

Simultaneous Probing of Metabolism and Oxygenation of Tumors In Vivo Using FLIM of NAD(P)H and PLIM of a New Polymeric Ir(III) Oxygen Sensor

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
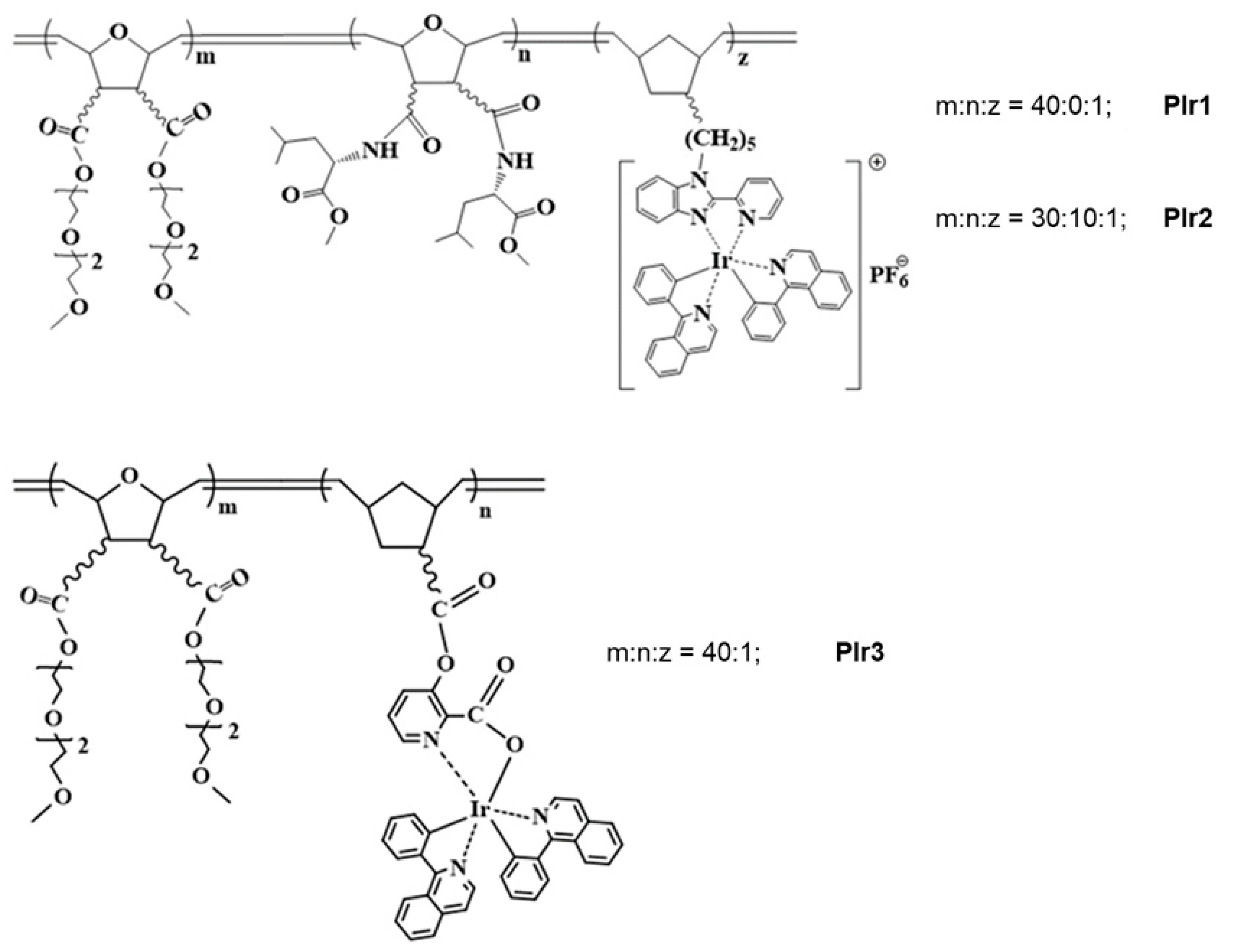
2.1. Synthesis of Polymeric Probes PIr1–PIr3
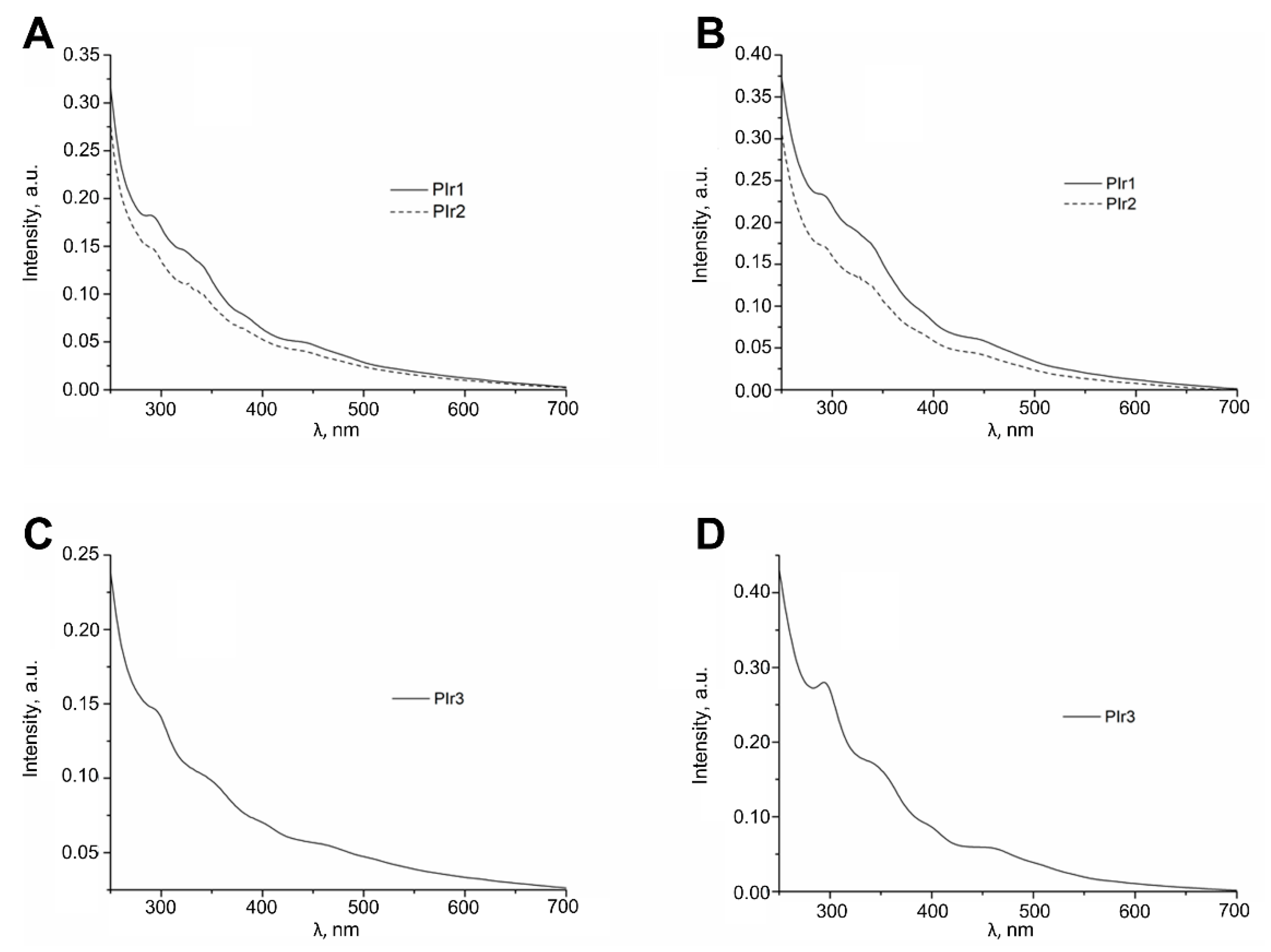
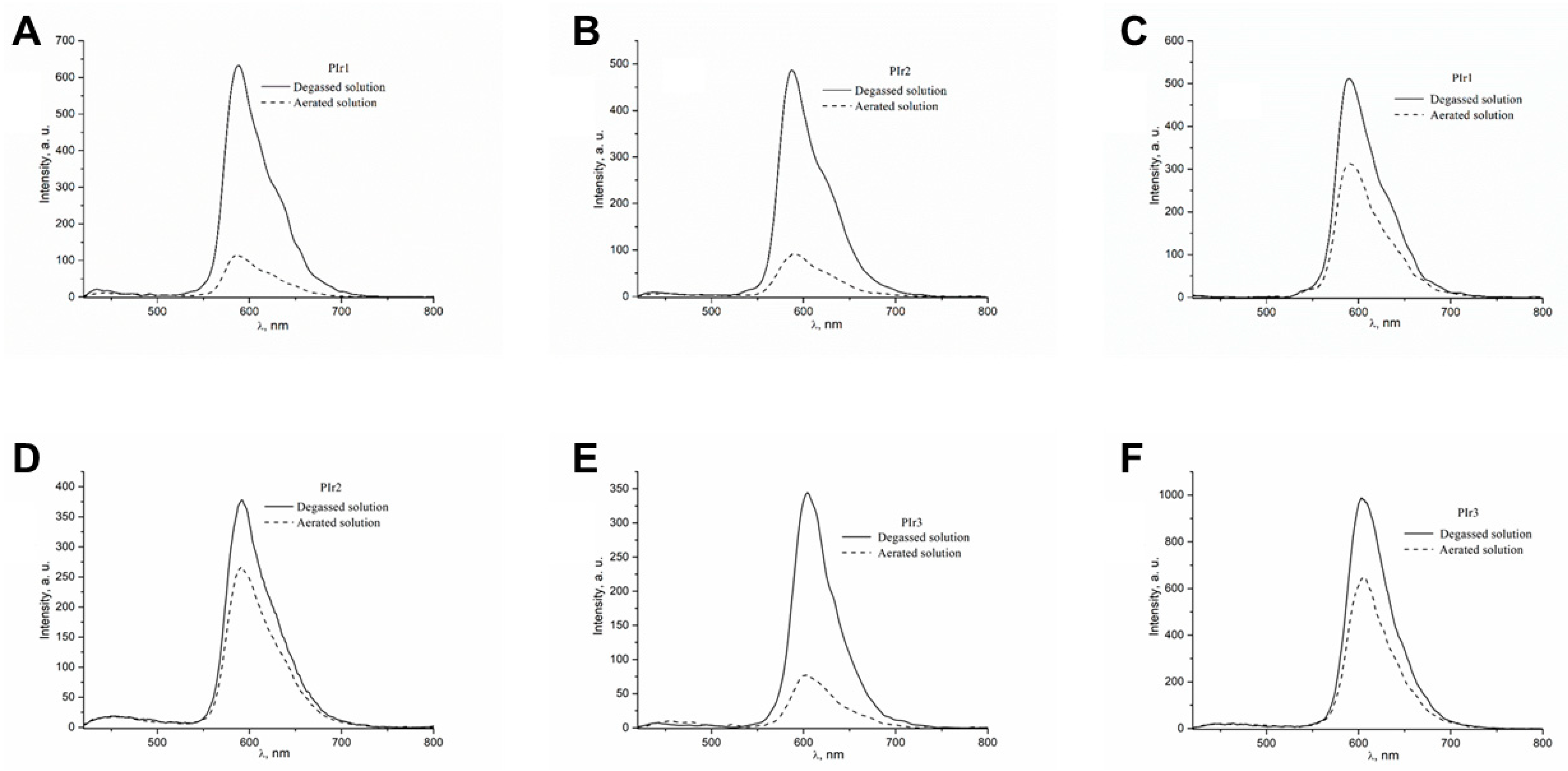
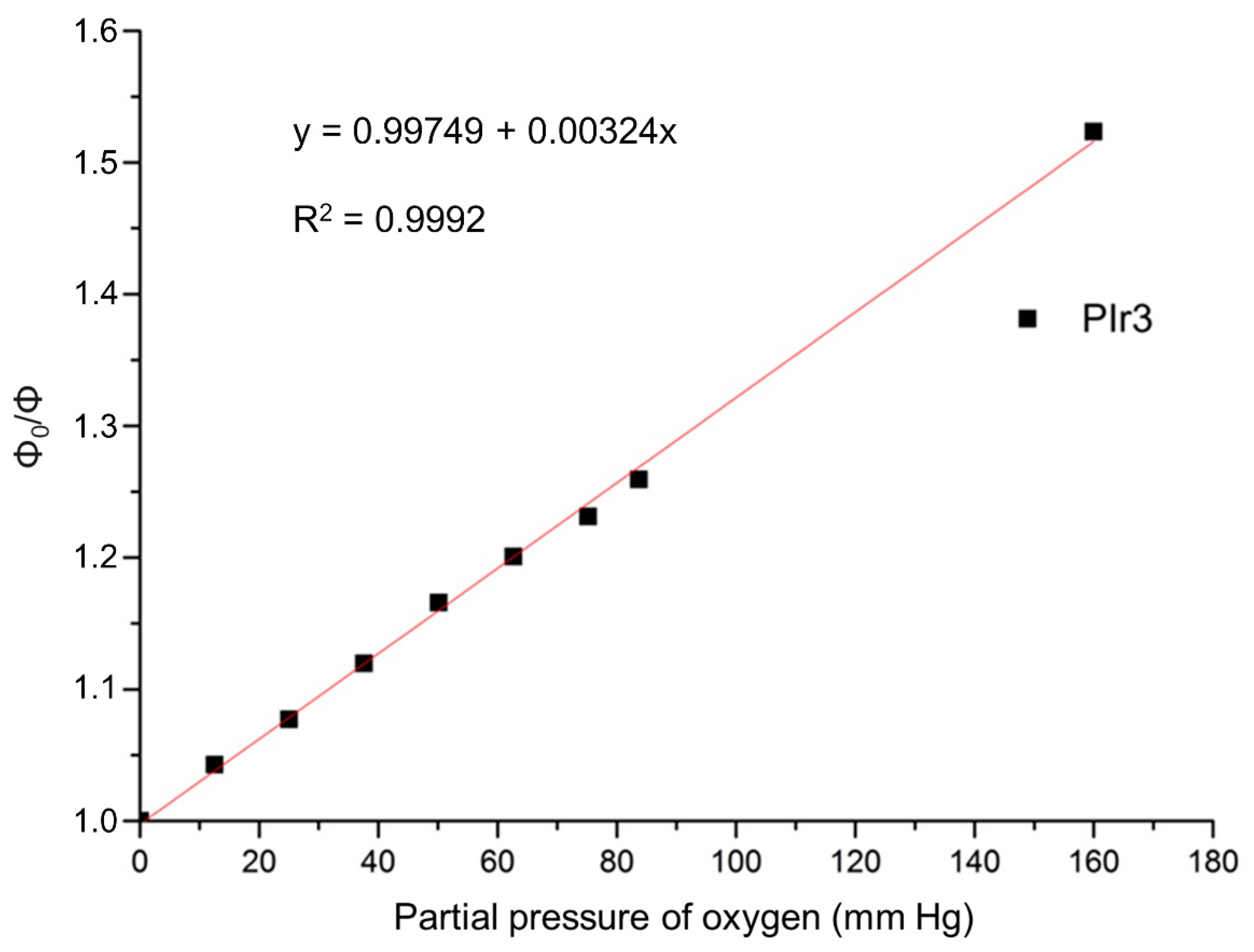
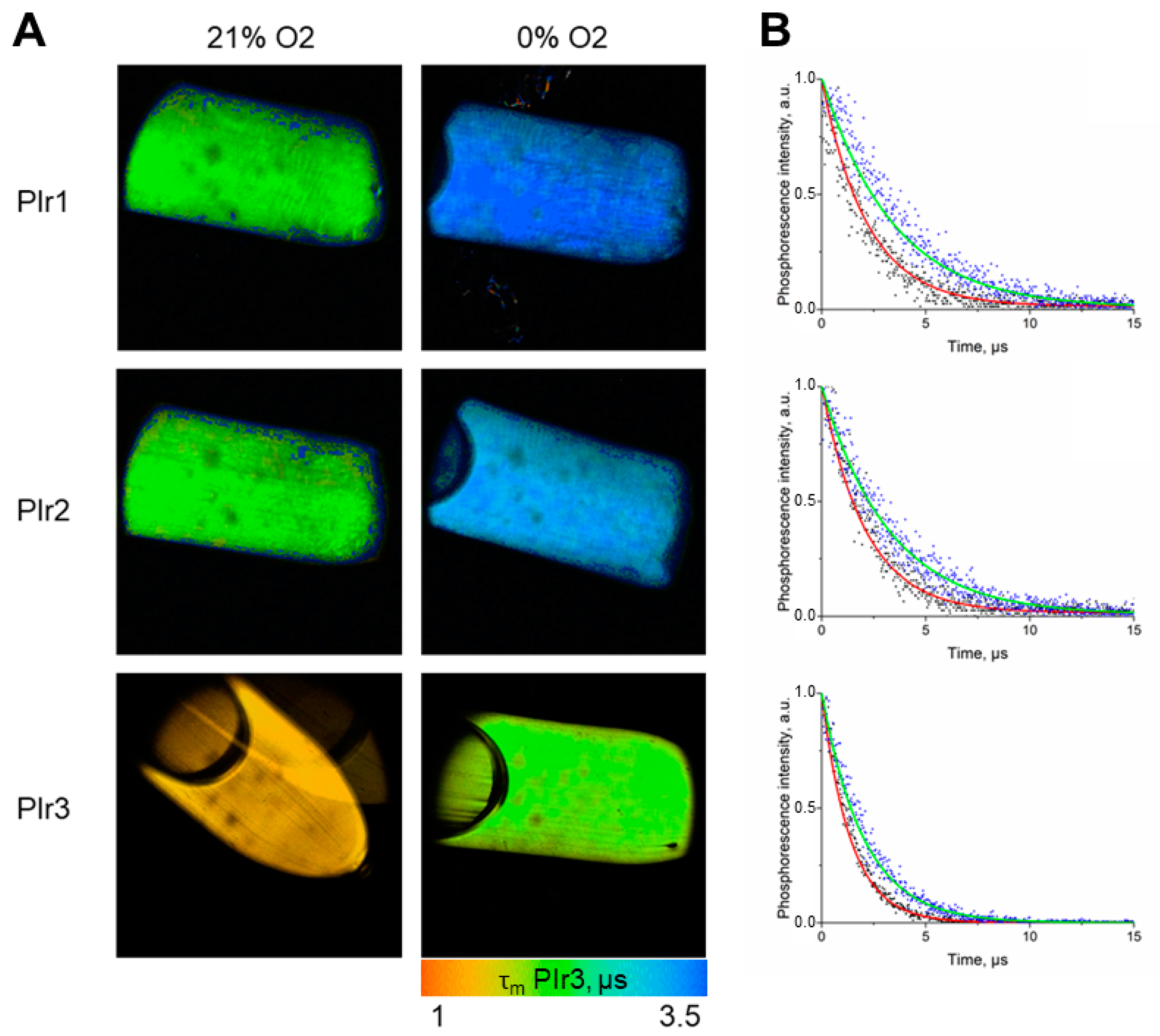
2.2. Photophysical Properties of the PIr1–PIr3 Probes in Solutions
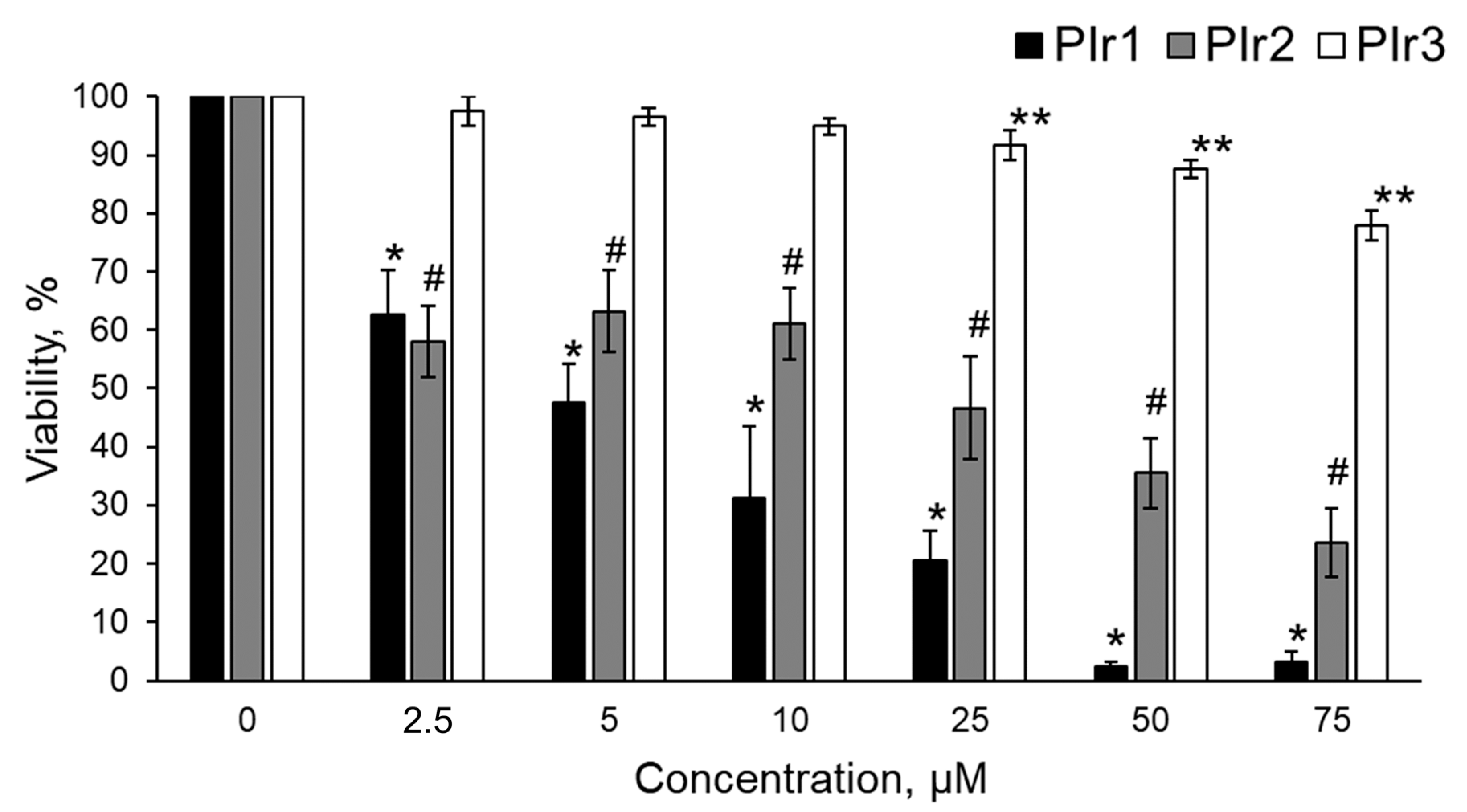
2.3. Cytotoxicity of Polymeric Probes PIr1–PIr3
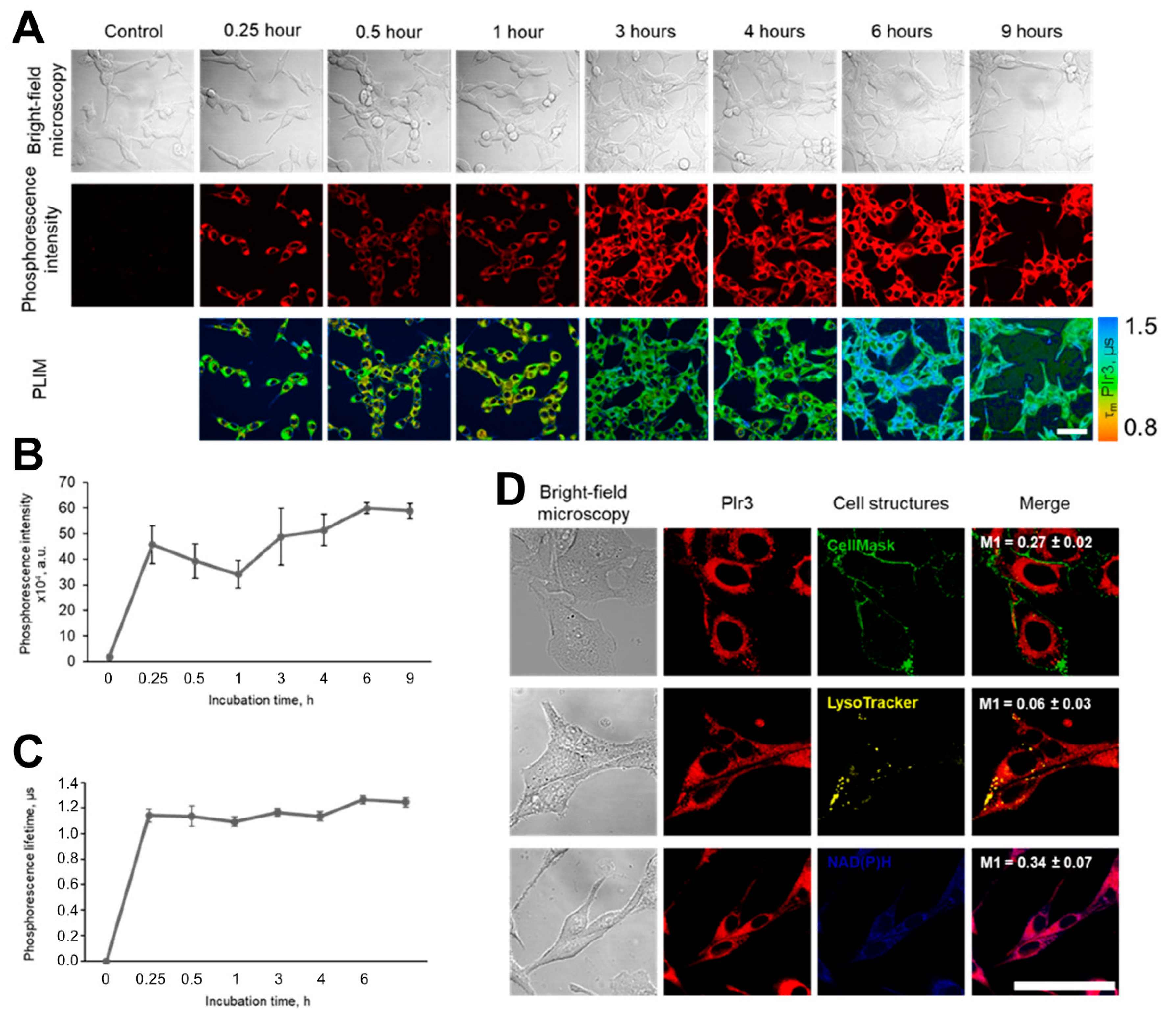
2.4. Cellular Uptake and Distribution of PIr3
2.5. Simultaneous Oxygen and Metabolic Imaging In Vitro
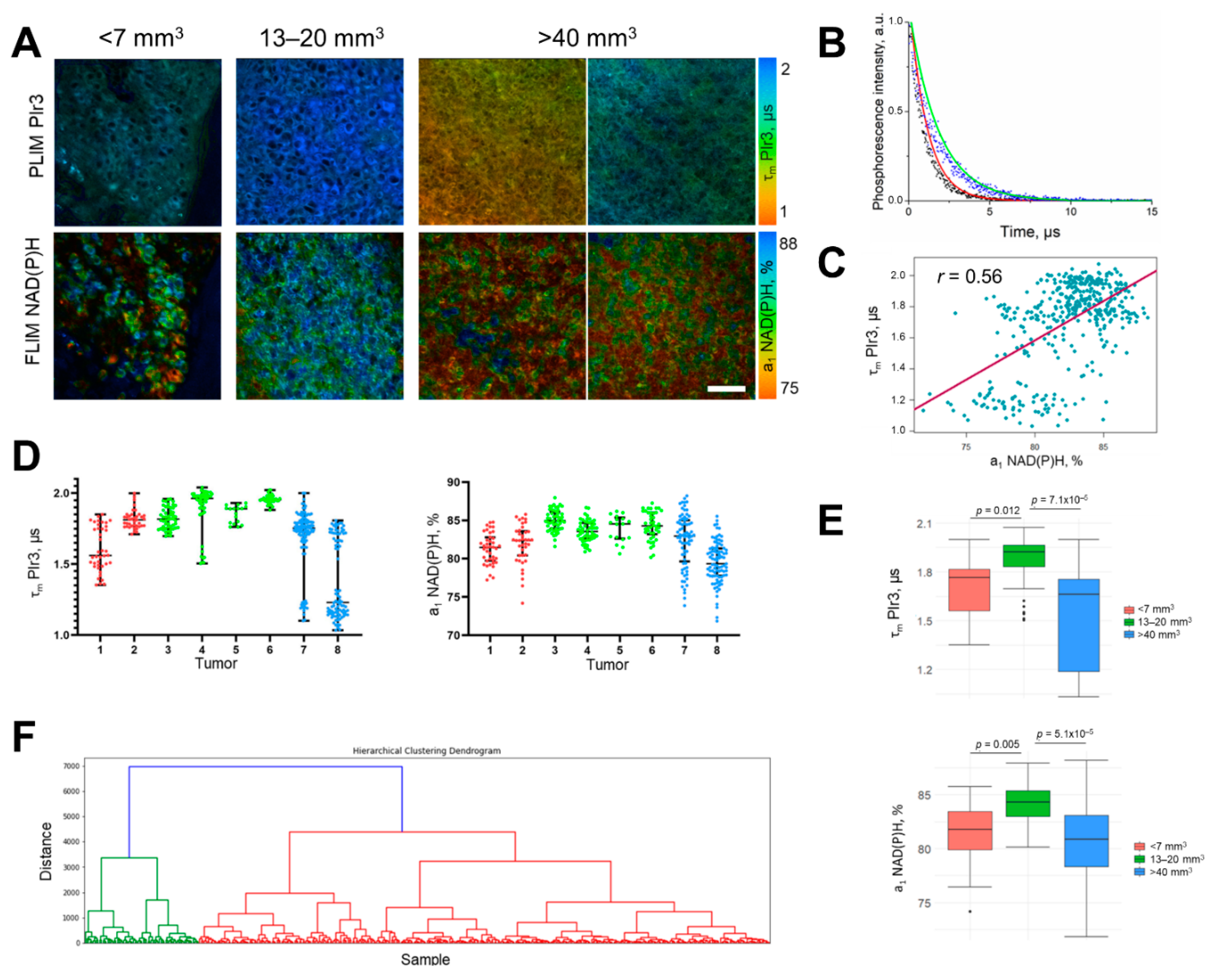
2.6. In Vivo Study of Oxygen and Metabolism in Tumors
2.7. In Vivo Microvascular Network Assessment by OCT-A
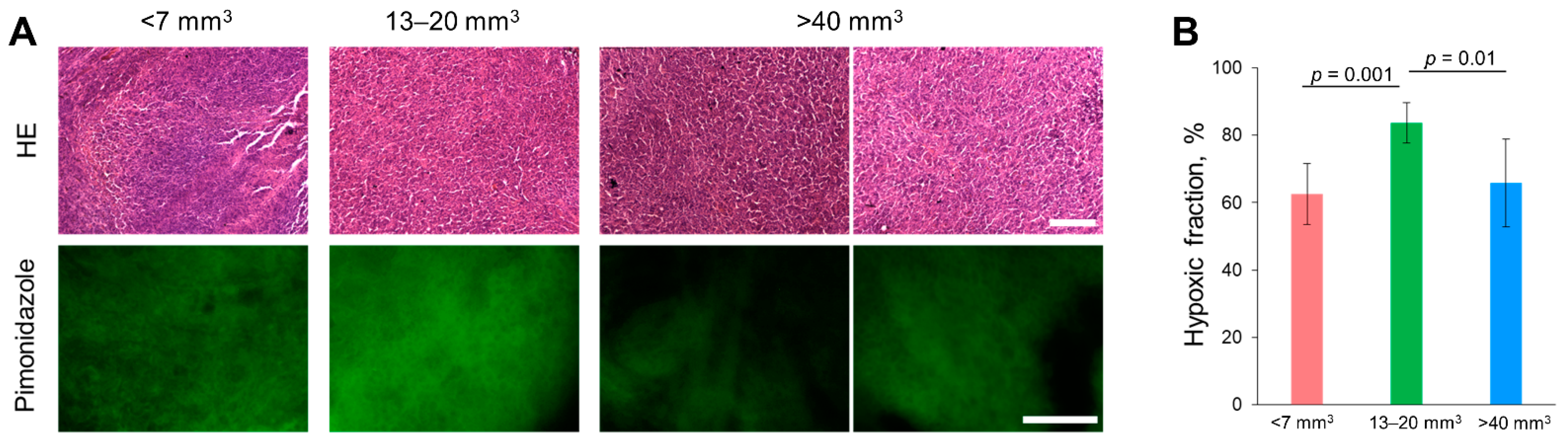
2.8. Histopathological and IHC Analysis of Tumors
3. Discussion
4. Materials and Methods
4.1. Synthesis of Polymeric Probes PIr1–PIr3
4.2. Absorption and Emission Spectra
4.3. Phosphorescence Lifetime Measurements
4.4. Cell Culturing
4.5. MTT Assay
4.6. Laser Scanning Microscopy
4.7. Subcellular Localization Assay
4.8. Tumor Model
4.9. Simultaneous PLIM and FLIM
4.10. Optical Coherence Tomography Angiography (OCT-A)
4.11. Histopathology and Immunohistochemistry
4.12. Clustering Analysis
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaupel, P.; Flood, A.B.; Swartz, H.M. Oxygenation Status of Malignant Tumors vs. Normal Tissues: Critical Evaluation and Updated Data Source Based on Direct Measurements with pO2 Microsensors. Appl. Magn. Reson. 2021, 52, 1451–1479. [Google Scholar] [CrossRef]
- Hompland, T.; Fjeldbo, C.S.; Lyng, H. Tumor Hypoxia as a Barrier in Cancer Therapy: Why Levels Matter. Cancers 2021, 13, 499. [Google Scholar] [CrossRef] [PubMed]
- Paredes, F.; Williams, H.C.; San Martin, A. Metabolic adaptation in hypoxia and cancer. Cancer Lett. 2021, 502, 133–142. [Google Scholar] [CrossRef]
- Kim, J.; DeBerardinis, R.J. Mechanisms and Implications of Metabolic Heterogeneity in Cancer. Cell Metab. 2019, 30, 434–446. [Google Scholar] [CrossRef]
- Vaupel, P.; Multhoff, G. Revisiting the Warburg effect: Historical dogma versus current understanding. J. Physiol. 2021, 599, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, S.; Breymaier, J.; Schaffer, P.; Calzia, E.; Shcheslavskiy, V.; Becker, W.; Rueck, A. Correlative NAD(P)H-FLIM and oxygen sensing-PLIM for metabolic mapping. J. Biophotonics 2016, 9, 800–811. [Google Scholar] [CrossRef]
- Kolenc, O.I.; Quinn, K.P. Evaluating Cell Metabolism Through Autofluorescence Imaging of NAD(P)H and FAD. Antioxid. Redox Signal. 2019, 30, 875–889. [Google Scholar] [CrossRef]
- Shirmanova, M.V.; Shcheslavskiy, V.I.; Lukina, M.M.; Becker, W.; Zagaynova, E.V. Exploring tumor metabolism with time-resolved fluorescence methods: From single cells to a whole tumor. In Multimodal Optical Diagnostics of Cancer, 1st ed.; Tuchin, V., Popp, J., Zakharov, V., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 133–155. [Google Scholar]
- Shcheslavskiy, V.I.; Neubauer, A.; Bukowiecki, R.; Dinter, F.; Becker, W. Combined fluorescence and phosphorescence lifetime imagin. Appl. Phys. Lett. 2016, 108, 091111. [Google Scholar] [CrossRef]
- Jahn, K.; Buschmann, V.; Hille, C. Simultaneous Fluorescence and Phosphorescence Lifetime Imaging Microscopy in Living Cells. Sci. Rep. 2015, 5, 14334. [Google Scholar] [CrossRef]
- Lukina, M.; Orlova, A.; Shirmanova, M.; Shirokov, D.; Pavlikov, A.; Neubauer, A.; Studier, H.; Becker, W.; Zagaynova, E.; Yoshihara, T.; et al. Interrogation of metabolic and oxygen states of tumors with fiber-based luminescence lifetime spectroscopy. Opt. Lett. 2017, 42, 731–734. [Google Scholar] [CrossRef]
- Yoshihara, T.; Hirakawa, Y.; Hosaka, M.; Nangaku, M.; Tobita, S. Oxygen imaging of living cells and tissues using luminescent molecular probes. J. Photochem. Photobiol. C Photochem. Rev. 2017, 30, 71–95. [Google Scholar] [CrossRef]
- Yoshihara, T.; Hosaka, M.; Terata, M.; Ichikawa, K.; Murayama, S.; Tanaka, A.; Mori, M.; Itabashi, H.; Takeuchi, T.; Tobita, S. Intracellular and in vivo oxygen sensing using phosphorescent Ir(III) complexes with a modified acetylacetonato ligand. Anal. Chem. 2015, 87, 2710–2717. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, T.; Murayama, S.; Tobita, S. Ratiometric molecular probes based on dual emission of a blue fluorescent coumarin and a red phosphorescent cationic iridium(III) complex for intracellular oxygen sensing. Sensors 2015, 15, 13503–13521. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, X.; Mao, H.; Wu, W.; Liu, B.; Jiang, X. Hypoxia-specific ultrasensitive detection of tumours and cancer cells in vivo. Nat. Commun. 2015, 6, 5834. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, S.; Ge, L.; Wu, W.; Jiang, X. Translatable high drug loading drug delivery systems based on biocompatible polymer nanocarriers. Biomacromolecules 2018, 19, 1732–1745. [Google Scholar] [CrossRef]
- Miller, T.; Hill, A.; Uezguen, S.; Weigandt, M.; Goepferich, A. Analysis of immediate stress mechanisms upon injection of polymeric micelles and related colloidal drug carriers: Implications on drug targeting. Biomacromolecules 2012, 13, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.-H.; Oh, Y.H.; Shin, I.-S.; Hong, J.-I. New approach toward fast response light-emitting electrochemical cells based on neutral iridium complexes via cation transport. Adv. Funct. Mater. 2009, 19, 711–717. [Google Scholar] [CrossRef]
- Lamansky, S.; Djurovich, P.; Murphy, D.; Abdel-Razzaq, F.; Kwong, R.; Tsyba, I.; Bortz, M.; Mui, B.; Bau, R.; Thompson, M.E. Synthesis and characterization of phosphorescent cyclometalated iridium complexes. Inorg. Chem. 2001, 40, 1704–1711. [Google Scholar] [CrossRef]
- Martínez-Alonso, M.; Cerdá, J.; Momblona, C.; Pertegás, A.; Junquera-Hernández, J.M.; Heras, A.; Rodríguez, A.M.; Espino, G.; Bolink, H.; Ortí, E. Highly stable and efficient light-emitting electrochemical cells based on cationic iridium complexes bearing arylazole ancillary ligands. Inorg. Chem. 2017, 56, 10298–10310. [Google Scholar] [CrossRef]
- Bochkarev, L.N.; Parshina, Y.P.; Gracheva, Y.V.; Kovylina, T.A.; Lermontova, S.A.; Klapshina, L.G.; Konev, A.N.; Lopatin, M.A.; Lukina, M.M.; Komarova, A.D.; et al. Red light-emitting water-soluble luminescent iridium-containing polynorbornenes. Synthesis, characterization and oxygen sensing properties in biological tissues in vivo. Molecules 2021, 26, 6349. [Google Scholar] [CrossRef]
- Montalti, M.; Credi, A.; Prodi, L.; Gandolfi, M.T. Handbook of Photochemistry, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2006; ISBN 0-8247-2377-5. [Google Scholar]
- Solomatina, A.I.; Su, S.-H.; Lukina, M.M.; Dudenkova, V.V.; Shcheslavskiy, V.I.; Wub, C.-H.; Chelushkin, P.S.; Chou, P.-T.; Koshevoy, I.O.; Tunik, S.P. Water-soluble cyclometalated platinum(II) and iridium(III) complexes: Synthesis, tuning of the photophysical properties, and in vitro and in vivo phosphorescence lifetime imaging. RSC Adv. 2018, 8, 17224–17236. [Google Scholar] [CrossRef]
- Mizukami, K.; Katano, A.; Shiozaki, S.; Yoshihara, T.; Goda, N.; Tobita, S. In vivo O2 imaging in hepatic tissues by phosphorescence lifetime imaging microscopy using Ir(III) complexes as intracellular probes. Sci. Rep. 2020, 10, 21053. [Google Scholar] [CrossRef]
- Feng, Z.; Tao, P.; Zou, L.; Gao, P.; Liu, Y.; Liu, X.; Wang, H.; Liu, S.; Dong, Q.; Li, J.; et al. Hyper-branched phosphorescent conjugated polymer dots with iridium(III) complex as the core for hypoxia imaging and photodynamic therapy. ACS Appl. Mater. Interfaces 2017, 9, 28319–28330. [Google Scholar] [CrossRef]
- Yoshihara, T.; Matsumura, N.; Tamura, T.; Shiozaki, S.; Tobita, S. Intracellular and Intravascular Oxygen Sensing of Pancreatic Tissues Based on Phosphorescence Lifetime Imaging Microscopy Using Lipophilic and Hydrophilic Iridium(III) Complexes. ACS Sens. 2022, 7, 545–554. [Google Scholar] [CrossRef]
- Kritchenkov, I.S.; Solomatina, A.I.; Kozina, D.O.; Porsev, V.V.; Sokolov, V.V.; Shirmanova, M.V.; Lukina, M.M.; Komarova, A.D.; Shcheslavskiy, V.I.; Belyaeva, T.N.; et al. Biocompatible Ir(III) Complexes as Oxygen Sensors for Phosphorescence Lifetime Imaging. Molecules 2021, 26, 2898. [Google Scholar] [CrossRef]
- Kalinina, S.; Breymayer, J.; Reeß, K.; Lilge, L.; Mandel, A.; Rück, A. Correlation of intracellular oxygen and cell metabolism by simultaneous PLIM of phosphorescent TLD1433 and FLIM of NAD(P)H. J. Biophotonics 2018, 11, e201800085. [Google Scholar] [CrossRef]
- Lukina, M.M.; Shimolina, L.E.; Kiselev, N.M.; Zagainov, V.E.; Komarov, D.V.; Zagaynova, E.V.; Shirmanova, M.V. Interrogation of tumor metabolism in tissue samples ex vivo using fluorescence lifetime imaging of NAD(P)H. Methods Appl. Fluoresc. 2020, 8, 014002. [Google Scholar] [CrossRef]
- Lukina, M.M.; Dudenkova, V.V.; Ignatova, N.I.; Druzhkova, I.N.; Shimolina, L.E.; Zagaynova, E.V.; Shirmanova, M.V. Metabolic cofactors NAD(P)H and FAD as potential indicators of cancer cell response to chemotherapy with paclitaxel. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1693–1700. [Google Scholar] [CrossRef]
- Shirshin, E.A.; Shirmanova, M.V.; Gayer, A.V.; Lukina, M.M.; Nikonova, E.E.; Yakimov, B.P.; Budylin, G.S.; Dudenkova, V.V.; Ignatova, N.I.; Komarov, D.V.; et al. Label-free sensing of cells with fluorescence lifetime imaging: The quest for metabolic heterogeneity. Proc. Natl. Acad. Sci. USA 2021, 119, e2118241119. [Google Scholar] [CrossRef]
- Walsh, A.J.; Cook, R.S.; Sanders, M.E.; Aurisicchio, L.; Ciliberto, G.; Arteaga, C.L.; Skala, M.C. Quantitative Optical Imaging of Primary Tumor Organoid Metabolism Predicts Drug Response in Breast Cancer. Cancer Res. 2014, 74, 5184–5194. [Google Scholar] [CrossRef] [Green Version]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Sebestyén, A.; Kopper, L.; Dankó, T.; Tímár, J. Hypoxia Signaling in Cancer: From Basics to Clinical Practice. Pathol. Oncol. 2021, 27, 1609802. [Google Scholar] [CrossRef] [PubMed]
- Hildingsson, S.; Gebre-Medhin, M.; Zschaeck, S.; Adrian, G. Hypoxia in relationship to tumor volume using hypoxia PET-imaging in head & neck cancer—A scoping review. Clin. Transl. Radiat. Oncol. 2022, 36, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Kiraga, Ł.; Cheda, Ł.; Taciak, B.; Różańska, K.; Tonecka, K.; Szulc, A.; Kilian, K.; Górka, E.; Rogulski, Z.; Rygiel, T.P.; et al. Changes in hypoxia level of CT26 tumors during various stages of development and comparing different methods of hypoxia determination. PLoS ONE 2018, 13, e0206706. [Google Scholar] [CrossRef]
- Lukina, M.M.; Sirotkina, M.A.; Orlova, A.G.; Dudenkova, V.V.; Komarova, A.D.; Plekhanov, A.A.; Snopova, L.B.; Zagaynova, E.V.; Papkovsky, D.B.; Shcheslavskiy, V.I.; et al. Effects of Irinotecan on Tumor Vasculature and Oxygenation: An in vivo Study on Colorectal Cancer Model. IEEE J. Sel. Top. Quantum Electron. 2021, 27, 1–8. [Google Scholar]
- Rich, L.J.; Miller, A.; Singh, A.K.; Seshadri, M. Photoacoustic Imaging as an Early Biomarker of Radio Therapeutic Efficacy in Head and Neck Cancer. Theranostics 2018, 8, 2064–2078. [Google Scholar] [CrossRef]
- Shibamoto, Y.; Yukawa, Y.; Tsutsui, K.; Takahashi, M.; Abe, M. Variation in the hypoxic fraction among mouse tumors of different types, sizes, and sites. Jpn. J. Cancer Res. 1986, 9, 908–915. [Google Scholar]
- Costantini, I.; Baria, E.; Sorelli, M.; Matuschke, F.; Giardini, F.; Menzel, M.; Mazzamuto, G.; Silvestri, L.; Cicchi, R.; Amunts, K.; et al. Autofluorescence enhancement for label-free imaging of myelinated fibers in mammalian brains. Sci. Rep. 2021, 11, 8038. [Google Scholar] [CrossRef]
- Tregidgo, C.; Levitt, J.A.; Suhling, K. Effect of refractive index on the fluorescence lifetime of green fluorescent protein. J. Biomed. Opt. 2008, 3, 031218. [Google Scholar] [CrossRef]
- Apreleva, S.V.; Wilson, D.F.; Vinogradov, S.A. Tomographic imaging of oxygen by phosphorescence lifetime. Appl. Opt. 2006, 45, 8547–8559. [Google Scholar]
- Fang, X.; Ju, B.; Liu, Z.; Wang, F.; Xi, G.; Sun, Z.; Chen, H.; Sui, C.; Wang, M.; Wu, C. Compact conjugated polymer dots with covalently incorporated metalloporphyrins for hypoxia bioimaging. ChemBioChem 2019, 20, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Lu, X.; Fan, Q.; Zhang, Z.; Song, W.; Li, B.; Huang, L.; Peng, J.; Huang, W. Water-soluble iridium(III)-containing conjugated polyelectrolytes with weakened energy transfer properties for multicolor protein sensing applications. Macromolecules 2011, 44, 8763–8770. [Google Scholar] [CrossRef]
- Liu, S.; Qiao, W.; Cao, G.; Chen, Y.; Ma, Y.; Huang, Y.; Liu, X.; Xu, W.; Zhao, Q.; Huang, W. Smart poly(N-isopropylacrylamide) containing iridium(III) complexes as water-soluble phosphorescent probe for sensing and bioimaging of homocysteine and cysteine. Macromol. Rapid Commun. 2013, 34, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, N.B.; Rys, A.Z.; Nassif, R.; Nayak, M.K.; Metera, K.; Chen, B.; Bazzi, H.S.; Sleiman, H.F. Ring-opening metathesis polymers for biodetection and signal amplification: Synthesis and self-assembly. Macromolecules 2010, 43, 5530–5537. [Google Scholar] [CrossRef]
- Bielawski, C.W.; Grubbs, R.H. Living ring-opening metathesis polymerization. Progr. Polym. Sci. 2007, 32, 1–29. [Google Scholar] [CrossRef]
- Hardy, C.G.; Zhang, J.; Yan, Y.; Ren, L.; Tang, C. Metallopolymers with transition metals in the side-chain by living and controlled polymerization techniques. Progr. Polym. Sci. 2014, 39, 1742–1796. [Google Scholar] [CrossRef]
- Hersey, J.S.; Meller, A.; Grinstaff, M.W. Functionalized nanofiber meshes enhance immunosorbent assays. Anal. Chem. 2015, 87, 11863–11870. [Google Scholar] [CrossRef]
- Sutthasupa, S.; Shiotsuki, M.; Matsuoka, H.; Masuda, T.; Sanda, F. Ring-opening metathesis block copolymerization of amino acid functionalized norbornene monomers. Effects of solvent and pH on micelle formation. Macromolecules 2010, 43, 1815–1822. [Google Scholar] [CrossRef]
- Lepeltier, M.; Dumur, F.; Marrot, J.; Contal, E.; Bertin, D.; Gigmes, D.; Mayer, C.R. Unprecedented combination of regioselective hydrodefluorination and ligand exchange reaction during the syntheses of tris-cyclometalated iridium(III) complexes. Dalton Trans. 2013, 42, 4479–4486. [Google Scholar] [CrossRef]
- Rozhkov, A.V.; Bochkarev, L.N.; Basova, G.V.; Ilichev, V.A.; Abakumov, G.A. Synthesis and luminescent properties of new europium-containing copolymers based on norbornene functional derivatives. Russ. Chem. Bull. 2012, 61, 2243–2251. [Google Scholar] [CrossRef]
- Kim, H.-T.; Seo, J.H.; Ahn, J.H.; Baek, M.-J.; Um, H.-D.; Lee, S.; Roh, D.-H.; Yum, J.-H.; Shin, T.J.; Seo, K.; et al. Customized energy down-shift using iridium complexes for enhanced performance of polymer solar cells. ACS Energy Lett. 2016, 1, 991–999. [Google Scholar] [CrossRef]
- Samanta, S.; Locklin, J. Formation of photochromic spiropyran polymer brushes via surface-initiated, ring-opening metathesis polymerization: Reversible photocontrol of wetting behavior and solvent dependent morphology changes. Langmuir 2008, 24, 9558–9565. [Google Scholar] [CrossRef] [PubMed]
- Scholl, M.; Ding, S.; Lee, C.W.; Grubbs, R.H. Synthesis and activity of a new generation of ruthenium-based olefin metathesis catalysts coordinated with 1,3-dimesityl-4,5-dihydroimidazol-2-ylidene ligands. Org. Lett. 1999, 1, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Love, J.A.; Morgan, J.P.; Trnka, T.M.; Grubbs, R.H. A practical and highly active ruthenium-based catalyst that effects the cross metathesis of acrylonitrile. Angew. Chem. 2002, 41, 4035–4037. [Google Scholar] [CrossRef]
- Demas, J.N.; Crosby, G.A. The Measurement of photoluminescence quantum yields. J. Phys. Chem. 1971, 75, 991–1024. [Google Scholar] [CrossRef]
- Shcheslavskiy, V.I.; Shirmanova, M.V.; Dudenkova, V.V.; Lukyanov, K.A.; Gavrina, A.I.; Shumilova, A.V.; Zagaynova, E.V.; Becker, W. Fluorescence time-resolved microimaging. Opt. Lett. 2018, 43, 3152–3155. [Google Scholar] [CrossRef]
- Sirotkina, M.A.; Gubarkova, E.V.; Plekhanov, A.A.; Sovetsky, A.A.; Elagin, V.V.; Matveyev, A.L.; Matveev, L.A.; Kuznetsov, S.S.; Zagaynova, E.V.; Gladkova, N.D.; et al. In vivo assessment of functional and morphological alterations in tumors under treatment using OCT-angiography combined with OCT-elastography. Biomed. Opt. Express 2020, 11, 1365–1382. [Google Scholar] [CrossRef]
- Tomayko, M.M.; Reynolds, C.P. Determination of subcutaneous tumor size in athymic (nude) mice. Cancer Chemother. Pharmacol. 1989, 24, 148–154. [Google Scholar] [CrossRef]
- Conway, J.R.W.; Warren, S.C.; Herrmann, D.; Murphy, K.J.; Aurélie, S.; Vennin, C.; Shearer, R.F.; Killen, M.J.; Magenau, A.; Mélénec, P.; et al. Intravital Imaging to Monitor Therapeutic Response in Moving Hypoxic Regions Resistant to PI3K Pathway Targeting in Pancreatic Cancer. Cell Rep. 2018, 23, 3312–3326. [Google Scholar] [CrossRef] [PubMed]
- Lukina, M.M.; Dudenkova, V.V.; Shimolina, L.E.; Snopova, L.B.; Zagaynova, E.V.; Shirmanova, M.V. In vivo metabolic and SHG imaging for monitoring of tumor response to chemotherapy. Cytom. Part A 2019, 95, 47–55. [Google Scholar] [CrossRef]
- Shirmanova, M.V.; Druzhkova, I.N.; Lukina, M.M.; Dudenkova, V.V.; Ignatova, N.I.; Snopova, L.B.; Shcheslavskiy, V.I.; Belousov, V.V.; Zagaynova, E.V. Chemotherapy with cisplatin: Insights into intracellular pH and metabolic landscape of cancer cells in vitro and in vivo. Sci. Rep. 2017, 7, 8911. [Google Scholar] [CrossRef] [PubMed]
- Moiseev, A.; Ksenofontov, S.; Sirotkina, M.; Kiseleva, E.; Gorozhantseva, M.; Shakhova, N.; Matveev, L.; Zaitsev, V.; Matveyev, A.; Zagaynova, E.; et al. Optical coherence tomography-based angiography device with real-time angiography B-scans visualization and hand-held probe for everyday clinical use. J. Biophotonics 2018, 11, e201700292. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | λmaxabs/nm (log ε) in CH2Cl2 | λmaxem/nm (in CH2Cl2) | Quantum Yield, % (in CH2Cl2) | Chromaticity Coordinates in the CIE Diagram (x; y) | |
|---|---|---|---|---|---|
| (a) | (b) | ||||
| PIr1 | 291 (5.12), 341 (4.97), 381 (4.76), 448 (4.54), 484 (4.39) | 588, 629 sh | 12.76 | 2.33 | 0.57; 0.41 |
| PIr2 | 291 (5.32), 343 (5.14), 387 (4.93), 439 (4.76), 480 (4.62) | 588, 629 sh | 14.64 | 2.95 | 0.58; 0.41 |
| PIr3 | 297 (5.03), 348 (4.86), 404 (4.08), 466 (4.63), 510 (4.53) | 605 | 16.45 | 4.03 | 0.64; 0.36 |
| Compound | λmaxabs/nm (log ε) in H2O | λmaxem/nm (in H2O) | Quantum Yield, % (in H2O) | Chromaticity Coordinates in the CIE Diagram (x; y) | |
|---|---|---|---|---|---|
| (a) | (b) | ||||
| PIr1 | 293 (4.74), 344 (4.57), 390 (4.31), 446 (4.11), 485 (3.94) | 588, 629 sh | 7.54 | 4.74 | 0.57; 0.40 |
| PIr2 | 291 (4.75), 340 (4.61), 390 (4.34), 445 (4.16), 481 (3.99) | 588, 629 sh | 8.42 | 6.18 | 0.57; 0.39 |
| PIr3 | 294 (4.62), 349 (4.39), 399 (4.12), 461 (3.94), 505 (3.74) | 605 | 8.79 | 5.77 | 0.64; 0.36 |
| PIr1 | PIr2 | PIr3 | |
|---|---|---|---|
| τ0, μs | 3.43 ± 0.07 | 3.15 ± 0.08 | 2.04 ± 0.03 |
| τ, μs | 2.13 ± 0.05 | 2.13 ± 0.05 | 1.41 ± 0.08 |
| τ in PBS, μs | 2.01 ± 0.09 | 1.98 ± 0.05 | 1.31 ± 0.05 |
| τ in 5% BSA, μs | 2.02 ± 0.07 | 1.95 ± 0.06 | 1.41 ± 0.05 |
| τ in DMEM with 10% FBS, μs | 1.91 ± 0.07 | 1.97 ± 0.07 | 1.33 ± 0.09 |
| Dynamic range r = τ0/τ | 1.61 ± 0.07 | 1.47 ± 0.07 | 1.44 ± 0.06 |
| Photon counts (for τ) | 5890 | 5600 | 13,823 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parshina, Y.P.; Komarova, A.D.; Bochkarev, L.N.; Kovylina, T.A.; Plekhanov, A.A.; Klapshina, L.G.; Konev, A.N.; Mozherov, A.M.; Shchechkin, I.D.; Sirotkina, M.A.; et al. Simultaneous Probing of Metabolism and Oxygenation of Tumors In Vivo Using FLIM of NAD(P)H and PLIM of a New Polymeric Ir(III) Oxygen Sensor. Int. J. Mol. Sci. 2022, 23, 10263. https://doi.org/10.3390/ijms231810263
Parshina YP, Komarova AD, Bochkarev LN, Kovylina TA, Plekhanov AA, Klapshina LG, Konev AN, Mozherov AM, Shchechkin ID, Sirotkina MA, et al. Simultaneous Probing of Metabolism and Oxygenation of Tumors In Vivo Using FLIM of NAD(P)H and PLIM of a New Polymeric Ir(III) Oxygen Sensor. International Journal of Molecular Sciences. 2022; 23(18):10263. https://doi.org/10.3390/ijms231810263
Chicago/Turabian StyleParshina, Yulia P., Anastasia D. Komarova, Leonid N. Bochkarev, Tatyana A. Kovylina, Anton A. Plekhanov, Larisa G. Klapshina, Aleksey N. Konev, Artem M. Mozherov, Ilya D. Shchechkin, Marina A. Sirotkina, and et al. 2022. "Simultaneous Probing of Metabolism and Oxygenation of Tumors In Vivo Using FLIM of NAD(P)H and PLIM of a New Polymeric Ir(III) Oxygen Sensor" International Journal of Molecular Sciences 23, no. 18: 10263. https://doi.org/10.3390/ijms231810263
APA StyleParshina, Y. P., Komarova, A. D., Bochkarev, L. N., Kovylina, T. A., Plekhanov, A. A., Klapshina, L. G., Konev, A. N., Mozherov, A. M., Shchechkin, I. D., Sirotkina, M. A., Shcheslavskiy, V. I., & Shirmanova, M. V. (2022). Simultaneous Probing of Metabolism and Oxygenation of Tumors In Vivo Using FLIM of NAD(P)H and PLIM of a New Polymeric Ir(III) Oxygen Sensor. International Journal of Molecular Sciences, 23(18), 10263. https://doi.org/10.3390/ijms231810263