escherichia coli and Enterococcus isolated from wild rabbits (Oryctolagus cuniculus) in Tunisia. Acta Vet. Hung. 2019, 67, 477–488. [Google Scholar] [CrossRef]
- Andreevskaya, M.; Jääskeläinen, E.; Johansson, P.; Ylinen, A.; Paulin, L.; Björkroth, J.; Auvinen, P. Food spoilage-associated Leuconostoc, Lactococcus, and Lactobacillus species display different survival strategies in response to competition. Appl. Environ. Microbiol. 2018, 84, 554–572. [Google Scholar] [CrossRef]
- Schubert, K.; Bichlmaier, A.M.; Mager, E.; Wolff, K.; Ruhland, G.; Fiedler, F. P45, an extracellular 45 kDa protein of Listeria monocytogenes with similarity to protein p60 and exhibiting peptidoglycan lyric activity. Arch. Microbiol. 2000, 173, 21–28. [Google Scholar] [CrossRef]
- Mori, S.; Mori, K.; Suzuki, I.; Kasumi, T. Phylogenetic Analysis of Lactococcus lactis subspecies Based on Decoding the Sequence of the pepT Tripeptidase Gene, the pepV Dipeptidase Gene and 16S rRNA. Syst. Appl. Microbiol. 2004, 27, 414–422. [Google Scholar] [CrossRef]
- Kanamaru, K.; Stephenson, S.; Perego, M. Overexpression of the PepF oligopeptidase inhibits sporulation initiation in Bacillus subtilis. J. Bacteriol. 2002, 184, 43–50. [Google Scholar] [CrossRef]
- Gómez, L.A.; Alvarez, F.I.; Molina, R.; Soto, R.; Daza-Castro, C.; Flores, M.; León, Y.; Oñate, A.A. A Zinc-dependent metalloproteinase in the intracellular adaptation of Brucella abortus in macrophages. bioRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Livingston, E.T.; Mursalin, M.H.; Coburn, P.S.; Astley, R.; Miller, F.C.; Amayem, O.; Lereclus, D.; Callegan, M.C. Immune inhibitor a metalloproteases contribute to virulence in Bacillus endophthalmitis. Infect. Immun. 2021, 89, IAI0020121. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.Y.; Ogunniyi, A.D.; Choi, M.H.; Pyo, S.N.; Rhee, D.K.; Paton, J.C. The ClpP protease of Streptococcus pneumoniae modulates virulence gene expression and protects against fatal pneumococcal challenge. Infect. Immun. 2004, 72, 5646–5653. [Google Scholar] [CrossRef]
- Kajfasz, J.K.; Martinez, A.R.; Rivera-ramos, I.; Abranches, J.; Koo, H.; Quivey, R.G. Role of Clp Proteins in Expression of Virulence Properties of Streptococcus mutans. J. Bacteriol. 2009, 191, 2060–2068. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Trappetti, C. Streptococcus pneumoniae Capsular Polysaccharide. In Gram-Positive Pathogens; ASM Press: Washington, DC, USA, 2019; pp. 304–315. [Google Scholar]
- Palmer, K.L.; Godfrey, P.; Griggs, A.; Kos, V.N.; Zucker, J.; Desjardins, C.; Cerqueira, G.; Gevers, D.; Walker, S.; Wortman, J.; et al. Comparative genomics of Enterococci: Variation in Enterococcus faecalis, clade structure in E. faecium, and defining characteristics of E. gallinarum and E. casseliflavus. MBio 2012, 3, e00318-11. [Google Scholar] [CrossRef]
- Domingos, R.M.; Teixeira, R.D.; Zeida, A.; Agudelo, W.A.; Alegria, T.G.P.; Da Silva Neto, J.F.; Vieira, P.S.; Murakami, M.T.; Farah, C.S.; Estrin, D.A.; et al. Substrate and product-assisted catalysis: Molecular aspects behind structural switches along organic hydroperoxide resistance protein catalytic cycle. ACS Catal. 2020, 10, 6587–6602. [Google Scholar] [CrossRef]
- Bateman, A.; Bycroft, M. The structure of a LysM domain from E. coli membrane-bound lytic murein transglycosylase D (MltD). J. Mol. Biol. 2000, 299, 1113–1119. [Google Scholar] [CrossRef]
- Rehman, M.A.; Yin, X.; Zaheer, R.; Goji, N.; Amoako, K.K.; McAllister, T.; Pritchard, J.; Topp, E.; Diarra, M.S. Genotypes and Phenotypes of Enterococci Isolated From Broiler Chickens. Front. Sustain. Food Syst. 2018, 2, 83. [Google Scholar] [CrossRef]
- Ruiz-Cruz, S.; Moreno-Blanco, A.; Espinosa, M.; Bravo, A. Transcriptional activation by MafR, a global regulator of Enterococcus faecalis. Sci. Rep. 2019, 9, 6146. [Google Scholar] [CrossRef]
- Ribardo, D.A.; Lambert, T.J.; McIver, K.S. Role of Streptococcus pyogenes two-component response regulators in the temporal control of Mga and the Mga-regulated virulence gene emm. Infect. Immun. 2004, 72, 3668–3673. [Google Scholar] [CrossRef] [PubMed]
- Warne, B.; Harkins, C.P.; Harris, S.R.; Vatsiou, A.; Stanley-Wall, N.; Parkhill, J.; Peacock, S.J.; Palmer, T.; Holden, M.T.G. The Ess/Type VII secretion system of Staphylococcus aureus shows unexpected genetic diversity. BMC Genom. 2016, 17, 222. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamed, H.; Ramachandran, R.; Narayanan, L.; Ozdemir, O.; Cooper, A.; Olivier, A.K.; Karsi, A.; Lawrence, M.L. Contributions of a LysR transcriptional regulator to listeria monocytogenes virulence and identification of its regulons. J. Bacteriol. 2020, 202, e00087-20. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. In Antimicrobial Resistance in Bacteria from Livestock and Companion Animals; ASM Press: Washington, DC, USA, 2018; pp. 185–227. [Google Scholar]
- Trost, M.; Wehmhöner, D.; Kärst, U.; Dieterich, G.; Wehland, J.; Jänsch, L. Comparative proteome analysis of secretory proteins from pathogenic and nonpathogenic Listeria species. Proteomics 2005, 5, 1544–1557. [Google Scholar] [CrossRef]
- McGowan, S.; Lucet, I.S.; Cheung, J.K.; Awad, M.M.; Whisstock, J.C.; Rood, J.I. The FxRxHrS Motif: A Conserved Region Essential for DNA Binding of the VirR Response Regulator from Clostridium perfringens. J. Mol. Biol. 2002, 322, 997–1011. [Google Scholar] [CrossRef]
- Del Papa, M.F.; Perego, M. Enterococcus faecalis virulence regulator FsrA binding to target promoters. J. Bacteriol. 2011, 193, 1527–1532. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Y.; Humphris, S.; Nie, W.; Zhang, P.; Wright, F.; Campbell, E.; Hu, B.; Fan, J.; Toth, I. Pectobacterium atrosepticum KDPG aldolase, Eda, participates in the Entner–Doudoroff pathway and independently inhibits expression of virulence determinants. Mol. Plant Pathol. 2021, 22, 271–283. [Google Scholar] [CrossRef]
- Del Mar Lleo, M.; Fontana, R.; Solioz, M. Identification of a gene (arpU) controlling muramidase-2 export in Enterococcus hirae. J. Bacteriol. 1995, 177, 5912–5917. [Google Scholar] [CrossRef][Green Version]
- Molloy, S. A competent escape for Listeria. Nat. Rev. Microbiol. 2012, 10, 670. [Google Scholar] [CrossRef]
- Ud-Din, A.I.M.S.; Roujeinikova, A. Methyl-accepting chemotaxis proteins: A core sensing element in Prokaryotes and Archaea. Cell. Mol. Life Sci. 2017, 74, 3293–3303. [Google Scholar] [CrossRef]
- Kuwana, R.; Yamamura, S.; Ikejiri, H.; Kobayashi, K.; Ogasawara, N.; Asai, K.; Sadaie, Y.; Takamatsu, H.; Watabe, K. Bacillus subtilis spoVIF (yjcC) gene, involved in coat assembly and spore resistance. Microbiology 2003, 149, 3011–3021. [Google Scholar] [CrossRef]
- Aertsen, A.; Michiels, C.W. Mrr instigates the SOS response after high pressure stress in Escherichia coli. Mol. Microbiol. 2005, 58, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- Guerardel, Y.; Sadovskaya, I.; Maes, E.; Furlan, S.; Chapot-Chartier, M.P.; Mesnage, S.; Rigottier-Gois, L.; Serror, P. Complete structure of the Enterococcal polysaccharide antigen (EPA) of vancomycin-resistant Enterococcus faecalis v583 reveals that EPA decorations are teichoic acids covalently linked to a rhamnopolysaccharide backbone. MBio 2020, 11, e00277-20. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Sim, J.R.; Yun, C.H.; Han, S.H. Lipoteichoic acids as a major virulence factor causing inflammatory responses via Toll-like receptor 2. Arch. Pharm. Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Benachour, A.; Ladjouzi, R.; Le Jeune, A.; Hébert, L.; Thorpe, S.; Courtin, P.; Chapot-Chartier, M.P.; Prajsnar, T.K.; Foster, S.J.; Mesnage, S. The Lysozyme-Induced Peptidoglycan N-Acetylglucosamine Deacetylase pgda (EF1843) Is Required for Enterococcus faecalis Virulence. J. Bacteriol. 2012, 194, 6066–6073. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, W.; Tomasz, A. Peptidoglycan N-acetylglucosamine deacetylase, a putative virulence factor in Streptococcus pneumoniae. Infect. Immun. 2002, 70, 7176–7178. [Google Scholar] [CrossRef]
- Meeske, A.J.; Riley, E.P.; Robins, W.P.; Uehara, T.; Mekalanos, J.J.; Kahne, D.; Walker, S.; Kruse, A.C.; Bernhardt, T.G.; Rudner, D.Z. SEDS proteins are a widespread family of bacterial cell wall polymerases. Nature 2016, 537, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Price, C.T.D.; Bukka, A.; Cynamon, M.; Graham, J.E. Glycine betaine uptake by the proXVWZ ABC transporter contributes to the ability of Mycobacterium tuberculosis to initiate growth in human macrophages. J. Bacteriol. 2008, 190, 3955–3961. [Google Scholar] [CrossRef]
- Tanaka, K.J.; Song, S.; Mason, K.; Pinkett, H.W. Selective substrate uptake: The role of ATP-binding cassette ( ABC ) importers in pathogenesis. BBA Biomembr. 2018, 1860, 868–877. [Google Scholar] [CrossRef]
- Orelle, C.; Mathieu, K.; Jault, J.M. Multidrug ABC transporters in bacteria. Res. Microbiol. 2019, 170, 381–391. [Google Scholar] [CrossRef]
- Kumar, S.; Mukherjee, M.M.; Varela, M.F. Modulation of Bacterial Multidrug Resistance Efflux Pumps of the Major Facilitator Superfamily. Int. J. Bacteriol. 2013, 2013, 204141. [Google Scholar] [CrossRef] [PubMed]
- Cotrim, C.A.; Jarrott, R.J.; Martin, J.L.; Drew, D. A structural overview of the zinc transporters in the cation diffusion facilitator family. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Jack, D.L.; Yang, N.M.; Saier, M.H. The drug/metabolite transporter superfamily. Eur. J. Biochem. 2001, 268, 3620–3639. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Wolz, C. Phages of Staphylococcus aureus and their impact on host evolution. Infect. Genet. Evol. 2014, 21, 593–601. [Google Scholar] [CrossRef]
- Romero, P.; López, R.; García, E. Genomic organization and molecular analysis of the inducible prophage EJ-1, a mosaic myovirus from an atypical Pneumococcus. Virology 2004, 322, 239–252. [Google Scholar] [CrossRef]
- Fortier, L.C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Menouni, R.; Hutinet, G.; Petit, M.A.; Ansaldi, M. Bacterial genome remodeling through bacteriophage recombination. FEMS Microbiol. Lett. 2015, 362, 1–10. [Google Scholar] [CrossRef]
- Deghorain, M.; Van Melderen, L. The Staphylococci phages family: An overview. Viruses 2012, 4, 3316–3335. [Google Scholar]
- Feiner, R.; Argov, T.; Rabinovich, L.; Sigal, N.; Borovok, I.; Herskovits, A.A. A new perspective on lysogeny: Prophages as active regulatory switches of bacteria. Nat. Rev. Microbiol. 2015, 13, 641–650. [Google Scholar] [CrossRef]
- Penadés, J.R.; Chen, J.; Quiles-Puchalt, N.; Carpena, N.; Novick, R.P. Bacteriophage-mediated spread of bacterial virulence genes. Curr. Opin. Microbiol. 2015, 23, 171–178. [Google Scholar] [CrossRef]
- Hall, R.J.; Whelan, F.J.; McInerney, J.O.; Ou, Y.; Domingo-Sananes, M.R. Horizontal Gene Transfer as a Source of Conflict and Cooperation in Prokaryotes. Front. Microbiol. 2020, 11, 1569. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.M.; Dunning Hotopp, J.C. Mobile elements and viral integrations prompt considerations for bacterial DNA integration as a novel carcinogen. Cancer Lett. 2014, 352, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Fatoba, D.O.; Amoako, D.G.; Akebe, A.L.K.; Ismail, A.; Essack, S.Y. Genomic analysis of antibiotic-resistant Enterococcus spp. reveals novel Enterococci strains and the spread of plasmid-borne Tet(M), Tet(L) and Erm(B) genes from chicken litter to agricultural soil in South Africa. J. Environ. Manag. 2022, 302, 114101. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wong, J.J.W.; Edwards, R.A.; Manchak, J.; Frost, L.S.; Glover, J.N.M. Structural basis of specific TraD–TraM recognition during F plasmid-mediated bacterial conjugation. Mol. Microbiol. 2008, 70, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, Y.; Tao, F.; Zhang, J.; Copetti, D.; Kudrna, D.; Talag, J.; Lee, S.; Wing, R.A.; Fan, C. DNA methylation changes facilitated evolution of genes derived from Mutator-like transposable elements. Genome Biol. 2016, 17, 92. [Google Scholar] [CrossRef] [PubMed]
- Kommineni, S.; Bretl, D.J.; Lam, V.; Chakraborty, R.; Hayward, M.; Simpson, P.; Cao, Y.; Bousounis, P.; Kristich, C.J.; Salzman, N.H. Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract. Nature 2015, 526, 719–722. [Google Scholar] [CrossRef]
- Li, F.; Alvarez-Martinez, C.; Chen, Y.; Choi, K.J.; Yeo, H.J.; Christie, P.J. Enterococcus faecalis PrgJ, a VirB4-Like ATPase, Mediates pCF10 conjugative transfer through substrate binding. J. Bacteriol. 2012, 194, 4041–4051. [Google Scholar] [CrossRef]
- Bhatty, M.; Camacho, M.I.; Gonzalez-Rivera, C.; Frank, K.L.; Dale, J.L.; Manias, D.A.; Dunny, G.M.; Christie, P.J. PrgU: A suppressor of sex pheromone toxicity in Enterococcus faecalis. Mol. Microbiol. 2017, 103, 398–412. [Google Scholar] [CrossRef]
- Alekseev, A.; Pobegalov, G.; Morozova, N.; Vedyaykin, A.; Cherevatenko, G.; Yakimov, A.; Baitin, D.; Khodorkovskii, M. A new insight into RecA filament regulation by RecX from the analysis of conformation-specific interactions. bioRxiv 2022. [Google Scholar] [CrossRef]
- Vellani, T.S.; Myers, R.S. Bacteriophage SPP1 Chu is an alkaline exonuclease in the SynExo family of viral two-component recombinases. J. Bacteriol. 2003, 185, 2465–2474. [Google Scholar] [CrossRef]
- Rodríguez-Rubio, L.; Gutiérrez, D.; Martínez, B.; Rodríguez, A.; Götz, F.; García, P. The tape measure protein of the Staphylococcus aureus bacteriophage vB_SauS-phiIPLA35 has an active muramidase domain. Appl. Environ. Microbiol. 2012, 78, 6369–6371. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; Alqarni, M.; Stockdale, S.; Spinelli, S.; Feyereisen, M.; Cambillau, C.; Van Sinderen, D. Functional and structural dissection of the tape measure protein of Lactococcal phage TP901-1. Sci. Rep. 2016, 6, 36667. [Google Scholar] [CrossRef]
- Durante-Rodríguez, G.; Mancheño, J.M.; Díaz, E.; Carmona, M. Refactoring the λ phage lytic/lysogenic decision with a synthetic regulator. Microbiologyopen 2016, 5, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Veesler, D.; Spinelli, S.; Mahony, J.; Lichière, J.; Blangy, S.; Bricogne, G.; Legrand, P.; Ortiz-Lombardia, M.; Campanacci, V.; Van Sinderen, D.; et al. Structure of the phage TP901-1 1.8 MDa baseplate suggests an alternative host adhesion mechanism. Proc. Natl. Acad. Sci. USA 2012, 109, 8954–8958. [Google Scholar] [CrossRef] [PubMed]
- Kostyuchenko, V.A.; Leiman, P.G.; Chipman, P.R.; Kanamaru, S.; Van Raaij, M.J.; Arisaka, F.; Mesyanzhinov, V.V.; Rossmann, M.G. Three-dimensional structure of bacteriophage T4 baseplate. Nat. Struct. Mol. Biol. 2003, 10, 688–693. [Google Scholar] [CrossRef]
- Quintela-Baluja, M.; Böhme, K.; Fernández-No, I.C.; Morandi, S.; Alnakip, M.E.; Caamaño-Antelo, S.; Barros-Velázquez, J.; Calo-Mata, P. Characterization of different food-isolated Enterococcus strains by MALDI-TOF mass fingerprinting. Electrophoresis 2013, 34, 2240–2250. [Google Scholar] [CrossRef]
- Shridhar, P.B.; Amachawadi, R.G.; Tokach, M.; Patel, I.; Gangiredla, J.; Mammel, M.; Nagaraja, T.G. Whole genome sequence analyses-based assessment of virulence potential and antimicrobial susceptibilities and resistance of Enterococcus faecium strains isolated from commercial swine and cattle probiotic products. J. Anim. Sci. 2022, 100, skac030. [Google Scholar] [CrossRef] [PubMed]
- Yuksekdag, Z.; Ahlatcı, N.S.; Hajikhani, R.; Darilmaz, D.O.; Beyatli, Y. Safety and metabolic characteristics of 17 Enterococcus faecium isolates. Arch. Microbiol. 2021, 203, 5683–5694. [Google Scholar] [CrossRef]
- Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Cañas, B.; Rama, J.L.R.; Villa, T.G.; Calo-Mata, P. Characterization of Bacteriophage Peptides of Pathogenic Streptococcus by LC-ESI-MS/MS: Bacteriophage Phylogenomics and Their Relationship to Their Host. Front. Microbiol. 2020, 11, 1241. [Google Scholar] [CrossRef]
- Carrera, M.; Cañas, B.; Gallardo, J.M. The sarcoplasmic fish proteome: Pathways, metabolic networks and potential bioactive peptides for nutritional inferences. J. Proteom. 2013, 78, 211–220. [Google Scholar] [CrossRef]
- Calo-Mata, P.; Carrera, M.; Böhme, K.; Barros-Velázquez, J. Novel Peptide Biomarker Discovery for Detection and Identification of Bacterial Pathogens by Bioactives compounds from seafood by-products. Artic. J. Anal. Bioanal. Tech. 2016, 7, 296. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
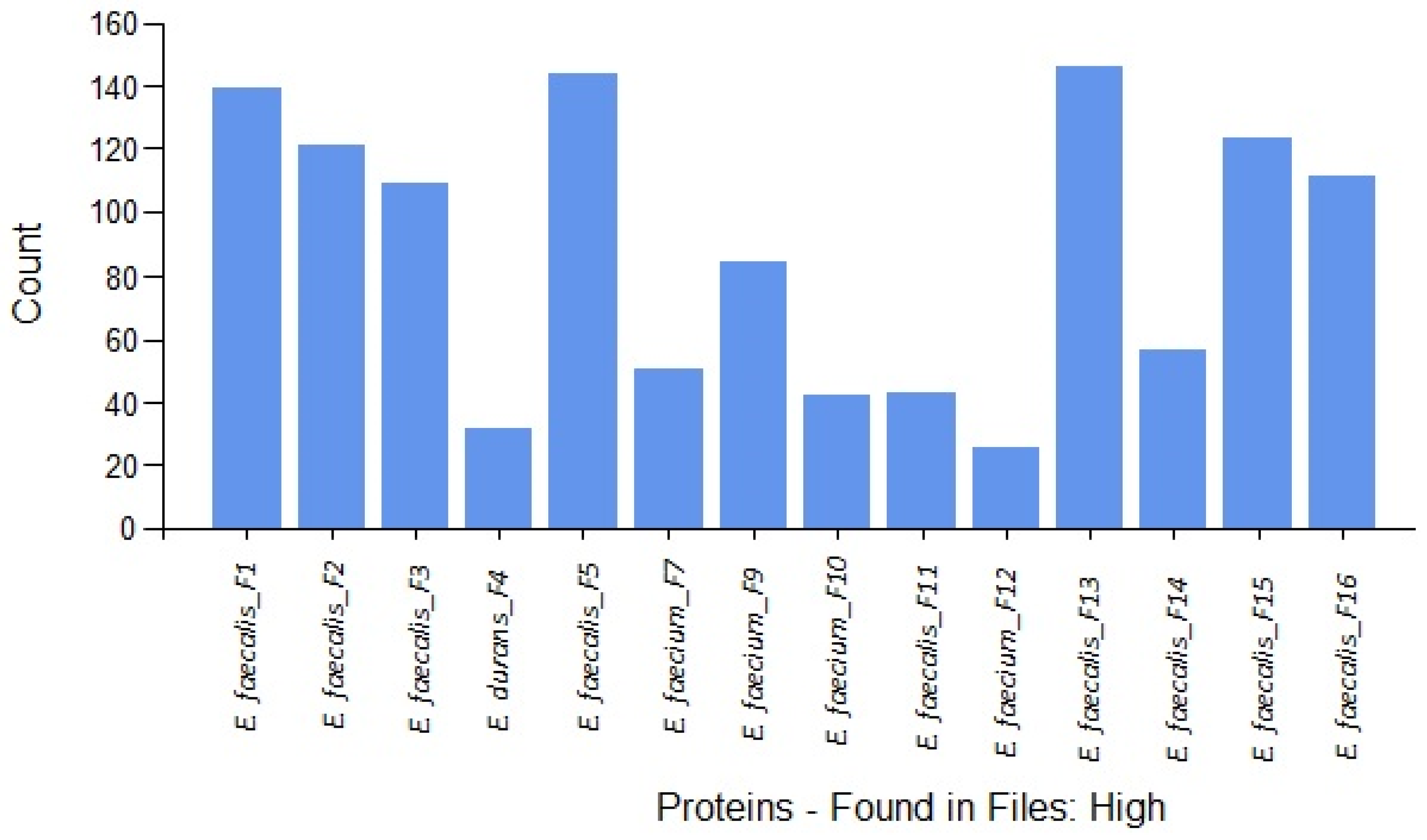
| Sample | Species | Strain | Source | GenBank Accession Number | Total Virulence Factor Peptides |
|---|---|---|---|---|---|
| F1 | Enterococcus faecalis | ISPA118 | Milk | OP113834 | 39 |
| F2 | Enterococcus faecalis | ISPA 10 | Dairy product | KC510240 | 44 |
| F3 | Enterococcus faecalis | ISPA T07 | Milk | - * | 37 |
| F4 | Enterococcus durans | CECT 411 | Milk powder | KC510230 | 5 |
| F5 | Enterococcus faecalis | ISPA 28 | Cheese | OP113835 | 50 |
| F7 | Enterococcus faecium | ISPA FRP2 | Cheese | KC510246 | 14 |
| F9 | Enterococcus faecium | ISPA ID114 | Milk | KC510249 | 19 |
| F10 | Enterococcus faecium | ISPA G07 | Cheese | KC510247 | 4 |
| F11 | Enterococcus faecalis | ISPA ID119 | Dairy product | KC510243 | 7 |
| F12 | Enterococcus faecium | ISPA G010 | Cheese | KC510248 | 2 |
| F13 | Enterococcus faecalis | MORA 13 | Dairy product | KC510244 | 32 |
| F14 | Enterococcus faecalis | ISPA ID 116 | Milk | OP113836 | 12 |
| F15 | Enterococcus faecalis | ISPA 148 | Dairy product | KC510245 | 25 |
| F16 | Enterococcus faecalis | ISPA 89 | Dairy product | KC510242 | 20 |
| Total peptides | 310 | ||||
| Function | Protein |
|---|---|
| Antibiotic resistance | TetR family transcriptional regulator |
| TetR family transcriptional regulator | |
| N-acetyltransferase domain-containing protein | |
| PadR domain-containing protein | |
| GNAT family acetyltransferase | |
| Cass2 domain-containing protein | |
| MarR family transcriptional regulator | |
| Penicillin-binding protein | |
| Daunorubicin resistance protein DrrC | |
| OmpR/PhoB-type domain-containing protein | |
| PASTA domain-containing protein | |
| VanY D-Ala-D-Ala carboxypeptidase | |
| Additional resistances and tolerances | Cold-shock protein |
| General stress protein | |
| CsbD-like protein | |
| YitT family protein | |
| Tellurite resistance protein TelA | |
| MerR family transcriptional regulator | |
| SugE protein | |
| Chaperone protein DnaK | |
| Antimicrobial compounds | Lantibiotic biosynthesis protein |
| Type 2 lantipeptide synthetase LanM | |
| LanM family lanthionine synthetase | |
| Thiopeptide-type bacteriocin biosynthesis domain protein | |
| Bacteriocin Protein | |
| Radical SAM additional 4Fe4S-binding SPASM domain-containing protein | |
| Toxin | Type II toxin–antitoxin system PemK/MazF family toxin |
| Type II toxin–antitoxin system RelE/ParE family toxin | |
| Toxin–antitoxin system, antitoxin component, AbrB family | |
| Toxin PIN | |
| Exfoliative toxin A/B | |
| Prevent-host-death family antitoxin (Phd antitoxin) | |
| LXG domain-containing protein | |
| Colonization and immune evasion factors | Internalin |
| Sortase | |
| LPXTG cell wall anchor domain-containing protein | |
| Adhesin BspA | |
| Fibronectin/fibrinogen-binding protein | |
| Collagen-binding protein | |
| Ig domain-containing protein | |
| DUF4097 domain-containing protein | |
| Flagellar hook-associated protein 2 | |
| Fn3-like domain-containing protein | |
| Fimbrial isopeptide formation D2 domain-containing protein | |
| SpaA domain-containing protein | |
| Endopeptidase NlpC/P60 family protein | |
| M24/M78/M28/M20/M25/M40 family peptidases | |
| Peptidase T | |
| DD-transpeptidase | |
| Dipeptidase PepV | |
| Endopeptidase La | |
| S8/S9/S74 Family Peptidases | |
| Dipeptidyl aminopeptidase | |
| Peptidase U32 | |
| Peptidase S74 | |
| Proline dipeptidase | |
| Oligoendopeptidase PepF/M3 family protein | |
| Peptidase C51 | |
| ImmA/IrrE family metallo-endopeptidase | |
| Isoaspartyl dipeptidase | |
| Signal peptidase I | |
| Immune inhibitor A | |
| ClpA protease | |
| ClpC protease | |
| Zinc protease | |
| Capsular polysaccharide biosynthesis protein CpsC | |
| N-acetylmuramoyl-L-alanine amidase | |
| LysM domain protein | |
| Hemolysin III family channel protein | |
| Autolysin modifier protein | |
| Mga domain-containing protein | |
| Toxin secretion/phage lysis holin | |
| Type VII secretion protein EssC | |
| LysR family transcriptional regulator | |
| LytTR family transcriptional regulator | |
| HTH-type transcriptional regulator KdgR | |
| ArpU family transcriptional regulator | |
| Competence protein ComEA helix-hairpin-helix repeat region | |
| Control of competence regulator ComK, YlbF/YmcA | |
| Regulatory protein YlbF | |
| Spore coat protein | |
| Sporulation protein YjcZ | |
| Restriction endonuclease type IV, Mrr | |
| SfeI restriction endonuclease | |
| Type-2 restriction enzyme | |
| O-antigen ligase | |
| Methyl-accepting chemotaxis protein (MCP) signaling domain | |
| N-acetylglucosamine-6-phosphate deacetylase | |
| Glycosyl/glycerophosphate transferases involved in teichoic acid biosynthesis TagF/TagB/EpsJ/RodC | |
| ABC transporters | Copper ABC transporter permease |
| Ferrichrome ABC transporter FhuC | |
| Cobalt ABC transporter permease | |
| Lantibiotic protection ABC transporter | |
| Multidrug ABC transporter | |
| Spermidine/putrescine ABC transporter | |
| Glycine betaine ABC transporter | |
| Peptide ABC transporter | |
| Amino acid ABC transporter | |
| Excinuclease ABC subunit A | |
| Sulfate ABC transporter ATP-binding protein | |
| C4-dicarboxylate ABC transporter | |
| Multiple sugar ABC transporter substrate-binding protein | |
| Sugar ABC transporter | |
| Carbohydrate ABC transporter substrate-binding protein | |
| Heme ABC transporter | |
| Thiol reductant ABC exporter subunit CydC | |
| Other transporters | Major facilitator superfamily (MFS) transporter |
| Multidrug resistance MFS transporter | |
| Cation diffusion facilitator family transporter | |
| EamA/RhaT family transporter | |
| Copper-transporting ATPase CopB | |
| Alternative virulence factors | Transposase |
| Tnp-DDE superfamily | |
| IS4 family transposase | |
| Transposase InsI for insertion sequence element IS30C | |
| IS30/IS4/IS6 family transposaseS | |
| Conjugative transposon protein | |
| Conjugal transfer protein TraG | |
| Mutator family transposase | |
| Tyrosine-type recombinase/integrase | |
| Enterococcus faecalis plasmid pPD1 bacI | |
| Putative plasmid replication protein | |
| PrgI family protein | |
| Pheromone response system RNA-binding regulator PrgU | |
| Regulatory protein RecX | |
| CRISPR-associated endonuclease Cas9 | |
| CRISPR-associated endonuclease Cas10 | |
| CRISPR-associated endoribonuclease Cas2 | |
| YqaJ domain-containing protein | |
| Luciferase family oxidoreductase, group 1 | |
| Phage proteins | Phage capsid proteins |
| Phage tail proteins | |
| Phage portal protein | |
| Phage/plasmid primase, P4 family domain protein | |
| PBSX family phage terminase | |
| Phage terminase | |
| Phage integraseç | |
| Cro/CI family transcriptional regulator | |
| BppU family phage baseplate upper protein | |
| phage infection protei YhgE |
| Function | Total Virulence Factor Peptides |
|---|---|
| Colonization and immune evasion factors | 104 |
| ABC transporters | 70 |
| Other transporters | 8 |
| Alternative virulence factors | 32 |
| Phage proteins | 49 |
| Antibiotic resistance | 21 |
| Additional resistances and tolerances | 11 |
| Antimicrobial compounds | 6 |
| Toxins | 9 |
| Total Peptides | 310 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abril, A.G.; Quintela-Baluja, M.; Villa, T.G.; Calo-Mata, P.; Barros-Velázquez, J.; Carrera, M. Proteomic Characterization of Virulence Factors and Related Proteins in Enterococcus Strains from Dairy and Fermented Food Products. Int. J. Mol. Sci. 2022, 23, 10971. https://doi.org/10.3390/ijms231810971
Abril AG, Quintela-Baluja M, Villa TG, Calo-Mata P, Barros-Velázquez J, Carrera M. Proteomic Characterization of Virulence Factors and Related Proteins in Enterococcus Strains from Dairy and Fermented Food Products. International Journal of Molecular Sciences. 2022; 23(18):10971. https://doi.org/10.3390/ijms231810971
Chicago/Turabian StyleAbril, Ana G., Marcos Quintela-Baluja, Tomás G. Villa, Pilar Calo-Mata, Jorge Barros-Velázquez, and Mónica Carrera. 2022. "Proteomic Characterization of Virulence Factors and Related Proteins in Enterococcus Strains from Dairy and Fermented Food Products" International Journal of Molecular Sciences 23, no. 18: 10971. https://doi.org/10.3390/ijms231810971
APA StyleAbril, A. G., Quintela-Baluja, M., Villa, T. G., Calo-Mata, P., Barros-Velázquez, J., & Carrera, M. (2022). Proteomic Characterization of Virulence Factors and Related Proteins in Enterococcus Strains from Dairy and Fermented Food Products. International Journal of Molecular Sciences, 23(18), 10971. https://doi.org/10.3390/ijms231810971