Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
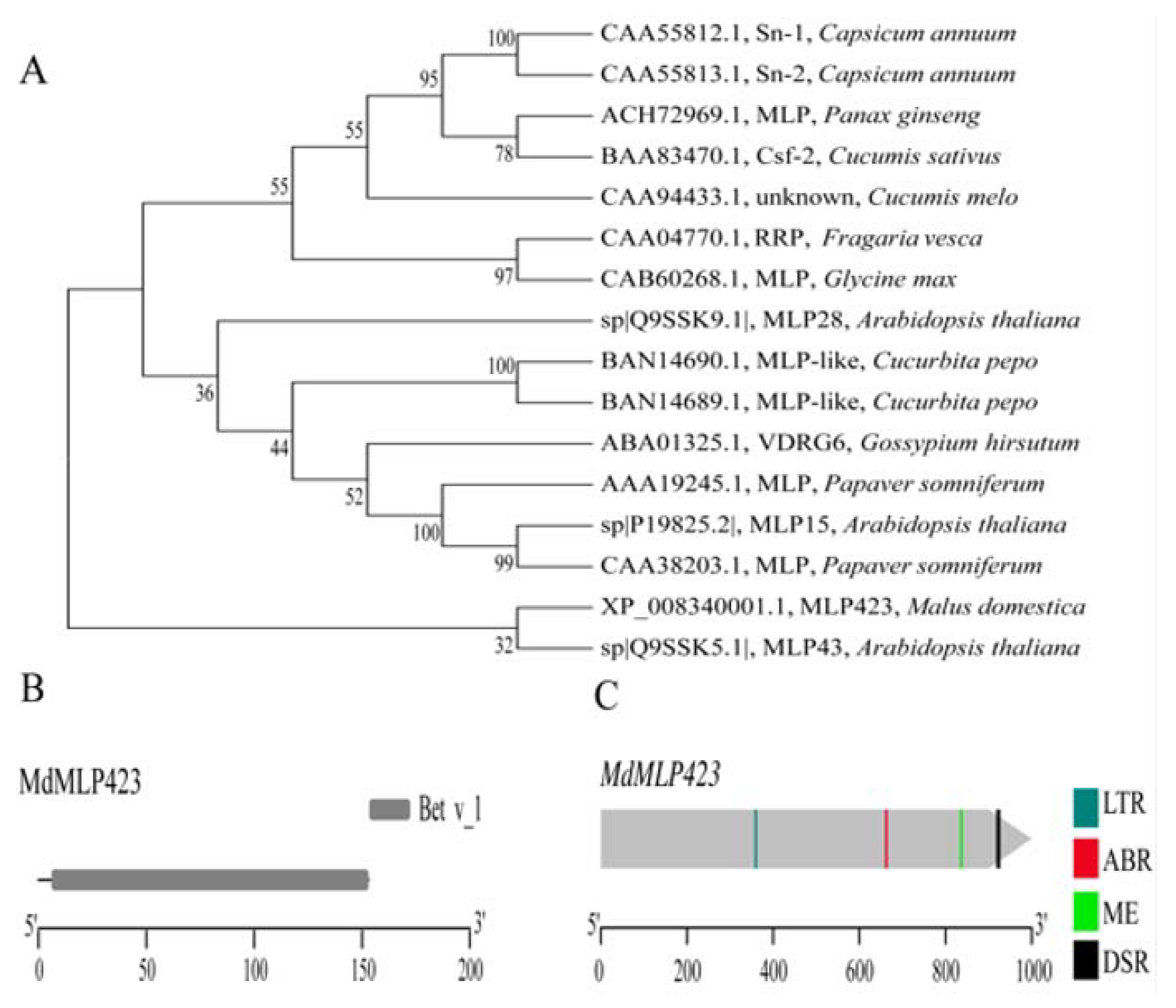
2.1. Isolation and Sequence Analysis of the MdMLP423 Gene
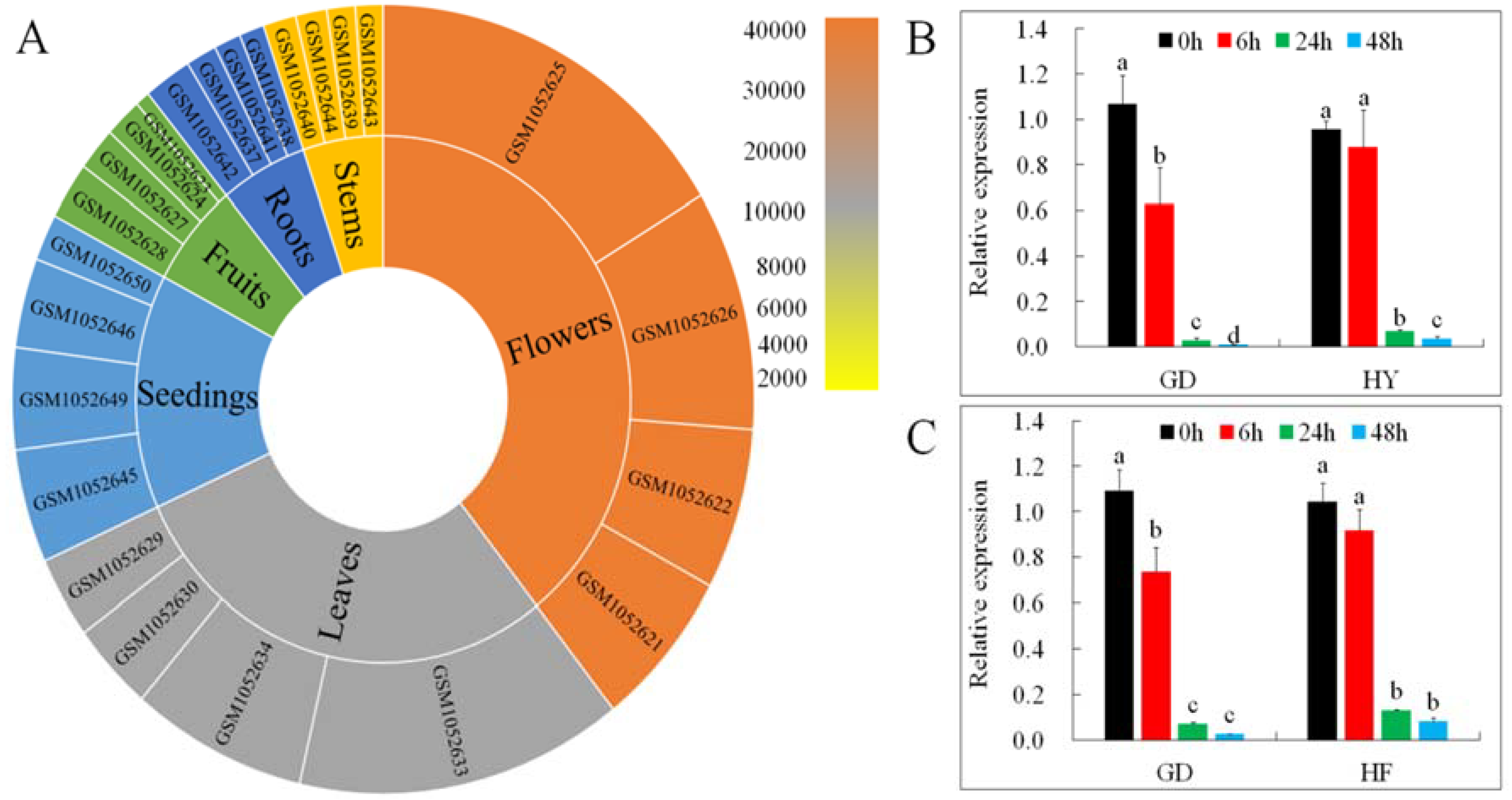
2.2. Expression of MdMLP423
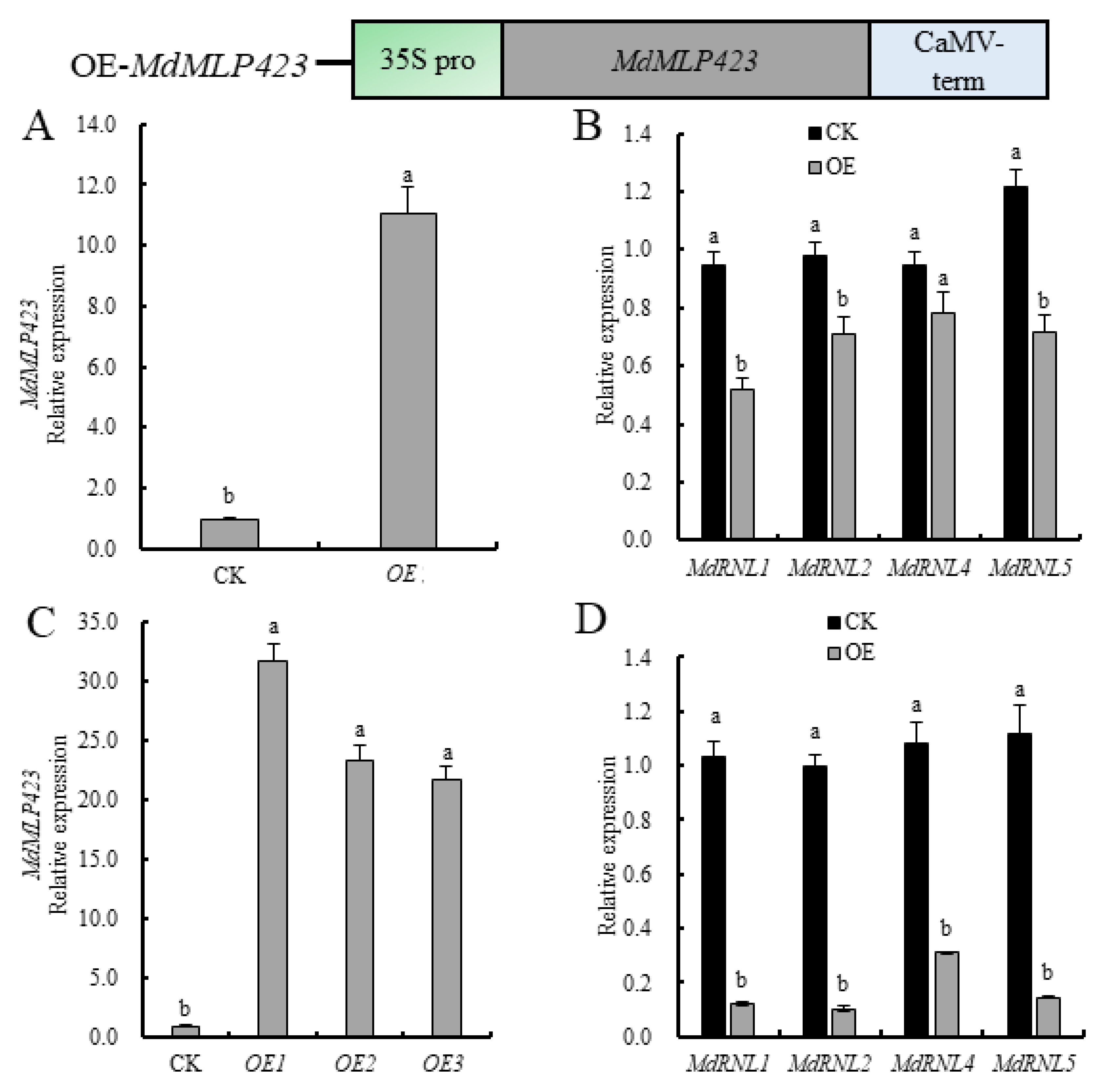
2.3. Overexpression of MdMLP423 in Apples
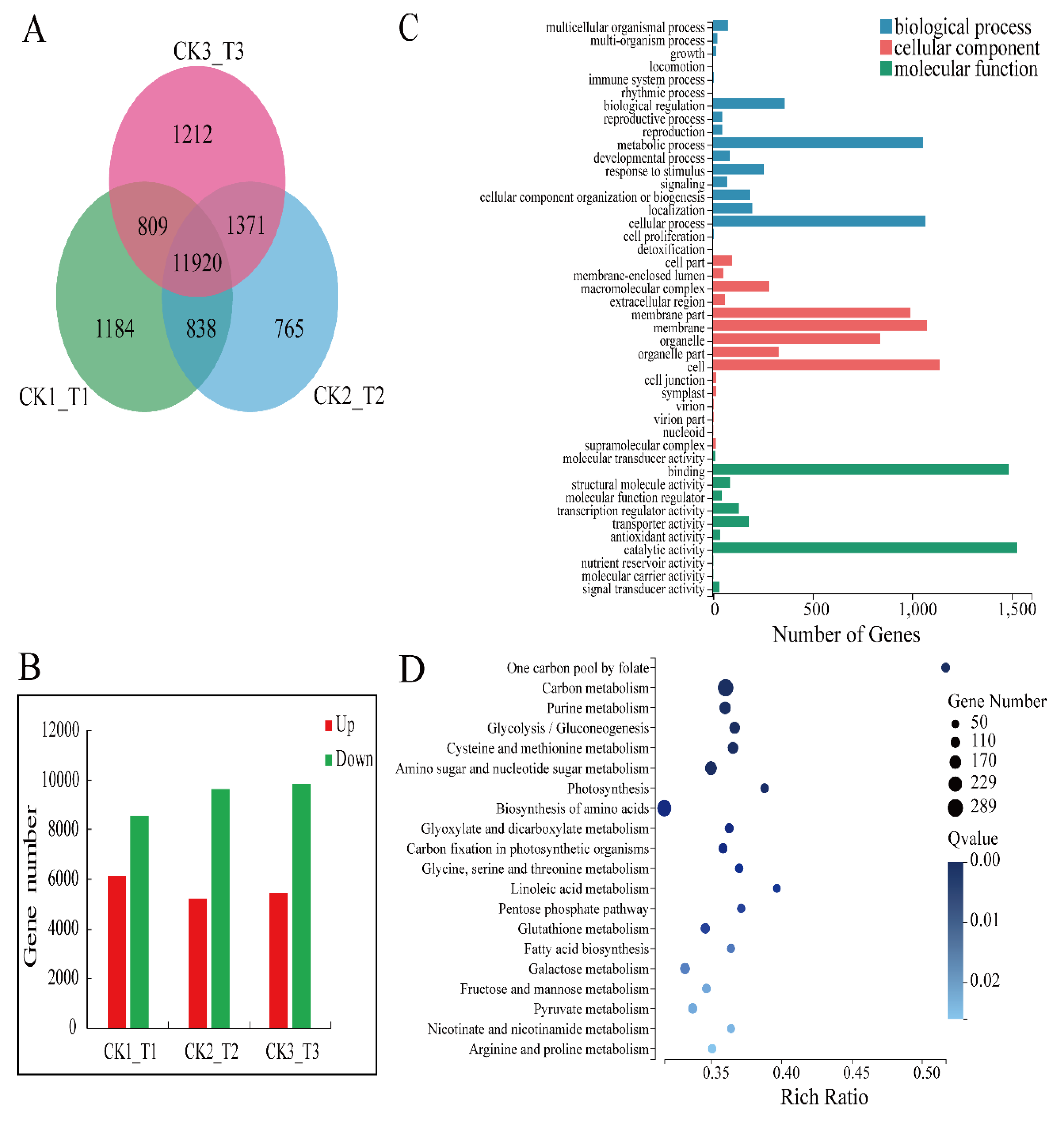
2.4. RNA-Seq Data, DEG Profiles, and Functional Annotation of DEGs
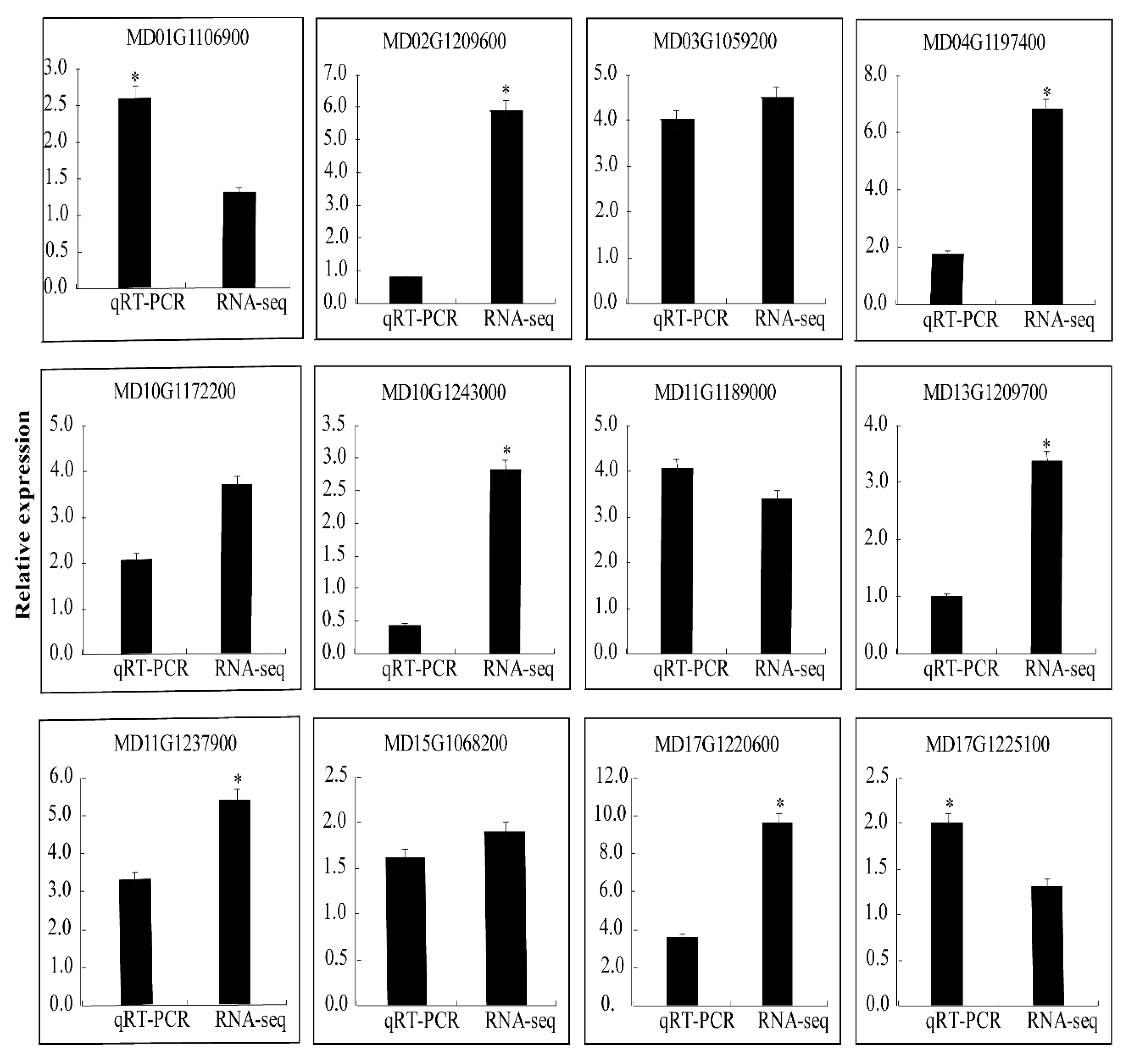
2.5. Validation by qRT-PCR
2.6. Changes in DEGs Related to Phytohormones
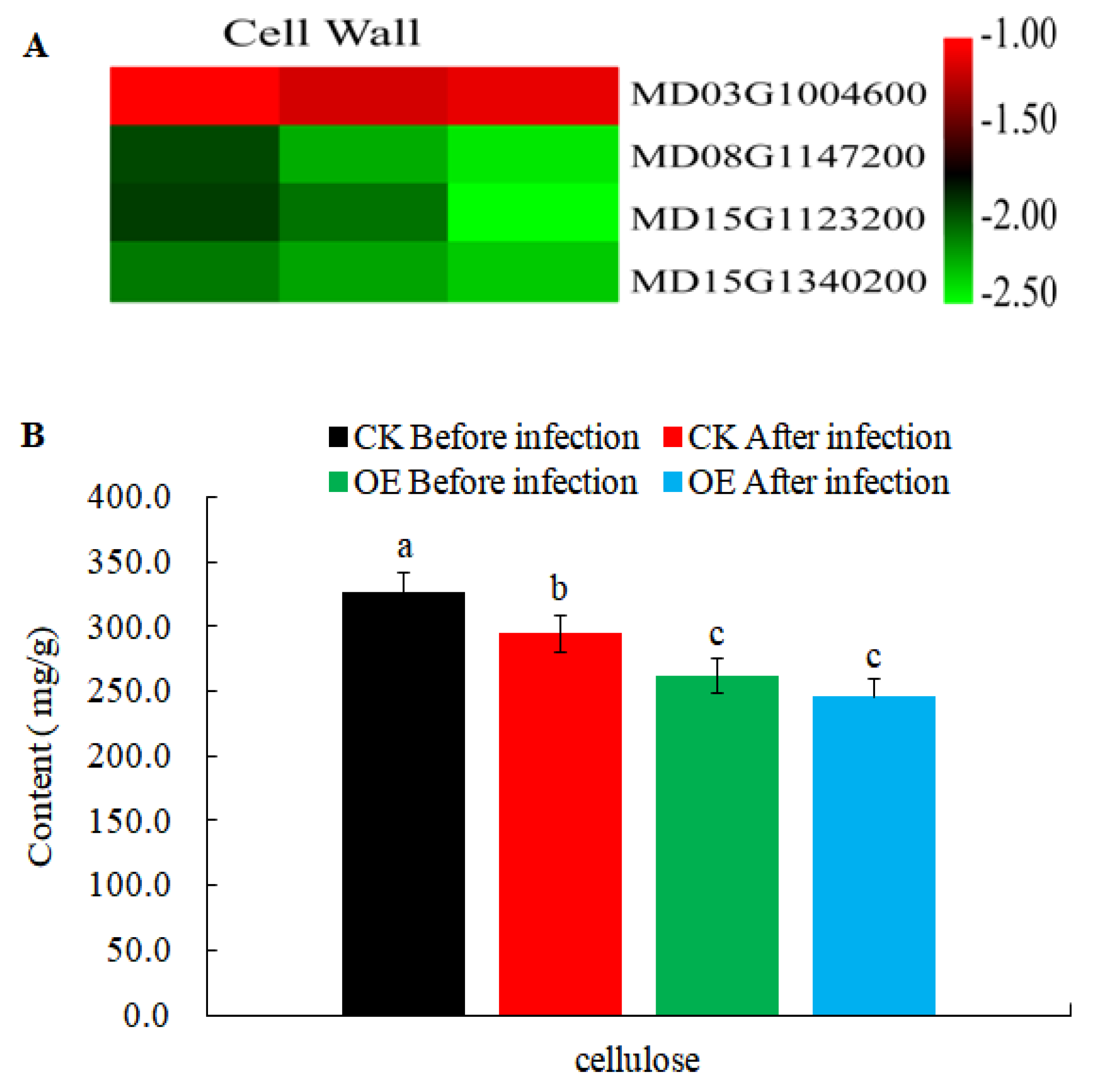
2.7. Changes in DEGs Related to Cell Wall
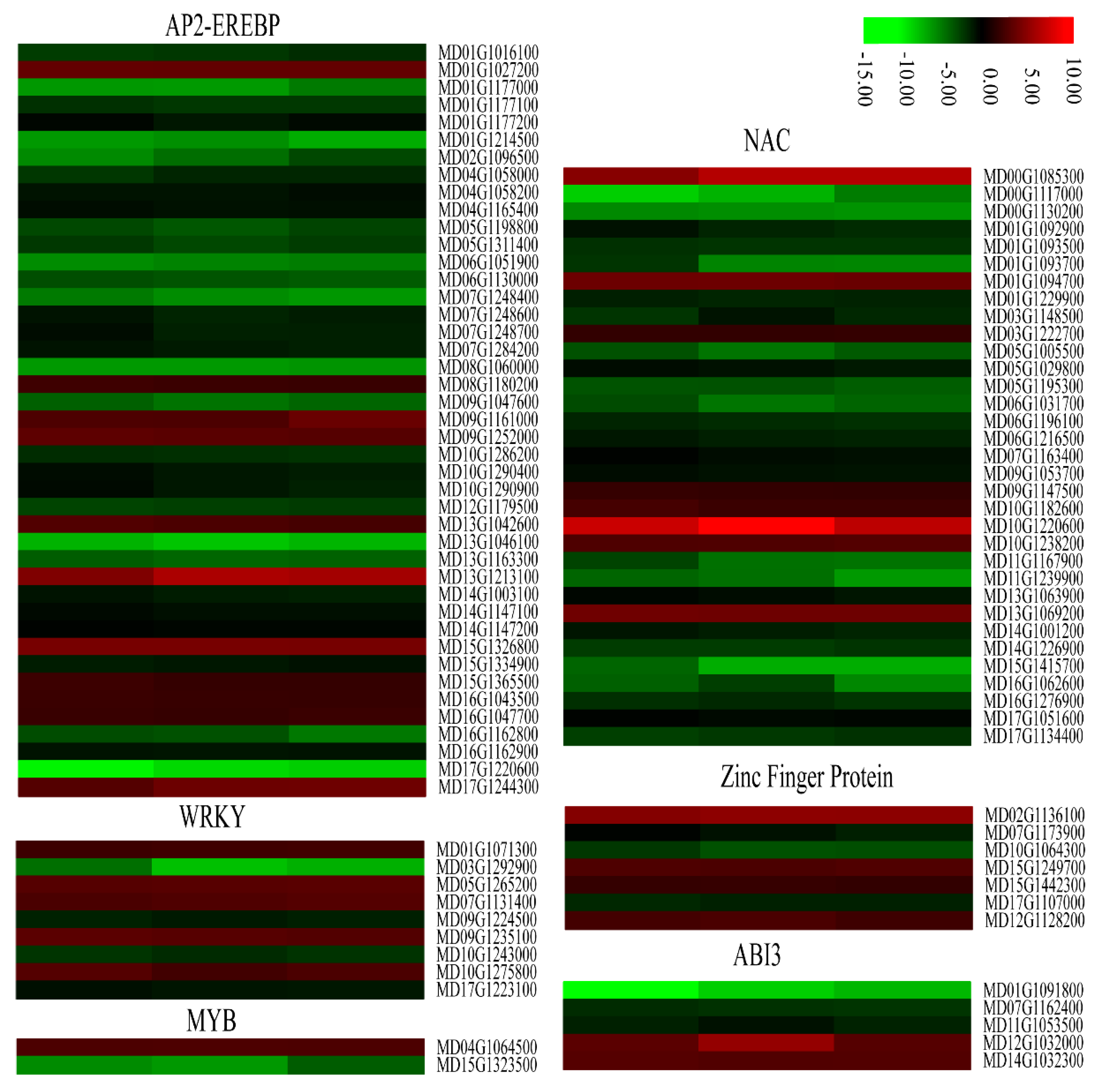
2.8. Changes in DEGs Related to Transcriptional Factors
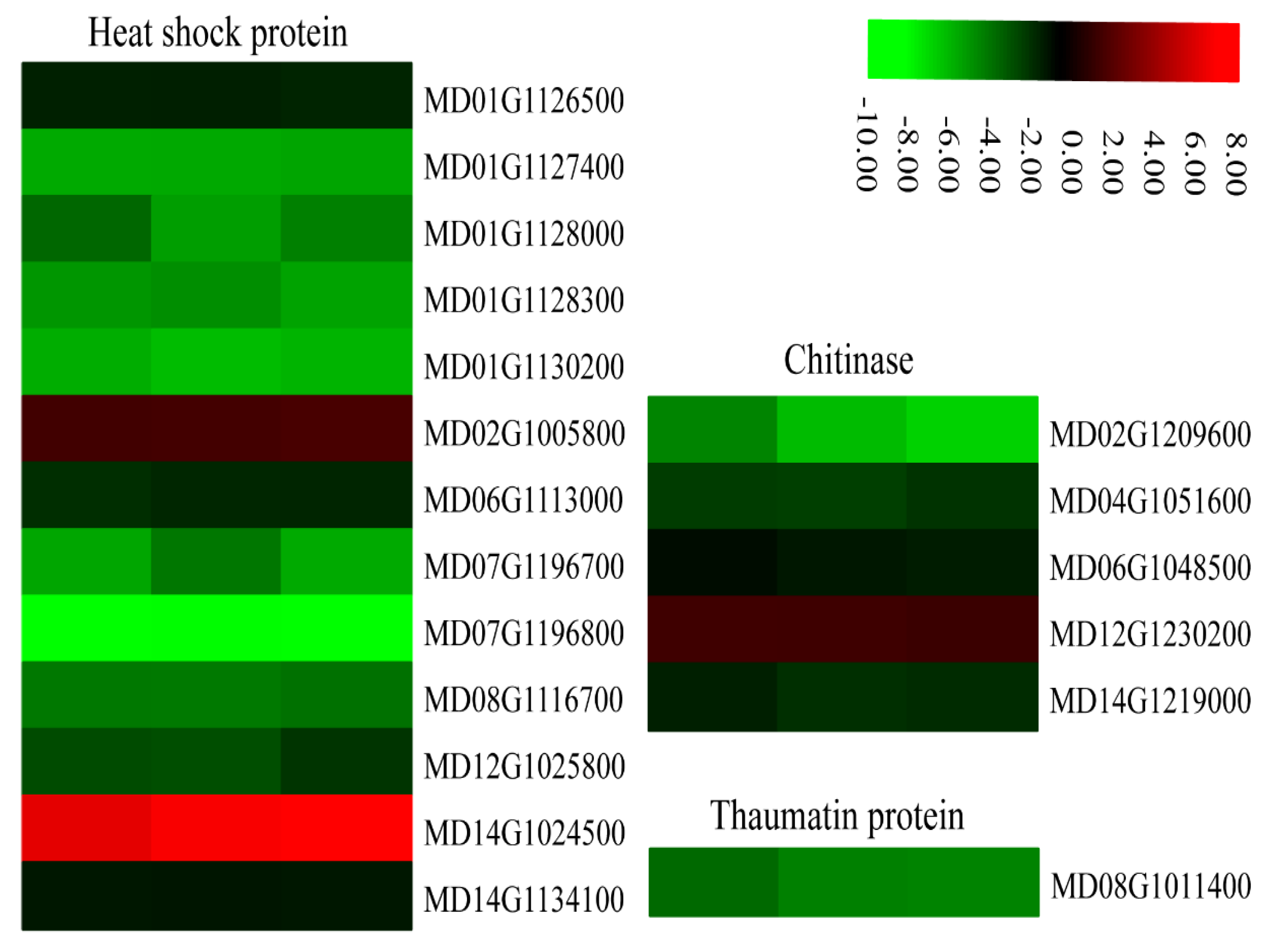
2.9. Changes in DEGs Related to Defense-Related Proteins
3. Discussion
3.1. The Function of MdMLP423
3.2. Changes in Resistance-Related Genes
4. Materials and Methods
4.1. Plant Materials and Treatment
4.2. Cloning and Sequence Analysis of MdMLP423
4.3. Agrobacterium-Mediated Transient Expression Assays with pRI101-AN Constructs
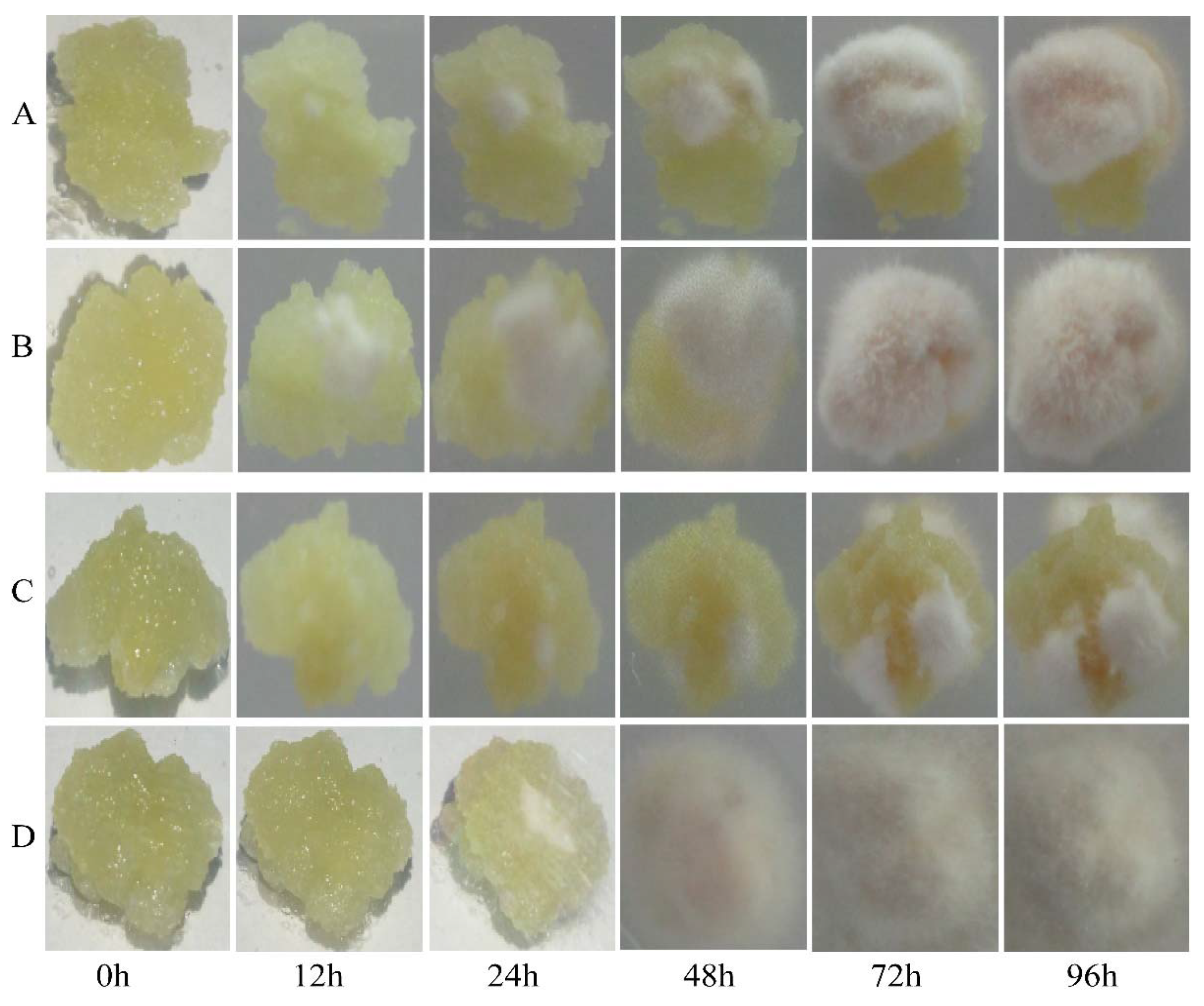
4.4. Disease Resistance Identification of Transgenic Materials
4.5. RNA Extraction and Sequencing
4.6. Quantitative Real Time PCR
4.7. Quantification of the JA, SA Contents, and Cellulose Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MLP/RRP | Major latex protein/ripening-related proteins |
| Gly-rich-loop | Glycine-rich loop |
| BB | Botryosphaeria berengeriana |
| AAAP | Alternaria alternata apple pathotype |
| DEG | Differentially expressed gene |
| HPI | Hours post inoculation |
| SA | Salicylic acid |
| JA | Jasmonic acid |
| ABA | Abscisic acid |
| GA | Gibberellin |
| ETH | Ethylene |
| GD | Golden Delicious apple |
| HF | Hanfu apple |
| HY | Huayue apple |
| NJ | Neighbor-joining |
| Mw | Molecular weight |
| pI | Isoelectric point |
| iTRAQ | Isobaric Tags for Relative and Absolute Quantitation |
| LTR | Low-temperature responsive element |
| ME | Meristem expression |
| DSR | Cis-acting element involved in defense and stress responsiveness |
| HPLC | High-performance liquid chromatography |
References
- Nessler, C.L.; Allen, R.D.; Galewsky, S. Identification and Characterization of Latex-Specific Proteins in Opium Poppy. Plant Physiol. 1985, 79, 499–504. [Google Scholar] [CrossRef]
- Nessler, C.L.; Kurz, W.G.W.; Pelcher, L.E. Isolation and analysis of the major latex protein genes of opium poppy. Plant Mol. Biol. 1990, 15, 951–953. [Google Scholar] [CrossRef]
- Suyama, T.; Yamada, K.; Mori, H.; Takeno, K.; Yamaki, S. Cloning cDNAs for Genes Preferentially Expressed during Fruit Growth in Cucumber. J. Am. Soc. Hortic. Sci. 1999, 124, 136–139. [Google Scholar] [CrossRef]
- Ruperti, B.; Bonghi, C.; Ziliotto, F.; Pagni, S.; Rasori, A.; Varotto, S.; Tonutti, P.; Giovannoni, J.J.; Ramina, A. Characterization of a major latex protein (MLP) gene down-regulated by ethylene during peach fruitlet abscission. Plant Sci. 2002, 163, 265–272. [Google Scholar] [CrossRef]
- Sun, H.; Kim, M.-K.; Pulla, R.K.; Kim, Y.-J.; Yang, D.-C. Isolation and expression analysis of a novel major latex-like protein (MLP151) gene from Panax ginseng. Mol. Biol. Rep. 2009, 37, 2215–2222. [Google Scholar] [CrossRef]
- Lytle, B.L.; Song, J.; De La Cruz, N.B.; Peterson, F.C.; Johnson, K.A.; Bingman, C.A.; Phillips, G.N.; Volkman, B.F. Structures of twoArabidopsis thalianamajor latex proteins represent novel helix-grip folds. Proteins Struct. Funct. Bioinform. 2009, 76, 237–243. [Google Scholar] [CrossRef]
- Inui, H.; Sawada, M.; Goto, J.; Yamazaki, K.; Kodama, N.; Tsuruta, H.; Eun, H. A major latex-like protein is a key factor in crop contamination by persistent organic pollutants. Plant Physiol. 2013, 161, 2128–2135. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef]
- Zhang, N.; Li, R.; Shen, W.; Jiao, S.; Zhang, J.-X.; Xu, W. Genome-wide evolutionary characterization and expression analyses of major latex protein (MLP) family genes in Vitis vinifera. Mol. Genet. Genom. 2018, 293, 1061–1075. [Google Scholar] [CrossRef]
- Radauer, C.; Lackner, P.; Breiteneder, H. The Bet v 1 fold: An ancient, versatile scaffold for binding of large, hydrophobic ligands. BMC Evol. Biol. 2008, 8, 286. [Google Scholar] [CrossRef]
- Park, C.-J.; Kim, K.-J.; Shin, R.; Park, J.M.; Shin, Y.-C.; Paek, K.-H. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. Plant J. 2004, 37, 186–198. [Google Scholar] [CrossRef]
- Pühringer, H.; Moll, W.-D.; Hoffmann-Sommergruber, K.; Watillon, B.; Katinger, H.; Machado, M.L.D.C. The promoter of an apple Ypr10 gene, encoding the major allergen Mald1, is stress- and pathogen-inducible. Plant Sci. 2000, 152, 35–50. [Google Scholar] [CrossRef]
- Lee, S.-R.; Cho, K.-O. Precambrian Crustal Evolution of the Korean Peninsula. J. Pet. Soc. Korea 2012, 21, 89–112. [Google Scholar] [CrossRef]
- Bantignies, B.; Séguin, J.; Muzac, I.; Dédaldéchamp, F.; Gulick, P.; Ibrahim, R. Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots. Plant Mol. Biol. 2000, 42, 871–881. [Google Scholar] [CrossRef]
- Fernandes, H.; Michalska, K.; Sikorski, M.; Jaskolski, M. Structural and functional aspects of PR-10 proteins. FEBS J. 2013, 280, 1169–1199. [Google Scholar] [CrossRef]
- Yang, C.-L.; Liang, S.; Wang, H.-Y.; Han, L.-B.; Wang, F.-X.; Cheng, H.-Q.; Wu, X.; Qu, Z.-L.; Wu, J.; Xia, G. Cotton Major Latex Protein 28 Functions as a Positive Regulator of the Ethylene Responsive Factor 6 in Defense against Verticillium dahliae. Mol. Plant 2015, 8, 399–411. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Chen, J.Y.; Dai, X.F. Cloning and characterization of the Gossypium hirsutum major latex protein gene and functional analysis in Arabidopsis thaliana. Planta 2010, 231, 861–873. [Google Scholar] [CrossRef]
- Li, S.L. Proteomics Analysis of Cotton and Whitefly Interaction and Functional Analysis of GhMLP423 Gene. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Zhu, L.; Ni, W.; Liu, S.; Cai, B.; Xing, H.; Wang, S. Transcriptomics Analysis of Apple Leaves in Response to Alternaria alternata Apple Pathotype Infection. Front. Plant Sci. 2017, 8, 22. [Google Scholar] [CrossRef]
- Zhang, C.X.; Yuan, G.P.; Han, X.L.; Tian, Y.; Zhang, L.Y.; Cong, P.H. Application of iTRAQ-based proteomic approach for screening of resistant associated proteins in ‘Huayue’ apple leaves induced by Alternaria mali. Acta Phytopathol. Sin. 2018, 48, 787–798. [Google Scholar]
- Zhang, Y. Proteomic Analysis of Tobacco Lines with Different Resistance to Brown Spot (Alternaria alternate) and Functional Study of NbMLP423. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2018. [Google Scholar]
- Devoto, A.; Turner, J.G. Regulation of jasmonate-mediated plant responses in Arabidopsis. Ann Bot. 2003, 92, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Chan, Z. Expression profiling of ABA pathway transcripts indicates crosstalk between abiotic and biotic stress responses in Arabidopsis. Genomics 2012, 100, 110–115. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Chen, X.; Ye, T.; Zhong, B.; Liu, R.; Wu, Y.; Chan, Z. Major latex protein-like protein 43 (MLP43) functions as a positive regulator during abscisic acid responses and confers drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2015, 67, 421–434. [Google Scholar] [CrossRef]
- Ecker, J.R. The ethylene signal transduction pathway in plants. Science 1995, 268, 667–675. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Gheysen, G.; Hofte, M. Hormone defense networking in rice: Tales from a different world. Trends Plant Sci. 2013, 18, 555–565. [Google Scholar] [CrossRef]
- Zhang, C.X.; Tian, Y.; Zhang, L.Y.; Zong, Z.R.; Cong, P.H. Comparative proteomic analysis of apple branches susceptible and resistant to ring rot disease. Eur. J. Plant Pathol. 2017, 148, 329–341. [Google Scholar]
- Yang, Z.Y.; Kang, G.D.; Wang, Q.; Xue, G.R.; Zhang, L.Y.; Tian, Y.; Yang, L.; Zhang, C.X.; Li, W.X.; Cong, P.H. The New Yellow Apple Cultivar ‘Huayue’. Acta Hortic. Sin. 2010, 37, 1877–1878. [Google Scholar]
- Ding, A.R.; Sun, X.H.; Hou, H.M.; Sun, X.; Bai, S.H.; Zhang, Y.G.; Dai, H.Y. Cloning and Expression Analysis of MdERF113 Gene from Apple Malus x domestica Infected with Botryosphaeria dothidea. J. Qingdao Agric. Univ. (Nat. Sci.) 2018, 35, 161–170. [Google Scholar]
- Zhang, Q.L.; Ma, C.; Zhang, Y.; Gu, Z.Y.; Li, W.; Duan, X.W.; Wang, S.N.; Hao, L.; Wang, Y.H.; Wang, S.Y.; et al. A single nucleotide polymorphism in the promoter of a hairpin RNA contributes to Alternaria leaf spot resistance in apple (Malus × domestica Borkh.). Plant Cell 2018, 30, 1924–1942. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, J.; Han, X.; Li, J.; Gao, Y.; Richards, C.M.; Zhang, C.; Tian, Y.; Liu, G.; Gul, H.; et al. A high-quality apple genome assembly reveals the association of a retrotransposon and red fruit colour. Nat. Commun. 2019, 10, 1494. [Google Scholar] [CrossRef] [PubMed]
- Verma, J.K.; Kishore, D.K.; Sharma, S.K.; Sharma, A. Evaluation of Biotic and Abiotic Factors for Production of Healthy Apple (Malus × domestica) Seedling. In Mocrobial Diversity and Biotechnology in Food Security; Kharwar, R.N., Ed.; Springer: New Delhi, India, 2014; Volume 21, pp. 243–249. [Google Scholar]
- Li, R. Investigation on Major Latex Protein423 (MdMLP423) from Apple in Response to Abiotic Stress. Master’s Thesis, Shandong Agricultural University, Taian, China, 2018. [Google Scholar]
- Schenk, P.; Kazan, K.; Wilson, I.; Anderson, J.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [PubMed]
- Siemens, D.H.; Garner, S.H.; Mitchell-Olds, T.; Callaway, R.M. Cost of defense in the context of plant competition: Brassica rapa may grow and defend. Ecology 2002, 83, 505–517. [Google Scholar] [CrossRef]
- Malter, D.; Wolf, S. Melon phloem-sap proteome: Developmental control and response to viral infection. Protoplasma 2011, 248, 217–224. [Google Scholar] [CrossRef]
- Qu, Z.L.; Wang, H.Y.; Xia, G.X. GhHbl: A nonsymbiotic hemoglobin gene of cotton responsive to infection by Verticillium dahliae. Biochim. Biophys. Acta 2005, 1730, 103–113. [Google Scholar] [CrossRef]
- Wang, F.-X.; Ma, Y.-P.; Yang, C.-L.; Zhao, P.-M.; Yao, Y.; Jian, G.-L.; Luo, Y.-M.; Xia, G. Proteomic analysis of the sea-island cotton roots infected by wilt pathogen Verticillium dahliae. Proteomics 2011, 11, 4296–4309. [Google Scholar] [CrossRef]
- Zhang, W.-W.; Jian, G.-L.; Jiang, T.-F.; Wang, S.-Z.; Qi, F.-J.; Xu, S.-C. Cotton gene expression profiles in resistant Gossypium hirsutum cv. Zhongzhimian KV1 responding to Verticillium dahliae strain V991 infection. Mol. Boil. Rep. 2012, 39, 9765–9774. [Google Scholar]
- Thomma, B.; Cammue, B.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef]
- Jin, Z.M.; Ping, B.Z.; Shen, H.J.; Du, H.Q.; Li, R.Q.; Zhu, L.; Zhang, D.B.; Yuan, Z. Characterisation and gene mapping of a britle culm mutant bc-s1 in rice. Chin. Bull. Bot. 2016, 51, 167–174. [Google Scholar]
- Koga, H.; Dohi, K.; Mori, M. Abscisic acid and low temperatures suppress the whole plant-specific resistance reaction of rice plants to the infection of Magnaporthe grisea. Physiol. Mol. Plant Pathol. 2004, 65, 3–9. [Google Scholar] [CrossRef]
- Xu, J.; Audenaert, K.; Hofte, M.; De Vleesschauwer, D. Abscisic Acid Promotes Susceptibility to the Rice Leaf Blight Pathogen Xanthomonas oryzae pv oryzae by Suppressing Salicylic Acid-Mediated Defenses. PLoS ONE 2013, 8, e67413. [Google Scholar]
- Fu, D.-Q.; Ghabrial, S.; Kachroo, A. GmRAR1 and GmSGT1 Are Required for Basal, R Gene—Mediated and Systemic Acquired Resistance in Soybean. Mol. Plant-Microbe Interactions 2009, 22, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.Y.; Yoshioka, K.; Desveaux, D. The roles of ABA in plant–pathogen interactions. J. Plant Res. 2011, 124, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2005, 5, 325–331. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Bruce, A.T.A.; Julián, P.P.; Manuel, M.P.P.; Marta, G.; Sánchez-Serrano, J.J.; Eric, A.S.; Roberto, S. ABA Is an Essential Signal for Plant Resistance to Pathogens Affecting JA Biosynthesis and the Activation of Defenses in Arabidopsis. Plant Cell 2007, 19, 1665–1681. [Google Scholar]
- Jensen, M.K.; Rung, J.H.; Gregersen, P.; Gjetting, T.; Fuglsang, A.T.; Hansen, M.; Joehnk, N.; Lyngkjaer, M.F.; Collinge, D.B.; Lyngkjær, M. The HvNAC6 transcription factor: A positive regulator of penetration resistance in barley and Arabidopsis. Plant Mol. Biol. 2007, 65, 137–150. [Google Scholar] [CrossRef]
- Kaneda, T.; Taga, Y.; Takai, R.; Lwano, M.; Matsui, H.; Takayama, S.; Isogai, A.; Che, F.S. The transcription factor OsNAC4 is a key positive regulator of plant hypersensitive cell death. EMBO J. 2009, 28, 926–936. [Google Scholar] [CrossRef]
- Seo, P.J.; Kim, M.J.; Park, J.Y.; Kim, S.Y.; Jeon, J.; Lee, Y.H.; Kim, J.; Park, C.M. Cold activation of a plasma membrane-tethered NAC transcription factor induces a pathogen resistance response in Arabidopsis. Plant J. 2010, 61, 661–671. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, H.C.; Kim, K.E.; Jung, M.S.; Han, H.J.; Kim, S.H.; Kwon, Y.S.; Bahk, S.; An, J.; Bae, D.W.; et al. A NAC transcription factor and SNI1 cooperatively suppress basal pathogen resistance in Arabidopsis thaliana. Nucleic Acids Res. 2012, 40, 9182–9192. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Ivanova, A.; Duncan, O.; Law, S.R.; Van Aken, O.; De Clercq, I.; Wang, Y.; Carrie, C.; Xu, L.; Kmiec, B.; et al. A membrane-bound NAC transcription factor, ANAC017, mediates mitochondrial retrograde signaling in Arabidopsis. Plant Cell 2013, 25, 3450–3471. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.D.; Xie, X.; Wei, F.J. Research progress of HSP70s in Poaceae. Plant Physiol. J. 2019, 55, 1054–1062. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 1, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Daccord, N.; Celton, J.-M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; Van De Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Xia, R.; Chen, H.; He, Y.H. TBtools, a Toolkit for Biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Chen, M.; Liu, X.W.; Shen, F.F.; Wang, P. A Highly Efficient System Establishment of Transient Expression in Lettuce. Acta Hortic. Sin. 2005, 2, 405–407. [Google Scholar]
- Zong, Z.R.; Tian, Y.; Zhang, L.Y.; Han, X.L.; Zhang, C.X.; Cong, P.H. Cloning and Function Identification of Apple Alternaria Blotch Resistant Gene Mal d 1. Acta Hortic. Sin. 2017, 44, 343–354. [Google Scholar]
- Lv, P.P. The Construction of Apple MdBAKIS Genes Expression Vector, Callus Transformation and Apple Ring Rot Disease Resistance Characterization. Master’s Thesis, Shandong Agricultural University, Taian, China, 2016. [Google Scholar]
- Yuan, G.; Bian, S.; Han, X.; He, S.; Liu, K.; Zhang, C.; Pei-Hua, C. An Integrated Transcriptome and Proteome Analysis Reveals New Insights into Russeting of Bagging and Non-Bagging “Golden Delicious” Apple. Int. J. Mol. Sci. 2019, 20, 4462. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔ CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Zhu, H.; Xu, B.; Huang, W.; Wang, S.; Ding, Y.; Liu, Z.; Li, G.; Chen, L.; Ding, C.; et al. Effect of Removing Superior Spikelets on Grain Filling of Inferior Spikelets in Rice. Front. Plant Sci. 2016, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.M. Study on Physiological and Molecular Mechanism of Fruit Texture Development and Pest-Harvest Regulation of Apple (Males Domestica Borkh). Ph.D. Thesis, Northwest A&F University, Yangling, China, 2016. [Google Scholar]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, S.; Yuan, G.; Bian, S.; Han, X.; Liu, K.; Cong, P.; Zhang, C. Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple. Int. J. Mol. Sci. 2020, 21, 1879. https://doi.org/10.3390/ijms21051879
He S, Yuan G, Bian S, Han X, Liu K, Cong P, Zhang C. Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple. International Journal of Molecular Sciences. 2020; 21(5):1879. https://doi.org/10.3390/ijms21051879
Chicago/Turabian StyleHe, Shanshan, Gaopeng Yuan, Shuxun Bian, Xiaolei Han, Kai Liu, Peihua Cong, and Caixia Zhang. 2020. "Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple" International Journal of Molecular Sciences 21, no. 5: 1879. https://doi.org/10.3390/ijms21051879
APA StyleHe, S., Yuan, G., Bian, S., Han, X., Liu, K., Cong, P., & Zhang, C. (2020). Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple. International Journal of Molecular Sciences, 21(5), 1879. https://doi.org/10.3390/ijms21051879