Claudin-1, A Double-Edged Sword in Cancer

 ,
, 
 ,
,  and
and
Abstract
1. Introduction
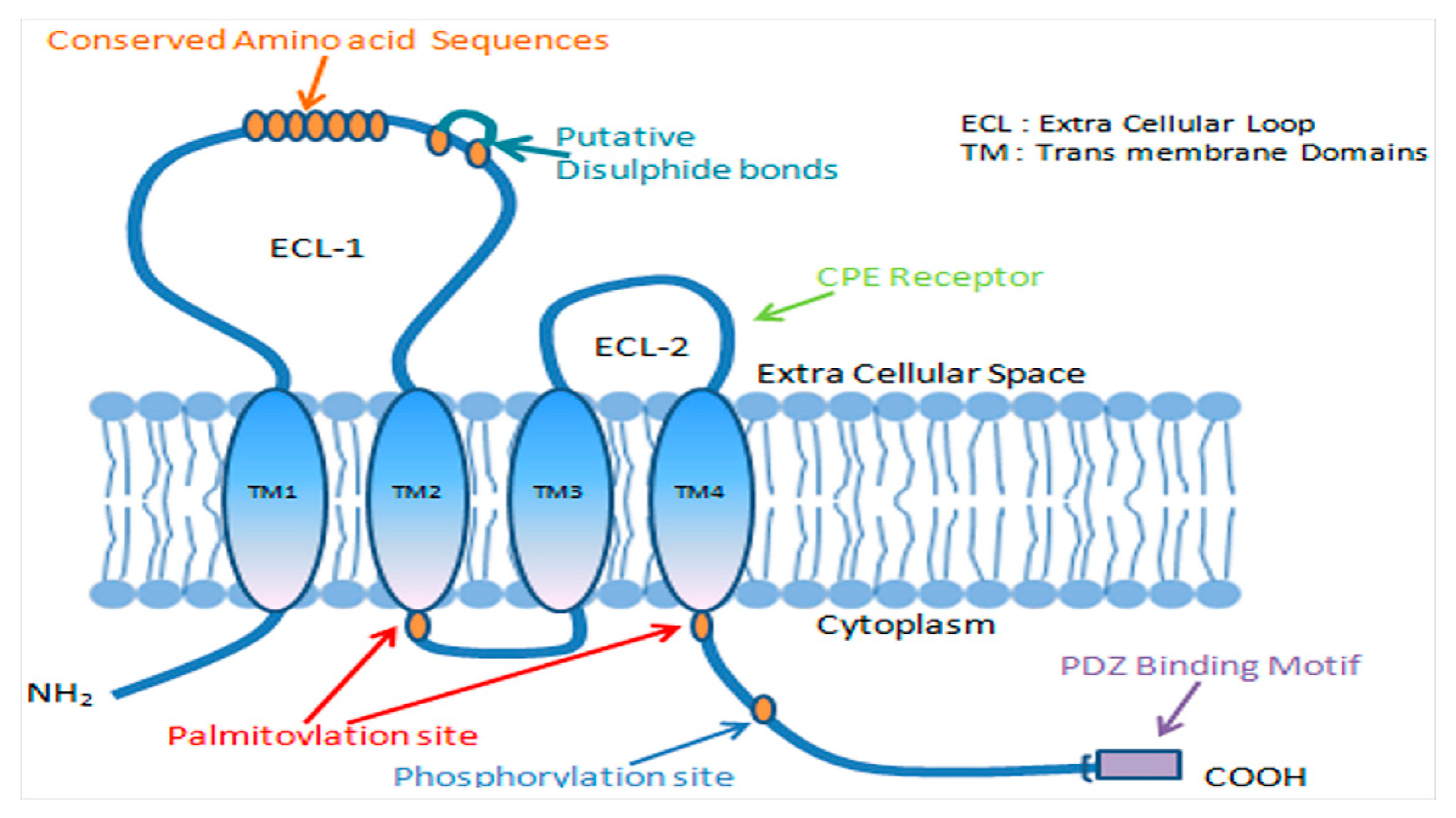
1.1. Tight Junctions
1.2. Claudins
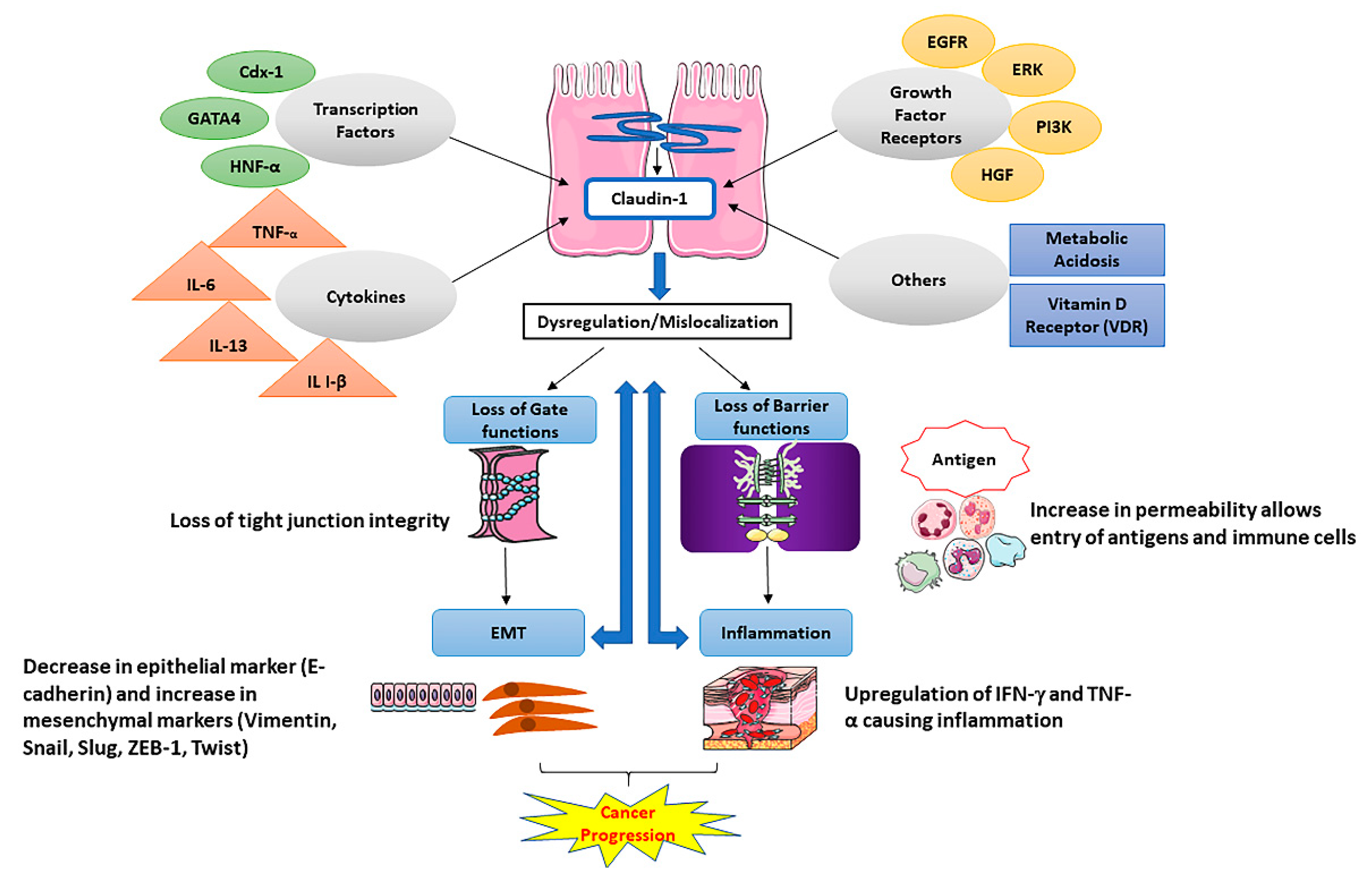
1.3. Claudins and Cancer
2. Claudin-1 and Cancer; Tumor Promoter or Suppressor
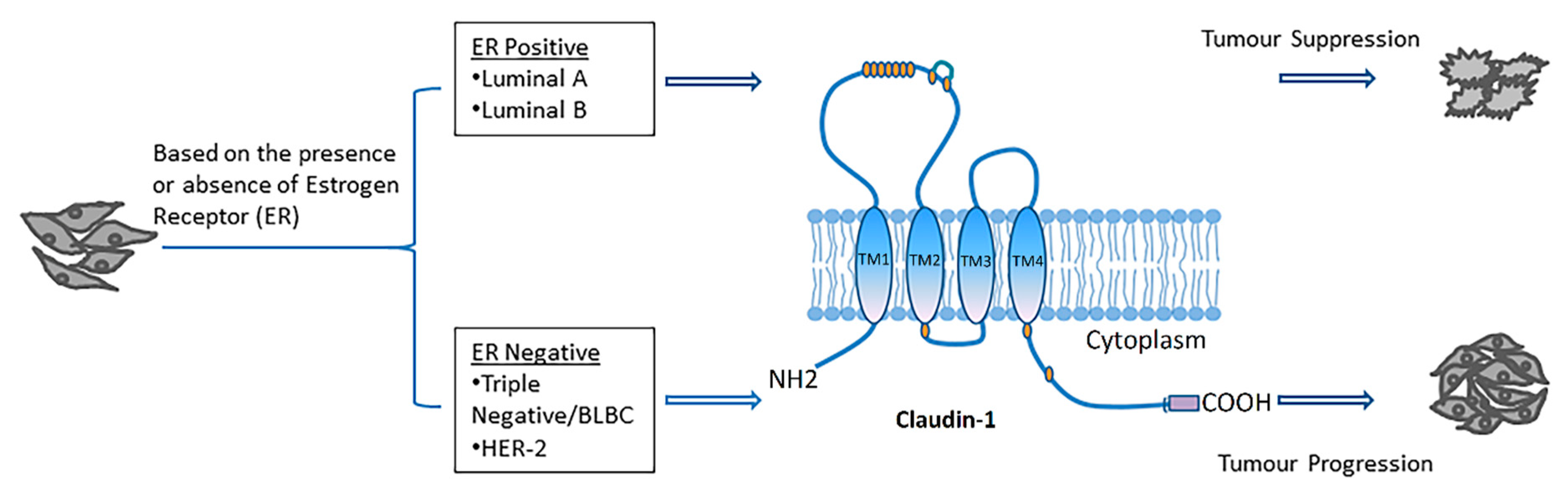
2.1. Claudin-1 and Breast Cancer
2.2. Claudin-1 and Thyroid Cancer
2.3. Claudin-1 and Colorectal Cancer
2.4. Claudin-1 and Gastric Cancer
2.5. Claudin-1 and Hypopharyngeal Squamous Cell Carcinoma
2.6. Claudin-1 and Hepatocellular Carcinoma
2.7. Claudin-1 and Lung Adenocarcinoma
2.8. Claudin-1 and Pancreatic Cancer
2.9. Claudin-1 and Ovarian Cancer
2.10. Claudin-1 and Oral Squamous Cell Carcinoma
2.11. Claudin-1 and Melanoma
2.12. Claudin-1 and Prostate Cancer
3. Claudin-1 and Tight Junction Barrier Function
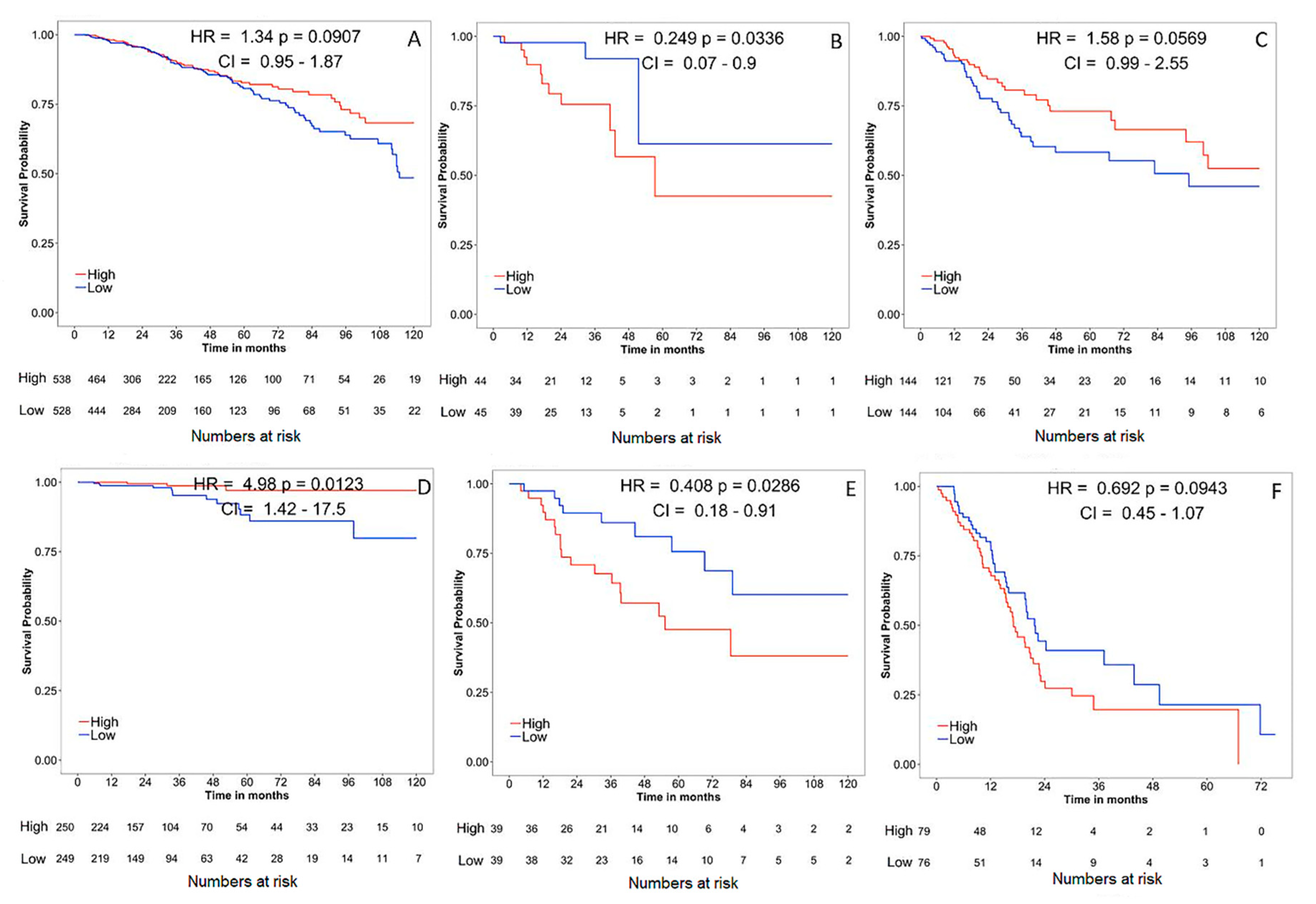
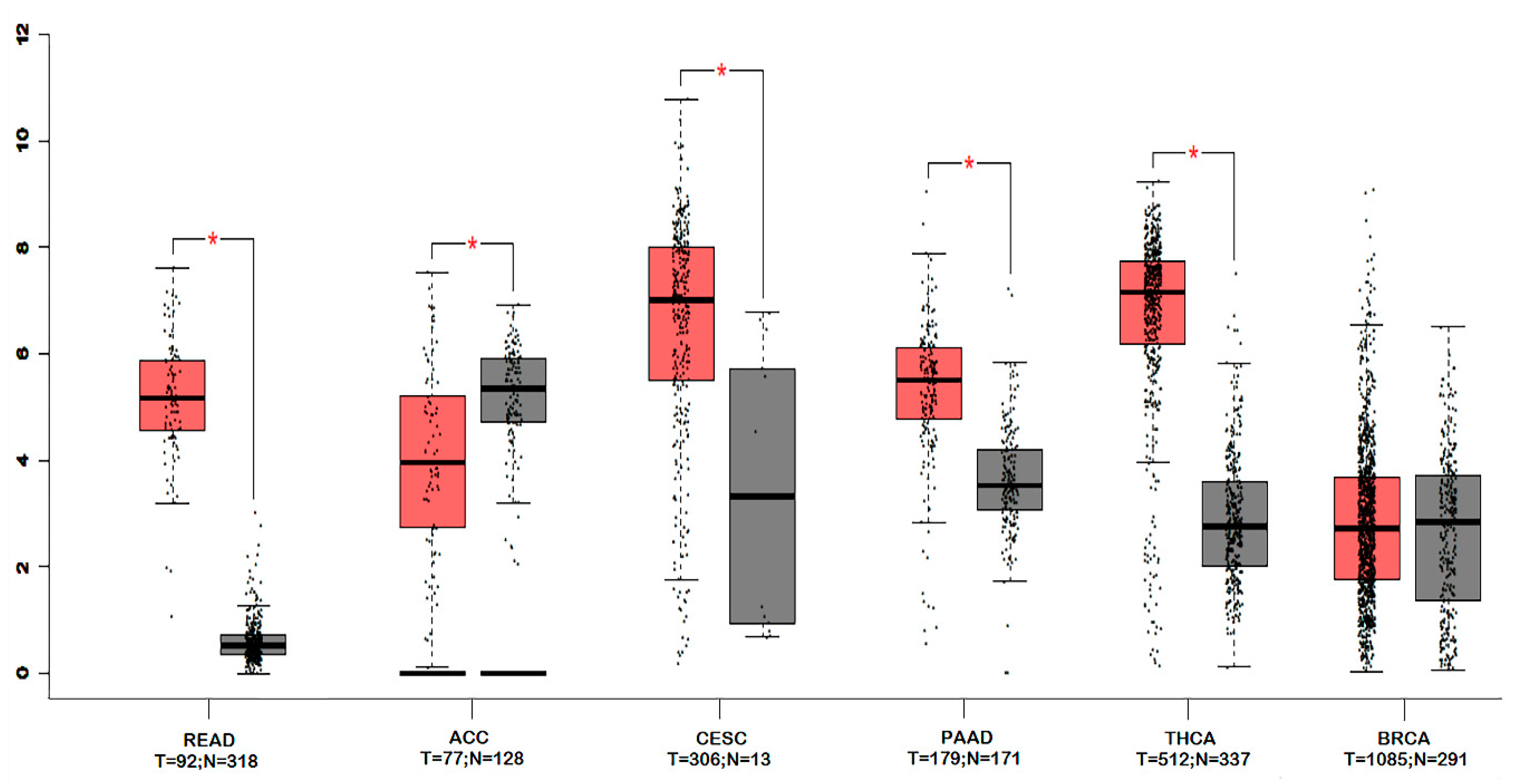
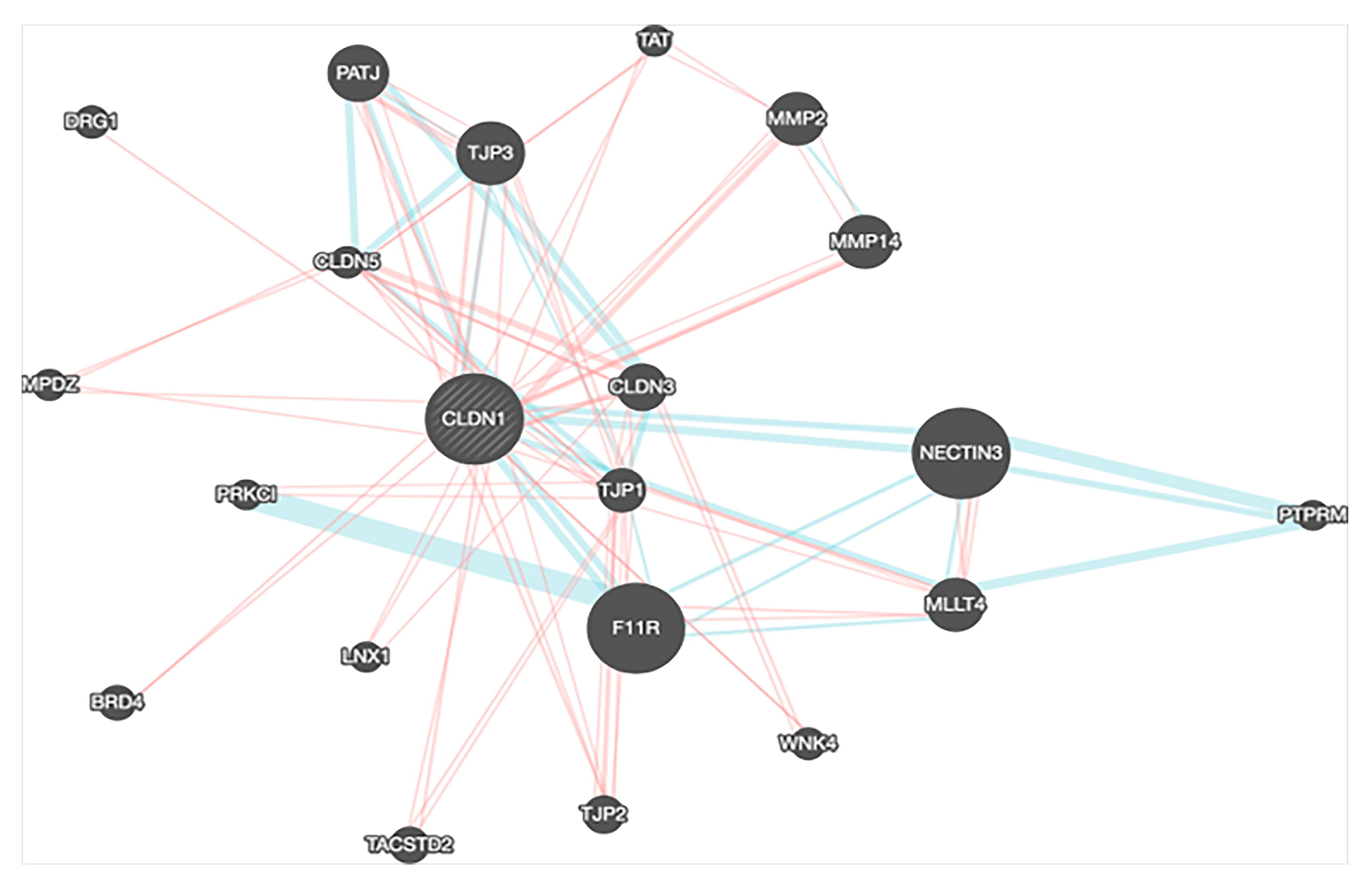
4. Gene Expression, Survival and Pathway Interaction Analysis of claudin-1 across different Cancers
5. Claudin-1 as a Drug Target
6. Claudins and Autosomal Recessive Disorders
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CLDN-1 | Claudin-1 protein |
| CLDN-1 | Claudin-1 gene |
| EMT | Epithelial-mesenchymal transition |
| CSCs | Cancer stem cells |
| HER2 | Human epidermal growth factor receptor 2 |
| BLBC | Basal-like breast cancer |
| ER | Estrogen receptor |
| ESCRT | Endosomal sorting complexes required for transport |
| EpCAM | Epithelial cell adhesion molecule |
| PTC | Papillary thyroid cancer |
| FTC | Follicular thyroid cancer |
| CRC | Colorectal cancer |
| mCRC | Metastatic colorectal cancer |
| UC | Ulcerative colitis |
| APC | Adenomatous polyposis coli |
| HNSCC | Head and neck squamous cell carcinoma |
| HSCC | Hypopharyngeal squamous cell carcinoma |
| HCC | Hepatocellular carcinoma |
| CTGF | Connective tissue growth factor |
| THBS1 | Thrombospondin 1 |
| DLC1 | Deleted in liver cancer 1 |
| OCLN | Occludin |
| ZO-1 | Zona occludins 1 |
| SPP1 | Secreted phosphoprotein 1 |
| CUTL1 | Cut-like homeobox 1 |
| TGF-α | Transforming growth factor-alpha |
| SLC2A3 | Solute carrier family 2 (facilitated glucose transporter) member 3 |
| PGF | Placental growth factor |
| EGFR | Epidermal growth factor |
| PC | Pancreatic cancer |
| MEK2 | Mitogen activated protein kinase 2 |
| miR-155 | microRNA-155 |
| BOT | Borderline tumors of the ovary |
| OSCC | Oral squamous cell carcinoma |
| MMP | Matrix metalloproteinase |
| BRCA | Breast invasive carcinoma |
| READ | Rectum adenocarcinoma |
| CESC | Cervical squamous cell carcinoma |
| THCA | Thyroid carcinoma |
| ACC | Adrenocortical carcinoma |
| PAAD | Pancreatic adenocarcinoma |
| mAbs | Monoclonal antibodies |
| cCPE | Clostridium perfringens enterotoxin |
| HCV | Hepatitis C virus |
| TCGA | The cancer genome atlas |
References
- Tsukita, S.; Yamazaki, Y.; Katsuno, T.; Tamura, A.; Tsukita, S. Tight junction-based epithelial microenvironment and cell proliferation. Oncogene 2008, 27, 6930–6938. [Google Scholar] [CrossRef]
- Itoh, M.; Bissell, M.J. The organization of tight junctions in epithelia: Implications for mammary gland biology and breast tumorigenesis. J. Mammary Gland Biol. Neoplasia 2003, 8, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Furuse, M.; Itoh, M. Multifunctional strands in tight junctions. Nat. Rev. Mol. Cell Biol. 2001, 2, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.; Winkler, L.; Mueller, S.L.; Haseloff, R.F.; Piontek, J.; Blasig, I.E. Structure and function of claudins. Biochim. Biophys. Acta 2008, 1778, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Furuse, M. Claudin-based barrier in simple and stratified cellular sheets. Curr. Opin. Cell Biol. 2002, 14, 531–536. [Google Scholar] [CrossRef]
- Anderson, J.M.; Van Itallie, C.M. Physiology and function of the tight junction. Cold Spring Harb. Perspect. Biol. 2009, 1, a002584. [Google Scholar] [CrossRef]
- Kirschner, N.; Rosenthal, R.; Furuse, M.; Moll, I.; Fromm, M.; Brandner, J.M. Contribution of tight junction proteins to ion, macromolecule, and water barrier in keratinocytes. J. Investig. Derm. 2013, 133, 1161–1169. [Google Scholar] [CrossRef]
- Soler, A.P.; Miller, R.D.; Laughlin, K.V.; Carp, N.Z.; Klurfeld, D.M.; Mullin, J.M. Increased tight junctional permeability is associated with the development of colon cancer. Carcinogenesis 1999, 20, 1425–1431. [Google Scholar] [CrossRef]
- Singh, A.B.; Sharma, A.; Dhawan, P. Claudin family of proteins and cancer: An overview. J. Oncol. 2010, 2010, 541957. [Google Scholar] [CrossRef]
- Kwon, M.J. Emerging roles of claudins in human cancer. Int. J. Mol. Sci. 2013, 14, 18148–18180. [Google Scholar] [CrossRef]
- Saraydaroglu, O.; Narter, S.; Ozsen, M.; Coskun, H. Non-epithelial tumors of the larynx: Case series of 12 years. Eur. Arch. Oto-Rhino-Laryngol. 2019, 276, 2843–2847. [Google Scholar] [CrossRef] [PubMed]
- Reinert, S. Principles of Management of Neoplastic Salivary Gland Disease. Maxillofacial Surgery, 3rd ed. 2017, pp. 657–667. Available online: https://www.sciencedirect.com/book/9780702060564/maxillofacial-surgery (accessed on 13 January 2020).
- Amasheh, S.; Milatz, S.; Krug, S.M.; Markov, A.G.; Gunzel, D.; Amasheh, M.; Fromm, M. Tight junction proteins as channel formers and barrier builders. Ann. N. Y. Acad. Sci. 2009, 1165, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Ikari, A.; Ito, M.; Okude, C.; Sawada, H.; Harada, H.; Degawa, M.; Sakai, H.; Takahashi, T.; Sugatani, J.; Miwa, M. Claudin-16 is directly phosphorylated by protein kinase a independently of a vasodilator-stimulated phosphoprotein-mediated pathway. J. Cell Physiol. 2008, 214, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Fujibe, M.; Chiba, H.; Kojima, T.; Soma, T.; Wada, T.; Yamashita, T.; Sawada, N. Thr203 of claudin-1, a putative phosphorylation site for map kinase, is required to promote the barrier function of tight junctions. Exp. Cell Res. 2004, 295, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Akizuki, R.; Shimobaba, S.; Matsunaga, T.; Endo, S.; Ikari, A. Claudin-5, -7, and -18 suppress proliferation mediated by inhibition of phosphorylation of akt in human lung squamous cell carcinoma. Biochim. Biophys Acta 2017, 1864, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Findley, M.K.; Koval, M. Regulation and roles for claudin-family tight junction proteins. Iubmb. Life 2009, 61, 431–437. [Google Scholar] [CrossRef]
- Lal-Nag, M.; Morin, P.J. The claudins. Genome Biol. 2009, 10, 235. [Google Scholar] [CrossRef]
- Hewitt, K.J.; Agarwal, R.; Morin, P.J. The claudin gene family: Expression in normal and neoplastic tissues. BMC Cancer 2006, 6, 186. [Google Scholar] [CrossRef]
- Pope, J.L.; Bhat, A.A.; Sharma, A.; Ahmad, R.; Krishnan, M.; Washington, M.K.; Beauchamp, R.D.; Singh, A.B.; Dhawan, P. Claudin-1 regulates intestinal epithelial homeostasis through the modulation of notch-signalling. Gut 2014, 63, 622–634. [Google Scholar] [CrossRef]
- Singh, A.B.; Uppada, S.B.; Dhawan, P. Claudin proteins, outside-in signaling, and carcinogenesis. Pflug. Arch. 2017, 469, 69–75. [Google Scholar] [CrossRef]
- Turksen, K. Claudins and cancer stem cells. Stem Cell Rev. 2011, 7, 797–798. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Flodby, P.; Luo, J.; Castillo, D.R.; Liu, Y.; Yu, F.X.; McConnell, A.; Varghese, B.; Li, G.; Chimge, N.O.; et al. Claudin-18-mediated yap activity regulates lung stem and progenitor cell homeostasis and tumorigenesis. J. Clin. Investig. 2018, 128, 970–984. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J. Claudin proteins in human cancer: Promising new targets for diagnosis and therapy. Cancer Res. 2005, 65, 9603–9606. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Moodie, A.; Blanchard, A.A.; Leygue, E.; Myal, Y. Claudin 1 in breast cancer: New insights. J. Clin. Med. 2015, 4, 1960–1976. [Google Scholar] [CrossRef]
- Usami, Y.; Chiba, H.; Nakayama, F.; Ueda, J.; Matsuda, Y.; Sawada, N.; Komori, T.; Ito, A.; Yokozaki, H. Reduced expression of claudin-7 correlates with invasion and metastasis in squamous cell carcinoma of the esophagus. Hum. Pathol. 2006, 37, 569–577. [Google Scholar] [CrossRef]
- Miyamoto, K.; Kusumi, T.; Sato, F.; Kawasaki, H.; Shibata, S.; Ohashi, M.; Hakamada, K.; Sasaki, M.; Kijima, H. Decreased expression of claudin-1 is correlated with recurrence status in esophageal squamous cell carcinoma. Biomed. Res. 2008, 29, 71–76. [Google Scholar] [CrossRef]
- Honda, H.; Pazin, M.J.; Ji, H.; Wernyj, R.P.; Morin, P.J. Crucial roles of sp1 and epigenetic modifications in the regulation of the cldn4 promoter in ovarian cancer cells. J. Biol. Chem. 2006, 281, 21433–21444. [Google Scholar] [CrossRef]
- Honda, H.; Pazin, M.J.; D’Souza, T.; Ji, H.; Morin, P.J. Regulation of the cldn3 gene in ovarian cancer cells. Cancer Biol. 2007, 6, 1733–1742. [Google Scholar] [CrossRef]
- Sheehan, G.M.; Kallakury, B.V.; Sheehan, C.E.; Fisher, H.A.; Kaufman, R.P., Jr.; Ross, J.S. Loss of claudins-1 and -7 and expression of claudins-3 and -4 correlate with prognostic variables in prostatic adenocarcinomas. Hum. Pathol. 2007, 38, 564–569. [Google Scholar] [CrossRef]
- Resnick, M.B.; Konkin, T.; Routhier, J.; Sabo, E.; Pricolo, V.E. Claudin-1 is a strong prognostic indicator in stage ii colonic cancer: A tissue microarray study. Mod. Pathol. 2005, 18, 511–518. [Google Scholar] [CrossRef]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl Acad Sci USA 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, P.; Singh, A.B.; Deane, N.G.; No, Y.; Shiou, S.R.; Schmidt, C.; Neff, J.; Washington, M.K.; Beauchamp, R.D. Claudin-1 regulates cellular transformation and metastatic behavior in colon cancer. J. Clin. Investig. 2005, 115, 1765–1776. [Google Scholar] [CrossRef]
- Oku, N.; Sasabe, E.; Ueta, E.; Yamamoto, T.; Osaki, T. Tight junction protein claudin-1 enhances the invasive activity of oral squamous cell carcinoma cells by promoting cleavage of laminin-5 gamma2 chain via matrix metalloproteinase (mmp)-2 and membrane-type mmp-1. Cancer Res. 2006, 66, 5251–5257. [Google Scholar] [CrossRef]
- Tokes, A.M.; Kulka, J.; Paku, S.; Szik, A.; Paska, C.; Novak, P.K.; Szilak, L.; Kiss, A.; Bogi, K.; Schaff, Z. Claudin-1, -3 and -4 proteins and mrna expression in benign and malignant breast lesions: A research study. Breast Cancer Res. 2005, 7, R296–R305. [Google Scholar] [CrossRef] [PubMed]
- Cohn, M.L.; Goncharuk, V.N.; Diwan, A.H.; Zhang, P.S.; Shen, S.S.; Prieto, V.G. Loss of claudin-1 expression in tumor-associated vessels correlates with acquisition of metastatic phenotype in melanocytic neoplasms. J. Cutan. Pathol. 2005, 32, 533–536. [Google Scholar] [CrossRef]
- Leotlela, P.D.; Wade, M.S.; Duray, P.H.; Rhode, M.J.; Brown, H.F.; Rosenthal, D.T.; Dissanayake, S.K.; Earley, R.; Indig, F.E.; Nickoloff, B.J.; et al. Claudin-1 overexpression in melanoma is regulated by pkc and contributes to melanoma cell motility. Oncogene 2007, 26, 3846–3856. [Google Scholar] [CrossRef]
- Morohashi, S.; Kusumi, T.; Sato, F.; Odagiri, H.; Chiba, H.; Yoshihara, S.; Hakamada, K.; Sasaki, M.; Kijima, H. Decreased expression of claudin-1 correlates with recurrence status in breast cancer. Int. J. Mol. Med. 2007, 20, 139–143. [Google Scholar] [CrossRef]
- Blanchard, A.A.; Skliris, G.P.; Watson, P.H.; Murphy, L.C.; Penner, C.; Tomes, L.; Young, T.L.; Leygue, E.; Myal, Y. Claudins 1, 3, and 4 protein expression in er negative breast cancer correlates with markers of the basal phenotype. Virchows Arch. 2009, 454, 647–656. [Google Scholar] [CrossRef]
- Lu, S.; Singh, K.; Mangray, S.; Tavares, R.; Noble, L.; Resnick, M.B.; Yakirevich, E. Claudin expression in high-grade invasive ductal carcinoma of the breast: Correlation with the molecular subtype. Mod. Pathol. 2013, 26, 485–495. [Google Scholar] [CrossRef]
- Nemeth, J.; Nemeth, Z.; Tatrai, P.; Peter, I.; Somoracz, A.; Szasz, A.M.; Kiss, A.; Schaff, Z. High expression of claudin-1 protein in papillary thyroid tumor and its regional lymph node metastasis. Pathol. Oncol. Res. 2010, 16, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Zwanziger, D.; Badziong, J.; Ting, S.; Moeller, L.; Schmid, K.; Siebolts, U.; Wickenhauser, C.; Dralle, H.; Fuhrer, D. The impact of claudin-1 on follicular thyroid carcinoma aggressiveness. Endocr. Relat. Cancer 2015, 22, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Kinugasa, T.; Akagi, Y.; Yoshida, T.; Ryu, Y.; Shiratuchi, I.; Ishibashi, N.; Shirouzu, K. Increased claudin-1 protein expression contributes to tumorigenesis in ulcerative colitis-associated colorectal cancer. Anticancer Res. 2010, 30, 3181–3186. [Google Scholar] [PubMed]
- Wu, Y.-L.; Zhang, S.; Wang, G.-R.; Chen, Y.-P. Expression transformation of claudin-1 in the process of gastric adenocarcinoma invasion. World J. Gastroenterol. 2008, 14, 4943–4948. [Google Scholar] [CrossRef]
- Nelhubel, G.A.; Karoly, B.; Szabo, B.; Lotz, G.; Kiss, A.; Tovari, J.; Kenessey, I. The prognostic role of claudins in head and neck squamous cell carcinomas. Pathol. Oncol. Res. 2014, 20, 99–106. [Google Scholar] [CrossRef]
- Li, W.J.; Zhang, Z.L.; Yu, X.M.; Cai, X.L.; Pan, X.L.; Yang, X.Y. Expression of claudin-1 and its relationship with lymphatic microvessel generation in hypopharyngeal squamous cell carcinoma. Genet. Mol. Res. 2015, 14, 11814–11826. [Google Scholar] [CrossRef]
- Holczbauer, A.; Gyongyosi, B.; Lotz, G.; Szijarto, A.; Kupcsulik, P.; Schaff, Z.; Kiss, A. Distinct claudin expression profiles of hepatocellular carcinoma and metastatic colorectal and pancreatic carcinomas. J. Histochem. Cytochem. 2013, 61, 294–305. [Google Scholar] [CrossRef]
- Higashi, Y.; Suzuki, S.; Sakaguchi, T.; Nakamura, T.; Baba, S.; Reinecker, H.C.; Nakamura, S.; Konno, H. Loss of claudin-1 expression correlates with malignancy of hepatocellular carcinoma. J. Surg. Res. 2007, 139, 68–76. [Google Scholar] [CrossRef]
- Paschoud, S.; Bongiovanni, M.; Pache, J.-C.; Citi, S. Claudin-1 and claudin-5 expression patterns differentiate lung squamous cell carcinomas from adenocarcinomas. Mod. Pathol. 2007, 20, 947–954. [Google Scholar] [CrossRef]
- Tsukahara, M.; Nagai, H.; Kamiakito, T.; Kawata, H.; Takayashiki, N.; Saito, K.; Tanaka, A. Distinct expression patterns of claudin-1 and claudin-4 in intraductal papillary–mucinous tumors of the pancreas. Pathol. Int. 2005, 55, 63–69. [Google Scholar] [CrossRef]
- English, D.P.; Santin, A.D. Claudins overexpression in ovarian cancer: Potential targets for clostridium perfringens enterotoxin (cpe) based diagnosis and therapy. Int. J. Mol. Sci. 2013, 14, 10412–10437. [Google Scholar] [CrossRef] [PubMed]
- El-Balat, A.; Schmeil, I.; Gasimli, K.; Sänger, N.; Karn, T.; Ahr, A.; Becker, S.; Arsenic, R.; Holtrich, U.; Engels, K. Claudin-1 is linked to presence of implants and micropapillary pattern in serous borderline epithelial tumours of the ovary. J. Clin. Pathol. 2018, 71, 1060. [Google Scholar] [CrossRef]
- Seo, K.W.; Kwon, Y.K.; Kim, B.H.; Kim, C.I.; Chang, H.S.; Choe, M.S.; Park, C.H. Correlation between claudins expression and prognostic factors in prostate cancer. Korean J. Urol. 2010, 51, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Väre, P.; Loikkanen, I.; Hirvikoski, P.; Vaarala, M.; Soini, Y. Low claudin expression is associated with high gleason grade in prostate adenocarcinoma. Oncol. Rep. 2008, 19, 25–31. [Google Scholar] [CrossRef] [PubMed]
- French, A.D.; Fiori, J.L.; Camilli, T.C.; Leotlela, P.D.; O’Connell, M.P.; Frank, B.P.; Subaran, S.; Indig, F.E.; Taub, D.D.; Weeraratna, A.T. Pkc and pka phosphorylation affect the subcellular localization of claudin-1 in melanoma cells. Int. J. Med. Sci. 2009, 6, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.-C.; Pan, S.-H.; Yang, S.-C.; Yu, S.-L.; Che, T.-F.; Lin, C.-W.; Tsai, M.-S.; Chang, G.-C.; Wu, C.-H.; Wu, Y.-Y.; et al. Claudin-1 is a metastasis suppressor and correlates with clinical outcome in lung adenocarcinoma. Am. J. Respir. Crit. Care Med. 2009, 179, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Kleinberg, L.; Holth, A.; Trope, C.G.; Reich, R.; Davidson, B. Claudin upregulation in ovarian carcinoma effusions is associated with poor survival. Hum. Pathol. 2008, 39, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Sharma, A.; Smith, J.J.; Krishnan, M.; Chen, X.; Eschrich, S.; Washington, M.K.; Yeatman, T.J.; Beauchamp, R.D.; Dhawan, P. Claudin-1 up-regulates the repressor zeb-1 to inhibit e-cadherin expression in colon cancer cells. Gastroenterology 2011, 141, 2140–2153. [Google Scholar] [CrossRef]
- Eftang, L.L.; Esbensen, Y.; Tannaes, T.M.; Blom, G.P.; Bukholm, I.R.; Bukholm, G. Up-regulation of cldn1 in gastric cancer is correlated with reduced survival. BMC Cancer 2013, 13, 586. [Google Scholar] [CrossRef]
- Li, W.; Dong, Q.; Li, L.; Zhang, Z.; Cai, X.; Pan, X. Prognostic significance of claudin-1 and cyclin b1 protein expression in patients with hypopharyngeal squamous cell carcinoma. Oncol. Lett. 2016, 11, 2995–3002. [Google Scholar] [CrossRef]
- Suh, Y.; Yoon, C.H.; Kim, R.K.; Lim, E.J.; Oh, Y.S.; Hwang, S.G.; An, S.; Yoon, G.; Gye, M.C.; Yi, J.M.; et al. Claudin-1 induces epithelial–mesenchymal transition through activation of the c-abl-erk signaling pathway in human liver cells. Oncogene 2013, 32, 4873–4882. [Google Scholar] [CrossRef] [PubMed]
- Stebbing, J.; Filipovic, A.; Giamas, G. Claudin-1 as a promoter of emt in hepatocellular carcinoma. Oncogene 2013, 32, 4871–4872. [Google Scholar] [CrossRef] [PubMed]
- Kondo, J.; Sato, F.; Kusumi, T.; Liu, Y.; Motonari, O.; Sato, T.; Kijima, H. Claudin-1 expression is induced by tumor necrosis factor-alpha in human pancreatic cancer cells. Int. J. Mol. Med. 2008, 22, 645–649. [Google Scholar] [PubMed]
- Cronin, K.A.; Lake, A.J.; Scott, S.; Sherman, R.L.; Noone, A.M.; Howlader, N.; Henley, S.J.; Anderson, R.N.; Firth, A.U.; Ma, J.; et al. Annual report to the nation on the status of cancer, part i: National cancer statistics. Cancer 2018, 124, 2785–2800. [Google Scholar] [CrossRef] [PubMed]
- Sinn, H.P.; Kreipe, H. A brief overview of the who classification of breast tumors, 4th edition, focusing on issues and updates from the 3rd edition. Breast Care (Basel) 2013, 8, 149–154. [Google Scholar] [CrossRef]
- Dai, X.; Li, T.; Bai, Z.; Yang, Y.; Liu, X.; Zhan, J.; Shi, B. Breast cancer intrinsic subtype classification, clinical use and future trends. Am. J. Cancer Res. 2015, 5, 2929–2943. [Google Scholar]
- Yeo, S.K.; Guan, J.L. Breast cancer: Multiple subtypes within a tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef]
- Dai, X.; Xiang, L.; Li, T.; Bai, Z. Cancer hallmarks, biomarkers and breast cancer molecular subtypes. J. Cancer 2016, 7, 1281–1294. [Google Scholar] [CrossRef]
- Blanchard, A.; Shiu, R.; Booth, S.; Sorensen, G.; DeCorby, N.; Nistor, A.; Wong, P.; Leygue, E.; Myal, Y. Gene expression profiling of early involuting mammary gland reveals novel genes potentially relevant to human breast cancer. Front. Biosci. 2007, 12, 2221–2232. [Google Scholar] [CrossRef][Green Version]
- Ricardo, S.; Gerhard, R.; Cameselle-Teijeiro, J.F.; Schmitt, F.; Paredes, J. Claudin expression in breast cancer: High or low, what to expect? Histol. Histopathol. 2012, 27, 1283–1295. [Google Scholar]
- Blanchard, A.A.; Zelinski, T.; Xie, J.; Cooper, S.; Penner, C.; Leygue, E.; Myal, Y. Identification of claudin 1 transcript variants in human invasive breast cancer. PLoS ONE 2016, 11, e0163387. [Google Scholar] [CrossRef] [PubMed]
- Swisshelm, K.; Machl, A.; Planitzer, S.; Robertson, R.; Kubbies, M.; Hosier, S. Semp1, a senescence-associated cdna isolated from human mammary epithelial cells, is a member of an epithelial membrane protein superfamily. Gene 1999, 226, 285–295. [Google Scholar] [CrossRef]
- Kramer, F.; White, K.; Kubbies, M.; Swisshelm, K.; Weber, B.H. Genomic organization of claudin-1 and its assessment in hereditary and sporadic breast cancer. Hum. Genet. 2000, 107, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hoevel, T.; Macek, R.; Swisshelm, K.; Kubbies, M. Reexpression of the tj protein cldn1 induces apoptosis in breast tumor spheroids. Int. J. Cancer 2004, 108, 374–383. [Google Scholar] [CrossRef]
- Zhou, B.; Blanchard, A.; Wang, N.; Ma, X.; Han, J.; Schroedter, I.; Leygue, E.; Myal, Y. Claudin 1 promotes migration and increases sensitivity to tamoxifen and anticancer drugs in luminal-like human breast cancer cells mcf7. Cancer Investig. 2015, 429–439. [Google Scholar] [CrossRef]
- Martinez-Estrada, O.M.; Culleres, A.; Soriano, F.X.; Peinado, H.; Bolos, V.; Martinez, F.O.; Reina, M.; Cano, A.; Fabre, M.; Vilaro, S. The transcription factors slug and snail act as repressors of claudin-1 expression in epithelial cells. Biochem. J. 2006, 394, 449–457. [Google Scholar] [CrossRef]
- Myal, Y.; Leygue, E.; Blanchard, A.A. Claudin 1 in breast tumorigenesis: Revelation of a possible novel “claudin high” subset of breast cancers. J. Biomed. Biotechnol. 2010, 2010, 956897. [Google Scholar] [CrossRef]
- Akasaka, H.; Sato, F.; Morohashi, S.; Wu, Y.; Liu, Y.; Kondo, J.; Odagiri, H.; Hakamada, K.; Kijima, H. Anti-apoptotic effect of claudin-1 in tamoxifen-treated human breast cancer mcf-7 cells. BMC Cancer 2010, 10, 548. [Google Scholar] [CrossRef]
- Tanaka, M.; Kamata, R.; Sakai, R. Phosphorylation of ephrin-b1 via the interaction with claudin following cell-cell contact formation. Embo J. 2005, 24, 3700–3711. [Google Scholar] [CrossRef]
- Schmidt, O.; Teis, D. The escrt machinery. Curr. Biol. 2012, 22, R116–R120. [Google Scholar] [CrossRef]
- Kovalenko, O.V.; Yang, X.H.; Hemler, M.E. A novel cysteine cross-linking method reveals a direct association between claudin-1 and tetraspanin cd9. Mol. Cell Proteom. 2007, 6, 1855–1867. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Mannan, P.; Lu, M.; Udey, M.C. Epithelial cell adhesion molecule (epcam) regulates claudin dynamics and tight junctions. J. Biol. Chem. 2013, 288, 12253–12268. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Ezzat, S.; Asa, S.L. Pathogenetic mechanisms in thyroid follicular-cell neoplasia. Nat. Rev. Cancer 2006, 6, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Kitahara, C.M.; Sosa, J.A. The changing incidence of thyroid cancer. Nat. Rev. Endocrinol. 2016, 12, 646–653. [Google Scholar] [CrossRef]
- Hucz, J.; Kowalska, M.; Jarzab, M.; Wiench, M. Gene expression of metalloproteinase 11, claudin 1 and selected adhesion related genes in papillary thyroid cancer. Endokrynol. Pol. 2006, 57, 18–25. [Google Scholar]
- Fluge, O.; Bruland, O.; Akslen, L.A.; Lillehaug, J.R.; Varhaug, J.E. Gene expression in poorly differentiated papillary thyroid carcinomas. Thyroid 2006, 16, 161–175. [Google Scholar] [CrossRef]
- Sobel, G.; Nemeth, J.; Kiss, A.; Lotz, G.; Szabo, I.; Udvarhelyi, N.; Schaff, Z.; Paska, C. Claudin 1 differentiates endometrioid and serous papillary endometrial adenocarcinoma. Gynecol. Oncol. 2006, 103, 591–598. [Google Scholar] [CrossRef]
- Tzelepi, V.N.; Tsamandas, A.C.; Vlotinou, H.D.; Vagianos, C.E.; Scopa, C.D. Tight junctions in thyroid carcinogenesis: Diverse expression of claudin-1, claudin-4, claudin-7 and occludin in thyroid neoplasms. Mod. Pathol. 2008, 21, 22–30. [Google Scholar] [CrossRef]
- Süren, D.; Yildirim, M.; Sayiner, A.; Alikanoğlu, A.S.; Atalay, I.; Gündüz, U.R.; Kaya, V.; Gündüz, Ş.; Oruç, M.T.; Sezer, C. Expression of claudin 1, 4 and 7 in thyroid neoplasms. Oncol. Lett. 2017, 13, 3722–3726. [Google Scholar] [CrossRef]
- Abd El Atti, R.M.; Shash, L.S. Potential diagnostic utility of cd56 and claudin-1 in papillary thyroid carcinoma and solitary follicular thyroid nodules. J. Egypt. Natl. Cancer Inst. 2012, 24, 175–184. [Google Scholar] [CrossRef]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683. [Google Scholar] [CrossRef] [PubMed]
- Ouban, A. Claudin-1 role in colon cancer: An update and a review. Histol. Histopathol. 2018, 11980. [Google Scholar]
- Pope, J.L.; Ahmad, R.; Bhat, A.A.; Washington, M.K.; Singh, A.B.; Dhawan, P. Claudin-1 overexpression in intestinal epithelial cells enhances susceptibility to adenamatous polyposis coli-mediated colon tumorigenesis. Mol. Cancer 2014, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Grone, J.; Weber, B.; Staub, E.; Heinze, M.; Klaman, I.; Pilarsky, C.; Hermann, K.; Castanos-Velez, E.; Ropcke, S.; Mann, B.; et al. Differential expression of genes encoding tight junction proteins in colorectal cancer: Frequent dysregulation of claudin-1, -8 and -12. Int. J. Colorectal Dis. 2007, 22, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Kinugasa, T.; Huo, Q.; Higashi, D.; Shibaguchi, H.; Kuroki, M.; Tanaka, T.; Futami, K.; Yamashita, Y.; Hachimine, K.; Maekawa, S.; et al. Selective up-regulation of claudin-1 and claudin-2 in colorectal cancer. Anticancer Res. 2007, 27, 3729–3734. [Google Scholar] [CrossRef]
- de Oliveira, S.S.; de Oliveira, I.M.; De Souza, W.; Morgado-Diaz, J.A. Claudins upregulation in human colorectal cancer. FEBS Lett. 2005, 579, 6179–6185. [Google Scholar] [CrossRef]
- Nakagawa, S.; Miyoshi, N.; Ishii, H.; Mimori, K.; Tanaka, F.; Sekimoto, M.; Doki, Y.; Mori, M. Expression of cldn1 in colorectal cancer: A novel marker for prognosis. Int. J. Oncol. 2011, 39, 791–796. [Google Scholar]
- Huo, Q.; Kinugasa, T.; Wang, L.; Huang, J.; Zhao, J.; Shibaguchi, H.; Kuroki, M.; Tanaka, T.; Yamashita, Y.; Nabeshima, K.; et al. Claudin-1 protein is a major factor involved in the tumorigenesis of colorectal cancer. Anticancer Res. 2009, 29, 851–857. [Google Scholar]
- Wang, K.; Xu, C.; Li, W.; Ding, L. Emerging clinical significance of claudin-7 in colorectal cancer: A review. Cancer Manag. Res. 2018, 10, 3741–3752. [Google Scholar] [CrossRef]
- Caruso, M.; Fung, K.Y.; Moore, J.; Brierley, G.V.; Cosgrove, L.J.; Thomas, M.; Cheetham, G.; Brook, E.; Fraser, L.M.; Tin, T.; et al. Claudin-1 expression is elevated in colorectal cancer precursor lesions harboring the braf v600e mutation. Transl. Oncol. 2014, 7, 456–463. [Google Scholar] [CrossRef]
- Schatoff, E.M.; Leach, B.I.; Dow, L.E. Wnt signaling and colorectal cancer. Curr. Colorectal Cancer Rep. 2017, 13, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Multilayered prevention and treatment of chronic inflammation, organ fibrosis and cancer associated with canonical wnt/betacatenin signaling activation (review). Int. J. Mol. Med. 2018, 713–725. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Kurzrock, R. Targeting the wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N.; Furuse, M.; Tsukita, S.; Niikawa, N.; Nakamura, Y.; Furukawa, Y. Involvement of claudin-1 in the beta-catenin/tcf signaling pathway and its frequent upregulation in human colorectal cancers. Oncol. Res. 2001, 12, 469–476. [Google Scholar] [CrossRef]
- Bhat, A.A.; Sharma, A.; Pope, J.; Krishnan, M.; Washington, M.K.; Singh, A.B.; Dhawan, P. Caudal homeobox protein cdx-2 cooperates with wnt pathway to regulate claudin-1 expression in colon cancer cells. PLoS ONE 2012, 7, e37174. [Google Scholar] [CrossRef]
- Cherradi, S.; Ayrolles-Torro, A.; Vezzo-Vié, N.; Gueguinou, N.; Denis, V.; Combes, E.; Boissière, F.; Busson, M.; Canterel-Thouennon, L.; Mollevi, C.; et al. Antibody targeting of claudin-1 as a potential colorectal cancer therapy. J. Exp. Clin. Cancer Res. 2017, 36, 89. [Google Scholar] [CrossRef]
- Jung, H.; Jun, K.H.; Jung, J.H.; Chin, H.M.; Park, W.B. The expression of claudin-1, claudin-2, claudin-3, and claudin-4 in gastric cancer tissue. J. Surg. Res. 2011, 167, e185–e191. [Google Scholar] [CrossRef]
- Huang, J.; Li, J.; Qu, Y.; Zhang, J.; Zhang, L.; Chen, X.; Liu, B.; Zhu, Z. The expression of claudin 1 correlates with beta-catenin and is a prognostic factor of poor outcome in gastric cancer. Int. J. Oncol. 2014, 44, 1293–1301. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, L.; He, C.; Qu, Y.; Li, J.; Zhang, J.; Du, T.; Chen, X.; Yu, Y.; Liu, B.; et al. Claudin-1 enhances tumor proliferation and metastasis by regulating cell anoikis in gastric cancer. Oncotarget 2015, 6, 1652–1665. [Google Scholar] [CrossRef]
- Tokuhara, Y.; Morinishi, T.; Matsunaga, T.; Ohsaki, H.; Kushida, Y.; Haba, R.; Hirakawa, E. Claudin-1, but not claudin-4, exhibits differential expression patterns between well- to moderately-differentiated and poorly-differentiated gastric adenocarcinoma. Oncol. Lett. 2015, 10, 93–98. [Google Scholar] [CrossRef]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Ouban, A.; Hamdan, H.; Hakam, A.; Ahmed, A.A. Claudin-1 expression in squamous cell carcinomas of different organs: Comparative study of cancerous tissues and normal controls. Int. J. Surg. Pathol. 2012, 20, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Ghouri, Y.A.; Mian, I.; Rowe, J.H. Review of hepatocellular carcinoma: Epidemiology, etiology, and carcinogenesis. J. Carcinog. 2017, 16, 1–18. [Google Scholar] [PubMed]
- Yoon, C.-H.; Kim, M.-J.; Park, M.-J.; Park, I.-C.; Hwang, S.-G.; An, S.; Choi, Y.-H.; Yoon, G.; Lee, S.-J. Claudin-1 acts through c-abl-protein kinase cdelta (pkcdelta) signaling and has a causal role in the acquisition of invasive capacity in human liver cells. J. Biol. Chem. 2010, 285, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; You, M.-L.; Chong, Q.-Y.; Pandey, V.; Zhuang, Q.-S.; Liu, D.-X.; Ma, L.; Zhu, T.; Lobie, P.E. Autocrine human growth hormone promotes invasive and cancer stem cell-like behavior of hepatocellular carcinoma cells by stat3 dependent inhibition of claudin-1 expression. Int. J. Mol. Sci. 2017, 18, 1274. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Jung, C.K.; Choi, H.J.; Jun, K.H.; Yoo, J.; Kang, S.J.; Lee, K.Y. Diagnostic utility of expression of claudins in non-small cell lung cancer: Different expression profiles in squamous cell carcinomas and adenocarcinomas. Pathol. Res. Pract. 2009, 205, 409–416. [Google Scholar] [CrossRef]
- Soini, Y. Claudins in lung diseases. Respir. Res. 2011, 12, 70. [Google Scholar] [CrossRef]
- Sun, B.-S.; Yao, Y.-Q.; Pei, B.-X.; Zhang, Z.-F.; Wang, C.-L. Claudin-1 correlates with poor prognosis in lung adenocarcinoma. Thorac. Cancer 2016, 7, 556–563. [Google Scholar] [CrossRef]
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014, 149185. [Google Scholar] [CrossRef]
- Shiozaki, A.; Bai, X.H.; Shen-Tu, G.; Moodley, S.; Takeshita, H.; Fung, S.Y.; Wang, Y.; Keshavjee, S.; Liu, M. Claudin 1 mediates tnfalpha-induced gene expression and cell migration in human lung carcinoma cells. PLoS ONE 2012, 7, e38049. [Google Scholar] [CrossRef]
- Bhat, A.A.; Ahmad, R.; Uppada, S.B.; Singh, A.B.; Dhawan, P. Claudin-1 promotes tnf-α-induced epithelial-mesenchymal transition and migration in colorectal adenocarcinoma cells. Exp. Cell Res. 2016, 349, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Cruet-Hennequart, S.; Troussard, A.; Fazli, L.; Costello, P.; Sutton, K.; Wheeler, J.; Gleave, M.; Sanghera, J.; Dedhar, S. Regulation of tumor angiogenesis by integrin-linked kinase (ilk). Cancer Cell 2004, 5, 79–90. [Google Scholar] [CrossRef]
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9–32. [Google Scholar] [PubMed]
- Liu, T.; Cheng, W.; Lai, D.; Huang, Y.; Guo, L. Characterization of primary ovarian cancer cells in different culture systems. Oncol. Rep. 2010, 23, 1277–1284. [Google Scholar] [PubMed]
- Zhu, Y.; Sundfeldt, K. Tight junction formation in epithelial ovarian adenocarcinoma. Acta Obs. Gynecol. Scand. 2007, 86, 1011–1019. [Google Scholar] [CrossRef]
- Tandon, P.; Dadhich, A.; Saluja, H.; Bawane, S.; Sachdeva, S. The prevalence of squamous cell carcinoma in different sites of oral cavity at our rural health care centre in loni, maharashtra—A retrospective 10-year study. Contemp. Oncol. 2017, 21, 178–183. [Google Scholar] [CrossRef]
- Habelhah, H.; Okada, F.; Kobayashi, M.; Nakai, K.; Choi, S.; Hamada, J.; Moriuchi, T.; Kaya, M.; Yoshida, K.; Fujinaga, K.; et al. Increased e1af expression in mouse fibrosarcoma promotes metastasis through induction of mt1-mmp expression. Oncogene 1999, 18, 1771–1776. [Google Scholar] [CrossRef][Green Version]
- Sappayatosok, K.; Phattarataratip, E. Overexpression of claudin-1 is associated with advanced clinical stage and invasive pathologic characteristics of oral squamous cell carcinoma. Head Neck Pathol. 2015, 9, 173–180. [Google Scholar] [CrossRef]
- Lourenco, S.V.; Coutinho-Camillo, C.M.; Buim, M.E.; Pereira, C.M.; Carvalho, A.L.; Kowalski, L.P.; Soares, F.A. Oral squamous cell carcinoma: Status of tight junction claudins in the different histopathological patterns and relationship with clinical parameters. A tissue-microarray-based study of 136 cases. J. Clin. Pathol. 2010, 63, 609–614. [Google Scholar] [CrossRef]
- De Vicente, J.C.; Fernández-Valle, Á.; Vivanco-Allende, B.; Santamarta, T.R.; Lequerica-Fernández, P.; Hernández-Vallejo, G.; Allonca-Campa, E. The prognostic role of claudins -1 and -4 in oral squamous cell carcinoma. Anticancer Res. 2015, 35, 2949–2959. [Google Scholar] [PubMed]
- Jerant, A.F.; Johnson, J.T.; Sheridan, C.D.; Caffrey, T.J. Early detection and treatment of skin cancer. Am. Fam. Physician 2000, 62, 357–368, 375–376, 381–382. [Google Scholar]
- Weeraratna, A.T.; Becker, D.; Carr, K.M.; Duray, P.H.; Rosenblatt, K.P.; Yang, S.; Chen, Y.; Bittner, M.; Strausberg, R.L.; Riggins, G.J.; et al. Generation and analysis of melanoma sage libraries: Sage advice on the melanoma transcriptome. Oncogene 2004, 23, 2264–2274. [Google Scholar] [CrossRef] [PubMed]
- Izraely, S.; Sagi-Assif, O.; Klein, A.; Meshel, T.; Ben-Menachem, S.; Zaritsky, A.; Ehrlich, M.; Prieto, V.G.; Bar-Eli, M.; Pirker, C.; et al. The metastatic microenvironment: Claudin-1 suppresses the malignant phenotype of melanoma brain metastasis. Int. J. Cancer 2015, 136, 1296–1307. [Google Scholar] [CrossRef]
- Wong, M.C.S.; Goggins, W.B.; Wang, H.H.X.; Fung, F.D.H.; Leung, C.; Wong, S.Y.S.; Ng, C.F.; Sung, J.J.Y. Global incidence and mortality for prostate cancer: Analysis of temporal patterns and trends in 36 countries. Eur. Urol. 2016, 70, 862–874. [Google Scholar] [CrossRef]
- Sugawara, T.; Iwamoto, N.; Akashi, M.; Kojima, T.; Hisatsune, J.; Sugai, M.; Furuse, M. Tight junction dysfunction in the stratum granulosum leads to aberrant stratum corneum barrier function in claudin-1-deficient mice. J. Dermatol. Sci. 2013, 70, 12–18. [Google Scholar] [CrossRef]
- Inai, T.; Kobayashi, J.; Shibata, Y. Claudin-1 contributes to the epithelial barrier function in mdck cells. Eur. J. Cell Biol. 1999, 78, 849–855. [Google Scholar] [CrossRef]
- McCarthy, K.M.; Francis, S.A.; McCormack, J.M.; Lai, J.; Rogers, R.A.; Skare, I.B.; Lynch, R.D.; Schneeberger, E.E. Inducible expression of claudin-1-myc but not occludin-vsv-g results in aberrant tight junction strand formation in mdck cells. J. Cell Sci. 2000, 113, 3387. [Google Scholar]
- Grosse, B.; Cassio, D.; Yousef, N.; Bernardo, C.; Jacquemin, E.; Gonzales, E. Claudin-1 involved in neonatal ichthyosis sclerosing cholangitis syndrome regulates hepatic paracellular permeability. Hepatology 2012, 55, 1249–1259. [Google Scholar] [CrossRef]
- Poritz, L.S.; Harris, L.R., 3rd; Kelly, A.A.; Koltun, W.A. Increase in the tight junction protein claudin-1 in intestinal inflammation. Dig. Dis. Sci. 2011, 56, 2802–2809. [Google Scholar] [CrossRef]
- Sladojevic, N.; Stamatovic, S.M.; Johnson, A.M.; Choi, J.; Hu, A.; Dithmer, S.; Blasig, I.E.; Keep, R.F.; Andjelkovic, A.V. Claudin-1-dependent destabilization of the blood–brain barrier in chronic stroke. J. Neurosci. 2019, 39, 743. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Fukasawa, M.; Kuniyasu, H.; Yagi, K.; Kondoh, M. Claudin-targeted drug development using anti-claudin monoclonal antibodies to treat hepatitis and cancer. Ann. N. Y. Acad. Sci. 2017, 1397, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Tada, M.; Iida, M.; Nagase, S.; Hata, T.; Watari, A.; Okada, Y.; Doi, T.; Fukasawa, M.; Yagi, K.; et al. Generation and characterization of a human–mouse chimeric antibody against the extracellular domain of claudin-1 for cancer therapy using a mouse model. Biochem. Biophys. Res. Commun. 2016, 477, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Nordin, B.E.; Robertson, A.; Seamark, R.F.; Bridges, A.; Philcox, J.C.; Need, A.G.; Horowitz, M.; Morris, H.A.; Deam, S. The relation between calcium absorption, serum dehydroepiandrosterone, and vertebral mineral density in postmenopausal women. J. Clin. Endocrinol. Metab. 1985, 60, 651–657. [Google Scholar] [CrossRef]
- Fukasawa, M.; Nagase, S.; Shirasago, Y.; Iida, M.; Yamashita, M.; Endo, K.; Yagi, K.; Suzuki, T.; Wakita, T.; Hanada, K.; et al. Monoclonal antibodies against extracellular domains of claudin-1 block hepatitis c virus infection in a mouse model. J. Virol. 2015, 89, 4866–4879. [Google Scholar] [CrossRef] [PubMed]
- Hollandsworth, H.M.; Lwin, T.M.; Amirfakhri, S.; Filemoni, F.; Batra, S.K.; Hoffman, R.M.; Dhawan, P.; Bouvet, M. Anti-claudin-1 conjugated to a near-infrared fluorophore targets colon cancer in pdox mouse models. J. Surg. Res. 2019, 242, 145–150. [Google Scholar] [CrossRef]
- Kominsky, S.L. Claudins: Emerging targets for cancer therapy. Expert Rev. Mol. Med. 2006, 8, 1–11. [Google Scholar] [CrossRef]
- Saeki, R.; Kondoh, M.; Kakutani, H.; Tsunoda, S.-i.; Mochizuki, Y.; Hamakubo, T.; Tsutsumi, Y.; Horiguchi, Y.; Yagi, K. A novel tumor-targeted therapy using a claudin-4-targeting molecule. Mol. Pharmacol. 2009, 76, 918. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Kawahigashi, Y.; Hata, T.; Li, X.; Watari, A.; Tada, M.; Ishii-Watabe, A.; Okada, Y.; Doi, T.; Fukasawa, M.; et al. Efficacy and safety evaluation of claudin-4-targeted antitumor therapy using a human and mouse cross-reactive monoclonal antibody. Pharmacol. Res. Perspect. 2016, 4, e00266. [Google Scholar] [CrossRef]
- Torres, J.B.; Knight, J.C.; Mosley, M.J.; Kersemans, V.; Koustoulidou, S.; Allen, D.; Kinchesh, P.; Smart, S.; Cornelissen, B. Imaging of claudin-4 in pancreatic ductal adenocarcinoma using a radiolabelled anti-claudin-4 monoclonal antibody. Mol. Imaging Biol. 2018, 20, 292–299. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Tachibana, K.; Krug, S.M.; Kunisawa, J.; Fromm, M.; Kondoh, M. Potential for tight junction protein-directed drug development using claudin binders and angubindin-1. Int. J. Mol. Sci. 2019, 20, 4016. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.B.; Lu, Y.; Choate, K.A.; Velazquez, H.; Al-Sabban, E.; Praga, M.; Casari, G.; Bettinelli, A.; Colussi, G.; Rodriguez-Soriano, J.; et al. Paracellin-1, a renal tight junction protein required for paracellular Mg2+ resorption. Science 1999, 285, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Sanjad, S.A.; Hariri, A.; Habbal, Z.M.; Lifton, R.P. A novel pcln-1 gene mutation in familial hypomagnesemia with hypercalciuria and atypical phenotype. Pediatr. Nephrol. 2007, 22, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Kobayashi, N.; Itoh, T.; Takasuga, A.; Nakamaru, T.; Hirotsune, S.; Sugimoto, Y. Null mutation of pcln-1/claudin-16 results in bovine chronic interstitial nephritis. Genome Res. 2000, 10, 659–663. [Google Scholar] [CrossRef]
- Hadj-Rabia, S.; Baala, L.; Vabres, P.; Hamel-Teillac, D.; Jacquemin, E.; Fabre, M.; Lyonnet, S.; de Prost, Y.; Munnich, A.; Hadchouel, M.; et al. Claudin-1 gene mutations in neonatal sclerosing cholangitis associated with ichthyosis: A tight junction disease. Gastroenterology 2004, 127, 1386–1390. [Google Scholar] [CrossRef]
- Sineni, C.J.; Yildirim-Baylan, M.; Guo, S.; Camarena, V.; Wang, G.; Tokgoz-Yilmaz, S.; Duman, D.; Bademci, G.; Tekin, M. A truncating cldn9 variant is associated with autosomal recessive nonsyndromic hearing loss. Hum. Genet. 2019, 138, 1071–1075. [Google Scholar] [CrossRef]
- Hadj-Rabia, S.; Brideau, G.; Al-Sarraj, Y.; Maroun, R.C.; Figueres, M.-L.; Leclerc-Mercier, S.; Olinger, E.; Baron, S.; Chaussain, C.; Nochy, D.; et al. Multiplex epithelium dysfunction due to cldn10 mutation: The helix syndrome. Genet. Med. 2018, 20, 190–201. [Google Scholar] [CrossRef]
- Wilcox, E.R.; Burton, Q.L.; Naz, S.; Riazuddin, S.; Smith, T.N.; Ploplis, B.; Belyantseva, I.; Ben-Yosef, T.; Liburd, N.A.; Morell, R.J.; et al. Mutations in the gene encoding tight junction claudin-14 cause autosomal recessive deafness dfnb29. Cell 2001, 104, 165–172. [Google Scholar] [CrossRef]
- Hampson, G.; Konrad, M.A.; Scoble, J. Familial hypomagnesaemia with hypercalciuria and nephrocalcinosis (fhhnc): Compound heterozygous mutation in the claudin 16 (cldn16) gene. BMC Nephrol. 2008, 9, 12. [Google Scholar] [CrossRef]
- Konrad, M.; Schaller, A.; Seelow, D.; Pandey, A.V.; Waldegger, S.; Lesslauer, A.; Vitzthum, H.; Suzuki, Y.; Luk, J.M.; Becker, C.; et al. Mutations in the tight-junction gene claudin 19 (cldn19) are associated with renal magnesium wasting, renal failure, and severe ocular involvement. Am. J. Hum. Genet. 2006, 79, 949–957. [Google Scholar] [CrossRef]
- Seker, M.; Fernandez-Rodriguez, C.; Martinez-Cruz, L.A.; Müller, D. Mouse models of human claudin-associated disorders: Benefits and limitations. Int. J. Mol. Sci. 2019, 20, 5504. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Subtypes | Expression of CLDN-1 | References |
|---|---|---|---|
| Breast Cancer | Luminal A | Downregulated | [40] |
| Luminal B | Downregulated | [40] | |
| Triple negative/Basal like | Upregulated | [40] | |
| HER2 enriched | Downregulated | [41] | |
| Claudin-low | Downregulated | [41] | |
| Thyroid Cancer | Papillary Thyroid Cancer | Upregulated | [42] |
| Follicular Thyroid Cancer | Upregulated | [43] | |
| Colorectal Cancer | Ulcerative Colitis associated Colorectal Cancer | Upregulated | [44] |
| Sporadic Colorectal Cancer | Upregulated | [34] | |
| Gastric Adenocarcinoma | - | Upregulated | [45] |
| Head and Neck Squamous Cell Carcinoma | - | Upregulated | [46] |
| Hypopharyngeal Squamous Cell Carcinoma | - | Upregulated | [47] |
| Hepatocellular Carcinoma | - | Downregulated | [48,49] |
| Lung Adenocarcinoma | - | Downregulated | [50] |
| Pancreatic Ductal Carcinoma | - | Upregulated | [51] |
| Epithelial Ovarian Cancer | - | Upregulated | [52,53] |
| Oral Squamous Cell Carcinoma | - | Upregulated | [35] |
| Melanoma | - | Upregulated | [38] |
| Prostate adenocarcinoma | - | Downregulated | [54,55] |
| Cancer Type | Activity | Findings | References |
|---|---|---|---|
| Melanoma | Tumor Promoter | Cytoplasmic expression of CLDN-1 contributes to the migratory capacity of melanoma cells | [56] |
| Oral Squamous Cell Carcinoma | Tumor Promoter | CLDN-1 enhances the invasive activity of OSC-4 and NOS-2 cell lines by activation of MT1-MMP and MMP-2 | [35] |
| Prostate Cancer | Tumor Suppressor | Loss of CLDN-1 associated with progression of Prostate cancer | [54] |
| Lung Cancer | Tumor Suppressor | Knockdown of CLDN-1 increased invasive and metastatic activity of lung adenocarcinoma cells | [57] |
| Breast Cancer | Tumor Promoter in ER-Subtypes | Increases cell migration and also exhibits an anti-apoptotic effect | [40] |
| Tumor Suppressor in ER+ Subtypes | Acts as a suppressor of mammary epithelial proliferation Increases apoptosis of breast cancer cells | ||
| Thyroid Cancer | Tumor Promoter | High expression of CLDN-1 found in follicular thyroid carcinoma (FTC-133) and Papillary Thyroid Carcinoma cells | [42,43] |
| Ovarian Cancer | Tumor Promoter | High expression of CLDN-1 correlated with shorter overall survival in ovarian carcinoma effusions | [58] |
| Colon Cancer | Tumor Promoter | High CLDN-1 expression in colon carcinoma and metastasis CLDN-1 upregulates the repressor ZEB-1 to reduce expression of E-cadherin in colon cancer cells | [34,59] |
| Gastric Cancer | Tumor Promoter | High expression of CLDN-1 in gastric cancer associated with poor survival | [60] |
| Hypopharyngeal Squamous Cell Carcinoma | Tumor Promoter | High expression of CLDN-1 associated with lymph node metastasis and degree of tumor differentiation | [61] |
| Hepatocellular Carcinoma | Tumor Promoter | CLDN-1 promoted epithelial-mesenchymal transition (EMT) in HCC cells by overexpression of mesenchymal markers (N-cadherin and vimentin) | [48,62,63] |
| Pancreatic Cancer | Tumor Promoter | TNF-α upregulated CLDN-1 expression, leading to increased proliferation of pancreatic cancer cells | [64] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhat, A.A.; Syed, N.; Therachiyil, L.; Nisar, S.; Hashem, S.; Macha, M.A.; Yadav, S.K.; Krishnankutty, R.; Muralitharan, S.; Al-Naemi, H.; et al. Claudin-1, A Double-Edged Sword in Cancer. Int. J. Mol. Sci. 2020, 21, 569. https://doi.org/10.3390/ijms21020569
Bhat AA, Syed N, Therachiyil L, Nisar S, Hashem S, Macha MA, Yadav SK, Krishnankutty R, Muralitharan S, Al-Naemi H, et al. Claudin-1, A Double-Edged Sword in Cancer. International Journal of Molecular Sciences. 2020; 21(2):569. https://doi.org/10.3390/ijms21020569
Chicago/Turabian StyleBhat, Ajaz A., Najeeb Syed, Lubna Therachiyil, Sabah Nisar, Sheema Hashem, Muzafar A. Macha, Santosh K. Yadav, Roopesh Krishnankutty, Shanmugakonar Muralitharan, Hamda Al-Naemi, and et al. 2020. "Claudin-1, A Double-Edged Sword in Cancer" International Journal of Molecular Sciences 21, no. 2: 569. https://doi.org/10.3390/ijms21020569
APA StyleBhat, A. A., Syed, N., Therachiyil, L., Nisar, S., Hashem, S., Macha, M. A., Yadav, S. K., Krishnankutty, R., Muralitharan, S., Al-Naemi, H., Bagga, P., Reddy, R., Dhawan, P., Akobeng, A., Uddin, S., Frenneaux, M. P., El-Rifai, W., & Haris, M. (2020). Claudin-1, A Double-Edged Sword in Cancer. International Journal of Molecular Sciences, 21(2), 569. https://doi.org/10.3390/ijms21020569