Biological Functions of HMGN Chromosomal Proteins



Abstract
1. Introduction
2. Genetically Altered HMGN Mice
3. HMGN Proteins in Development
3.1. Embryogenesis
3.2. Neuronal Development
4. Ocular Development
5. Reproductive System
6. Pancreas
7. HMGN Proteins in Disease
8. Cancer
9. Neurological Disorders
10. Immune Functions
11. Perspective: Biological Function of the HMGN Protein Family
Funding
Acknowledgments
Conflicts of Interest
References
- Hock, R.; Furusawa, T.; Ueda, T.; Bustin, M. HMG chromosomal proteins in development and disease. Trends Cell Biol. 2007, 17, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Romero, R.; Eirin-Lopez, J.M.; Ausio, J. Evolution of high mobility group nucleosome-binding proteins and its implications for vertebrate chromatin specialization. Mol. Biol. Evol. 2015, 32, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Catez, F.; Lim, J.H.; Hock, R.; Postnikov, Y.V.; Bustin, M. HMGN dynamics and chromatin function. Biochem. Cell Biol. 2003, 81, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, H.; Herrera, J.E.; Bustin, M.; Postnikov, Y. Targeting of high mobility group-14/-17 proteins in chromatin is independent of DNA sequence. J. Biol. Chem. 2000, 275, 37937–37944. [Google Scholar] [CrossRef]
- Postnikov, Y.; Bustin, M. Regulation of chromatin structure and function by HMGN proteins. Biochim. Biophys. Acta 2010, 1799, 62–68. [Google Scholar] [CrossRef]
- Catez, F.; Brown, D.T.; Misteli, T.; Bustin, M. Competition between histone H1 and HMGN proteins for chromatin binding sites. EMBO Rep. 2002, 3, 760–766. [Google Scholar] [CrossRef]
- Rochman, M.; Postnikov, Y.; Correll, S.; Malicet, C.; Wincovitch, S.; Karpova, T.S.; McNally, J.G.; Wu, X.; Bubunenko, N.A.; Grigoryev, S.; et al. The interaction of NSBP1/HMGN5 with nucleosomes in euchromatin counteracts linker histone-mediated chromatin compaction and modulates transcription. Mol. Cell 2009, 35, 642–656. [Google Scholar] [CrossRef]
- Catez, F.; Ueda, T.; Bustin, M. Determinants of histone H1 mobility and chromatin binding in living cells. Nat. Struct. Mol. Biol. 2006, 13, 305–310. [Google Scholar] [CrossRef]
- Postnikov, Y.V.; Bustin, M. Functional interplay between histone H1 and HMG proteins in chromatin. Biochim. Biophys. Acta 2016, 1859, 462–467. [Google Scholar] [CrossRef]
- Deng, T.; Zhu, Z.I.; Zhang, S.; Postnikov, Y.; Huang, D.; Horsch, M.; Furusawa, T.; Beckers, J.; Rozman, J.; Klingenspor, M.; et al. Functional compensation among HMGN variants modulates the DNase I hypersensitive sites at enhancers. Genome Res. 2015, 25, 1295–1308. [Google Scholar] [CrossRef]
- He, B.; Deng, T.; Zhu, I.; Furusawa, T.; Zhang, S.; Tang, W.; Postnikov, Y.; Ambs, S.; Li, C.C.; Livak, F.; et al. Binding of HMGN proteins to cell specific enhancers stabilizes cell identity. Nat. Commun. 2018, 9, 5240. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, H.; Rochman, M.; Furusawa, T.; Kuehn, M.R.; Horigome, S.; Haketa, K.; Sugita, Y.; Inada, T.; Komai, M.; Bustin, M. The nucleosomal binding protein NSBP1 is highly expressed in the placenta and modulates the expression of differentiation markers in placental Rcho-1 cells. J. Cell. Biochem. 2009, 106, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Furusawa, T.; Kurahashi, T.; Tessarollo, L.; Bustin, M. The nucleosome binding protein HMGN3 modulates the transcription profile of pancreatic beta cells and affects insulin secretion. Mol. Cell. Biol. 2009, 29, 5264–5276. [Google Scholar] [CrossRef] [PubMed]
- Kugler, J.E.; Deng, T.; Bustin, M. The HMGN family of chromatin-binding proteins: Dynamic modulators of epigenetic processes. Biochim. Biophys. Acta 2012, 1819, 652–656. [Google Scholar] [CrossRef]
- Landsman, D.; McBride, O.W.; Soares, N.; Crippa, M.P.; Srikantha, T.; Bustin, M. Chromosomal protein HMG-14. Identification, characterization, and chromosome localization of a functional gene from the large human multigene family. J. Biol. Chem. 1989, 264, 3421–3427. [Google Scholar]
- Landsman, D.; McBride, O.W.; Bustin, M. Human non-histone chromosomal protein HMG-17: Identification, characterization, chromosome localization and RFLPs of a functional gene from the large multigene family. Nucleic Acids Res. 1989, 17, 2301–2314. [Google Scholar] [CrossRef]
- Furusawa, T.; Cherukuri, S. Developmental function of HMGN proteins. Biochim. Biophys. Acta 2010, 1799, 69–73. [Google Scholar] [CrossRef]
- Kugler, J.E.; Horsch, M.; Huang, D.; Furusawa, T.; Rochman, M.; Garrett, L.; Becker, L.; Bohla, A.; Holter, S.M.; Prehn, C.; et al. High mobility group N proteins modulate the fidelity of the cellular transcriptional profile in a tissue-and variant-specific manner. J. Biol. Chem. 2013, 288, 16690–16703. [Google Scholar] [CrossRef]
- Gao, X.L.; Tian, W.J.; Liu, B.; Wu, J.; Xie, W.; Shen, Q. High mobility group nucleosomal binding domain 2 protects against microcephaly by maintaining global chromatin accessibility during corticogenesis. J. Biol. Chem. 2019. [Google Scholar] [CrossRef]
- Deng, T.; Postnikov, Y.; Zhang, S.; Garrett, L.; Becker, L.; Racz, I.; Holter, S.M.; Wurst, W.; Fuchs, H.; Gailus-Durner, V.; et al. Interplay between H1 and HMGN epigenetically regulates OLIG1&2 expression and oligodendrocyte differentiation. Nucleic Acids Res. 2017, 45, 3031–3045. [Google Scholar]
- Mohamed, O.A.; Bustin, M.; Clarke, H.J. High-mobility group proteins 14 and 17 maintain the timing of early embryonic development in the mouse. Dev. Biol. 2001, 229, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, T.; Lim, J.H.; Catez, F.; Birger, Y.; Mackem, S.; Bustin, M. Down-regulation of nucleosomal binding protein HMGN1 expression during embryogenesis modulates Sox9 expression in chondrocytes. Mol. Cell. Biol. 2006, 26, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, S.; Lehtonen, E. HMG-17 is an early marker of inductive interactions in the developing mouse kidney. Differentiation 2001, 67, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, T.; Ko, J.H.; Birger, Y.; Bustin, M. Expression of nucleosomal protein HMGN1 in the cycling mouse hair follicle. Gene Expr. Patterns 2009, 9, 289–295. [Google Scholar] [CrossRef]
- Deng, T.; Zhu, Z.I.; Zhang, S.; Leng, F.; Cherukuri, S.; Hansen, L.; Marino-Ramirez, L.; Meshorer, E.; Landsman, D.; Bustin, M. HMGN1 modulates nucleosome occupancy and DNase I hypersensitivity at the CpG island promoters of embryonic stem cells. Mol. Cell. Biol. 2013, 33, 3377–3389. [Google Scholar] [CrossRef]
- Garza-Manero, S.; Sindi, A.A.A.; Mohan, G.; Rehbini, O.; Jeantet, V.H.M.; Bailo, M.; Latif, F.A.; West, M.P.; Gurden, R.; Finlayson, L.; et al. Maintenance of active chromatin states by HMGN2 is required for stem cell identity in a pluripotent stem cell model. Epigenetics Chromatin 2019, 12, 73. [Google Scholar] [CrossRef]
- Birger, Y.; West, K.L.; Postnikov, Y.V.; Lim, J.H.; Furusawa, T.; Wagner, J.P.; Laufer, C.S.; Kraemer, K.H.; Bustin, M. Chromosomal protein HMGN1 enhances the rate of DNA repair in chromatin. EMBO J. 2003, 22, 1665–1675. [Google Scholar] [CrossRef]
- Birger, Y.; Davis, J.; Furusawa, T.; Rand, E.; Piatigorsky, J.; Bustin, M. A role for chromosomal protein HMGN1 in corneal maturation. Differentiation 2006, 74, 19–29. [Google Scholar] [CrossRef]
- Korner, U.; Bustin, M.; Scheer, U.; Hock, R. Developmental role of HMGN proteins in Xenopus laevis. Mech. Dev. 2003, 120, 1177–1192. [Google Scholar] [CrossRef]
- Pash, J.M.; Bhorjee, J.S.; Patterson, B.M.; Bustin, M. Persistence of chromosomal proteins HMG-14/-17 in myotubes following differentiation-dependent reduction of HMG mRNA. J. Biol. Chem. 1990, 265, 4197–4199. [Google Scholar]
- Crippa, M.P.; Nickol, J.M.; Bustin, M. Developmental changes in the expression of high mobility group chromosomal proteins. J. Biol. Chem. 1991, 266, 2712–2714. [Google Scholar]
- Bustin, M.; Misteli, T. Nongenetic functions of the genome. Science 2016, 352, aad6933. [Google Scholar] [CrossRef]
- Furusawa, T.; Rochman, M.; Taher, L.; Dimitriadis, E.K.; Nagashima, K.; Anderson, S.; Bustin, M. Chromatin decompaction by the nucleosomal binding protein HMGN5 impairs nuclear sturdiness. Nat. Commun. 2015, 6, 6138. [Google Scholar] [CrossRef]
- Ciappio, E.D.; Krausz, K.W.; Rochman, M.; Furusawa, T.; Bonzo, J.A.; Tessarollo, L.; Gonzalez, F.J.; Bustin, M. Metabolomics reveals a role for the chromatin-binding protein HMGN5 in glutathione metabolism. PLoS ONE 2014, 9, e84583. [Google Scholar] [CrossRef]
- Nagao, M.; Lanjakornsiripan, D.; Itoh, Y.; Kishi, Y.; Ogata, T.; Gotoh, Y. High mobility group nucleosome-binding family proteins promote astrocyte differentiation of neural precursor cells. Stem Cells 2014, 32, 2983–2997. [Google Scholar] [CrossRef]
- Ito, Y.; Bustin, M. Immunohistochemical localization of the nucleosome-binding protein HMGN3 in mouse brain. J. Histochem. Cytochem. 2002, 50, 1273–1275. [Google Scholar] [CrossRef][Green Version]
- West, K.L.; Castellini, M.A.; Duncan, M.K.; Bustin, M. Chromosomal proteins HMGN3a and HMGN3b regulate the expression of glycine transporter 1. Mol. Cell. Biol. 2004, 24, 3747–3756. [Google Scholar] [CrossRef]
- Barkess, G.; Postnikov, Y.; Campos, C.D.; Mishra, S.; Mohan, G.; Verma, S.; Bustin, M.; West, K.L. The chromatin-binding protein HMGN3 stimulates histone acetylation and transcription across the Glyt1 gene. Biochem. J. 2012, 442, 495–505. [Google Scholar] [CrossRef]
- Moretti, F.; Rolando, C.; Winker, M.; Ivanek, R.; Rodriguez, J.; Von Kriegsheim, A.; Taylor, V.; Bustin, M.; Pertz, O. Growth Cone Localization of the mRNA Encoding the Chromatin Regulator HMGN5 Modulates Neurite Outgrowth. Mol. Cell. Biol. 2015, 35, 2035–2050. [Google Scholar] [CrossRef]
- Pash, J.; Popescu, N.; Matocha, M.; Rapoport, S.; Bustin, M. Chromosomal protein HMG-14 gene maps to the Down syndrome region of human chromosome 21 and is overexpressed in mouse trisomy 16. Proc. Natl. Acad. Sci. USA 1990, 87, 3836–3840. [Google Scholar] [CrossRef]
- Pash, J.; Smithgall, T.; Bustin, M. Chromosomal protein HMG-14 is overexpressed in Down syndrome. Exp. Cell Res. 1991, 193, 232–235. [Google Scholar] [CrossRef]
- Potier, M.C.; Rivals, I.; Mercier, G.; Ettwiller, L.; Moldrich, R.X.; Laffaire, J.; Personnaz, L.; Rossier, J.; Dauphinot, L. Transcriptional disruptions in Down syndrome: A case study in the Ts1Cje mouse cerebellum during post-natal development. J. Neurochem. 2006, 97 (Suppl. 1), 104–109. [Google Scholar] [CrossRef] [PubMed]
- Abuhatzira, L.; Shamir, A.; Schones, D.E.; Schaffer, A.A.; Bustin, M. The chromatin-binding protein HMGN1 regulates the expression of methyl CpG-binding protein 2 (MECP2) and affects the behavior of mice. J. Biol. Chem. 2011, 286, 42051–42062. [Google Scholar] [CrossRef] [PubMed]
- Lucey, M.M.; Wang, Y.; Bustin, M.; Duncan, M.K. Differential expression of the HMGN family of chromatin proteins during ocular development. Gene Expr. Patterns 2008, 8, 433–437. [Google Scholar] [CrossRef]
- Li, D.D.; Guo, C.H.; Yue, L.; Duan, C.C.; Yang, Z.Q.; Cao, H.; Guo, B.; Yue, Z.P. Expression, regulation and function of Hmgn3 during decidualization in mice. Mol. Cell. Endocrinol. 2015, 413, 13–25. [Google Scholar] [CrossRef]
- Kurahashi, T.; Furusawa, T.; Ueda, T.; Bustin, M. The nucleosome binding protein HMGN3 is expressed in pancreatic alpha-cells and affects plasma glucagon levels in mice. J. Cell. Biochem. 2010, 109, 49–57. [Google Scholar]
- Lee, H.J.; Kim, J.Y.; Song, I.H.; Park, I.A.; Yu, J.H.; Ahn, J.H.; Gong, G. High mobility group B1 and N1 (HMGB1 and HMGN1) are associated with tumor-infiltrating lymphocytes in HER2-positive breast cancers. Virchows Arch. 2015, 467, 701–709. [Google Scholar] [CrossRef]
- Park, I.A.; Heo, S.H.; Song, I.H.; Kim, Y.A.; Park, H.S.; Bang, W.S.; Park, S.Y.; Jo, J.H.; Lee, H.J.; Gong, G. Endoplasmic reticulum stress induces secretion of high-mobility group proteins and is associated with tumor-infiltrating lymphocytes in triple-negative breast cancer. Oncotarget 2016, 7, 59957–59964. [Google Scholar] [CrossRef][Green Version]
- Wei, F.; Yang, F.; Jiang, X.; Yu, W.; Ren, X. High-mobility group nucleosome-binding protein 1 is a novel clinical biomarker in non-small cell lung cancer. Tumour Biol. 2015, 36, 9405–9410. [Google Scholar] [CrossRef]
- Morande, P.E.; Borge, M.; Abreu, C.; Galletti, J.; Zanetti, S.R.; Nannini, P.; Bezares, R.F.; Pantano, S.; Dighiero, G.; Oppezzo, P.; et al. Surface localization of high-mobility group nucleosome-binding protein 2 on leukemic B cells from patients with chronic lymphocytic leukemia is related to secondary autoimmune hemolytic anemia. Leuk. Lymphoma 2015, 56, 1115–1122. [Google Scholar] [CrossRef]
- Bustin, M.; Alfonso, P.J.; Pash, J.M.; Ward, J.M.; Gearhart, J.D.; Reeves, R.H. Characterization of transgenic mice with an increased content of chromosomal protein HMG-14 in their chromatin. DNA Cell Biol. 1995, 14, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.A.; Chapuy, B.; Lin, C.Y.; Tivey, T.; Li, H.; Townsend, E.C.; van Bodegom, D.; Day, T.A.; Wu, S.C.; Liu, H.; et al. Triplication of a 21q22 region contributes to B cell transformation through HMGN1 overexpression and loss of histone H3 Lys27 trimethylation. Nat. Genet. 2014, 46, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Mowery, C.T.; Reyes, J.M.; Cabal-Hierro, L.; Higby, K.J.; Karlin, K.L.; Wang, J.H.; Kimmerling, R.J.; Cejas, P.; Lim, K.; Li, H.; et al. Trisomy of a Down Syndrome Critical Region Globally Amplifies Transcription via HMGN1 Overexpression. Cell Rep. 2018, 25, 1898–1911.e5. [Google Scholar] [CrossRef] [PubMed]
- Schauwecker, S.M.; Kim, J.J.; Licht, J.D.; Clevenger, C.V. Histone H1 and Chromosomal Protein HMGN2 Regulate Prolactin-induced STAT5 Transcription Factor Recruitment and Function in Breast Cancer Cells. J. Biol. Chem. 2017, 292, 2237–2254. [Google Scholar] [CrossRef] [PubMed]
- Medler, T.R.; Craig, J.M.; Fiorillo, A.A.; Feeney, Y.B.; Harrell, J.C.; Clevenger, C.V. HDAC6 Deacetylates HMGN2 to Regulate Stat5a Activity and Breast Cancer Growth. Mol. Cancer Res. 2016, 14, 994–1008. [Google Scholar] [CrossRef]
- Kugler, J.; Postnikov, Y.V.; Furusawa, T.; Kimura, S.; Bustin, M. Elevated HMGN4 expression potentiates thyroid tumorigenesis. Carcinogenesis 2017, 38, 391–401. [Google Scholar] [CrossRef]
- Lim, J.H.; Catez, F.; Birger, Y.; West, K.L.; Prymakowska-Bosak, M.; Postnikov, Y.V.; Bustin, M. Chromosomal protein HMGN1 modulates histone H3 phosphorylation. Mol. Cell 2004, 15, 573–584. [Google Scholar] [CrossRef]
- Rubinstein, Y.R.; Furusawa, T.; Lim, J.H.; Postnikov, Y.V.; West, K.L.; Birger, Y.; Lee, S.; Nguyen, P.; Trepel, J.B.; Bustin, M. Chromosomal protein HMGN1 modulates the expression of N-cadherin. FEBS J. 2005, 272, 5853–5863. [Google Scholar] [CrossRef]
- Postnikov, Y.V.; Furusawa, T.; Haines, D.C.; Factor, V.M.; Bustin, M. Loss of the nucleosome-binding protein HMGN1 affects the rate of N-nitrosodiethylamine-induced hepatocarcinogenesis in mice. Mol. Cancer Res. 2014, 12, 82–90. [Google Scholar] [CrossRef]
- Birger, Y.; Catez, F.; Furusawa, T.; Lim, J.H.; Prymakowska-Bosak, M.; West, K.L.; Postnikov, Y.V.; Haines, D.C.; Bustin, M. Increased tumorigenicity and sensitivity to ionizing radiation upon loss of chromosomal protein HMGN1. Cancer Res. 2005, 65, 6711–6718. [Google Scholar] [CrossRef][Green Version]
- West, K.L. HMGN proteins play roles in DNA repair and gene expression in mammalian cells. Biochem. Soc. Trans. 2004, 32, 918–919. [Google Scholar] [CrossRef] [PubMed]
- Gerlitz, G. HMGNs, DNA repair and cancer. Biochim. Biophys. Acta 2010, 1799, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Strahler, J.R.; Dodgson, J.B. Neither HMG-14a nor HMG-17 gene function is required for growth of chicken DT40 cells or maintenance of DNaseI-hypersensitive sites. Nucleic Acids Res. 1997, 25, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, M.; Gonzalez, R.W.; Patil, H.; Ueda, T.; Lim, J.H.; Kraemer, K.H.; Bustin, M.; Bergel, M. The nucleosome-binding protein HMGN2 modulates global genome repair. FEBS J. 2009, 276, 6646–6657. [Google Scholar] [CrossRef] [PubMed]
- Chavez, C.; Cruz-Becerra, G.; Fei, J.; Kassavetis, G.A.; Kadonaga, J.T. The tardigrade damage suppressor protein binds to nucleosomes and protects DNA from hydroxyl radicals. Elife 2019, 8, e47682. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Gerlitz, G.; Furusawa, T.; Catez, F.; Nussenzweig, A.; Oh, K.S.; Kraemer, K.H.; Shiloh, Y.; Bustin, M. Activation of ATM depends on chromatin interactions occurring before induction of DNA damage. Nat. Cell Biol. 2009, 11, 92–96. [Google Scholar] [CrossRef]
- Gerlitz, G.; Bustin, M. Nucleosome binding proteins potentiate ATM activation and DNA damage response by modifying chromatin. Cell Cycle 2009, 8, 1641. [Google Scholar] [CrossRef][Green Version]
- Masaoka, A.; Gassman, N.R.; Kedar, P.S.; Prasad, R.; Hou, E.W.; Horton, J.K.; Bustin, M.; Wilson, S.H. HMGN1 protein regulates poly(ADP-ribose) polymerase-1 (PARP-1) self-PARylation in mouse fibroblasts. J. Biol. Chem. 2012, 287, 27648–27658. [Google Scholar] [CrossRef]
- Gardiner, K.; Herault, Y.; Lott, I.T.; Antonarakis, S.E.; Reeves, R.H.; Dierssen, M. Down syndrome: From understanding the neurobiology to therapy. J. Neurosci. 2010, 30, 14943–14945. [Google Scholar] [CrossRef]
- Cabin, D.E.; McKee-Johnson, J.W.; Matesic, L.E.; Wiltshire, T.; Rue, E.E.; Mjaatvedt, A.E.; Huo, Y.K.; Korenberg, J.R.; Reeves, R.H. Physical and comparative mapping of distal mouse chromosome 16. 5 p5. Genome Res. 1998, 8, 940–950. [Google Scholar] [CrossRef][Green Version]
- Ling, K.H.; Hewitt, C.A.; Tan, K.L.; Cheah, P.S.; Vidyadaran, S.; Lai, M.I.; Lee, H.C.; Simpson, K.; Hyde, L.; Pritchard, M.A.; et al. Functional transcriptome analysis of the postnatal brain of the Ts1Cje mouse model for Down syndrome reveals global disruption of interferon-related molecular networks. BMC Genom. 2014, 15, 624. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.D.; Liu, N.; Xu, X.J. Bioinformatics analysis of biomarkers and transcriptional factor motifs in Down syndrome. Braz. J. Med. Biol. Res. 2014, 47, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Carney, R.M.; Wolpert, C.M.; Ravan, S.A.; Shahbazian, M.; Ashley-Koch, A.; Cuccaro, M.L.; Vance, J.M.; Pericak-Vance, M.A. Identification of MeCP2 mutations in a series of females with autistic disorder. Pediatr. Neurol. 2003, 28, 205–211. [Google Scholar] [CrossRef]
- Yamazaki, M.; Itoh, S.; Sasaki, N.; Tanabe, K.; Uchiyama, M. Modification of the dialysis membrane method for drug release from suppositories. Pharm. Res. 1993, 10, 927–929. [Google Scholar] [CrossRef]
- Ramocki, M.B.; Tavyev, Y.J.; Peters, S.U. The MECP2 duplication syndrome. Am. J. Med. Genet. A 2010, 152A, 1079–1088. [Google Scholar] [CrossRef]
- Bustin, M.; Reisch, J.; Einck, L.; Klippel, J.H. Autoantibodies to nucleosomal proteins: Antibodies to HMG-17 in autoimmune diseases. Science 1982, 215, 1245–1247. [Google Scholar] [CrossRef]
- Neuer, G.; Bautz, F.A.; Bustin, M.; Michels, H.; Truckenbrodt, H. Sera from JRA patients contain antibodies against a defined epitope in chromosomal protein HMG-17. Autoimmunity 1994, 17, 23–30. [Google Scholar] [CrossRef]
- Neuer, G.; Bustin, M.; Michels, H.; Truckenbrodt, H.; Bautz, F.A. Autoantibodies to the chromosomal protein HMG-17 in juvenile rheumatoid arthritis. Arthritis Rheumatol. 1992, 35, 472–475. [Google Scholar] [CrossRef]
- Vlachoyiannopoulos, P.G.; Boumba, V.A.; Tzioufas, A.G.; Seferiadis, C.; Tsolas, O.; Moutsopoulos, H.M. Autoantibodies to HMG-17 nucleosomal protein in patients with scleroderma. J. Autoimmun. 1994, 7, 193–201. [Google Scholar] [CrossRef]
- Zheng, S.; Ren, L.; Li, H.; Shen, X.; Yang, X.; Li, N.; Wang, X.; Guo, X.; Wang, X.; Huang, N. High-mobility group nucleosome-binding domain 2 protein inhibits the invasion of Klebsiella pneumoniae into mouse lungs in vivo. Mol. Med. Rep. 2015, 12, 1279–1285. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, N.; Wu, Q.; Wang, B. HMGN2: A novel antimicrobial effector molecule of human mononuclear leukocytes? J. Leukoc. Biol. 2005, 78, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Miao, J.; Zhang, F.; Xiong, F.; Zhu, F.; Li, J.; Wang, X.; Chen, S.; Chen, J.; Huang, N.; et al. Non-histone nuclear protein HMGN2 differently regulates the urothelium barrier function by altering expression of antimicrobial peptides and tight junction protein genes in UPEC J96-infected bladder epithelial cell monolayer. Acta Biochim. Pol. 2018, 65, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhu, F.; Yang, J.; Zhang, W.; Liu, K.; Ren, L.; Xiong, F.; Lu, K.; Li, D.; Zeng, R.; et al. High-mobility group protein N2 induces autophagy by activating AMPK/ULK1 pathway and thereby boosts UPEC proliferation within bladder epithelial cells. Biochem. Biophys. Res. Commun. 2019, 513, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, X.F.; Zhou, X.E.; Shi, Y.E.; Deng, L.X.; Ma, Y.; Wang, X.Y.; Li, J.Y.; Huang, N. Antibacterial mechanism of high-mobility group nucleosomal-binding domain 2 on the Gram-negative bacteria Escherichia coli. J. Zhejiang Univ. Sci. B 2017, 18, 410–420. [Google Scholar] [CrossRef]
- Wang, X.; Chen, S.; Ren, H.; Chen, J.; Li, J.; Wang, Y.; Hua, Y.; Wang, X.; Huang, N. HMGN2 regulates non-tuberculous mycobacteria survival via modulation of M1 macrophage polarization. J. Cell. Mol. Med. 2019, 23, 7985–7998. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Yang, H.; Tracey, K.J.; Bustin, M.; Oppenheim, J.J. High mobility group box-1 protein induces the migration and activation of human dendritic cells and acts as an alarmin. J. Leukoc. Biol. 2007, 81, 59–66. [Google Scholar] [CrossRef]
- Yang, D.; Postnikov, Y.V.; Li, Y.; Tewary, P.; de la Rosa, G.; Wei, F.; Klinman, D.; Gioannini, T.; Weiss, J.P.; Furusawa, T.; et al. High-mobility group nucleosome-binding protein 1 acts as an alarmin and is critical for lipopolysaccharide-induced immune responses. J. Exp. Med. 2012, 209, 157–171. [Google Scholar] [CrossRef]
- Wei, F.; Yang, D.; Tewary, P.; Li, Y.; Li, S.; Chen, X.; Howard, O.M.; Bustin, M.; Oppenheim, J.J. The Alarmin HMGN1 contributes to antitumor immunity and is a potent immunoadjuvant. Cancer Res. 2014, 74, 5989–5998. [Google Scholar] [CrossRef]
- Nie, Y.; Yang, D.; Trivett, A.; Han, Z.; Xin, H.; Chen, X.; Oppenheim, J.J. Development of a Curative Therapeutic Vaccine (TheraVac) for the Treatment of Large Established Tumors. Sci. Rep. 2017, 7, 14186. [Google Scholar] [CrossRef]
- Alam, M.M.; Yang, D.; Trivett, A.; Meyer, T.J.; Oppenheim, J.J. HMGN1 and R848 Synergistically Activate Dendritic Cells Using Multiple Signaling Pathways. Front. Immunol. 2018, 9, 2982. [Google Scholar] [CrossRef]
- Chen, C.Y.; Ueha, S.; Ishiwata, Y.; Yokochi, S.; Yang, D.; Oppenheim, J.J.; Ogiwara, H.; Shichino, S.; Deshimaru, S.; Shand, F.H.W.; et al. Combined treatment with HMGN1 and anti-CD4 depleting antibody reverses T cell exhaustion and exerts robust anti-tumor effects in mice. J. Immunother Cancer 2019, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zeng, Z.; Li, J.; Zheng, Y.; Wei, F.; Ren, X. PD-1/PD-L1 Axis, Rather Than High-Mobility Group Alarmins or CD8+ Tumor-Infiltrating Lymphocytes, Is Associated with Survival in Head and Neck Squamous Cell Carcinoma Patients Who Received Surgical Resection. Front. Oncol. 2018, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, M.; Li, W.; Wu, C.; Salovska, B.; Liu, Y. Combining Rapid Data Independent Acquisition and CRISPR Gene Deletion for Studying Potential Protein Functions: A Case of HMGN1. Proteomics 2019, 19, e1800438. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Bustin, M.; Oppenheim, J.J. Harnessing the alarmin HMGN1 for anticancer therapy. Immunotherapy 2015, 7, 1129–1131. [Google Scholar] [CrossRef]
- Yang, D.; Han, Z.; Alam, M.M.; Oppenheim, J.J. High-mobility group nucleosome binding domain 1 (HMGN1) functions as a Th1-polarizing alarmin. Semin. Immunol. 2018, 38, 49–53. [Google Scholar] [CrossRef]
- Zhang, S.; Deng, T.; Tang, W.; He, B.; Furusawa, T.; Ambs, S.; Bustin, M. Epigenetic regulation of REX1 expression and chromatin binding specificity by HMGNs. Nucleic Acids Res. 2019, 47, 4449–4461. [Google Scholar] [CrossRef]


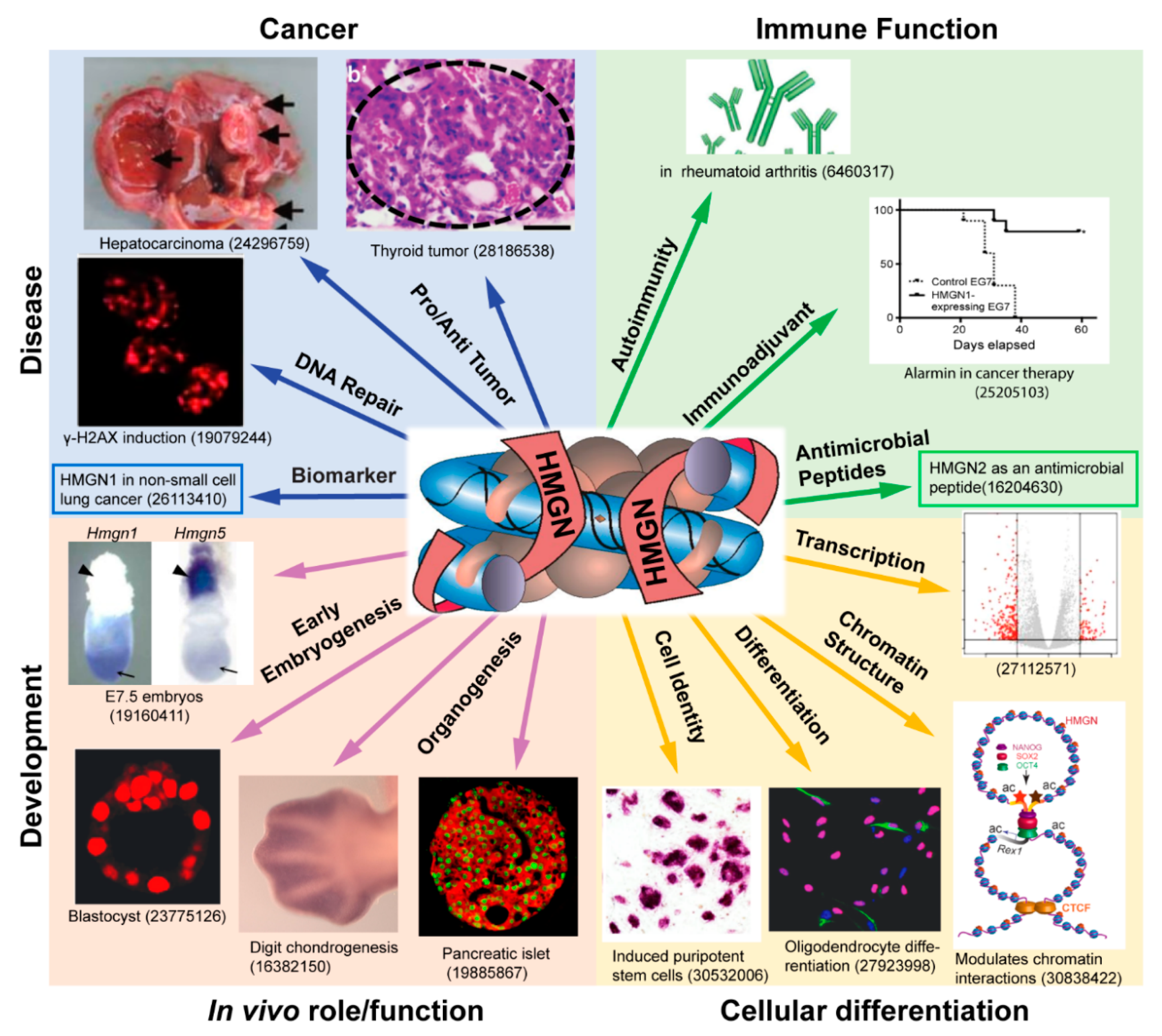{kind=link}
| Gene | Modification | References |
|---|---|---|
| Hmgn1 | Knockout | Birger, Y. et al., 2003 |
| Hmgn2 | Knockout | Deng, T. et al., 2015 |
| Hmgn2# | Knockout | Gao, X.L. et al., 2019 |
| Hmgn3 | Knockout | Ueda, T. et al., 2009 |
| Hmgn5 | Knockout | Ciappio, E.D. et al., 2014 |
| Hmgn1/Hmgn2 | Knockout | Deng, T. et al., 2015 |
| Human HMGN1 | Transgenic | Bustin, M. et al., 1995 |
| Human HMGN4 | Transgenic | Kugler, J. et al., 2017 |
| Hmgn5 | Transgenic | Furusawa, T. et al., 2016 |
| HMGN | Species | Study Model | Summary | Reference (PMID) |
|---|---|---|---|---|
| HMGN1 | Mouse | Myoblast differentiation | Overexpression of human HMGN1 inhibited the differentiation of mouse C2C12 myoblast cells into myotubes | 8514795 |
| HMGN1 | Mouse | Transcription in MEFs | HMGN1 is a negative regulator of N-Cadherin gene. N-Cadherin was also higher in the early embryos of HMGN1 KO compared to WT | 16279949 |
| HMGN1 | Mouse | Cornea differentiation and maturation | In corneal epithelium of Hmgn1 KO mice, corneal differentiation, maturation and gene expression were affected. In the wildtype mice, HMGN1 is colocalized with p63 and p63 expression was absent in the central region of the Hmgn1 KO cornea | 16466397 |
| HMGN1 | Mouse | Chondrocyte differentiation | In developing limb bud, Hmgn1 expression is inversely correlated to Sox9 expression and it is down regulated during chondrogenesis | 16382150 |
| HMGN1 | Mouse | Myogenesis and adipogenesis | Myogenesis or adipogenesis were induced in Hmgn1 knock down (KD) NIH3T3 cell line by overexpressing MyoD or C/EBPalpha. Hmgn1 Knockdown cells did not show any differences when compared with control cells | 16451822 |
| HMGN1 | Mouse | Digit formation | At E13.5 mouse autopod, interdigital expression of hmgn1 mRNA is highest at the digit-interdigit junction and is greatly down-regulated in the Raldh2 KO mice | 20034106 |
| HMGN1 | Mouse | embryonic stem cells (ESCs) | HMGN1 is strongly expressed in oocytes and throughout all preimplantation stages. Loss of HMGN1 reduces the DNase I hypersensitivity (DHS) at CpG island promoters in ES cells | 23775126 |
| HMGN1 | Human | Transgenic mice | HMGN1 is negative regulator for methyl CpG-binding protein 2. Hence, mice overexpressing HMGN1 displayed abnormalities in activity and anxiety and to social deficits | 22009741 |
| HMGN1 | Mouse | Down syndrome model mouse. | HMGN1 is overexpressed in Down syndrome and in all down syndrome model mice | 2140193 |
| 16635258 | ||||
| HMGN2 | Mouse | Kidney differentiation | During metanephric kidney development, HMGN2 expression is highly linked to induction and differentiation in kidney organogenesis | 11683498 |
| HMGN1 HMGN2 | Mouse | Myoblast differentiation | During myogenesis the level of Hmgn1 and Hmgn2 mRNA were decreased and this downregulation is a differentiation related event | 1689723 |
| HMGN1 HMGN2 | Chicken | Erythroid cells | Erythroid cells from 5-day chicken embryos contain higher level of Hmgn mRNAs than cells from 14-day embryos. Change from primitive to definitive erythroid lineage during embryogenesis is accompanied by a change in the expression of HMGN proteins. | 1993650 |
| HMGN1 HMGN2 | Rat Human | Osteoblast differentiation and monocyte differentiation | In both primary cultures of calvarial-derived rat osteoblasts and human leukemia HL-60 cells, HMGN1 and HMGN2 are preferentially expressed in the proliferating state, and the levels are down-regulated during differentiation. | 8496248 |
| HMGN1 HMGN2 | Mouse | Early embryonic development | HMGN1 and HMGN2 were detected throughout oogenesis and embryonic transcripts accumulate beyond the two-cell stage. Depletion of HMGNs in one-cell or two-cell mouse embryos delays subsequent progression. | 11133167 |
| HMGN1 HMGN2 | Xenopus | Early embryonic development | Depletion of Hmgn1 or Hmgn2 resulted in developmental defects after mid blastula transition (MBT) in Xenopus embryos, and re-expression of HMGN2 rescued these phenotypes | 14568106 |
| HMGN1 HMGN2 | Bovine | Maternal-zygotic transition | During bovine embryo early stage development, HMGN1 and HMGN2 are expressed in oocytes but are downregulated after fertilization and almost disappear by the 8-cell stage | 14960490 |
| HMGN1 HMGN2 | Mouse | Hair follicle cycle | Hmgn mRNAs was expressed at the onset of hair follicle development and during the adult hair follicle cycle | 19303948 |
| HMGN1 HMGN2 | Mouse | MEF, mouse behavior | Genome wide mapping of DHSs sites in MEFs from HMGN1 KO, HMGN2 KO and HMGN DKO revealed that loss of both, but not single HMGN variant leads to significant remodeling of DHS especially at enhancer regions | 26156321 |
| HMGN1 HMGN2 | Mouse | B lymphocyte | Loss of HMGN1 and HMGN2 proteins affect gene expression by modulating chromatin regulatory sites during activation of naïve B cells | 27112571 |
| HMGN1 HMGN2 | Mouse | Oligodendrocyte differentiation | Loss of HMGNs affect the expression of OLIG2 and OLIG1, thereby affecting oligodendrocyte development and nerve myelination, and mouse behavior | 27923998 |
| HMGN1 HMGN2 | Mouse | MEFs, induced pluripotent stem cells (iPSCs) | MEFs from Hmgn1/2 DKO mice showed enhanced rate of iPSCs induction by OSKM factors, and into neurons induced by ASCL1 | 30532006 |
| HMGN1 HMGN2 HMGN3 | Mouse | Eye development | HMGN1, HMGN2 and HMGN3 displayed dynamic and specific expression patterns during ocular development (E10.5, E13.5 and newborn) and adult eye | 18502697 |
| HMGN1 HMGN2 HMGN3 | Mouse | Astrocyte differentiation | Ectopic expression of either HMGN1, HMGN2 or HMGN3 in neural precursor cells (NPCs) or in late embryonic neocortex promoted the generation of astrocytes while knockdown suppressed astrocyte differentiation. | 25069414 |
| HMGN2 | Bovine | Early embryonic development | An in vitro fertilization study of bovine embryos revealed that embryos with enhanced levels of HMGN2 failed to develop into blastocyst due to the altered chromatin remodeling which might be caused by hyperacetylation of Histone H3 at lysine 14 residue | 17712799 |
| HMGN2 | Mouse | Tooth development | HMGN2 binds to Pitx2 and inhibits Pitx2 mediated Amelogenin expression during tooth development | 23975681 |
| HMGN2 | Mouse | Corticogenesis (Cerebral cortex development) | Hmgn2 KO mice exhibited microcephalus phenotype with reduced cortical surface area and almost normal radial corticogenesis | 31699896 |
| HMGN3 | Xenopus | Anuran metamorphosis | Hmgn3 expression is upregulated during thyroid hormone dependent anuran metamorphosis. | 11921340 |
| HMGN3 | Mouse | Retina | HMGN3 is highly expressed in glia cells and eye. HMGN3 binds to glycine transporter 1 (Glyt1) gene and upregulates its expression. | 15082770 |
| HMGN3 | Mouse | Pancreas (beta cells) | Loss of HMGN3 abrogates glucose stimulated insulin secretion and HMGN3 KO mice possess diabetic phenotype. HMGN3 along with PDX1regulates Glu2 gene | 19651901 |
| HMGN3 | Bovine | Early embryonic development | Hmgn3 mRNA was expressed at similar levels in bovine matured oocytes and 2–4-cell embryos, but expression was higher in 8-16-cell embryos, morulae and blastocysts | 19393058 |
| HMGN3 | Mouse | Pancreas (alpha cells) | HMGN3 is highly expressed in pancreatic alpha cells, and that Hmgn3 KO mice displayed reduced glucagon levels in their blood | 19885867 |
| HMGN3 | Mouse | Decidualization | HMGN3 is highly expressed in decidua and decidualizing stromal cells, and also in the artificially stimulated decidualization | 26112184 |
| HMGN5 | Mouse | Placenta development | Hmgn5 mRNA is strongly expressed in ectoplacental cone in E7.5 embryo and its expression retained in developing placenta and in trophoblast giant cells | 19160411 |
| HMGN5 | Mouse | Metabolomics | Metabolomic analysis of liver extracts and urine from Hmgn5 KO revealed that loss of HMGN5 leads to downregulation of glutathione peroxidase 6 (Gpx6) and hexokinase 1 (Hk1), thereby affects glutathione metabolism | 24392144 |
| HMGN5 | Mouse | Heart development | Transgenic mice overexpressing HMGN5 (both ubiquitous or heart specific) are normal at birth but develop hypertrophic heart with large cardiomyocytes, deformed nuclei and disrupted lamina and die of cardiac malfunction | 25609380 |
| HMGN | Study Model | Disease | Role/Function | Summary | Reference (PMID) |
|---|---|---|---|---|---|
| HMGN1 | Mice | Cancer | Immunoadjuvant | Augments antitumor immunity of alpha-fetoprotein expressing lentiviral vaccines in hepatocellular carcinoma mice | 31281528 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | Treatment with HMGN1 and anti-CD4 depleting antibody reverses T cell exhaustion and exerts robust anti-tumor effects in mice | 30696484 |
| HMGN1 | Cell culture | Cancer | Immunoadjuvant | HMGN1 promotes dendritic cells (DC) recruitment through interacting with a Gαi protein-coupled receptor and activates DCs predominantly through triggering toll like receptor 4 (TLR4) | 29503123 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | Intratumoral delivery of HMGN1 and R848 along with cytoxan (TheraVac) eradicated large established CT26, Renca and EG7 tumors in mice | 29079801 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | Intratumoral injections cured mice harboring large established subcutaneous Hepa1-6 hepatomas | 28881713 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | HMGN1 contributes to antitumor immunity | 25205103 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | HMGN1 induced DC maturation via TLR4, recruitment of antigen presenting cells at sites of injection and promoted antigen-specific immune response upon co-administration with antigens | 22184635 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | DNA(TA) encapsulated within peptide hydrogels encodes for a melanoma-specific gp100 antigen fused to the HMGN1 enhanced immunostimulatory effects in tumor mice model | 25890750 |
| HMGN1 | Mice | Cancer | Protumor activity | Down syndrome associated triplication of a 21q22 region contributes to B cell proliferation with increased risk of B cell acute lymphoblastic leukemia through HMGN1 overexpression | 24747640 |
| HMGN1 | Cell culture | Cancer | Protumor activity | High expression of HMGN1 in highly metastatic cells of MDA-MB-435 suggests the association of HMGN1 with metastasis of breast cancer cells | 16979889 |
| HMGN1 | Patient’s studies | Cancer | Protumor activity | Endoplasmic reticulum stress induces secretion of HMGN1 and is associated with tumor-infiltrating lymphocytes in triple-negative breast cancer | 27494867 |
| HMGN1 | Patient’s studies | Cancer | Protumor activity | High cytoplasmic expression of HMGN1 was associated with a high histological grade, high levels of tumor infiltering lymphocytes (TILs) in HER2-positive breast cancer tissues | 26445971 |
| HMGN1 | Patient’s studies | Cancer | Biomarker | Patients with high serum HMGN1 had a poorer overall survival after curative pneumonectomy in non-small cell lung cancer | 26113410 |
| HMGN1 | Cell culture | Cancer | Transcriptional regulator | HMGN1 and HMGN2 are synthesized throughout the S-phase and persists through the cell cycle in Burkitt’s lymphoma | 8670211 |
| HMGN1 | Mice | Cancer | Antitumor activity | Loss of HMGN1 accelerates the progression of N-nitrosodiethylamine-induced hepatocarcinogenesis | 24296759 |
| HMGN1 | Mice | Cancer | Antitumor activity | Loss of HMGN1 increases the tumorigenicity and sensitivity to Ionizing radiations | 16061652 |
| HMGN1 | Patient’s studies | Down Syndrome | Transcriptional regulator | Gene dosage studies in cultured cells and brain tissue samples obtained from Down syndrome patients suggest that HMGN1 as contributing factor in the etiology of Down syndrome | 1825298 |
| HMGN1 | Mice | Down Syndrome | Gene locus | HMGN1 maps to the 21q22.3 in humans, the region associated with the pathogenesis of Down syndrome, and to chromosome 16 in mice | 2140193 |
| HMGN1 | Mice | Down Syndrome | Transcriptional regulator | Overexpression of the HMGN1 (encoded on chr21q22) reiterates transcriptional changes seen with triplication of Down syndrome critical region on distal chromosome 21 | 30428356 |
| HMGN1 | Mice | Diabetic nephropathy | Transcriptional regulator | HMGN1 expression correlates with epithelial mesenchymal transition, renal fibrosis in diabetic nephropathy mice | 31203283 |
| HMGN1 | Mice | Diabetic nephropathy | Transcriptional regulator | Insulin growth factor 1 receptor inhibitor attenuates diabetic nephropathy by suppressing HMGN1-TLR4 pathway | 29384065 |
| HMGN1 | Rats | Amnesia | Biomarker | Increased HMGN1 phosphorylation by protein kinase CK2 decreases amnesia in aged rats | 14635996 |
| HMGN1 | Patient’s studies | Systemic sclerosis | Immunogen in autoimmunity | Autoantibodies against HMGN1 and HMGN2 were found in systemic sclerosis patients’ sera | 7869312 |
| HMGN1 | Patient’s studies | Drug induced lupus | Immunogen in autoimmunity | Autoantibodies against HMGN1 and HMGN2 were found in drug induced lupus patients | 7907477 |
| HMGN1 | Patient’s studies | Primary pulmonary hypertension | Immunogen in autoimmunity | Autoantibodies against HMGN1 were found in sera from primary pulmonary hypertension patients | 8918589 |
| HMGN1 | Mice | Immune Tolerance | TLR-4 agonist | HMGN1 induces innate immune tolerance in a TLR4 dependent Sirtuin-1 mediated deacetylation in human blood peripheral mononuclear cells | 29593748 |
| HMGN2 | Mice | Cancer | Antitumor activity | The tumor volumes in SaO2 and U2-OS osteocarcinoma subcutaneous nude mouse models treated with overexpressing HMGN2 lentivirus were significantly decreased | 25530340 |
| HMGN2 | Mice | Cancer | Antitumor activity | Overexpression of HMGN2 significantly decreased migration capacity, increased apoptosis of MCF7 breast cancer cells both in vitro and in vivo | 30655878 |
| HMGN2 | Cell culture | Cancer | Protumor activity | HMGN2 was increased in tumor cell line MDA-MB-468 and also in metastatic oral squamous cell carcinoma tissues | 29541221 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | Isopentenyl pyrophosphate activated γδ T cells expresses high level of HMGN2 which contributes to antitumor activity | 29401165 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | Overexpression of HMGN2 in by lentivirus, inhibits the growth and migration of osteosarcoma cells | 25530340 |
| HMGN2 | Cell culture | Cancer | Signal transducer | Surface expressed of HMGN2 on leukemic B cells binds to autoantigen erythrocyte protein band 3 and contributes to initiation of autoimmune hemolytic anemia | 25156469 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | Phytohemagglutinin activated CD8 T cells releases high level of HMGN2 and kill tumor cells | 25060707 |
| HMGN2 | Cell culture | Cancer | Protumor activity | Prolactin induced H1 disassociation mediated by HMGN2 induce STAT5 recruitment and breast cancer pathogenesis | 28035005 |
| HMGN2 | Patient studies | Cancer | Protumor activity | Expression of HMGN2 was higher in human leukemia cells with chronic myelogenic leukemia | 8528142 |
| HMGN2 | Cell culture | Cancer | Protumor activity | HDAC6 mediated deacetylation of HMGN2 enhances Stat5a transcriptional activity, thereby regulating prolactin-induced gene transcription and breast cancer growth | 27358110 |
| HMGN2 | Cell culture | Cancer | Biomarker | HMGN1 and HMGN2 are synthesized throughout the S-phase and persists through the cell cycle in Burkitt’s lymphoma | 8670211 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | HMGN2 is a direct target of MicroRNA-23a which plays crucial role in epithelial mesenchymal transition (EMT) in lung cancer cells suggests its role | 23437179 |
| HMGN2 | Mice | Cancer | Antitumor activity | HMGN2 exerts inhibitory effect on tumors in human tongue carcinoma transplanted nude mice | 24665631 |
| HMGN2 | Mice | Cancer | Antitumor activity | HMGN2 treatment inhibited the growth, increased apoptosis of Tca8113 cells in vitro and in vivo | 24348831 |
| HMGN2 | Cell culture | Cancer | Antimicrobial activity | 31-aa peptide (F3) in the HMGN2 sequence homed to HL-60 and MDA-MB-435 tumors upon I.v. injection | 12032302 |
| HMGN2 | Patient studies | Cancer | Immunogen in autoimmunity | High throughput screening of autoantibodies in ovarian cancer ascitic fluids revealed HMGN2 as an antigen | 31428516 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | HMGN2 regulates the expression of antimicrobial peptides in Uropathogenic Escherichia coli J96-infected bladder epithelial cell monolayer | 29549670 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 displayed anti-bacterial and anti-biofilm activity against Escherichia coli K12 | 28471113 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 inhibits Pseudomonas aeruginosa internalization in A549 cells | 28408162 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 protects bladder epithelial cells from Klebsiella invasion | 21720014 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | Knockdown of HMGN2 affected the survival of non-tuberculosis mycobacteria in macrophage | 31596045 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | HMGN2 is a target of miR-155, which is downregulated in Klebsiella pneumoniae infection suggested the role in controlling infection | 27534887 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | Knockdown of HMGN2 increases the internalization of Klebsiella pneumoniae by respiratory epithelial cells | 27460641 |
| HMGN2 | Mice | Infection | Antimicrobial activity | HMGN2 is crucial for LPS induced expression of murine β-defensin-3 and -4 in mice | 21594618 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 is crucial for LPS induced expression of murine β-defensin-3 and -4 in A549 cells | 21518253 |
| HMGN2 | Mice | Infection | Antimicrobial activity | HMGN2 inhibits the invasion of Klebsiella pneumoniae into mouse lungs in vivo | 25760831 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 inhibited hepatitis B virus infection and replication in HepG2.2.15 cell line | 19150374 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 inhibited internalization of Klebsiella pneumoniae into cultured bladder epithelial cells | 21778192 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | N-terminal amino sequence was PKRKAEGDAK of HMGN2 is a novel antimicrobial peptide | 16204630 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 showed potent antimicrobial activity against E coli ML-35p, P aeruginosa ATCC 27853 | 16115376 |
| HMGN2 | Patient’s studies | Systemic lupus erythromatosus | Immunogen in autoimmunity | Autoantibodies against HMGN2 were found in systemic lupus erythromatosis (SLE) patients | 8037838 |
| HMGN2 | Patient’s studies | Systemic lupus erythromatosus | Immunogen in autoimmunity | Autoantibodies to HMGN2 were found in different autoimmune diseases and are more frequent in Systemic lupus erythromatosus | 8318042 |
| HMGN2 | Patient’s studies | Systemic lupus erythromatosus | Immunogen in autoimmunity | Autoantibodies to HMGN2 were present in serums from patients with systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), and mixed connective tissue disease (MCTD) | 6460317 |
| HMGN2 | Patient’s studies | Drug induced lupus | Immunogen in autoimmunity | Autoantibodies against HMGN1 and HMGN2 were found in drug induced lupus patients | 7907477 |
| HMGN2 | Patient’s studies | Juvenile Rheumatoid Arthritis | Immunogen in autoimmunity | Autoantibodies to HMGN2 were detected in ANA-positive patients with periarticular-onset Juvenile Rheumatoid Arthritis | 7517709 |
| HMGN2 | Patient’s studies | Juvenile Rheumatoid Arthritis | Immunogen in autoimmunity | Autoantibodies to HMGN2 were detected in patients with Juvenile Rheumatoid Arthritis | 1567496 |
| HMGN2 | Patient’s studies | Periodontitis | Biomarker | HMGN2 in saliva of periodontitis patients was higher especially with severe periodontitis | 28591948 |
| HMGN2 | Mice | Microcephaly | Transcriptional regulator | HMGN2 protects against Microcephaly by maintaining chromatin accessibility during corticogenesis | 31699896 |
| HMGN3 | Mice | Diabetes | Anti-diabetic | Hmgn3-/- mice which have a mild diabetic phenotype, with reduced glucagon levels in the blood | 19885867 |
| HMGN3 | Mice | Diabetes | Anti-diabetic | In mice, loss of HMGN3 blunt glucose-stimulated insulin secretion and have a diabetic phenotype. HMGN3 regulates Glut4 gene in pancreatic beta cells | 19651901 |
| HMGN4 | Patient’s studies | Cancer | Biomarker | Patients with high expression of HMGN4 have high grade tumors, shorter overall survival in hepatocellular carcinoma | 30272824 |
| HMGN4 | Mice | Cancer | Protumor activity | Mouse and human cells overexpressing HMGN4 displayed increased tumorigenicity in thyroid tumor mouse model | 28186538 |
| HMGN5 | Cell culture | Cancer | Protumor activity | microRNA-183-3p directly inhibits the expression of HMGN5 and prostate cancer progression | 31314587 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 expression was upregulated in pancreatic ductal adenocarcinoma tissues and cell lines. | 30128022 |
| HMGN5 | Cell culture | Cancer | Protumor activity | miR-488 inhibits cell growth and metastasis in renal cell carcinoma by abrogating HMGN5 expression | 29713189 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 expression was significantly upregulated in bladder transitional cell carcinoma tissues | 29509244 |
| HMGN5 | Cell culture | Cancer | Protumor activity | Silencing HMGN5 promotes chemosensitivity of human bladder cancer cells to cisplatin | 29163683 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 was significantly upregulated in esophageal squamous cell carcinoma cells. Silencing of HMGN5 significantly inhibited cell growth and induced cell apoptosis of ESCC cells | 28914995 |
| HMGN5 | Cell culture | Cancer | Protumor activity | microRNA-495 inhibits the proliferation and invasion and induces the apoptosis of osteosarcoma cells by inhibiting HMGN5 | 28627703 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-140-5p expression enhances osteosarcoma chemoresistance by targeting HMGN5 and autophagy | 28341864 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-409-3p decreases glioma cell invasion and proliferation by targeting HMGN5 | 28109076 |
| HMGN5 | Cell culture | Cancer | Protumor activity | microRNA-340 inhibits tumorigenic potential of prostate cancer cells by abrogating HMGN5 expression | 26394192 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing enhances apoptosis, suppresses invasion and increases chemosensitivity to temozolomide in meningiomas | 26315299 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing suppresses the viability and invasion of human urothelial bladder cancer 5637 cells | 25796505 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 is over expressed in prostate cancer cells and activates MAPK and its expression is correlated with chemosensitivity | 25572120 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 is highly expressed in breast cancer cells and prompts proliferation and invasion | 25315189 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing in cells exhibit increased apoptosis rate and decreased colonogenic survival in response to 2–8 Gy ionizing radiation | 25307178 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 was positively correlated with pathologic staging and TNM staging of osteosarcoma | 24687550 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 increases drug resistance in osteosarcoma cell lines U-2OS and MG63 through upregulating autophagy | 24664583 |
| HMGN5 | Cell culture | Cancer | Protumor activity | Silencing of HMGN5 induces apoptosis through the modulation of a mitochondrial pathway and Bcl-2 family proteins in prostate cancer cells | 22504871 |
| HMGN5 | Cell culture | Cancer | Protumor activity | Knockdown of HMGN5 slowed down the cell cycle at the G0/G1 phase in human lung cancer cell lines A549 and H1299 | 22994738 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing inhibits the growth and invasion of clear cell renal cell carcinoma cells and its expression is associated with tumor grade | 22420896 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 promotes the viability of bladder cancer cells through increased cell proliferation, invasion ability of metastatic bladder cancer cells through the upregulation of MMP-9 | 21695596 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown causes cell cycle arrest in human glioma U251 and U87 cells and it is highly expressed in high grade and low-grade tumor tissues | 21373965 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown causes cell cycle arrest in prostate cancer cell line DU145 | 20531280 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown inhibits proliferation and tumorigenicity of BGC823 and SGC7901 gastric cancer cell lines | 30854121 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-186 suppresses the growth and metastasis of bladder cancer by targeting HMGN5 | 26290438 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown inhibits proliferation and tumorogenecity of BGC823 and SGC7901 gastric cancer cell lines | 30854121 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-186 suppresses the growth and metastasis of bladder cancer by targeting HMGN5 | 26290438 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanduri, R.; Furusawa, T.; Bustin, M. Biological Functions of HMGN Chromosomal Proteins. Int. J. Mol. Sci. 2020, 21, 449. https://doi.org/10.3390/ijms21020449
Nanduri R, Furusawa T, Bustin M. Biological Functions of HMGN Chromosomal Proteins. International Journal of Molecular Sciences. 2020; 21(2):449. https://doi.org/10.3390/ijms21020449
Chicago/Turabian StyleNanduri, Ravikanth, Takashi Furusawa, and Michael Bustin. 2020. "Biological Functions of HMGN Chromosomal Proteins" International Journal of Molecular Sciences 21, no. 2: 449. https://doi.org/10.3390/ijms21020449
APA StyleNanduri, R., Furusawa, T., & Bustin, M. (2020). Biological Functions of HMGN Chromosomal Proteins. International Journal of Molecular Sciences, 21(2), 449. https://doi.org/10.3390/ijms21020449