Proteomic Analysis of Renal Biomarkers of Kidney Allograft Fibrosis—A Study in Renal Transplant Patients

 , ,
, ,
Abstract
1. Introduction
2. Results
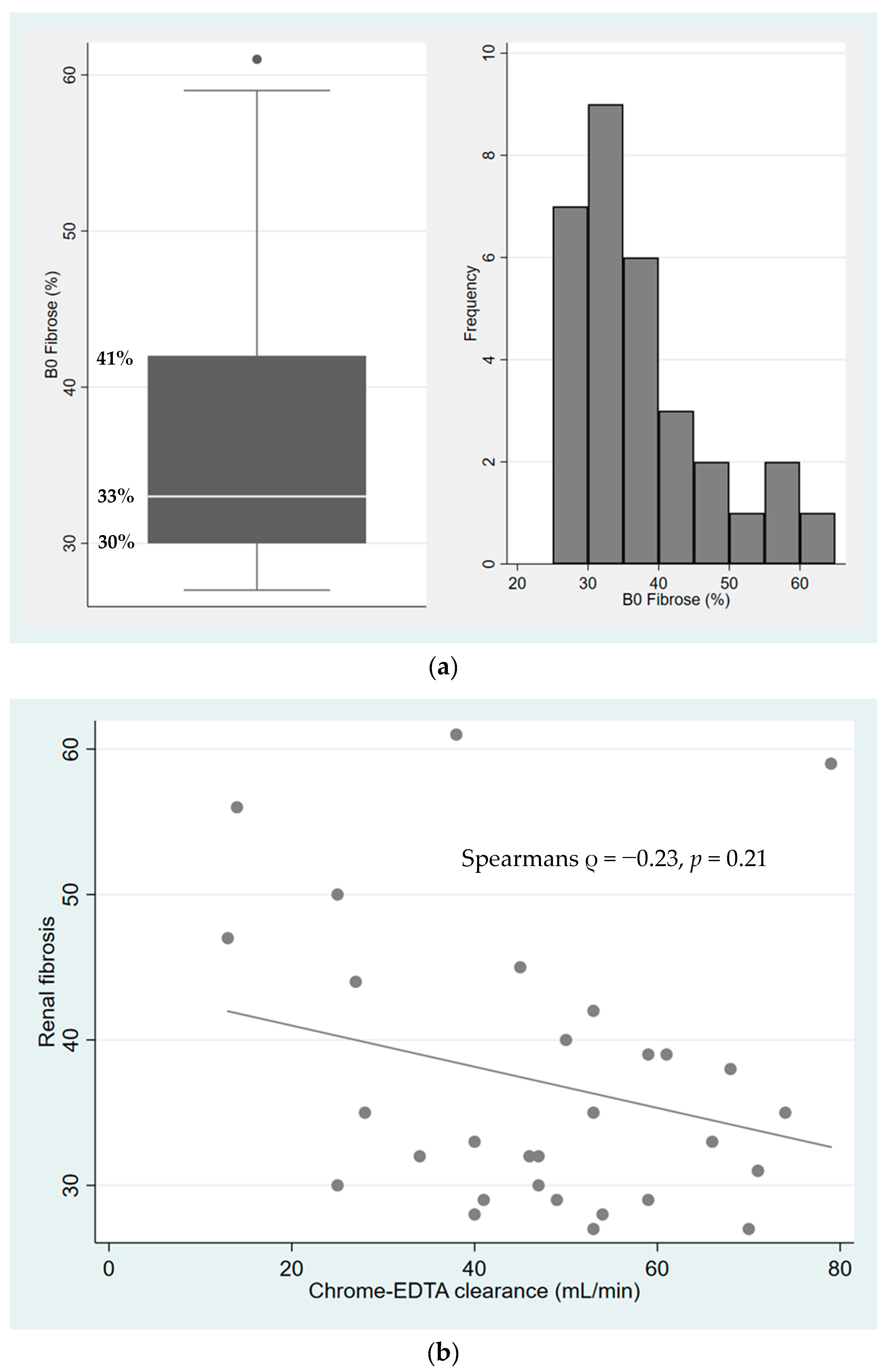
2.1. Characterization of Fibrosis in Kidney Allograft Biopsies
2.2. Kidney Allograft Proteomics
2.3. Proteins Correlated with the Degree of Fibrosis
3. Discussion
4. Methods
4.1. Participants
4.2. Proteomics
4.2.1. Sample Preparation
4.2.2. HILIC Fractionation
4.2.3. NanoLC-MS/MS
4.3. Data Analysis
4.4. Statistical Analysis
4.5. Ethics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wolfe, R.A.; Ashby, V.B.; Milford, E.L.; Ojo, A.; Ettenger, R.E.; Agodoa, L.Y.; Held, P.J.; Port, F.K. Comparison of Mortality in All Patients on Dialysis, Patients on Dialysis Awaiting Transplantation, and Recipients of a First Cadaveric Transplant. N. Engl. J. Med. 1999, 341, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.H.; Garratt, A.M.; Cody, D.J.; Catto, G.R.D.; Kumar, A.; Edward, N.; MacLeod, A.M. Patients’ perception of health on renal replacement therapy: Evaluation using a new instrument. Nephrol. Dial. Transplant. 1995, 10, 684–689. [Google Scholar] [PubMed]
- Hariharan, S.; Bresnahan, B.A.; Taranto, S.E.; McIntosh, M.J.; Stablein, N.; Johnson, C.P. Improved Graft Survival after Renal Transplantation in the United States, 1988 to 1996. N. Engl. J. Med. 2000, 342, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Solez, K.; Colvin, R.B.; Racusen, L.C.; Sis, B.; Halloran, P.F.; Birk, P.E.; Campbell, P.M.; Cascalho, M.; Collins, A.B.; Demetris, A.J.; et al. Banff ’05 Meeting Report: Differential Diagnosis of Chronic Allograft Injury and Elimination of Chronic Allograft Nephropathy (‘CAN’). Am. J. Transplant. 2007, 7, 518–526. [Google Scholar] [CrossRef]
- Serón, D.; Moreso, F.; Ser, D. Protocol biopsies in renal transplantation: Prognostic value of structural monitoring. Kidney Int. 2007, 72, 690–697. [Google Scholar] [CrossRef]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef]
- Fletcher, J.T.; Nankivell, B.J.; Alexander, S.I. Chronic allograft nephropathy. Pediatr. Nephrol. 2009, 24, 1465–1471. [Google Scholar] [CrossRef]
- Schwarz, A.; Gwinner, W.; Hiss, M.; Radermacher, J.; Mengel, M.; Haller, H. Safety and Adequacy of Renal Transplant Protocol Biopsies. Am. J. Transplant. 2005, 5, 1992–1996. [Google Scholar] [CrossRef]
- Farris, A.B.; Colvin, R.B. Renal interstitial fibrosis: Mechanisms and evaluation. Curr. Opin. Nephrol. Hypertens. 2012, 21, 289–300. [Google Scholar] [CrossRef]
- Rhee, E.P. How Omics Data Can Be Used in Nephrology. Am. J. Kidney Dis. 2018, 72, 129–135. [Google Scholar] [CrossRef]
- Knepper, M.A. Common sense approaches to urinary biomarker study design. J. Am. Soc. Nephrol. 2009, 20, 1175–1178. [Google Scholar] [CrossRef] [PubMed]
- Nakorchevsky, A.; Hewel, J.A.; Kurian, S.; Mondala, T.S.; Campbell, D.; Head, S.R.; Marsh, C.; Yates, J.R.; Salomon, D.R. Molecular mechanisms of chronic kidney transplant rejection via large-scale proteogenomic analysis of tissue biopsies. J. Am. Soc. Nephrol. 2010, 21, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Bagoly, Z.; Katona, É.; Muszbek, L. Factor XIII and inflammatory cells. Thromb. Res. 2012, 129, S77–S81. [Google Scholar] [CrossRef] [PubMed]
- Bereczky, Z.; Katona, Ë.; Muszbek, L. Fibrin Stabilization (Factor XIII), Fibrin Structure and Thrombosis. Pathophysiol. Haemost. Thromb. 2003, 33, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Kohler, H.P.; Schroeder, V. Factor XIII: Structure and Function. Semin. Thromb. Hemost. 2016, 42, 422–428. [Google Scholar] [CrossRef]
- Dardik, R.; Krapp, T.; Rosenthal, E.; Loscalzo, J.; Inbal, A. Effect of FXIII on Monocyte and Fibroblast Function. Cell. Physiol. Biochem. 2007, 19, 113–120. [Google Scholar] [CrossRef]
- Welch, M.D.; DePace, A.H.; Verma, S.; Iwamatsu, A.; Mitchison, T.J. The human Arp2/3 complex is composed of evolutionarily conserved subunits and is localized to cellular regions of dynamic actin filament assembly. J. Cell Biol. 1997, 138, 375–384. [Google Scholar] [CrossRef]
- Magdalena, J.; Millard, T.H.; Etienne-Manneville, S.; Launay, S.; Warwick, H.K.; Machesky, L.M. Involvement of the Arp2/3 complex and Scar2 in Golgi polarity in scratch wound models. Mol. Biol. Cell. 2003, 14, 670–684. [Google Scholar] [CrossRef][Green Version]
- Barrientos, A.; De Barros, M.H.; Valnot, I.; Rötig, A.; Rustin, P.; Tzagoloff, A. Cytochrome oxidase in health and disease. Gene 2002, 286, 53–63. [Google Scholar] [CrossRef]
- Pacheu-Grau, D.; Bareth, B.; Dudek, J.; Juris, L.; Vögtle, F.-N.; Wissel, M.; Leary, S.C.; Dennerlein, S.; Rehling, P.; Deckers, M. Cooperation between COA6 and SCO2 in COX2 Maturation during Cytochrome c Oxidase Assembly Links Two Mitochondrial Cardiomyopathies. Cell Metab. 2015, 21, 823–833. [Google Scholar] [CrossRef]
- Puigmulé, M.; Lopez-Hellin, J.; Suñè, G.; Tornavaca, O.; Camano, S.; Tejedor, A.; Meseguer, A. Differential proteomic analysis of cyclosporine A-induced toxicity in renal proximal tubule cells. Nephrol. Dial. Transplant. 2009, 24, 2672–2686. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.L.; Cao, D.; Zhang, D.; Handschumacher, R.E.; Pizzorno, G. Uridine Phosphorylase Association with Vimentin. Intracellular distribution and localization. J. Boil. Chem. 2001, 276, 13302–13307. [Google Scholar] [CrossRef] [PubMed]
- Pizzorno, G.; Cao, D.; Leffert, J.J.; Russell, R.L.; Zhang, D.; E Handschumacher, R. Homeostatic control of uridine and the role of uridine phosphorylase: A biological and clinical update. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2002, 1587, 133–144. [Google Scholar] [CrossRef]
- Reuter, S.; Reiermann, S.; Wörner, R.; Schröter, R.; Edemir, B.; Buck, F.; Henning, S.; Peter-Katalinić, J.; Vollenbröker, B.; Amann, K.; et al. IF/TA-related metabolic changes—Proteome analysis of rat renal allografts. Nephrol. Dial. Transplant. 2010, 25, 2492–2501. [Google Scholar] [CrossRef]
- Kunter, U.; Floege, J.; Von Jurgensonn, A.S.; Stojanovic, T.; Merkel, S.; Grone, H.J.; Ferran., C. Expression of A20 in the vessel wall of rat-kidney allografts correlates with protection from transplant arteriosclerosis. Transplant. 2003, 75, 3–9. [Google Scholar] [CrossRef]
- Späth, M.R.; Bartram, M.P.; Palacio-Escat, N.; Hoyer, K.J.R.; Debes, C.; Demir, F.; Schroeter, C.B.; Mandel, A.M.; Grundmann, F.; Ciarimboli, G.; et al. The proteome microenvironment determines the protective effect of preconditioning in cisplatin-induced acute kidney injury. Kidney Int. 2019, 95, 333–349. [Google Scholar] [CrossRef]
- Racusen, L.C.; Solez, K.; Colvin, R.B.; Bonsib, S.M.; Castro, M.C.; Cavallo, T.; Croker, B.P.; Demetris, A.J.; Drachenberg, C.B.; Fogo, A.B.; et al. The Banff 97 working classification of renal allograft pathology. Kidney Int. 1999, 55, 713–723. [Google Scholar] [CrossRef]
- Ellingsen, A.R.; Nyengaard, J.R.; Østerby, R.; Jørgensen, K.A.; Petersen, S.E.; Marcussen, N. Measurements of cortical interstitium in biopsies from human kidney grafts: How representative and how reproducible? Nephrol. Dial. Transplant. 2002, 17, 788–792. [Google Scholar] [CrossRef]
- Nicholson, M.L.; McCulloch, T.A.; Harper, S.J.; Wheatley, T.J.; Edwards, C.M.; Feehally, J.; Furness, P.N. Early measurement of interstitial fibrosis predicts long-term renal function and graft survival in renal transplantation. BJS 1996, 83, 1082–1085. [Google Scholar] [CrossRef]
- Williamson, J.; Edwards, A.V.G.; Verano-Braga, T.; Schwämmle, V.; Kjeldsen, F.; Jensen, O.N.; Larsen, M.R. High-performance hybrid Orbitrap mass spectrometers for quantitative proteome analysis: Observations and implications. Proteomics 2016, 16, 907–914. [Google Scholar] [CrossRef]
- Mortensen, L.A.; Thiesson, H.C.; Tougaard, B.; Egfjord, M.; Fischer, A.S.L.; Bistrup, C. The effect of spironolactone on calcineurin inhibitor induced nephrotoxicity: A multicenter randomized, double-blind, clinical trial (the SPIREN trial). BMC Nephrol. 2018, 19, 105. [Google Scholar] [CrossRef]
- Solez, K.; Axelsen, R.A.; Benediktsson, H.; Burdick, J.F.; Cohen, A.H.; Colvin, R.B.; Croker, B.P.; Droz, D.; Dunnill, M.S.; Halloran, P.F.; et al. International standardization of criteria for the histologic diagnosis of renal allograft rejection: The Banff working classification of kidney transplant pathology. Kidney Int. 1993, 44, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Meinshausen, N.; Buhlmann, P. Stability selection. J. R. Stat. Soc. B 2010, 72, 417–473. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Item | Value |
|---|---|
| Male sex | 22 (71%) |
| Body Mass Index (kg/m2) | 26 (24–28) |
| Age (years) | 60 (50–66) |
| Smoking (never/previous/current) | 16/9/6 (52%/29%/19%) |
| Age of current transplant (years) | 3.0 (1.6–9.5) |
| Donor type | |
| Living related | 12 |
| Deceased | 19 |
| Previous dialysis | 24 (77%) |
| Previous rejection | 3 (10%) |
| Borderline | 1 (3%) |
| Grade 1A | 2 (6%) |
| Cause of renal failure | |
| Hypertension | 6 (19%) |
| Diabetes | 2 (6%) |
| Polycystic kidney disease | 9 (29%) |
| IgA nephritis | 2 (6%) |
| Glomerulonepritis | 3 (10%) |
| Congenital malformation | 3 (10%) |
| Reflux nefropathy | 1 (3%) |
| Unknown | 5 (16%) |
| Comorbidity | |
| Hypertension | 29 (94%) |
| Diabetes | 2 (6%) |
| Apoplexia | 1 (3%) |
| Myocardial infarction | 3 (10%) |
| Heart failure | 0 (0%) |
| Current medication | |
| ACE-inhibitors | 14 (45%) |
| Angiotensin II receptor blockers | 2 (6%) |
| Calcium channel antagonists | 18 (58%) |
| β-adrenergic antagonists | 17 (55%) |
| α-adrenergic antagonists | 5 (16%) |
| Loop diuretics | 4 (13%) |
| Thiazide diuretics | 2 (6%) |
| Cholesterol lowering drugs | 4 (13%) |
| Calcineurin inhibition: | |
| -Tacrolimus | 25 (81%) |
| -Cyclosporine A | 6 (19%) |
| Antimetabolite: | |
| -Mycophenolate mofetil | 29 (94%) |
| -Azathioprine | 2 (6%) |
| Prednisolone | 4 (13%) |
| Systolic blood pressure (mmHg) | 131 (118–141) |
| Diastolic blood pressure (mmHg) | 79 (75–83) |
| Chrome-EDTA clearance (mL/min) | 49 (39–60) |
| Fibrosis (%) | 33 (30–41) |
| Protein | Accesion Number A | Estimate B | SE C | p-Value | Adjusted p-Value D | Missing values E |
|---|---|---|---|---|---|---|
| Uridine phosphorylase 1 | Q16831 | 19.4 | 2.5 | 3.06 × 10-7 | 8.21 × 10-4 | 8 |
| Cytochrome c oxidase assembly factor 6 homolog | Q5JTJ3.2 | −44.9 | 6.9 | 6,98 × 10-7 | 1.88 × 10-3 | 0 |
| Coagulation factor XIII A chain | P00488 | 18.7 | 3.4 | 9.09 × 10-6 | 2.44 × 10-2 | 0 |
| Actin-related protein 2/3 complex subunit 2 | O15144 | 34.3 | 6.5 | 1.78 × 10-5 | 4.77 × 10-2 | 0 |
| WD repeat-containing protein 82 | Q6UXN9 | 34.8 | 6.0 | 3.38 × 10-5 | 9.08 × 10-2 | 11 |
| Actin-related protein 2/3 complex subunit 1B | O15143 | 21.6 | 4.5 | 5.17 × 10-5 | 1.39 × 10-1 | 0 |
| Heterogeneous nuclear ribonucleoprotein U | Q00839 | 48.3 | 10.0 | 5.38 × 10-5 | 1.44 × 10-1 | 0 |
| Mitochondrial aconitate hydratase | A2A274 | −47.2 | 10.1 | 8.20 × 10-5 | 2.20 × 10-1 | 0 |
| Hematopoietic lineage cell-specific protein | P14317 | 11.3 | 2.5 | 9.49 × 10-5 | 2.54 × 10-1 | 0 |
| Lamina-associated polypeptide 2, isoforms beta/gamma | P42167 | 25.7 | 5.6 | 1.06 × 10-4 | 2.84 × 10-1 | 0 |
| NADH dehydrogenase 1 alpha subcomplex subunit 2 | O43678 | −38.4 | 8.5 | 1.13 × 10-4 | 3.01 × 10-1 | 0 |
| 60S ribosomal protein L7a | P62424 | 56.2 | 12.4 | 1.17 × 10-4 | 3.14 × 10-1 | 0 |
| Cathepsin G | P08311 | 9.9 | 2.2 | 1.18 × 10-4 | 3.16 × 10-1 | 0 |
| Perilipin-3 | O60664 | 33.6 | 7.3 | 1.21 × 10-4 | 3.24 × 10-1 | 3 |
| Dimethylglycine dehydrogenase, mitochondrial | Q9UI17 | −26.6 | 5.9 | 1.22 × 10-4 | 3.26 × 10-1 | 0 |
| 60S ribosomal protein L5 | P46777 | 36.3 | 8.1 | 1.28 × 10-4 | 3.43 × 10-1 | 0 |
| Proteasome subunit beta type-8 | P28062 | 26.8 | 6.0 | 1.35 × 10-4 | 3.61 × 10-1 | 0 |
| HLA class I histocompatibility antigen, B-42 alpha chain | P30480 | 17.6 | 3.9 | 1.37 × 10-4 | 3.65 × 10-1 | 0 |
| Xaa-Pro aminopeptidase 2 | O43895 | −27.5 | 6.2 | 1.41 × 10-4 | 3.78 × 10-1 | 0 |
| Coronin-1A | P31146 | 5.9 | 1.3 | 1.80 × 10-4 | 3.94 × 10-1 | 0 |
| Tubulin beta chain | P07437 | 36.3 | 8.2 | 1.54 × 10-4 | 4.10 × 10-1 | 0 |
| Splicing factor U2AF 65 kDa subunit | P26368 | 25.7 | 5.8 | 1.54 × 10-4 | 4.11 × 10-1 | 0 |
| Short/branched chain specific acyl-CoA dehydrogenase, mitochondrial | P45954 | −26.9 | 6.1 | 1.58 × 10-4 | 4.20 × 10-1 | 0 |
| X-ray repair cross-complementing protein 6 | P12956 | 40.2 | 9.1 | 1.63 × 10-4 | 4.33 × 10-1 | 0 |
| Delta(3,5)-Delta(2,4)-dienoyl-CoA isomerase, mitochondrial | Q13011 | −37.2 | 8.5 | 1.72 × 10-4 | 4.58 × 10-1 | 0 |
| Sodium-dependent neutral amino acid transporter B(0)AT1 | Q695T7 | −21.4 | 4.9 | 1.79 × 10-4 | 4.77 × 10-1 | 0 |
| Protein | Accession Number | Correlation | Biological Process | Selection Probability |
|---|---|---|---|---|
| Cytochrome c oxidase assembly factor 6 homolog | Q5JTJ3.2 | Negative | Cell respiration | 0.567 |
| Apoptosis inhibitor 5 | Q9BZZ5.2 | Positive | Inhibition of (fibroblast) apoptosis | 0.234 |
| Coagulation factor XIII A chain | P00488 | Positive | Coagulation/cross-linking ECM proteins/inhibition of fibroblast apoptosis | 0.217 |
| Microtubule-associated protein 1B | P46821 | Positive | Neuronal cell structure maintainance | 0.205 |
| Splicing factor U2AF 65 kDa subunit | P26368 | Positive | mRNA processing | 0.199 |
| Symplekin | Q92797 | Positive | Cell adhesion/mRNA processing | 0.156 |
| Threonine synthase-like 1 | Q8IYQ7 | Negative | Unknown | 0.131 |
| AP-2 complex subunit sigma | P53680 | Positive | Protein transport | 0.118 |
| Actin-related protein 2/3 complex subunit 2 | O15144 | Positive | Actin cytoskeleton organization | 0.107 |
| GO Term | Description | p-Value | FDR q-Value | Enrichment (N, B, n, b) |
|---|---|---|---|---|
| Categories positively associated with fibrosis | ||||
| GO:0010468 | regulation of gene expression | 2.09 × 10-7 | 1.97 × 10-3 | 1.89 (2240,546,113,52) |
| GO:0051704 | multi-organism process | 2.37 × 10-7 | 1.11 × 10-3 | 2.22 (2240, 348, 113, 39) |
| GO:0050789 | regulation of biological process | 5.94 × 10-7 | 1.86 × 10-3 | 1.34 (2240, 1371, 113, 93) |
| GO:0010629 | negative regulation of gene expression | 6.29 × 10-7 | 1.48 × 10-3 | 2.47 (2240, 249, 113, 31) |
| GO:0044419 | interspecies interaction between organisms | 8.74 × 10-7 | 1.64 × 10-3 | 2.38 (2240, 266, 113, 32) |
| GO:0002376 | immune system process | 1.27 × 10-6 | 1.99 × 10-3 | 1.96 (2240, 446, 113, 44) |
| GO:0002682 | regulation of immune system process | 1.41 × 10-6 | 1.90 × 10-3 | 2.38 (2240, 258, 113, 31) |
| GO:0045321 | leukocyte activation | 1.46 × 10-6 | 1.72 × 10-3 | 2.43 (2240, 245, 113, 30) |
| GO:0006952 | defense response | 1.83 × 10-6 | 1.91 × 10-3 | 2.87 (2240, 159, 113, 23) |
| GO:0002252 | immune effector process | 3.34 × 10-6 | 3.14 × 10-3 | 2.21 (2240, 296, 113, 33) |
| GO:0048518 | positive regulation of biological process | 3.65 × 10-6 | 3.12 × 10-3 | 1.55 (2240, 844, 113, 66) |
| GO:0006955 | immune response | 5.25 × 10-6 | 4.11 × 10-3 | 2.98 (2240, 133, 113, 20) |
| GO:0050794 | regulation of cellular process | 5.83 × 10-6 | 4.21 × 10-3 | 1.36 (2240, 1239, 113, 85) |
| GO:0007166 | cell surface receptor signaling pathway | 8.47 × 10-6 | 5.68 × 10-3 | 2.16 (2240, 294, 113, 32) |
| GO:0001775 | cell activation | 9.28 × 10-6 | 5.81 × 10-3 | 2.23 (2240, 267, 113, 30) |
| GO:0002684 | positive regulation of immune system process | 9.39 × 10-6 | 5.52 × 10-3 | 2.54 (2240, 187, 113, 24) |
| Categories negatively associated with fibrosis | ||||
| GO:0055114 | oxidation-reduction process | 1.94 × 10-15 | 1.79 × 10-11 | 3.75 (2198, 318, 70, 38) |
| GO:0019752 | carboxylic acid metabolic process | 1.16 × 10-11 | 5.37 × 10-8 | 3.16 (2198, 348, 70, 35) |
| GO:0043436 | oxoacid metabolic process | 4.53 × 10-11 | 1.39 × 10-7 | 3.02 (2198, 364, 70, 38) |
| GO:0044281 | small molecule metabolic process | 6.03 × 10-11 | 1.39 × 10-7 | 2.40 (2198, 576, 70, 44) |
| GO:0006082 | organic acid metabolic process | 8.68 × 10-11 | 1.60 × 10-7 | 2.95 (2198, 372, 70, 35) |
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| Age > 18 years | Former intolerance of spironolactone |
| Tacrolimus/cyclosporine treatment | Potassium binder or digoxin treatment |
| Proteinuria < 3 g/day | Pregnancy or planned pregnancy |
| Creatinine clearance ≥ 30 mL/min | Clinically relevant organic, systemic or psychological disorder |
| Plasma potassium < 5.5 mmol/L | Expectation of non-compliance |
| Negative pregnancy test at inclusion for women of childbearing potential and adequate contraception throughout the trial |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mortensen, L.A.; Svane, A.M.; Burton, M.; Bistrup, C.; Thiesson, H.C.; Marcussen, N.; Beck, H.C. Proteomic Analysis of Renal Biomarkers of Kidney Allograft Fibrosis—A Study in Renal Transplant Patients. Int. J. Mol. Sci. 2020, 21, 2371. https://doi.org/10.3390/ijms21072371
Mortensen LA, Svane AM, Burton M, Bistrup C, Thiesson HC, Marcussen N, Beck HC. Proteomic Analysis of Renal Biomarkers of Kidney Allograft Fibrosis—A Study in Renal Transplant Patients. International Journal of Molecular Sciences. 2020; 21(7):2371. https://doi.org/10.3390/ijms21072371
Chicago/Turabian StyleMortensen, Line Aas, Anne Marie Svane, Mark Burton, Claus Bistrup, Helle Charlotte Thiesson, Niels Marcussen, and Hans Christian Beck. 2020. "Proteomic Analysis of Renal Biomarkers of Kidney Allograft Fibrosis—A Study in Renal Transplant Patients" International Journal of Molecular Sciences 21, no. 7: 2371. https://doi.org/10.3390/ijms21072371
APA StyleMortensen, L. A., Svane, A. M., Burton, M., Bistrup, C., Thiesson, H. C., Marcussen, N., & Beck, H. C. (2020). Proteomic Analysis of Renal Biomarkers of Kidney Allograft Fibrosis—A Study in Renal Transplant Patients. International Journal of Molecular Sciences, 21(7), 2371. https://doi.org/10.3390/ijms21072371