4,4′-Diaminodiphenyl Sulfone (DDS) as an Inflammasome Competitor


Abstract
1. Introduction
2. Materials and Methods
2.1. Respiratory Tract Specimens and Symptoms of COVID-19
2.2. The Topological Properties of DDS Provide Diverse Clues
2.3. The Longevity of Leprosy-Affected Males in Spite of Several Pandemic Viral Diseases
2.4. Cohort Study for Treatment of Alzheimer’s Disease with 4,4′-Diaminodiphenylsulfone
3. Results
3.1. DDS Has the Clinical Manifestations of COVID-19 Infection in Humans
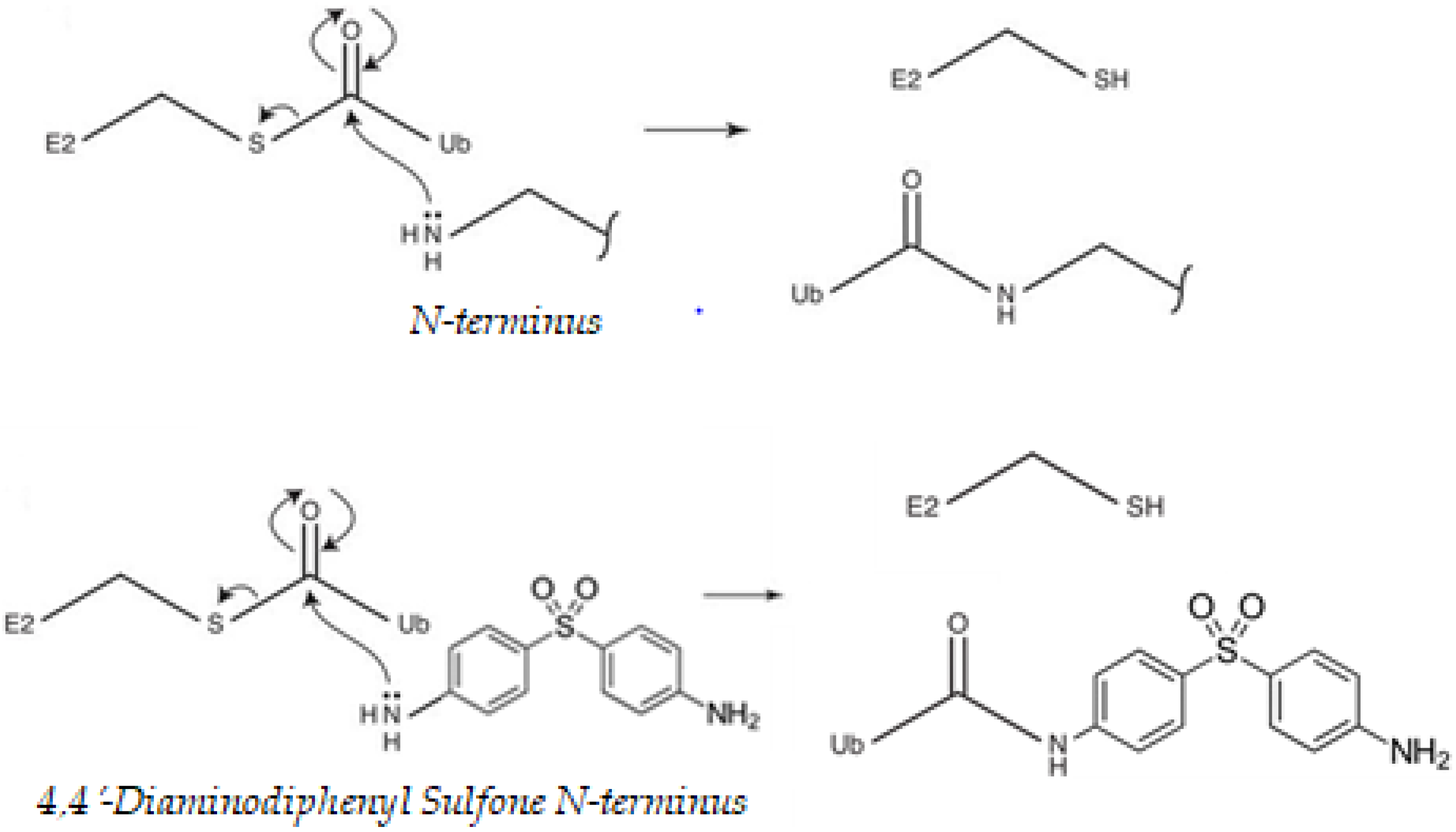
3.2. DDS Has Nucleophilic Sites for Ubiquitination
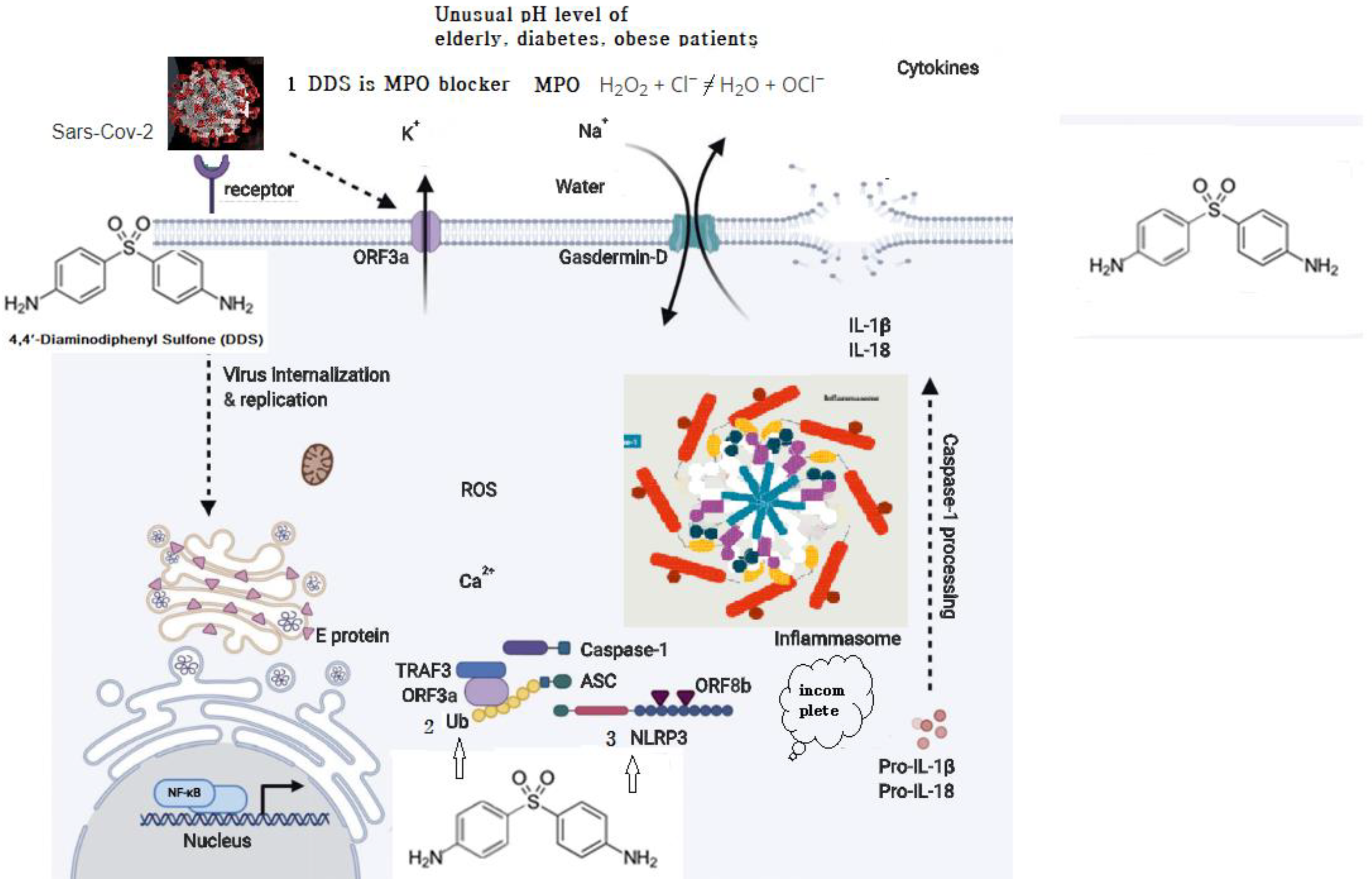
3.3. DDS Should Be Effective for Molecular Regulation of Inflammasome Activators
- Myeloperoxidase is a kind of oxidoreductase that catalyzes the chemical reaction of the following reaction: H2O2 + Cl− = H2O + OCl−. DDS binds to myeloperoxidase and regulates the production of hypochlorite, thereby reducing the inflammatory response of cells.
- Nucleophilic properties of DDS compete with Ub.
- Nucleophilic properties of DDS compete with NLRP3. ORF8b activates NLRP3 through direct interaction of the leucine-rich repeat domain of NLRP3.
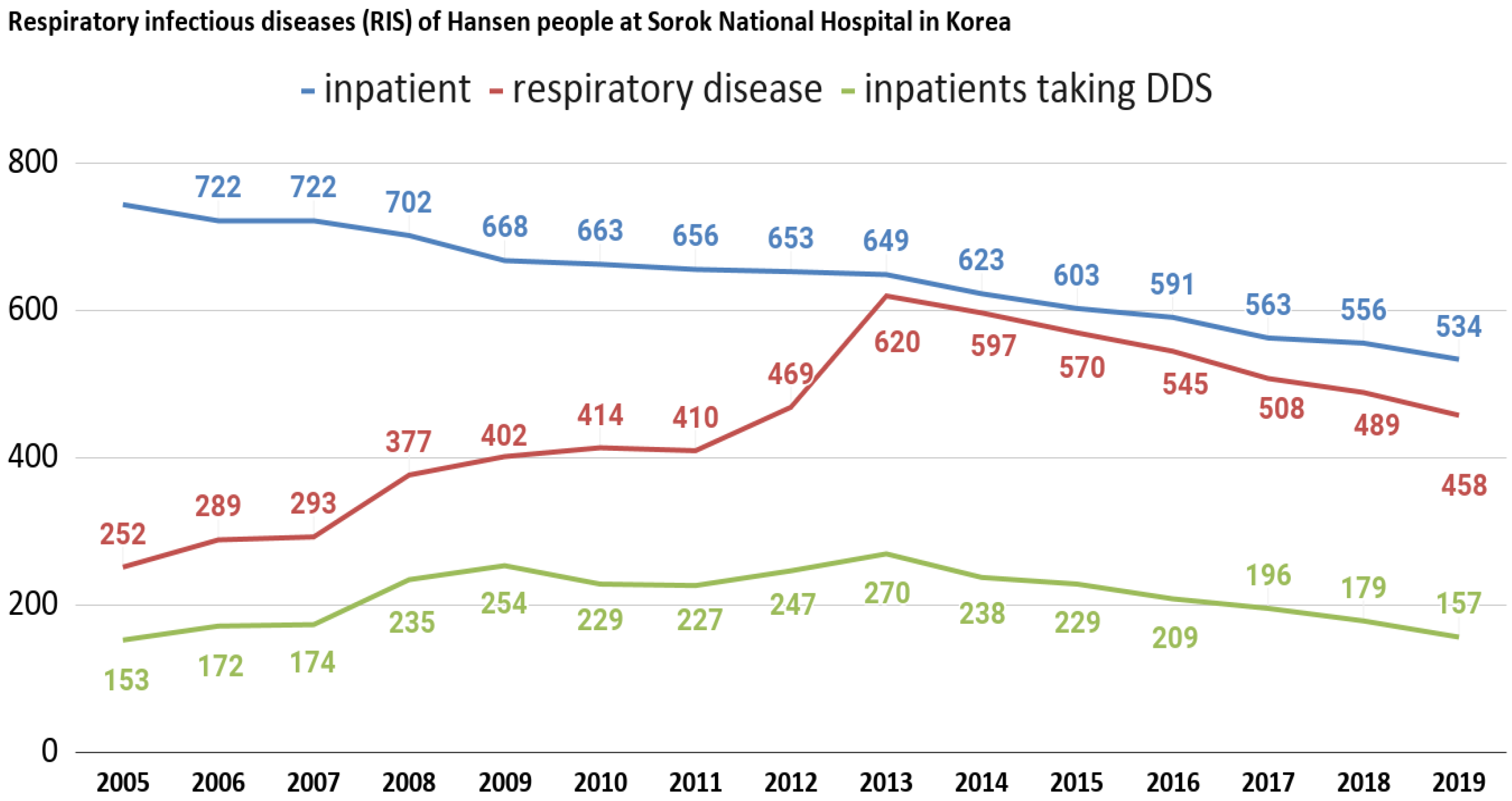
3.4. General Profiles of Leprosy-Affected Elderly in Korea
3.5. A Prospective Cohort Study for Neuroinflammation
4. Discussion
4.1. This Special Issue on “Myeloperoxidase”
4.2. Molecular Regulation of Inflammasome Activators
4.3. Myeloperoxidase and Red Blood Cells’ Deformity
4.4. Peroxymonocarbonate and Neurotoxicity
4.5. DDS Performs Various Anti-Inflammatory Reactions
4.6. Korean TLR2Arg677Trp Variation
4.7. COVID-19 Is a Generalized Systemic Immune Disease
4.8. Other Anti-Inflammatory Reactants
4.9. The Evidence of Viral Virulence Inhibition by DDS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| DDS | 4,4′-Diaminodiphenyl sulfone (dapsone) |
| LL | Lepromatous leprosy |
| MADDS | Monoacetyldapsone |
| MCI | Mild cognitive impairment |
| PD | Parkinson’s disease |
| NLRP3 | NOD-, LRR- and pyrin domains-containing protein 3 |
| MPO | Myeloperoxidase |
| TLR | Toll-like receptor |
References
- Mahase, E. Coronavirus: Covid-19 has killed more people than SARS and MERS combined, despite lower case fatality rate. BMJ 2020. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.L.; Baric, R.S. Recombination, reservoirs, and the modular spike: Mechanisms of coronavirus cross-species transmission. J. Virol. 2010, 84, 3134–3146. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Yuan, F.F.; Boehm, I.; Chan, P.K.S.; Marks, K.; Tang, J.W.; Hui, D.S.C.; Sung, J.J.Y.; Dyer, W.B.; Geczy, A.F.; Sullivan, J.S. High Prevalence of the CD14-159CC Genotype in Patients Infected with Severe Acute Respiratory Syndrome-Associated Coronavirus. Clin. Vaccine Immunol. 2007, 14, 1644–1645. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.; Ross, R.; Frydas, I.; Kritas, S. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by COVID-19: Anti-inflammatory strategies. J. Biol. Regul. Homeost Agents 2020, 34, 1. [Google Scholar] [PubMed]
- Ichinohe, T.; Yamazaki, T.; Koshiba, T.; Yanagi, Y. Mitochondrial protein mitofusin 2 is required for NLRP3 inflammasome activation after RNA virus infection. Proc. Natl. Acad. Sci. USA 2013, 110, 17963–17968. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Juliana, C.; Hong, S.; Datta, P.; Hwang, I.; Fernandes-Alnemri, T.; Yu, J.-W.; Alnemri, E.S. The mitochondrial antiviral protein MAVS associates with NLRP3 and regulates its inflammasome activity. J. Immunol. 2013, 191, 4358–4366. [Google Scholar] [CrossRef]
- Friker, L.L.; Scheiblich, H.; Hochheiser, I.V.; Brinkschulte, R.; Riedel, D.; Latz, E.; Geyer, M.; Heneka, M.T. β-Amyloid Clustering around ASC Fibrils Boosts Its Toxicity in Microglia. Cell Rep. 2020, 30, 3743–3754.e3746. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P.-Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Lowe, J. Treatment of leprosy with diamino-diphenyl sulphone by mouth. Lancet 1950, 258, 145–150. [Google Scholar] [CrossRef]
- Zhu, Y.I.; Stiller, M.J. Dapsone and sulfones in dermatology: Overview and update. J. Am. Acad. Dermatol. 2001, 45, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Wozel, V.G. Innovative use of dapsone. Dermatol. Clin. 2010, 28, 599–610. [Google Scholar] [CrossRef]
- Zhang, F.R.; Liu, H.; Irwanto, A.; Fu, X.A.; Li, Y.; Yu, G.Q.; Yu, Y.X.; Chen, M.F.; Low, H.Q.; Li, J.H.; et al. HLA-B*13:01 and the Dapsone Hypersensitivity Syndrome. N. Engl. J. Med. 2013, 369, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.; Lorenz, M.; Wozel, G. Hypersensitivity Reactions to Dapsone: A Systematic Review. Acta Derm. Venereol. 2012, 92, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; Chung, Y.-S.; Jo, H.J.; Lee, N.-J.; Kim, M.S.; Woo, S.H.; Park, S.; Kim, J.W.; Kim, H.M.; Han, M.-G. Identification of Coronavirus Isolated from a Patient in Korea with COVID-19. Osong Public Health Res. Perspect. 2020, 11, 3–7. [Google Scholar] [CrossRef]
- Bouaziz, J.D.; Duong, T.A.; Jachiet, M.; Velter, C.; Lestang, P.; Cassius, C.; Arsouze, A.; Domergue Than Trong, E.; Bagot, M.; Begon, E.; et al. Vascular skin symptoms in COVID-19: A French observational study. J. Eur. Acad. Dermatol. Venereol. 2020. [Google Scholar] [CrossRef]
- Rajendran, N.D.; Natarajan Mookan, I.S.; Mookan, S.B.; Munusamy, G.; Gurudeeban, S.; Kaliamurthi, S. A theoretical study of chemical bonding and topological and electrostatic properties of the anti-leprosy drug dapsone. J. Mol. Modeling 2020, 26. [Google Scholar] [CrossRef] [PubMed]
- Parr, R.G. Density functional theory of atoms and molecules. In Horizons of Quantum Chemistry; Springer: Berlin/Heidelberg, Germany, 1980; pp. 5–15. [Google Scholar]
- Cho, Y.; Shim, E.; Lee, K.-S.; Park, S.C. Mortality profiles of leprosy-affected elderly in Korea: A demographic perspective. Asia Pac. E J. Health Soc. Sci. 2014, 3, 1–5. Available online: https://sites.google.com/site/asiapacificejournalofhss/journal-issues/june-2014 (accessed on 18 August 2020).
- Ahn, Y.-H.; Park, H.; Kweon, S.-S. Causes of Death among Persons Affected by Leprosy in Korea, 2010–2013. Am. J. Trop. Med. Hyg. 2020, 102, 42–47. [Google Scholar] [CrossRef]
- Lee, J.; Choi, S.; Lee, C.J.; Oh, S. Supplementary Material for: Recovery of Dementia Syndrome following Treatment of Brain Inflammation. Dement. Geriatr. Cogn. Disord. EXTRA 2020, 10, 1–12. [Google Scholar] [CrossRef]
- JH, R.; Smith, T. Increased incidence in leprosy of hypersensitivity reactions to dapsone after introduction of multidrug therapy. Lepr. Rev. 1989, 60, 267–273. [Google Scholar]
- Alves-Rodrigues, E.N.; Ribeiro, L.C.; Silva, M.D.; Takiuchi, A.; Fontes, C.J.F. Dapsone syndrome with acute renal failure during leprosy treatment: Case report. Braz. J. Infect. Dis. 2005, 9, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Singhal, T. A Review of Coronavirus Disease-2019 (COVID-19). Indian J. Pediatr. 2020, 87, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Hu, W.; Niu, L.; Liu, H.; Xu, H.; Xiao, S.-Y. Pulmonary pathology of early phase 2019 novel coronavirus (COVID-19) pneumonia in two patients with lung cancer. J. Thorac. Oncol. 2020, 15, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Frater, J.L.; Zini, G.; D’Onofrio, G.; Rogers, H.J. COVID-19 and the clinical hematology laboratory. Int. J. Lab. Hematol. 2020, 42, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Bangash, M.N.; Patel, J.; Parekh, D. COVID-19 and the liver: Little cause for concern. Lancet Gastroenterol. Hepatol. 2020, 5, 529–530. [Google Scholar] [CrossRef]
- Liu, F.; Long, X.; Zou, W.; Fang, M.; Wu, W.; Li, W.; Zhang, B.; Zhang, W.; Chen, X.; Zhang, Z. Highly ACE2 Expression in Pancreas May Cause Pancreas Damage after SARS-CoV-2 Infection. medRxiv 2020. [Google Scholar] [CrossRef]
- Zhu, K.; He, F.; Jin, N.; Lou, J.; Cheng, H. Complete atrioventricular block associated with dapsone therapy: A rare complication of dapsone-induced hypersensitivity syndrome. J. Clin. Pharm. Ther. 2009, 34, 489–492. [Google Scholar] [CrossRef]
- Ghishan, F.K. The sulfone syndrome complicated by pancreatitis and pleural effusion in an adolescent receiving dapsone for treatment of acne vulgaris. J. Pediatr. Gastroenterol. Nutr. 1998, 26, 103–105. [Google Scholar]
- Kissling, S.; Rotman, S.; Gerber, C.; Halfon, M.; Lamoth, F.; Comte, D.; Lhopitallier, L.; Sadallah, S.; Fakhouri, F. Collapsing glomerulopathy in a COVID-19 patient. Kidney Int. 2020, 98, 228–231. [Google Scholar] [CrossRef]
- Zheng, Y.-Y.; Ma, Y.-T.; Zhang, J.-Y.; Xie, X. COVID-19 and the cardiovascular system. Nat. Rev. Cardiol. 2020, 17, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Kim, H.I.; Kim, O.H.; Cha, K.C.; Kim, H.; Lee, K.H.; Hwang, S.O.; Cha, Y.S. Clinical outcomes of adverse cardiovascular events in patients with acute dapsone poisoning. Clin. Exp. Emerg. Med. 2016, 3, 41. [Google Scholar] [CrossRef] [PubMed]
- Oxley, T.J.; Mocco, J.; Majidi, S.; Kellner, C.P.; Shoirah, H.; Singh, I.P.; De Leacy, R.A.; Shigematsu, T.; Ladner, T.R.; Yaeger, K.A.; et al. Large-Vessel Stroke as a Presenting Feature of Covid-19 in the Young. N. Engl. J. Med. 2020, 382, e60. [Google Scholar] [CrossRef] [PubMed]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 382, 2268–2270. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E. Dapsone as treatment adjunct in ARDS. Exp. Lung Res. 2020, 46, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Choi, S.-H.; Lee, C.J.; Oh, S.-S. Recovery of Dementia Syndrome following Treatment of Brain Inflammation. Dement. Geriatr. Cogn. Disord. Extra 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Moura, S.L.; Fernandes, G.F.S.; Machado, F.B.C.; Ferrão, L.F.A. Theoretical and experimental electronic spectra of neutral, monoprotonated and diprotonated dapsone. Theor. Chem. Acc. 2020, 139, 1–12. [Google Scholar] [CrossRef]
- Mendes, A.P.; Schalcher, T.R.; Barros, T.G.; Almeida, E.D.; Maia, C.S.; Barros, C.A.; Monteiro, M.C.; Borges, R.S. A Geometric and Electronic Study of Dapsone. J. Comput. Theor. Nanosci. 2011, 8, 1428–1431. [Google Scholar] [CrossRef]
- McDowell, G.S.; Philpott, A. Non-canonical ubiquitylation: Mechanisms and consequences. Int. J. Biochem. Cell. Biol. 2013, 45, 1833–1842. [Google Scholar] [CrossRef]
- McClellan, A.J.; Laugesen, S.H.; Ellgaard, L. Cellular functions and molecular mechanisms of non-lysine ubiquitination. Open Biol. 2019, 9, 190147. [Google Scholar] [CrossRef]
- Hyne, J.B.; Greidanus, J.W. Nuclear magnetic resonance study of intramolecular electronic effects in diphenyl sulfides, sulfoxides, and sulfones. Can. J. Chem. 1969, 47, 803–812. [Google Scholar] [CrossRef]
- Foley, J.F. Serine ubiquitylation. Sci. Signal. 2016, 9, ec303. [Google Scholar] [CrossRef]
- Kume, K.; Iizumi, Y.; Shimada, M.; Ito, Y.; Kishi, T.; Yamaguchi, Y.; Handa, H. Role of N-end rule ubiquitin ligases UBR1 and UBR2 in regulating the leucine-mTOR signaling pathway. Genes Cells 2010, 15, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Simmons, G.; Reeves, J.D.; Rennekamp, A.J.; Amberg, S.M.; Piefer, A.J.; Bates, P. Characterization of severe acute respiratory syndrome-associated coronavirus (SARS-CoV) spike glycoprotein-mediated viral entry. Proc. Natl. Acad. Sci. USA 2004, 101, 4240–4245. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Ducatelle, R.; Hoorens, J. Significance of lysosomes in the morphogenesis of coronaviruses. Arch. Virol. 1984, 79, 1–12. [Google Scholar] [CrossRef]
- Bozeman, P.M.; Learn, D.B.; Thomas, E.L. Inhibition of the human leukocyte enzymes myeloperoxidase and eosinophil peroxidase by dapsone. Biochem. Pharmacol. 1992, 44, 553–563. [Google Scholar] [CrossRef]
- Van Zyl, J.M.; Basson, K.; Kriegler, A.; van der Walt, B.J. Mechanisms by which clofazimine and dapsone inhibit the myeloperoxidase system: A possible correlation with their anti-inflammatory properties. Biochem. Pharmacol. 1991, 42, 599–608. [Google Scholar] [CrossRef]
- Uetrecht, J.P.; Shear, N.H.; Zahid, N. N-chlorination of sulfamethoxazole and dapsone by the myeloperoxidase system. Drug Metab. Dispos. 1993, 21, 830–834. [Google Scholar]
- Kim, S.-K.; Lee, S.-B.; Kang, T.-J.; Chae, G.-T. Detection of gene mutations related with drug resistance inMycobacterium lepraefrom leprosy patients using Touch-Down (TD) PCR. FEMS Immunol. Med. Microbiol. 2003, 36, 27–32. [Google Scholar] [CrossRef]
- Kang, T.-J.; Chae, G.-T. Detection of Toll-like receptor 2 (TLR2) mutation in the lepromatous leprosy patients. FEMS Immunol. Med. Microbiol. 2001, 31, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Yap, J.K.Y.; Moriyama, M.; Iwasaki, A. Inflammasomes and Pyroptosis as Therapeutic Targets for COVID-19. J. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.-S.; Nabar, N.R.; Huang, N.-N.; Kehrl, J.H. SARS-Coronavirus Open Reading Frame-8b triggers intracellular stress pathways and activates NLRP3 inflammasomes. Cell Death Discov. 2019, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Chakraborty, A.; Panda, A.; Ghosh, R.; Biswas, A. DNA minor groove binding of a well known anti-mycobacterial drug dapsone: A spectroscopic, viscometric and molecular docking study. Arch. Biochem. Biophys. 2019, 665, 107. [Google Scholar] [CrossRef]
- Lee, K.H.; Park, J.H.; Kim, D.H.; Hwang, J.; Lee, G.; Hyun, J.S.; Heo, S.T.; Choi, J.H.; Kim, M.; Kim, M. Dapsone as a potential treatment option for Henoch-Schönlein Purpura (HSP). Med. Hypotheses 2017, 108, 42–45. [Google Scholar] [CrossRef]
- Ahmed, F. Epigenetics: Tales of adversity. Nature 2010, 468, S20. [Google Scholar] [CrossRef]
- Demoinet, E.; Li, S.; Roy, R. AMPK blocks starvation-inducible transgenerational defects in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2017, 114, E2689–E2698. [Google Scholar] [CrossRef]
- Yamaza, H.; Chiba, T.; Higami, Y.; Shimokawa, I. Lifespan extension by caloric restriction: An aspect of energy metabolism. Microsc. Res. Tech. 2002, 59, 325–330. [Google Scholar] [CrossRef]
- Gibson, M.; Rogers, C.; Murrell, D. Successful dapsone therapy in inherited Epidermolysis Bullosa. J. Eur. Acad. Dermatol. Venereol. 2020, 34. [Google Scholar] [CrossRef] [PubMed]
- Ghaoui, N.; Hanna, E.; Abbas, O.; Kibbi, A.G.; Kurban, M. Update on the use of dapsone in dermatology. Int. J. Dermatol. 2020, 59. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Ruiz, A.; Zavala, C.; Montes, S.; Ortiz-Plata, A.; Salgado-Ceballos, H.; Orozco-Suarez, S.; Nava-Ruiz, C.; Pérez-Neri, I.; Perez-Severiano, F.; Ríos, C. Antioxidant, antiinflammatory and antiapoptotic effects of dapsone in a model of brain ischemia/reperfusion in rats. J. Neurosci. Res. 2008, 86, 3410–3419. [Google Scholar] [CrossRef] [PubMed]
- Mahale, A.; Kumar, R.; Sarode, L.P.; Gakare, S.; Prakash, A.; Ugale, R.R. Dapsone prolong delayed excitotoxic neuronal cell death by interacting with proapoptotic/survival signaling proteins. J. Stroke Cerebrovasc. Dis. 2020, 29, 104848. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Li, L.; Li, Z.; Ni, C.; Cao, Y.; Liu, T.; Tian, M.; Chui, D.; Guo, X. Protective effect of dapsone on cognitive impairment induced by propofol involves hippocampal autophagy. Neurosci. Lett. 2017, 649, 5–92. [Google Scholar] [CrossRef] [PubMed]
- Ríos, C.; Orozco-Suarez, S.; Salgado-Ceballos, H.; Mendez-Armenta, M.; Nava-Ruiz, C.; Santander, I.; Barón-Flores, V.; Caram-Salas, N.; Diaz-Ruiz, A. Anti-apoptotic effects of dapsone after spinal cord injury in rats. Neurochem. Res. 2015, 40, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Namba, Y.; Kawatsu, K.; Izumi, S.; Ueki, A.; Ikeda, K. Neurofibrillary tangles and senile plaques in brain of elderly leprosy patients. Lancet 1992, 340, 978. [Google Scholar] [CrossRef]
- Kimura, T.; Goto, M. Existence of senile plaques in the brains of elderly leprosy patients. Lancet 1993, 342, 1364. [Google Scholar] [CrossRef]
- Chui, D.-H.; Tabira, T.; Izumi, S.; Koya, G.; Ogata, J. Decreased beta-amyloid and increased abnormal Tau deposition in the brain of aged patients with leprosy. Am. J. Pathol. 1994, 145, 771. [Google Scholar]
- Appleby, B.S.; Cummings, L.J. Discovering new treatments for Alzheimer’s disease by repurposing approved medications. Curr. Top. Med. Chem. 2013, 13, 2306–2327. [Google Scholar] [CrossRef]
- Bain, A. Alzheimer disease: Dapsone phase 2 trial results reported. Immune Netw. Ltd. Press Release 2002. [Google Scholar]
- Imbimbo, B.P.; Solfrizzi, V.; Panza, F. Are NSAIDs useful to treat Alzheimer’s disease or mild cognitive impairment? Front. Aging Neurosci. 2010, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Reilly, T.P.; Lash, L.H.; Doll, M.A.; Hein, D.W.; Woster, P.M.; Svensson, C.K. A role for bioactivation and covalent binding within epidermal keratinocytes in sulfonamide-induced cutaneous drug reactions. J. Investig. Dermatol. 2000, 114, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Hurst, J.K.; Barrette, W.C.; Michel, B.R.; Rosen, H. Hypochlorous acid and myeloperoxidase-catalyzed oxidation of iron-slfur clusters in bacterial respiratory dehydrogenases. Eur. J. Biochem. 1991, 202, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Posadas, S.; Pichler, W. Delayed drug hypersensitivity reactions-new concepts. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2007, 37, 989. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.C.S. Are hypersensitivity reactions to dapsone becoming more frequent? Lepr. Rev. 1988, 59. [Google Scholar] [CrossRef]
- Stepan, A.F.; Walker, D.P.; Bauman, J.; Price, D.A.; Baillie, T.A.; Kalgutkar, A.S.; Aleo, M.D. Structural alert/reactive metabolite concept as applied in medicinal chemistry to mitigate the risk of idiosyncratic drug toxicity: A perspective based on the critical examination of trends in the top 200 drugs marketed in the United States. Chem. Res. Toxicol. 2011, 24, 1345–1410. [Google Scholar] [CrossRef]
- Roy, M.; Pal, I.; Nath, A.K.; Dey, S.G. Peroxidase activity of heme bound amyloid β peptides associated with Alzheimer’s disease. Chem. Commun. 2020, 56, 4505–4518. [Google Scholar] [CrossRef]
- Kiko, T.; Nakagawa, K.; Satoh, A.; Tsuduki, T.; Furukawa, K.; Arai, H. Amyloid β Levels in Human Red Blood Cells. PLoS ONE 2012, 7, e49620. [Google Scholar] [CrossRef]
- Blair-Johnson, M.; Fiedler, T.; Fenna, R. Human myeloperoxidase: Structure of a cyanide complex and its interaction with bromide and thiocyanate substrates at 1.9 Å resolution. Biochemistry 2001, 40, 13990–13997. [Google Scholar] [CrossRef]
- Shamova, E.V.; Gorudko, I.V.; Grigorieva, D.V.; Sokolov, A.V.; Kokhan, A.U.; Melnikova, G.B.; Yafremau, N.A.; Gusev, S.A.; Sveshnikova, A.N.; Vasilyev, V.B. The effect of myeloperoxidase isoforms on biophysical properties of red blood cells. Mol. Cell. Biochem. 2020, 464, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Varadarajan, S.; Yatin, S.; Kanski, J.; Jahanshahi, F.; Butterfield, D.A. Methionine residue 35 is important in amyloid β-peptide-associated free radical oxidative stress. Brain Res. Bull. 1999, 50, 133–141. [Google Scholar] [CrossRef]
- Vogt, W. Oxidation of methionyl residues in proteins: Tools, targets, and reversal. Free Radic. Biol. Med. 1995, 18, 93–105. [Google Scholar] [CrossRef]
- Enache, T.A.; Oliveira-Brett, A.M. Alzheimer’s disease amyloid beta peptides in vitro electrochemical oxidation. Bioelectrochemistry 2017, 114, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Francioso, A.; Baseggio Conrado, A.; Blarzino, C.; Foppoli, C.; Montanari, E.; Dinarelli, S.; Giorgi, A.; Mosca, L.; Fontana, M. One-and Two-Electron Oxidations of β-Amyloid25-35 by Carbonate Radical Anion (CO3•−) and Peroxymonocarbonate (HCO4−): Role of Sulfur in Radical Reactions and Peptide Aggregation. Molecules 2020, 25, 961. [Google Scholar] [CrossRef]
- Lee, Y.-I.; Kang, H.; Ha, Y.W.; Chang, K.-Y.; Cho, S.-C.; Song, S.O.; Kim, H.; Jo, A.; Khang, R.; Choi, J.-Y. Diaminodiphenyl sulfone–induced parkin ameliorates age-dependent dopaminergic neuronal loss. Neurobiol. Aging 2016, 41, 1–10. [Google Scholar] [CrossRef]
- Kwon, M.-J.; Joo, H.-G. Dapsone modulates lipopolysaccharide-activated bone marrow cells by inducing cell death and down-regulating tumor necrosis factor-α production. J. Vet. Sci. 2018, 19, 744–749. [Google Scholar] [CrossRef]
- Rashidian, A.; Rashki, A.; Abdollahi, A.; Haddadi, N.-S.; Chamanara, M.; Mumtaz, F.; Dehpour, A.R. Dapsone reduced acetic acid-induced inflammatory response in rat colon tissue through inhibition of NF-kB signaling pathway. Immunopharmacol. Immunotoxicol. 2019, 41, 607–613. [Google Scholar] [CrossRef]
- Abe, M.; Shimizu, A.; Yokoyama, Y.; Takeuchi, Y.; Ishikawa, O. A possible inhibitory action of diaminodiphenyl sulfone on tumour necrosis factor-α production from activated mononuclear cells on cutaneous lupus erythematosus. Clin. Exp. Dermatol. Exp. Dermatol. 2008, 33, 759–763. [Google Scholar] [CrossRef]
- Sheibani, M.; Nezamoleslami, S.; Faghir-Ghanesefat, H.; hossein Emami, A.; Dehpour, A.R. Cardioprotective effects of dapsone against doxorubicin-induced cardiotoxicity in rats. Cancer Chemother. Pharmacol. 2020, 85, 563–571. [Google Scholar] [CrossRef]
- Riviere, G.R.; Riviere, K.; Smith, K. Molecular and immunological evidence of oral Treponema in the human brain and their association with Alzheimer’s disease. Oral Microbiol. Immunol. 2002, 17, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Kamer, A.R.; Dasanayake, A.P.; Craig, R.G.; Glodzik-Sobanska, L.; Bry, M.; De Leon, M.J. Alzheimer’s disease and peripheral infections: The possible contribution from periodontal infections, model and hypothesis. J. Alzheimers Dis. 2008, 13, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Leira, Y.; Dominguez, C.; Seoane, J.; Seoane-Romero, J.; Pías-Peleteiro, J.M.; Takkouche, B.; Blanco, J.; Aldrey, J.M. Is periodontal disease associated with Alzheimer’s disease? A systematic review with meta-analysis. Neuroepidemiology 2017, 48, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Dominy, S.S.; Lynch, C.; Ermini, F.; Benedyk, M.; Marczyk, A.; Konradi, A.; Nguyen, M.; Haditsch, U.; Raha, D.; Griffin, C. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 2019, 5, eaau3333. [Google Scholar] [CrossRef]
- Ebaid, D.; Crewther, S.G. Visual information processing in young and older adults. Front. Aging Neurosci. 2019, 11, 116. [Google Scholar] [CrossRef]
- Sakamoto, W.; Fujii, Y.; Kanehira, T.; Asano, K.; Izumi, H. A novel assay system for myeloperoxidase activity in whole saliva. Clin. Biochem. 2008, 41, 584–590. [Google Scholar] [CrossRef]
- Moon, S.-Y.; Joo, H.-G. Anti-inflammatory effects of 4,4′-diaminodiphenyl sulfone (dapsone) in lipopolysaccharide-treated spleen cells: Selective inhibition of inflammation-related cytokines. Korean J. Vet. Res. 2015, 55, 199–204. [Google Scholar] [CrossRef]
- Park, Y.H.; Remmers, E.F.; Lee, W.; Ombrello, A.K.; Chung, L.K.; Shilei, Z.; Stone, D.L.; Ivanov, M.I.; Loeven, N.A.; Barron, K.S. Ancient familial Mediterranean fever mutations in human pyrin and resistance to Yersinia pestis. Nat. Immunol. 2020, 1–11. [Google Scholar] [CrossRef]
- Calligaris, L.; Marchetti, F.; Tommasini, A.; Ventura, A. The efficacy of anakinra in an adolescent with colchicine-resistant familial Mediterranean fever. Eur. J. Pediatr. 2008, 167, 695–696. [Google Scholar] [CrossRef]
- Salehzadeh, F.; Jahangiri, S.; Mohammadi, E. Dapsone as an alternative therapy in children with familial Mediterranean fever. Iran. J. Pediatr. 2012, 22, 23. [Google Scholar]
- Blackwell, J.M.; Jamieson, S.E.; Burgner, D. HLA and infectious diseases. Clin. Microbiol. Rev. 2009, 22, 370–385. [Google Scholar] [CrossRef] [PubMed]
- Matzaraki, V.; Kumar, V.; Wijmenga, C.; Zhernakova, A. The MHC locus and genetic susceptibility to autoimmune and infectious diseases. Genome Biol. 2017, 18, 76. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.; Shao, C.; Huang, J.; Gan, J.; Huang, X.; Bucci, E.; Piacentini, M.; Ippolito, G.; Melino, G. COVID-19 infection: The perspectives on immune responses. Cell Death Differ. 2020, 27, 1451–1454. [Google Scholar] [CrossRef] [PubMed]
- Bochud, P.-Y.; Hawn, T.R.; Aderem, A. Cutting Edge: A Toll-Like Receptor 2 Polymorphism That Is Associated with Lepromatous Leprosy Is Unable to Mediate Mycobacterial Signaling. J. Immunol. 2003, 170, 3451–3454. [Google Scholar] [CrossRef]
- Dasu, M.R.; Devaraj, S.; Park, S.; Jialal, I. Increased Toll-Like Receptor (TLR) Activation and TLR Ligands in Recently Diagnosed Type 2 Diabetic Subjects. Diabetes Care 2010, 33, 861–868. [Google Scholar] [CrossRef]
- Gomes, T.A.; Leite, N.; Tureck, L.; de Souza, R.; Titski, A.; Milano-Gai, G.; Lazarotto, L.; da Silva, L.; Furtado-Alle, L. Association between Toll-like receptors (TLR) and NOD-like receptor (NLR) polymorphisms and lipid and glucose metabolism. Gene 2019, 685, 211. [Google Scholar] [CrossRef]
- Fukata, M.; Vamadevan, A.; Abreu, M. Toll-like receptors (TLRs) and Nod-like receptors (NLRs) in inflammatory disorders. Semin. Immunol. 2009, 21, 242–253. [Google Scholar] [CrossRef]
- Xiao, Y.; Yan, H.; Diao, H.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Mao, X.; Luo, Y.; Chen, D. Early Gut Microbiota Intervention Suppresses DSS-Induced Inflammatory Responses by Deactivating TLR/NLR Signalling in Pigs. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Endoh, M.; Kunishita, T.; Tabira, T. No effect of anti-leprosy drugs in the prevention of Alzheimer’s disease and β-amyloid neurotoxicity. J. Neurol. Sci. 1999, 165, 28–30. [Google Scholar] [CrossRef]
- Manjili, R.H.; Zarei, M.; Habibi, M.; Manjili, M.H. COVID-19 as an Acute Inflammatory Disease. J. Immunol. 2020, 205, 12–19. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-L.; Li, Z.-Y.; Bi, X.-Y.; Zhao, H.; Zhao, J.-J.; Zhou, J.-G.; Han, Y.; Huang, Z.; Zhang, Y.-F.; Cai, J.-Q. Human leukocyte antigen gene polymorphisms are associated with systemic inflammation in hepatitis B virus-related hepatocellular carcinoma. Cancer Manag. Res. 2018, 10, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Ko, J.-H.; Kim, Y.; Kim, Y.-J.; Kim, J.-M.; Chung, Y.-S.; Kim, H.M.; Han, M.-G.; Kim, S.Y.; Chin, B.S. Viral Load Kinetics of SARS-CoV-2 Infection in First Two Patients in Korea. J. Korean Med. Sci. 2020, 35. [Google Scholar] [CrossRef] [PubMed]
- Bester, J.; Matshailwe, C.; Pretorius, E. Simultaneous presence of hypercoagulation and increased clot lysis time due to IL-1β, IL-6 and IL-8. Cytokine 2018, 110, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, D.; Sperhake, J.-P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S. Autopsy findings and venous thromboembolism in patients with COVID-19: A prospective cohort study. Ann. Intern. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rubino, F.; Amiel, S.A.; Zimmet, P.; Alberti, G.; Bornstein, S.; Eckel, R.H.; Mingrone, G.; Boehm, B.; Cooper, M.E.; Chai, Z.; et al. New-Onset Diabetes in Covid-19. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.; Nilsson-Payant, B.E.; Gupta, V.; Wang, P.; Duan, X.; Tang, X.; Zhu, J.; Zhao, Z.; Jaffré, F.; et al. A Human Pluripotent Stem Cell-based Platform to Study SARS-CoV-2 Tropism and Model Virus Infection in Human Cells and Organoids. Cell Stem. Cell 2020. [Google Scholar] [CrossRef]
- Feldstein, L.R.; Rose, E.B.; Horwitz, S.M.; Collins, J.P.; Newhams, M.M.; Son, M.B.F.; Newburger, J.W.; Kleinman, L.C.; Heidemann, S.M.; Martin, A.A.; et al. Multisystem Inflammatory Syndrome in U.S. Children and Adolescents. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Oxford University. Low-Cost Dexamethasone Reduces Death by up to One Third in Hospitalised Patients with Severe Respiratory Complications of COVID-19; Juillet, G., Ed.; Oxford University: Oxford, UK, 2020. [Google Scholar]
- Shang, L.; Zhao, J.; Hu, Y.; Du, R.; Cao, B. On the use of corticosteroids for 2019-nCoV pneumonia. Lancet 2020, 395, 683. [Google Scholar] [CrossRef]
- Haberman, R.; Axelrad, J.; Chen, A.; Castillo, R.; Yan, D.; Izmirly, P.; Neimann, A.; Adhikari, S.; Hudesman, D.; Scher, J.U. Covid-19 in immune-mediated inflammatory diseases—Case series from New York. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Kivela, P. Paradigm Shift for COVID-19 Response: Identifying High-risk Individuals and Treating Inflammation. West. J. Emerg. Med. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Gordeuk, V.; Delanghe, J.; Langlois, M.; Boelaert, J. Iron status and the outcome of HIV infection: An overview. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2001, 20, 111. [Google Scholar] [CrossRef]
- Duval, X.; Clayette, P.; Dereuddre-Bosquet, N.; Fretier, P.; Martin, M.; Salmon-Céron, D.; Gras, G.; Vildé, J.; Dormont, D. Dapsone and HIV-1 replication in primary cultures of lymphocytes and monocyte-derived macrophages. Aids 1997, 11, 943–944. [Google Scholar]
- Salmon-Ceron, D.; Fontbonne, A.; Saba, J.; May, T.; Raffi, F.; Chidiac, C.; Patey, O.; Aboulker, J.; Schwartz, D.; Vilde, J. Lower survival in AIDS patients receiving dapsone compared with aerosolized pentamidine for secondary prophylaxis of Pneumocystis carinii pneumonia. J. Infect. Dis. 1995, 172, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Hughes, W.T. Use of Dapsone in the Prevention and Treatment of Pneumocystis carinii Pneumonia: A Review. Clin. Infect. Dis. 1998, 27, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.M.; Lefèvre, P.; Hovette, P.; Issifi, S.; Mongin, M. Dapsone for thrombocytopenic purpura related to human immunodeficiency virus infection. Am. J. Med. 1991, 90, 675–677. [Google Scholar] [CrossRef]
- Opravil, M.; Hirschel, B.; Lazzarin, A.; Heald, A.; Pechère, M.; Rüttimann, S.; Iten, A.; von Overbeck, J.; Oertle, D.; Praz, G.; et al. Once-Weekly Administration of Dapsone/Pyrimethamine vs. Aerosolized Pentamidine as Combined Prophylaxis for Pneumocystis carinii Pneumonia and Toxoplasmic Encephalitis in Human Immunodeficiency Virus-Infected Patients. Clin. Infect. Dis. 1995, 20, 531–541. [Google Scholar] [CrossRef]
- Boelaert, J.R.; Piette, J.; Weinberg, G.A.; Sappey, C.; Weinberg, E.D.; Rich, E.A.; Abbud, R.A. Iron and Oxidative Stress as a Mechanism for the Enhanced Production of Human Immunodeficiency Virus by Alveolar Macrophages from Otherwise Healthy Cigarette Smokers [with Reply]. J. Infect. Dis. 1996, 173, 1045–1047. [Google Scholar] [CrossRef]
- Drakesmith, H.; Prentice, A. Viral infection and iron metabolism. Nat. Rev. Microbiol. 2008, 6, 541. [Google Scholar] [CrossRef]
- Banjoko, S.O.; Oseni, F.A.; Togun, R.A.; Onayemi, O.; Emma-Okon, B.O.; Fakunle, J.B. Iron status in HIV-1 infection: Implications in disease pathology. BMC Clin. Pathol. 2012, 12, 26. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Clinical Manifestations | Sars-Cov-2 Symptoms | DDS Symptoms |
|---|---|---|
| skin vascular symptom slymphadenopathy fever | (i) violaceous macules with porcelain appearance, (ii) livedo of the trunk with chest pain and cough, (iii) violaceous macule and Raynaud’s phenomenon 10 days after fever and cough, (iv) necrotic purpura in a patient treated with leflunomide and systemic steroids for rheumatoid arthritis, (v) chilblain appearance and Raynaud’s phenomenon in a patient with anosmia, fever and cough, (vi) eruptive cherry angioma 21 days after COVID-19 healing of clinical symptoms [16] | “glandular fever”: (i) fever, (ii) lymphadenopathy, (iii) generalized rash, and (iv) hepatitis occurring after dapsone intake [22] rash, exanthema/erythema, erythroderma, mucosal involvement [14] papular or exfoliative dermatitis, generalized lymphadenopathy [23] |
| hypersensitivity reactions | SARS-CoV-2 symptoms are like a severe idiosyncratic DDS reaction characterized by the clinical triad of fever, rash, and systemic involvement, which can cause severe organ (heart, kidney, lung, brain, etc.) dysfunction [24] | the syndrome is a severe idiosyncratic DDS reaction characterized by the clinical triad of fever, rash, and systemic involvement (most commonly of the liver and the hematologic system), which can cause severe organ dysfunction [13] |
| hematology laboratory | focal fibrin clusters mixed with mononuclear inflammatory cells, decreased eosinophils, decreased lymphocytes, increased neutrophils [25] lymphopenia, leukocytosis, neutrophilia, thrombocytopenia [26] | leukocytosis, eosinophilia [14], resembling a mononucleosis infection [23] |
| anemia | thrombocytopenia, consumptive coagulopathy [26] | hemolytic anemia and methemoglobinemia [12] |
| liver disease, pancreatic disease | clinically significant liver injury is uncommon [27], pancreatic cells highly express ACE2 [28] | hepatitis/liver toxicity [23], cholangitis, colitis, thyroiditis [29], pancreatitis and pleural effusion [30] |
| renal disease | severe collapsing focal segmental glomerulosclerosis, acute tubular necrosis [31] | acute renal failure [23], |
| cardiac disease | acute myocardial injury and chronic damage to the cardiovascular system [32] | myocarditis, dapsone-induced hypersensitivity syndrome-associated complete atrioventricular block [29], myocardial injury [33] |
| pulmonary disease | coronavirus disease (COVID-19)-related pneumonia [24,32] | pneumonitis [29], pneumonia or multiple organ failure [33] |
| neurologic disease | large-vessel stroke [34] encephalopathy, prominent agitation and confusion, and corticospinal tract signs [35,36] | recovery of dementia syndrome following treatment of brain inflammation [37] |
| Cumulative Recurrence Risk Index | Infectious Respiratory Diseases | |||||
|---|---|---|---|---|---|---|
| Year | Inactive Patient | Number of Recurrent Patients | Annual Recurrence Rate (%) | DDS Intake (100 mg) | Infected Person | Remarks (Report) |
| 2002 SARS-CoV | 16,712 | 10 | 0.05 | -(0 person) | (no prevalence) | |
| 2003 | 16,283 | 6 | 0.03 | - | - | |
| 2004 | 15,797 | 6 | 0.03 | - | - | |
| 2005 | 15,350 | 5 | 0.03 | - | - | |
| 2006 | 14,851 | 5 | 0.03 | - | - | |
| 2007 | 14,321 | 3 | 0.02 | - | - | |
| 2008 | 13,875 | 3 | 0.02 | - | - | |
| 2009 Influenza A virus subtype H1N1 | 13,422 | 3 | 0.02 | -(0 person) | (no prevalence) | |
| 2010 | 13,033 | 7 | 0.05 | - | - | |
| 2011 | 12,582 | 3 | 0.02 | - | - | |
| 2012 | 12,068 | 1 | 0.01 | - | - | |
| 2013 | 11,595 | 1 | 0.01 | - | - | |
| 2014 | 11,104 | 1 | 0.01 | - | - | |
| 2015 MERS | 10,653 | 1 | 0.01 | -(0 person) | (no prevalence) | |
| 2016 | 10,236 | - | - | - | - | |
| 2017 | 9908 | - | - | - | - | |
| 2018 | 9503 | 1 | 0.01 | 2019–2020.6. 3814(person)/9134(total) | - | - |
| 2019 SARS-CoV-2 | 9160 | 2 | 0.02 | -(0 person) | (no prevalence) | |
| –2020. 6.30. SARS-CoV-2 | -(0 person) | (no prevalence) | ||||
| Total | Age | Average Age | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–19 | 20–39 | 40–59 | 60–79 | 80– | SUM | Treatment Place | |||||
| Home | Town | Nursing | Hospital | ||||||||
| total | 9288 (100%) | - | 18 (0.2%) | 545 (5.9%) | 5147 (55.4%) | 3578 (38.5%) | 79 | 78 | 80 | 81 | 78 |
| Active leprosy patients | 128 (100%) | - | 7 (5.5%) | 9 (7.0%) | 79 (61.7%) | 33 (25.8%) | 71 | 73 | 68 | 65 | 77 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-h.; An, H.K.; Sohn, M.-G.; Kivela, P.; Oh, S. 4,4′-Diaminodiphenyl Sulfone (DDS) as an Inflammasome Competitor. Int. J. Mol. Sci. 2020, 21, 5953. https://doi.org/10.3390/ijms21175953
Lee J-h, An HK, Sohn M-G, Kivela P, Oh S. 4,4′-Diaminodiphenyl Sulfone (DDS) as an Inflammasome Competitor. International Journal of Molecular Sciences. 2020; 21(17):5953. https://doi.org/10.3390/ijms21175953
Chicago/Turabian StyleLee, Jong-hoon, Ha Kyeu An, Mun-Gi Sohn, Paul Kivela, and Sangsuk Oh. 2020. "4,4′-Diaminodiphenyl Sulfone (DDS) as an Inflammasome Competitor" International Journal of Molecular Sciences 21, no. 17: 5953. https://doi.org/10.3390/ijms21175953
APA StyleLee, J.-h., An, H. K., Sohn, M.-G., Kivela, P., & Oh, S. (2020). 4,4′-Diaminodiphenyl Sulfone (DDS) as an Inflammasome Competitor. International Journal of Molecular Sciences, 21(17), 5953. https://doi.org/10.3390/ijms21175953