Regulation of GABAA Receptor Subunit Expression in Substance Use Disorders


Abstract
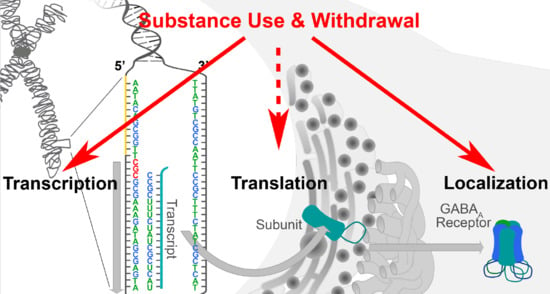
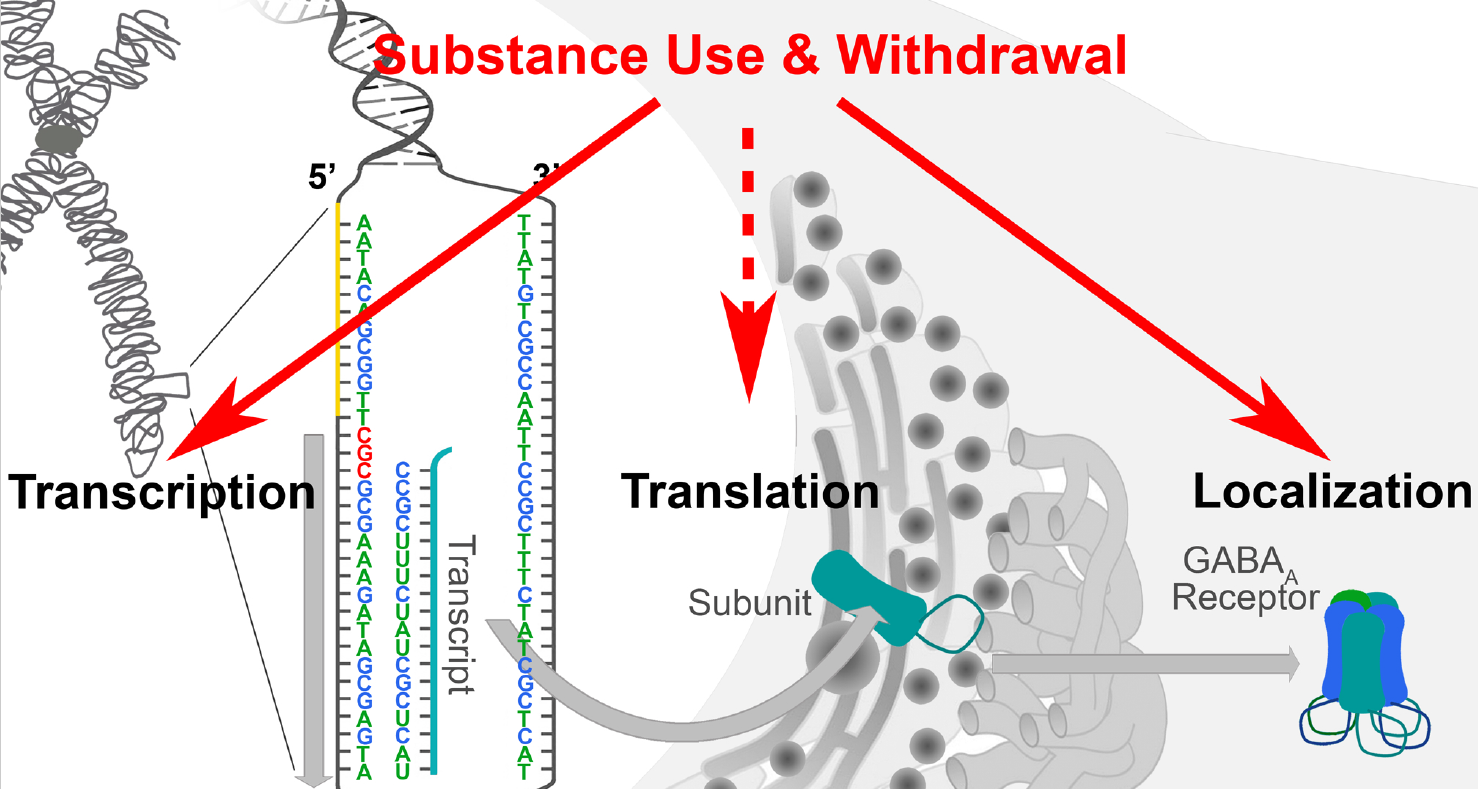
1. Introduction
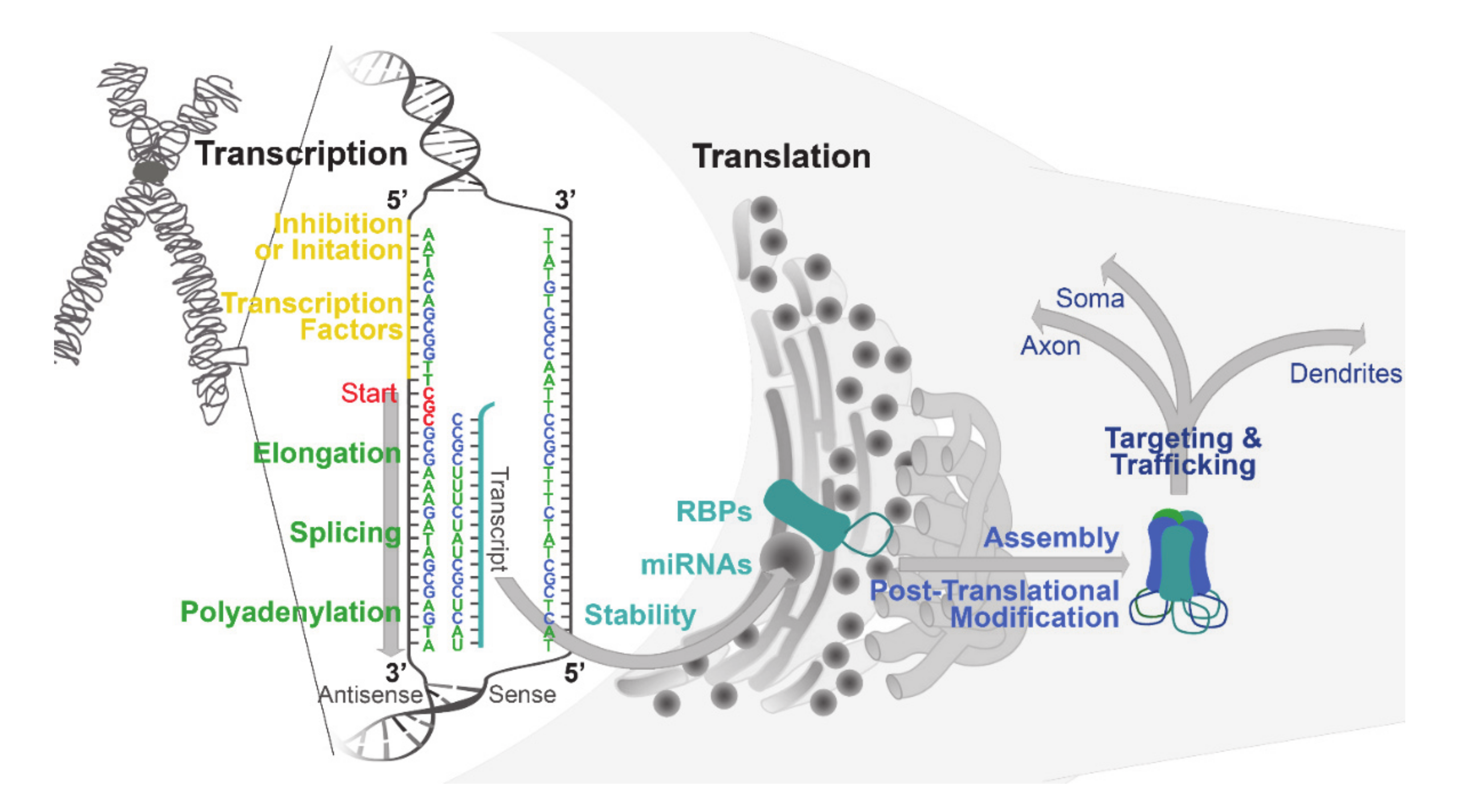
2. Basal Mechanisms Regulating GABAA Receptor Expression: Transcription, Translation, and Beyond
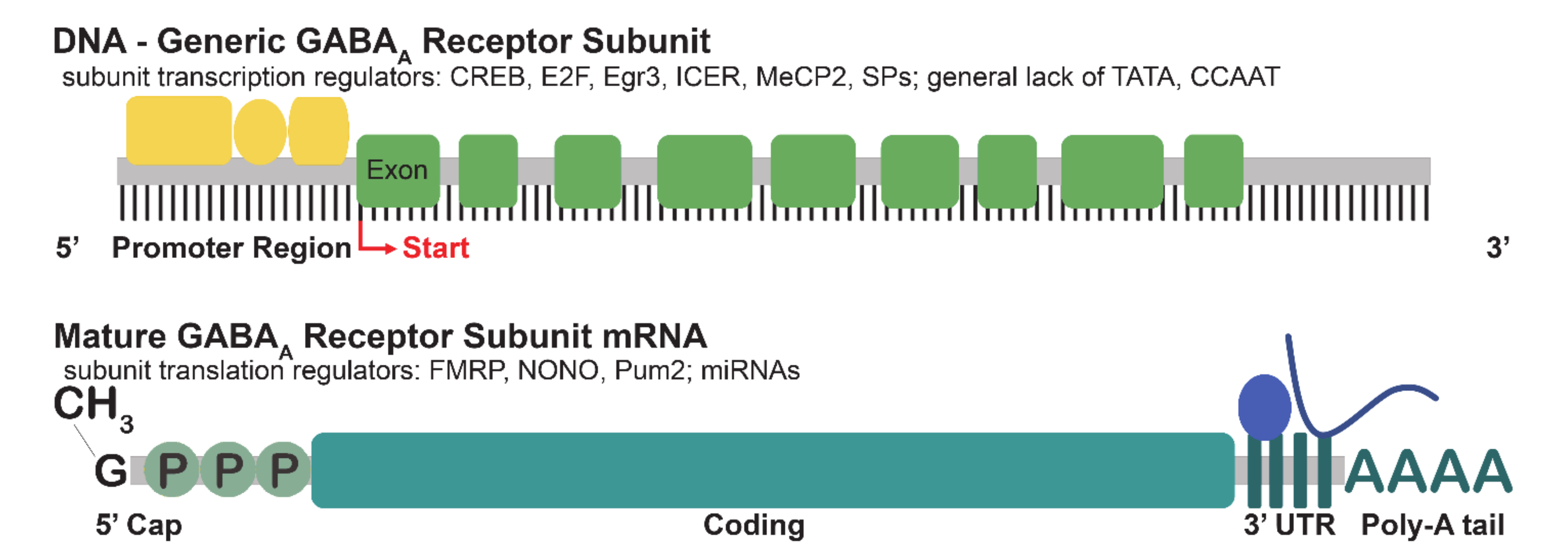
2.1. Transcription
2.2. Translation
3. Experience-Dependent Plasticity Regulates GABAA Receptor Subunit Expression
4. Substance Use Regulates GABAA Receptor Subunit Expression
4.1. Benzodiazepine Use
4.2. Alcohol Use
4.3. Use of Other Highly Abused Substances
4.4. Withdrawal
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Steiger, J.L.; Russek, S.J. GABAA receptors: Building the bridge between subunit mRNAs, their promoters, and cognate transcription factors. Pharmacol. Ther. 2004, 101, 259–281. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, W.; Fuchs, K.; Tretter, V.; Ebert, V.; Jechlinger, M.; Höger, H.; Adamiker, D. Structure and subunit composition of GABAA receptors. Neurochem. Int. 1999, 34, 379–385. [Google Scholar] [CrossRef]
- Backus, K.H.; Arigoni, M.; Drescher, U.; Scheurer, L.; Malherbe, P.; Möhler, H.; Benson, J.A. Stoichiometry of a recombinant GABAA receptor deduced from mutation-induced rectification. Neuroreport 1993, 5, 285–288. [Google Scholar] [CrossRef]
- Laurie, D.J.; Wisden, W.; Seeburg, P.H. The distribution of thirteen GABAA receptor subunit mRNAs in the rat brain. III. Embryonic and postnatal development. J. Neurosci. 1992, 12, 4151–4172. [Google Scholar] [CrossRef]
- Ganguly, K.; Schinder, A.F.; Wong, S.T.; Poo, M. GABA Itself Promotes the Developmental Switch of Neuronal GABAergic Responses from Excitation to Inhibition. Cell 2001, 105, 521–532. [Google Scholar] [CrossRef]
- Wisden, W.; Laurie, D.J.; Monyer, H.; Seeburg, P.H. The Distribution of 13 GABA, Receptor Subunit mRNAs in the Rat Brain. I. Telencephalon, Diencephalon, Mesencephalon. J. Neurosci. 1992, 12, 1040–1062. [Google Scholar] [CrossRef]
- Pirker, S.; Schwarzer, C.; Wieselthaler, A.; Sieghart, W.; Sperk, G. GABAA receptors: Immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience 2000, 101, 815–850. [Google Scholar] [CrossRef]
- Hörtnagl, H.; Tasan, R.O.; Wieselthaler, A.; Kirchmair, E.; Sieghart, W.; Sperk, G. Patterns of mRNA and protein expression for 12 GABAA receptor subunits in the mouse brain. Neuroscience 2013, 236, 345–372. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wu, Z.; Ning, G.; Guo, Y.; Ali, R.; Macdonald, R.L.; De Blas, A.L.; Luscher, B.; Chen, G. GABAA receptor alpha subunits play a direct role in synaptic versus extrasynaptic targeting. J. Biol. Chem. 2012, 287, 27417–27430. [Google Scholar] [CrossRef]
- Körber, C.; Richter, A.; Kaiser, M.; Schlicksupp, A.; Mükusch, S.; Kuner, T.; Kirsch, J.; Kuhse, J. Effects of distinct collybistin isoforms on the formation of GABAergic synapses in hippocampal neurons. Mol. Cell. Neurosci. 2012, 50, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Panzanelli, P.; Gunn, B.G.; Schlatter, M.C.; Benke, D.; Tyagarajan, S.K.; Scheiffele, P.; Belelli, D.; Lambert, J.J.; Rudolph, U.; Fritschy, J.-M. Distinct mechanisms regulate GABAA receptor and gephyrin clustering at perisomatic and axo-axonic synapses on CA1 pyramidal cells. J. Physiol. 2011, 589, 4959–4980. [Google Scholar] [CrossRef]
- Kneussel, M.; Brandstätter, J.H.; Gasnier, B.; Feng, G.; Sanes, J.R.; Betz, H. Gephyrin-independent clustering of postsynaptic GABA(A) receptor subtypes. Mol. Cell. Neurosci. 2001, 17, 973–982. [Google Scholar] [CrossRef]
- Eyre, M.D.; Renzi, M.; Farrant, M.; Nusser, Z. Setting the Time Course of Inhibitory Synaptic Currents by Mixing Multiple GABAA Receptor Subunit Isoforms. J. Neurosci. 2012, 32, 5853–5867. [Google Scholar] [CrossRef] [PubMed]
- Bright, D.P.; Renzi, M.; Bartram, J.; McGee, T.P.; MacKenzie, G.; Hosie, A.M.; Farrant, M.; Brickley, S.G. Profound desensitization by ambient GABA limits activation of δ-containing GABAA receptors during spillover. Version 2. J. Neurosci. 2011, 31, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Picton, A.J.; Fisher, J.L. Effect of the alpha subunit subtype on the macroscopic kinetic properties of recombinant GABA(A) receptors. Brain Res. 2007, 1165, 40–49. [Google Scholar] [CrossRef]
- Bianchi, M.T.; Haas, K.F.; Macdonald, R.L. α1 and α6 subunits specify distinct desensitization, deactivation and neurosteroid modulation of GABAA receptors containing the δ subunit. Neuropharmacology 2002, 43, 492–502. [Google Scholar] [CrossRef]
- Mortensen, M.; Patel, B.; Smart, T.G. GABA Potency at GABAA Receptors Found in Synaptic and Extrasynaptic Zones. Front. Cell. Neurosci. 2012, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schofield, C.M.; Huguenard, J.R. GABA Affinity Shapes IPSCs in Thalamic Nuclei. J. Neurosci. 2007, 27, 7954–7962. [Google Scholar] [CrossRef] [PubMed]
- Keramidas, A.; Harrison, N.L. The activation mechanism of α1β2γ2S and α3β3γ2S GABAA receptors. J. Gen. Physiol. 2010, 135, 59–75. [Google Scholar] [CrossRef]
- Dixon, C.; Sah, P.; Lynch, J.W.; Keramidas, A. GABAA Receptor α and γ Subunits Shape Synaptic Currents via Different Mechanisms. J. Biol. Chem. 2014, 289, 5399–5411. [Google Scholar] [CrossRef]
- Mizukami, K.; Grayson, D.R.; Ikonomovic, M.D.; Sheffield, R.; Armstrong, D.M. GABAA receptor β2 and β3 subunits mRNA in the hippocampal formation of aged human brain with Alzheimer-related neuropathology. Mol. Brain Res. 1998, 56, 268–272. [Google Scholar] [CrossRef]
- Ali, N.J.; Olsen, R.W. Chronic benzodiazepine treatment of cells expressing recombinant GABAA receptors uncouples allosteric binding: Studies on possible mechanisms. J. Neurochem. 2001, 79, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Russek, S.J. Evolution of GABAA receptor diversity in the human genome. Gene 1999, 227, 213–222. [Google Scholar] [CrossRef]
- Brooks-Kayal, A.R.; Shumate, M.D.; Jin, H.; Lin, D.D.; Rikhter, T.Y.; Holloway, K.L.; Coulter, D.A. Human Neuronal γ-Aminobutyric AcidA Receptors: Coordinated Subunit mRNA Expression and Functional Correlates in Individual Dentate Granule Cells. J. Neurosci. 1999, 19, 8312–8318. [Google Scholar] [CrossRef] [PubMed]
- Buckle, V.J.; Fujita, N.; Ryder-Cook, A.S.; Derry, J.M.; Barnard, P.J.; Lebo, R.V.; Schofield, P.R.; Seeburg, P.H.; Bateson, A.N.; Darlison, M.G.; et al. Chromosomal localization of GABAA receptor subunit genes: Relationship to human genetic disease. Neuron 1989, 3, 647–654. [Google Scholar] [CrossRef]
- McLean, P.J.; Farb, D.H.; Russek, S.J. Mapping of the alpha 4 subunit gene (GABRA4) to human chromosome 4 defines an alpha 2-alpha 4-beta 1-gamma 1 gene cluster: Further evidence that modern GABAA receptor gene clusters are derived from an ancestral cluster. Genomics 1995, 26, 580–586. [Google Scholar] [CrossRef]
- Wilcox, A.S.; Warrington, J.A.; Gardiner, K.; Berger, R.; Whiting, P.; Altherr, M.R.; Wasmuth, J.J.; Patterson, D.; Sikela, J.M. Human chromosomal localization of genes encoding the gamma 1 and gamma 2 subunits of the gamma-aminobutyric acid receptor indicates that members of this gene family are often clustered in the genome. Proc. Natl. Acad. Sci. USA 1992, 89, 5857–5861. [Google Scholar] [CrossRef]
- Lemon, B.; Tjian, R. Orchestrated response: A symphony of transcription factors for gene control. Genes Dev. 2000, 14, 2551–2569. [Google Scholar] [CrossRef]
- Joyce, C.J. In silico comparative genomic analysis of GABAA receptor transcriptional regulation. BMC Genomics 2007, 8, 203. [Google Scholar] [CrossRef]
- Bateson, A.N.; Ultsch, A.; Darlison, M.G. Isolation and sequence analysis of the chicken GABAA receptor α1-subunit gene promoter. Gene 1995, 153, 243–247. [Google Scholar] [CrossRef]
- Nair, B.; Johar, K.; Priya, A.; Wong-Riley, M.T.T. Specificity protein 4 (Sp4) transcriptionally regulates inhibitory GABAergic receptors in neurons. Biochim. Biophys. Acta 2016, 1863, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Brooks-Kayal, A.R.; Russek, S.J. Regulation of GABAA Receptor Gene Expression and Epilepsy. In Jasper’s Basic Mechanisms of the Epilepsies; Noebels, J.L., Avoli, M., Rogawski, M.A., Olsen, R.W., Delgado-Escueta, A.V., Eds.; National Center for Biotechnology Information: Bethesda, MD, USA, 2012. [Google Scholar]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef]
- Kandel, E.R. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain 2012, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Carlezon, W.A., Jr.; Duman, R.S.; Nestler, E.J. The many faces of CREB. Trends Neurosci. 2005, 28, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.C.; Kyzar, E.J.; Zhang, H. Epigenetic Basis of the Dark Side of Alcohol Addiction. Neuropharmacology 2017, 122, 74–84. [Google Scholar] [CrossRef]
- Fuchs, K.; Celepirovic, N. The 5′-flanking region of the rat GABAA receptor α2-subunit gene (Gabra2). J. Neurochem. 2002, 82, 1512–1523. [Google Scholar] [CrossRef]
- Hadingham, K.L.; Wingrove, P.; Le Bourdelles, B.; Palmer, K.J.; I Ragan, C.; Whiting, P.J. Cloning of cDNA sequences encoding human alpha 2 and alpha 3 gamma-aminobutyric acidA receptor subunits and characterization of the benzodiazepine pharmacology of recombinant alpha 1-, alpha 2-, alpha 3-, and alpha 5-containing human gamma-aminobutyric acidA receptors. Mol. Pharmacol. 1993, 43, 970–975. [Google Scholar]
- Mu, W.; Burt, D.R. The mouse GABAA receptor α3 subunit gene and promoter. Mol. Brain Res. 1999, 73, 172–180. [Google Scholar] [CrossRef]
- De Groen, P.C.; Eggen, B.J.; Gispen, W.H.; Schotman, P.; Schrama, L.H. Cloning and promoter analysis of the human B-50/GAP-43 gene. J. Mol. Neurosci. 1995, 6, 109–119. [Google Scholar] [CrossRef]
- Ma, L.; Song, L.; Radoi, G.E.; Harrison, N.L. Transcriptional regulation of the mouse gene encoding the alpha-4 subunit of the GABAA receptor. J. Biol. Chem. 2004, 279, 40451–40461. [Google Scholar] [CrossRef]
- Roberts, D.S.; Raol, Y.H.; Bandyopadhyay, S.; Lund, I.V.; Budreck, E.C.; Passini, M.J.; Wolfe, J.H.; Brooks-Kayal, A.R.; Russek, S.J. Egr3 stimulation of GABRA4 promoter activity as a mechanism for seizure-induced up-regulation of GABA(A) receptor alpha4 subunit expression. Proc. Natl. Acad. Sci. USA 2005, 102, 11894–11899. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.V.; Cole, A.J.; Baraban, J.M. Role of monoamine systems in activation of zif268 by cocaine. J. Psychiatry Neurosci. 1992, 17, 94–102. [Google Scholar] [PubMed]
- Blackwood, C.A.; McCoy, M.T.; Ladenheim, B.; Cadet, J.L. Escalated Oxycodone Self-Administration and Punishment: Differential Expression of Opioid Receptors and Immediate Early Genes in the Rat Dorsal Striatum and Prefrontal Cortex. Front. Neurosci. 2019, 13, 1392. [Google Scholar] [CrossRef]
- Wang, W.; Stock, R.E.; Gronostajski, R.M.; Wong, Y.W.; Schachner, M.; Kilpatrick, D.L. A role for nuclear factor I in the intrinsic control of cerebellar granule neuron gene expression. J. Biol. Chem. 2004, 279, 53491–53497. [Google Scholar] [CrossRef] [PubMed]
- McLean, P.J.; Shpektor, D.; Bandyopadhyay, S.; Russek, S.J.; Farb, D.H. A minimal promoter for the GABA(A) receptor alpha6-subunit gene controls tissue specificity. J. Neurochem. 2000, 74, 1858–1869. [Google Scholar] [CrossRef] [PubMed]
- Cholewa-Waclaw, J.; Shah, R.; Webb, S.; Chhatbar, K.; Ramsahoye, B.; Pusch, O.; Yu, M.; Greulich, P.; Waclaw, B.; Bird, A. Quantitative modelling predicts the impact of DNA methylation on RNA polymerase II traffic. Proc. Natl. Acad. Sci. USA 2019, 116, 14995–15000. [Google Scholar] [CrossRef]
- Chahrour, M.; Jung, S.Y.; Shaw, C.A.; Zhou, X.; Wong, S.T.C.; Qin, J.; Zoghbi, H.Y. MeCP2, a Key Contributor to Neurological Disease, Activates and Represses Transcription. Science 2008, 320, 1224–1229. [Google Scholar] [CrossRef]
- Kozinetz, C.A.; Skender, M.L.; Macnaughton, N.; Almes, M.J.; Schultz, R.J.; Percy, A.K.; Glaze, D.G. Epidemiology of Rett syndrome: A population-based registry. Pediatrics 1993, 91, 445–450. [Google Scholar]
- Zhang, Z.-W.; Zak, J.D.; Liu, H. MeCP2 Is Required for Normal Development of GABAergic Circuits in the Thalamus. J. Neurophysiol. 2010, 103, 2470–2481. [Google Scholar] [CrossRef][Green Version]
- Samaco, R.C.; Hogart, A.; LaSalle, J.M. Epigenetic overlap in autism-spectrum neurodevelopmental disorders: MECP2 deficiency causes reduced expression of UBE3A and GABRB3. Hum. Mol. Genet. 2005, 14, 483–492. [Google Scholar] [CrossRef]
- Medrihan, L.; Tantalaki, E.; Aramuni, G.; Sargsyan, V.; Dudanova, I.; Missler, M.; Zhang, W. Early defects of GABAergic synapses in the brain stem of a MeCP2 mouse model of Rett syndrome. J. Neurophysiol. 2008, 99, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Oyarzabal, A.; Xiol, C.; Castells, A.A.; Grau, C.; O’Callaghan, M.; Fernández, G.; Alcántara, S.; Pineda, M.; Armstrong, J.; Altafaj, X.; et al. Comprehensive Analysis of GABAA-A1R Developmental Alterations in Rett Syndrome: Setting the Focus for Therapeutic Targets in the Time Frame of the Disease. Int. J. Mol. Sci. 2020, 21, 518. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.E. Autism Spectrum Disorders and Drug Addiction: Common Pathways, Common Molecules, Distinct Disorders? Front Neurosci. 2016, 10, 20. [Google Scholar] [CrossRef]
- Schieweck, R.; Kiebler, M.A. Posttranscriptional Gene Regulation of the GABA Receptor to Control Neuronal Inhibition. Front. Mol. Neurosci. 2019, 12, 152. [Google Scholar] [CrossRef]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The Mechanism of Eukaryotic Translation Initiation and Principles of its Regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y.; Dohmae, N.; Hirokawa, N. Kinesin Transports RNA: Isolation and Characterization of an RNA-Transporting Granule. Neuron 2004, 43, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Fritzsche, R.; Karra, D.; Bennett, K.L.; Ang, F.Y.; Heraud-Farlow, J.E.; Tolino, M.; Doyle, M.; Bauer, K.E.; Thomas, S.; Planyavsky, M.; et al. Interactome of Two Diverse RNA Granules Links mRNA Localization to Translational Repression in Neurons. Cell Rep. 2013, 5, 1749–1762. [Google Scholar] [CrossRef]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Andreassi, C.; Riccio, A. To localize or not to localize: mRNA fate is in 3′UTR ends. Trends Cell Biol. 2009, 19, 465–474. [Google Scholar] [CrossRef]
- Floor, S.N.; Doudna, J.A. Tunable protein synthesis by transcript isoforms in human cells. Elife 2016, 5, e10921. [Google Scholar] [CrossRef]
- Blair, J.D.; Hockemeyer, D.; Doudna, J.A.; Bateup, H.S.; Floor, S.N. Widespread translational remodeling during human neuronal differentiation. Cell Rep. 2017, 21, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Follwaczny, P.; Schieweck, R.; Riedemann, T.; Demleitner, A.; Straub, T.; Klemm, A.H.; Bilban, M.; Sutor, B.; Popper, B.; Kiebler, M. Pumilio2-deficient mice show a predisposition for epilepsy. Dis. Model. Mech. 2017, 10, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Mircsof, D.; The DDD Study; Langouet, M.; Rio, M.; Moutton, S.; Siquier-Pernet, K.; Bole-Feysot, C.; Cagnard, N.; Nitschké, P.; Gaspar, L.; et al. Mutations in NONO lead to syndromic intellectual disability and inhibitory synaptic defects. Nat. Neurosci. 2015, 18, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Neubert, T.A.; Jordan, B.A. RNA Binding Proteins Accumulate at the Postsynaptic Density with Synaptic Activity. J. Neurosci. 2012, 32, 599–609. [Google Scholar] [CrossRef]
- Bryant, C.D.; Yazdani, N. RNA-binding proteins, neural development and the addictions. Genes Brain Behav. 2016, 15, 169–186. [Google Scholar] [CrossRef]
- Gantois, I.; Vandesompele, J.; Speleman, F.; Reyniers, E.; D’Hooge, R.; Severijnen, L.-A.; Willemsen, R.; Tassone, F.; Kooy, R.F. Expression profiling suggests underexpression of the GABAA receptor subunit δ in the fragile X knockout mouse model. Neurobiol. Dis. 2006, 21, 346–357. [Google Scholar] [CrossRef]
- Chen, E.; Sharma, M.R.; Shi, X.; Agrawal, R.K.; Joseph, S. Fragile X Mental Retardation Protein Regulates Translation by Binding Directly to the Ribosome. Mol. Cell 2014, 54, 407–417. [Google Scholar] [CrossRef]
- Antar, L.N.; Dictenberg, J.B.; Plociniak, M.; Afroz, R.; Bassell, G.J. Localization of FMRP-associated mRNA granules and requirement of microtubules for activity-dependent trafficking in hippocampal neurons. Genes Brain Behav. 2005, 4, 350–359. [Google Scholar] [CrossRef]
- Liu, B.; Li, Y.; Stackpole, E.E.; Novak, A.; Gao, Y.; Zhao, Y.; Zhao, X.; Richter, J.D. Regulatory discrimination of mRNAs by FMRP controls mouse adult neural stem cell differentiation. Proc. Natl. Acad. Sci. USA 2018, 115, E11397–E11405. [Google Scholar] [CrossRef]
- Adusei, D.C.; Pacey, L.K.K.; Chen, D.; Hampson, D.R. Early developmental alterations in GABAergic protein expression in fragile X knockout mice. Neuropharmacology 2010, 59, 167–171. [Google Scholar] [CrossRef]
- D’Hulst, C.; De Geest, N.; Reeve, S.P.; Van Dam, D.; De Deyn, P.P.; Hassan, B.A.; Kooy, R.F. Decreased expression of the GABAA receptor in fragile X syndrome. Brain Res. 2006, 1121, 238–245. [Google Scholar] [CrossRef]
- Hong, A.; Zhang, A.; Ke, Y.; El Idrissi, A.; Shen, C.-H. Downregulation of GABA(A) β subunits is transcriptionally controlled by Fmr1p. J. Mol. Neurosci. 2012, 46, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Curia, G.; Papouin, T.; Séguéla, P.; Avoli, M. Downregulation of tonic GABAergic inhibition in a mouse model of fragile X syndrome. Cereb. Cortex 2009, 19, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Bushati, N.; Cohen, S.M. microRNA Functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Sartor, G.C.; St. Laurent, G.; Wahlestedt, C. The Emerging Role of Non-Coding RNAs in Drug Addiction. Front. Genet. 2012, 3, 106. [Google Scholar] [CrossRef]
- Zhao, C.; Huang, C.; Weng, T.; Xiao, X.; Ma, H.; Liu, L. Computational prediction of MicroRNAs targeting GABA receptors and experimental verification of miR-181, miR-216 and miR-203 targets in GABA-A receptor. BMC Res. Notes 2012, 5, 91. [Google Scholar] [CrossRef]
- Sengupta, J.; Pochiraju, S.; Pochiraju, S.; Kannampalli, P.; Bruckert, M.; Addya, S.; Yadav, P.; Miranda, A.; Shaker, R.; Banerjee, B. MicroRNA-mediated GABAAα-1 receptor subunit downregulation in adult spinal cord following neonatal cystitis-induced chronic visceral pain in rats. Pain 2013, 154, 59–70. [Google Scholar] [CrossRef]
- Bekdash, R.A.; Harrison, N.L. Downregulation of Gabra4 expression during alcohol withdrawal is mediated by specific microRNAs in cultured mouse cortical neurons. Brain Behav. 2015, 5, e00355. [Google Scholar] [CrossRef]
- Janeczek, P.; Colson, N.; Dodd, P.R.; Lewohl, J.M. Sex Differences in the Expression of the α5 Subunit of the GABAA Receptor in Alcoholics with and without Cirrhosis of the Liver. Alcohol. Clin. Exp. Res. 2020, 44, 423–434. [Google Scholar] [CrossRef]
- Bali, P.; Kenny, P.J. MicroRNAs and Drug Addiction. Front. Genet. 2013, 4, 335–344. [Google Scholar] [CrossRef]
- Li, M.D.; van der Vaart, A.D. MicroRNAs in addiction: Adaptation’s middlemen? Mol. Psychiatry 2011, 16, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.W.; Kenny, P.J. MicroRNAs regulate synaptic plasticity underlying drug addiction. Genes Brain Behav. 2018, 17, e12424. [Google Scholar] [CrossRef] [PubMed]
- Zito, K.; Svoboda, K. Activity-Dependent Synaptogenesis in the Adult Mammalian Cortex. Neuron 2002, 35, 1015–1017. [Google Scholar] [CrossRef]
- Wong, R.O.L.; Ghosh, A. Activity-dependent regulation of dendritic growth and patterning. Nat. Rev. Neurosci. 2002, 3, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, N.C. Electrical activity in early neuronal development. Nature 2006, 444, 707–712. [Google Scholar] [CrossRef]
- Katz, L.C.; Shatz, C.J. Synaptic Activity and the Construction of Cortical Circuits. Science 1996, 274, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Bloodgood, B.L.; Hauser, J.L.; Lapan, A.D.; Koon, A.C.; Kim, T.-K.; Hu, L.S.; Malik, A.N.; Greenberg, M.E. Activity-dependent regulation of inhibitory synapse development by Npas4. Nature 2008, 455, 1198–1204. [Google Scholar] [CrossRef]
- Hartzell, A.L.; Martyniuk, K.M.; Brigidi, G.S.; Heinz, D.A.; Djaja, N.A.; Payne, A.; Bloodgood, B.L. NPAS4 recruits CCK basket cell synapses and enhances cannabinoid-sensitive inhibition in the mouse hippocampus. eLife 2018, 7, e35927. [Google Scholar] [CrossRef] [PubMed]
- Benevento, L.A.; Bakkum, B.W.; Cohen, R.S. gamma-Aminobutyric acid and somatostatin immunoreactivity in the visual cortex of normal and dark-reared rats. Brain Res. 1995, 689, 172–182. [Google Scholar] [CrossRef]
- Hensch, T.K. Critical period plasticity in local cortical circuits. Nat. Rev. Neurosci. 2005, 6, 877–888. [Google Scholar] [CrossRef]
- Marty, S.; Wehrlé, R.; Sotelo, C. Neuronal Activity and Brain-Derived Neurotrophic Factor Regulate the Density of Inhibitory Synapses in Organotypic Slice Cultures of Postnatal Hippocampus. J. Neurosci. 2000, 20, 8087–8095. [Google Scholar] [CrossRef] [PubMed]
- Turrigiano, G.G.; Nelson, S.B. Homeostatic plasticity in the developing nervous system. Nat. Rev. Neurosci. 2004, 5, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Renier, N.; Adams, E.L.; Kirst, C.; Wu, Z.; Azevedo, R.; Kohl, J.; Autry, A.E.; Kadiri, L.; Venkataraju, K.U.; Zhou, Y.; et al. Mapping of Brain Activity by Automated Volume Analysis of Immediate Early Genes. Cell 2016, 165, 1789–1802. [Google Scholar] [CrossRef] [PubMed]
- Bullitt, E. Expression of c-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J. Comp. Neurol. 1990, 296, 517–530. [Google Scholar] [CrossRef]
- Roberts, D.S.; Hu, Y.; Lund, I.V.; Brooks-Kayal, A.R.; Russek, S.J. Brain-derived Neurotrophic Factor (BDNF)-induced Synthesis of Early Growth Response Factor 3 (Egr3) Controls the Levels of Type A GABA Receptorα4 Subunits in Hippocampal Neurons. J. Biol. Chem. 2006, 281, 29431–29435. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A Simple Role for BDNF in Learning and Memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef]
- Logrip, M.L.; Barak, S.; Warnault, V.; Ron, D. Corticostriatal BDNF and alcohol addiction. Brain Res. 2015, 1628, 60–67. [Google Scholar] [CrossRef]
- Koskela, M.; Bäck, S.; Voikar, V.; Richie, C.T.; Domanskyi, A.; Harvey, B.K.; Airavaara, M. Update of neurotrophic factors in neurobiology of addiction and future directions. Neurobiol. Dis. 2017, 97, 189–200. [Google Scholar] [CrossRef]
- Bloodgood, B.L.; Sharma, N.; Browne, H.A.; Trepman, A.Z.; Greenberg, M.E. The activity-dependent transcription factor NPAS4 regulates domain-specific inhibition. Nature 2013, 503, 121–125. [Google Scholar] [CrossRef]
- Hasin, D.S.; O’Brien, C.P.; Auriacombe, M.; Borges, G.; Bucholz, K.; Budney, A.; Compton, W.M.; Crowley, T.; Ling, W.; Petry, N.M.; et al. DSM-5 Criteria for Substance Use Disorders: Recommendations and Rationale. Am. J. Psychiatry 2013, 170, 834–851. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurobiology of addiction: A neurocircuitry analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef]
- Lüscher, C.; Robbins, T.W.; Everitt, B.J. The transition to compulsion in addiction. Nat. Rev. Neurosci. 2020, 21, 247–263. [Google Scholar] [CrossRef]
- Sanchis-Segura, C.; Spanagel, R. Behavioural assessment of drug reinforcement and addictive features in rodents: An overview. Addict Biol. 2006, 11, 2–38. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of Addiction. Neuropsychopharmacol 2010, 35, 217–238. [Google Scholar] [CrossRef]
- Mameli, M.; Halbout, B.; Creton, C.; Engblom, D.; Parkitna, J.R.; Spanagel, R.; Lüscher, C. Cocaine-evoked synaptic plasticity: Persistence in the VTA triggers adaptations in the NAc. Nat. Neurosci. 2009, 12, 1036–1041. [Google Scholar] [CrossRef]
- Bali, P.; Kenny, P.J. Transcriptional mechanisms of drug addiction. Dialogues Clin. Neurosci. 2019, 21, 379–387. [Google Scholar] [PubMed]
- Olsen, R.W. GABAA Receptor: Positive and Negative Allosteric Modulators. Neuropharmacology 2018, 136, 10–22. [Google Scholar] [CrossRef]
- Forman, S.A.; Miller, K.W. Mapping General Anesthetic Sites in Heteromeric Gamma-Aminobutyric Acid Type A Receptors Reveals a Potential For Targeting Receptor Subtypes. Anesth. Analg. 2016, 123, 1263–1273. [Google Scholar] [CrossRef]
- Centanni, S.W.; Burnett, E.J.; Trantham-Davidson, H.; Chandler, L.J. Loss of δ-GABAA receptor-mediated tonic currents in the adult prelimbic cortex following adolescent alcohol exposure. Addict Biol. 2017, 22, 616–628. [Google Scholar] [CrossRef]
- Wei, W.; Faria, L.C.; Mody, I. Low ethanol concentrations selectively augment the tonic inhibition mediated by delta subunit-containing GABAA receptors in hippocampal neurons. J. Neurosci. 2004, 24, 8379–8382. [Google Scholar] [CrossRef]
- Herman, M.; Roberto, M. Cell type-specific tonic GABA signaling in the rat central amygdala is selectively altered by acute and chronic ethanol. Addict. Biol. 2016, 21, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Jacob, T.C.; Michels, G.; Silayeva, L.; Haydon, J.; Succol, F.; Moss, S.J. Benzodiazepine treatment induces subtype-specific changes in GABAA receptor trafficking and decreases synaptic inhibition. Proc. Natl. Acad. Sci. USA 2012, 109, 18595–18600. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, U.; Möhler, H. Analysis of GABAA receptor function and dissection of the pharmacology of benzodiazepines and general anesthetics through mouse genetics. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 475–498. [Google Scholar] [CrossRef] [PubMed]
- Poncer, J.-C.; Dürr, R.; Gähwiler, B.H.; Thompson, S.M. Modulation of Synaptic GABAA Receptor Function by Benzodiazepines in Area CA3 of Rat Hippocampal Slice Cultures. Neuropharmacology 1996, 35, 1169–1179. [Google Scholar] [CrossRef]
- Kang, I.; Miller, L.G. Decreased GABAA receptor subunit mRNA concentrations following chronic lorazepam administration. Br. J. Pharmacol. 1991, 103, 1285–1287. [Google Scholar] [CrossRef]
- Auta, J.; Gatta, E.; Davis, J.M.; Pandey, S.C.; Guidotti, A. Potential role for histone deacetylation in chronic diazepam-induced downregulation of α1-GABAA receptor subunit expression. Pharmacol. Res. Perspect. 2018, 6, e00416. [Google Scholar] [CrossRef]
- Follesa, P.; Cagetti, E.; Mancuso, L.; Biggio, F.; Manca, A.; Maciocco, E.; Massa, F.; Desole, M.S.; Carta, M.; Busonero, F.; et al. Increase in expression of the GABA(A) receptor alpha(4) subunit gene induced by withdrawal of, but not by long-term treatment with, benzodiazepine full or partial agonists. Brain Res. Mol. Brain Res. 2001, 92, 138–148. [Google Scholar] [CrossRef]
- Pandey, S.C.; Ugale, R.; Zhang, H.; Tang, L.; Prakash, A. Brain Chromatin Remodeling: A Novel Mechanism of Alcoholism. J. Neurosci. 2008, 28, 3729–3737. [Google Scholar] [CrossRef]
- Kumar, S.; Porcu, P.; Werner, D.F.; Matthews, U.B.; Diaz-Granados, J.L.; Helfand, R.S.; Morrow, A.L. The role of GABAA receptors in the acute and chronic effects of ethanol: A decade of progress. Psychopharmacology 2009, 205, 529. [Google Scholar] [CrossRef]
- Lewohl, J.M.; Crane, D.I.; Dodd, P.R. Expression of the alpha 1, alpha 2 and alpha 3 isoforms of the GABAA receptor in human alcoholic brain. Brain Res. 1997, 751, 102–112. [Google Scholar] [CrossRef]
- Matthews, D.B.; Devaud, L.L.; Fritschy, J.M.; Sieghart, W.; Morrow, A.L. Differential regulation of GABA(A) receptor gene expression by ethanol in the rat hippocampus versus cerebral cortex. J. Neurochem. 1998, 70, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Sanna, E.; Mosallino, M.C.; Busonero, F.; Talani, G.; Tranquilli, S.; Mameli, M.; Spiga, S.; Follesa, P.; Biggio, G. Changes in GABAA Receptor Gene Expression Associated with Selective Alterations in Receptor Function and Pharmacology after Ethanol Withdrawal. J. Neurosci. 2003, 23, 11711–11724. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.L.; Ferreri, M.C.; Gravielle, M.C. GABA-induced uncoupling of GABA/benzodiazepine site interactions is mediated by increased GABAA receptor internalization and associated with a change in subunit composition. Neuroscience 2014, 257, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Reynolds, J.N. Uncoupling of GABA-benzodiazepine receptors in chick cerebral cortical neurons requires co-activation of both receptor sites. Brain Res. 1992, 591, 327–331. [Google Scholar] [CrossRef]
- Lindemeyer, A.K.; Shen, Y.; Yazdani, F.; Shao, X.M.; Spigelman, I.; Davies, D.L.; Olsen, R.W.; Liang, J. α2 Subunit—Containing GABAA Receptor Subtypes Are Upregulated and Contribute to Alcohol-Induced Functional Plasticity in the Rat Hippocampus. Mol. Pharmacol. 2017, 92, 101–112. [Google Scholar] [CrossRef]
- File, S.E.; Wilks, L.J.; Mabbutt, P.S. Withdrawal, tolerance and sensitization after a single dose of lorazepam. Pharmacol. Biochem. Behav. 1988, 31, 937–940. [Google Scholar] [CrossRef]
- Berkel, T.D.M.; Pandey, S.C. Emerging Role of Epigenetic Mechanisms in Alcohol Addiction. Alcohol. Clin. Exp. Res. 2017, 41, 666–680. [Google Scholar] [CrossRef]
- Nott, A.; Cheng, J.; Gao, F.; Lin, Y.-T.; Gjoneska, E.; Ko, T.; Minhas, P.; Zamudio, A.V.; Meng, J.; Zhang, F.; et al. Histone deacetylase 3 associates with MeCP2 to regulate FOXO and social behavior. Nat. Neurosci. 2016, 19, 1497–1505. [Google Scholar] [CrossRef]
- Tesone-Coelho, C.; Varela, P.; Escosteguy-Neto, J.C.; Cavarsan, C.; Mello, L.E.; Santos-Junior, J.G. Effects of ethanol on hippocampal neurogenesis depend on the conditioned appetitive response. Addict. Biol. 2013, 18, 774–785. [Google Scholar] [CrossRef]
- Roca, D.J.; Rozenberg, I.; Farrant, M.; Farb, D.H. Chronic agonist exposure induces down-regulation and allosteric uncoupling of the gamma-aminobutyric acid/benzodiazepine receptor complex. Mol. Pharmacol. 1990, 37, 37–43. [Google Scholar]
- Holt, R.A.; Bateson, A.N.; Martin, I.L. Decreased GABA Enhancement of Benzodiazepine Binding after a Single Dose of Diazepam. J. Neurochem. 1999, 72, 2219–2222. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.; Lyon, T.; Skolnick, P. Chronic exposure to benzodiazepine receptor ligands uncouples the gamma-aminobutyric acid type A receptor in WSS-1 cells. Mol. Pharmacol. 1994, 46, 1056–1062. [Google Scholar] [PubMed]
- Hernandez, T.D.; Heninger, C.; Wilson, M.A.; Gallager, D.W. Relationship of agonist efficacy to changes in GABA sensitivity and anticonvulsant tolerance following chronic benzodiazepine ligand exposure. Eur. J. Pharmacol. 1989, 170, 145–155. [Google Scholar] [CrossRef]
- Foitzick, M.F.; Medina, N.B.; Iglesias García, L.C.; Gravielle, M.C. Benzodiazepine exposure induces transcriptional down-regulation of GABAA receptor α1 subunit gene via L-type voltage-gated calcium channel activation in rat cerebrocortical neurons. Neurosci. Lett. 2020, 721, 134801. [Google Scholar] [CrossRef]
- Jacob, T.C.; Moss, S.J.; Jurd, R. GABAA receptor trafficking and its role in the dynamic modulation of neuronal inhibition. Nat. Rev. Neurosci. 2008, 9, 331–343. [Google Scholar] [CrossRef]
- Feng, J.; Nestler, E.J. Epigenetic Mechanisms of Drug Addiction. Curr. Opin. Neurobiol. 2013, 23, 521–528. [Google Scholar] [CrossRef]
- Starkman, B.G.; Sakharkar, A.J.; Pandey, S.C. Epigenetics—Beyond the Genome in Alcoholism. Alcohol. Res. 2012, 34, 293–305. [Google Scholar]
- Liu, C.; Marioni, R.E.; Hedman, Å.K.; Pfeiffer, L.; Tsai, P.-C.; Reynolds, L.M.; Just, A.C.; Duan, Q.; Boer, C.G.; Tanaka, T.; et al. A DNA methylation biomarker of alcohol consumption. Mol. Psychiatry 2018, 23, 422–433. [Google Scholar] [CrossRef]
- Jin, Z.; Bhandage, A.K.; Bazov, I.; Kononenko, O.; Bakalkin, G.; Korpi, E.R.; Birnir, B. Expression of specific ionotropic glutamate and GABA-A receptor subunits is decreased in central amygdala of alcoholics. Front. Cell. Neurosci. 2014, 8, 288. [Google Scholar] [CrossRef]
- Mitsuyama, H.; Little, K.Y.; Sieghart, W.; Devaud, L.L.; Morrow, A.L. GABA(A) receptor alpha1, alpha4, and beta3 subunit mRNA and protein expression in the frontal cortex of human alcoholics. Alcohol. Clin. Exp. Res. 1998, 22, 815–822. [Google Scholar]
- Floyd, D.W.; Friedman, D.P.; Daunais, J.B.; Pierre, P.J.; Grant, K.A.; McCool, B.A. Long-Term Ethanol Self-Administration by Cynomolgus Macaques Alters the Pharmacology and Expression of GABAA Receptors in Basolateral Amygdala. J. Pharmacol. Exp. Ther. 2004, 311, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Hemby, S.E.; O’Connor, J.A.; Acosta, G.; Floyd, D.; Anderson, N.; McCool, B.A.; Friedman, D.; Grant, K.A. Ethanol-Induced Regulation of GABAA Subunit mRNAs in Prefrontal Fields of Cynomolgus Monkeys. Alcohol. Clin. Exp. Res. 2006, 30, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Blednov, Y.A.; Walker, D.; Alva, H.; Creech, K.; Findlay, G.; Harris, R.A. GABAA Receptor α1 and β2 Subunit Null Mutant Mice: Behavioral Responses to Ethanol. J. Pharmacol. Exp. Ther. 2003, 305, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Blednov, Y.A.; Jung, S.; Alva, H.; Wallace, D.; Rosahl, T.; Whiting, P.-J.; Harris, R.A. Deletion of the α1 or β2 Subunit of GABAAReceptors Reduces Actions of Alcohol and Other Drugs. J. Pharmacol. Exp. Ther. 2003, 304, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Boehm, S.L.; Ponomarev, I.; Jennings, A.W.; Whiting, P.J.; Rosahl, T.W.; Garrett, E.M.; Blednov, Y.A.; Harris, R.A. γ-Aminobutyric acid A receptor subunit mutant mice: New perspectives on alcohol actions. Biochem. Pharmacol. 2004, 68, 1581–1602. [Google Scholar] [CrossRef]
- June, H.L.; Foster, K.L.; Eiler, W.J.A.; Goergen, J.; Cook, J.B.; Johnson, N.; Mensah-Zoe, B.; Simmons, J.O.; Yin, W.; Cook, J.M.; et al. Dopamine and Benzodiazepine-Dependent Mechanisms Regulate the EtOH-Enhanced Locomotor Stimulation in the GABA A α 1 Subunit Null Mutant Mice. Neuropsychopharmacology 2007, 32, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Mihalek, R.M.; Bowers, B.J.; Wehner, J.M.; Kralic, J.E.; Vandoren, M.J.; Morrow, A.L.; Homanics, G.E. GABAA-Receptor δ Subunit Knockout Mice Have Multiple Defects in Behavioral Responses to Ethanol. Alcohol. Clin. Exp. Res. 2001, 25, 1708–1718. [Google Scholar]
- Liu, J.; Yang, A.R.; Kelly, T.; Puche, A.; Esoga, C.; Elnabawi, A.; Merchenthaler, I.; Sieghart, W.; June, H.L.; Aurelian, L. Binge alcohol drinking is associated with GABAA α2-regulated Toll-like receptor 4 (TLR4) expression in the central amygdala. Proc. Natl. Acad. Sci. USA 2011, 108, 4465–4470. [Google Scholar] [CrossRef]
- Rewal, M.; Jurd, R.; Gill, T.M.; He, D.; Ron, D.; Janak, P.H. α4-Containing GABAA Receptors in the Nucleus Accumbens Mediate Moderate Intake of Alcohol. J. Neurosci. 2009, 29, 543–549. [Google Scholar] [CrossRef]
- Edenberg, H.J.; Dick, D.M.; Xuei, X.; Tian, H.; Almasy, L.; Bauer, L.O.; Crowe, R.R.; Goate, A.; Hesselbrock, V.; Jones, K.; et al. Variations in GABRA2, encoding the alpha 2 subunit of the GABA(A) receptor, are associated with alcohol dependence and with brain oscillations. Am. J. Hum. Genet. 2004, 74, 705–714. [Google Scholar] [CrossRef]
- Bierut, L.J.; Agrawal, A.; Bucholz, K.K.; Doheny, K.F.; Laurie, C.; Pugh, E.; Fisher, S.; Fox, L.; Howells, W.; Bertelsen, S.; et al. A genome-wide association study of alcohol dependence. Proc. Natl. Acad. Sci. USA 2010, 107, 5082–5087. [Google Scholar] [CrossRef] [PubMed]
- Lewohl, J.M.; Nunez, Y.O.; Dodd, P.R.; Tiwari, G.R.; Harris, R.A.; Mayfield, R.D. Up-Regulation of MicroRNAs in Brain of Human Alcoholics. Alcohol. Clin. Exp. Res. 2011, 35, 1928–1937. [Google Scholar] [CrossRef] [PubMed]
- Sinirlioglu, Z.A.; Coskunpinar, E.; Akbas, F. miRNA and mRNA expression profiling in rat brain following alcohol dependence and withdrawal. Cell. Mol. Biol. 2017, 63, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kralic, J.E.; O’Buckley, T.K.; Grobin, A.C.; Morrow, A.L. Chronic ethanol consumption enhances internalization of α1 subunit-containing GABAA receptors in cerebral cortex. J. Neurochem. 2003, 86, 700–708. [Google Scholar] [CrossRef]
- Kumar, S.; Suryanarayanan, A.; Boyd, K.N.; Comerford, C.E.; Lai, M.A.; Ren, Q.; Morrow, A.L. Ethanol Reduces GABAA α1 Subunit Receptor Surface Expression by a Protein Kinase Cγ-Dependent Mechanism in Cultured Cerebral Cortical Neurons. Mol. Pharmacol. 2010, 77, 793–803. [Google Scholar] [CrossRef]
- Moss, S.J.; Doherty, C.A.; Huganir, R.L. Identification of the cAMP-dependent protein kinase and protein kinase C phosphorylation sites within the major intracellular domains of the beta 1, gamma 2S, and gamma 2L subunits of the gamma-aminobutyric acid type A receptor. J. Biol. Chem. 1992, 267, 14470–14476. [Google Scholar]
- McDonald, B.J.; Moss, S.J. Differential phosphorylation of intracellular domains of gamma-aminobutyric acid type A receptor subunits by calcium/calmodulin type 2-dependent protein kinase and cGMP-dependent protein kinase. J. Biol. Chem. 1994, 269, 18111–18117. [Google Scholar]
- Parakala, M.L.; Zhang, Y.; Modgil, A.; Chadchankar, J.; Vien, T.N.; Ackley, M.A.; Doherty, J.J.; Davies, P.A.; Moss, S.J. Metabotropic, but not allosteric, effects of neurosteroids on GABAergic inhibition depend on the phosphorylation of GABAA receptors. J. Biol. Chem. 2019, 294, 12220–12230. [Google Scholar] [CrossRef]
- Kumar, S.; Lane, B.M.; Morrow, A.L. Differential Effects of Systemic Ethanol Administration on Protein Kinase Cϵ, γ, and β Isoform Expression, Membrane Translocation, and Target Phosphorylation: Reversal by Chronic Ethanol Exposure. J. Pharmacol. Exp. Ther. 2006, 319, 1366–1375. [Google Scholar] [CrossRef]
- Mcdonald, B.J.; Moss, S.J. Conserved phosphorylation of the intracellular domains of GABAA receptorβ2 and β3 subunits by cAMP-dependent protein kinase, cGMP-dependent protein kinase, protein kinase C and Ca2+/calmodulin type II-dependent protein kinase. Neuropharmacology 1997, 36, 1377–1385. [Google Scholar] [CrossRef]
- Oh, S.; Jang, C.-G.; Ma, T.; Ho, I.K. Activation of protein kinase C by phorbol dibutyrate modulates GABAA receptor binding in rat brain slices. Brain Res. 1999, 850, 158–165. [Google Scholar] [CrossRef]
- Qi, Z.-H.; Song, M.; Wallace, M.; Wang, D.; Newton, P.; McMahon, T.; Chou, W.-H.; Zhang, C.; Shokat, K.; Messing, R.O. Protein Kinase Cϵ Regulates γ-Aminobutyrate Type A Receptor Sensitivity to Ethanol and Benzodiazepines through Phosphorylation of γ2 Subunits. J. Biol. Chem. 2007, 282, 33052–33063. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Pergadia, M.L.; Saccone, S.F.; Hinrichs, A.L.; Lessov-Schlaggar, C.N.; Saccone, N.L.; Neuman, R.J.; Breslau, N.; Johnson, E.O.; Hatsukami, R.; et al. Gamma-aminobutyric acid receptor genes and nicotine dependence: Evidence for association from a case–control study. Addiction 2008, 103, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Balan, I.; Warnock, K.T.; Puche, A.C.; Gondre-Lewis, M.C.; June, H.; Aurelian, L. The GABAA Receptor α2 Subunit Activates a Neuronal TLR4 Signal in the Ventral Tegmental Area that Regulates Alcohol and Nicotine Abuse. Brain Sci. 2018, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- Dixon, C.I.; Morris, H.V.; Breen, G.; Desrivières, S.; Jugurnauth, S.; Steiner, R.C.; Vallada, H.; Guindalini, C.; Laranjeira, R.; Messas, G.; et al. Cocaine effects on mouse incentive-learning and human addiction are linked to α2 subunit-containing GABAA receptors. Proc. Natl. Acad. Sci. USA 2010, 107, 2289–2294. [Google Scholar] [CrossRef]
- Chen, Q.; Lee, T.H.; Wetsel, W.C.; Sun, Q.; Liu, Y.; Davidson, C.; Xiong, X.; Ellinwood, E.H.; Zhang, X. Reversal of cocaine sensitization-associated changes in GAD67 and GABAA receptor α2 subunit expression, and PKC ζ activity. Biochem. Biophys. Res. Commun. 2007, 356, 733–738. [Google Scholar] [CrossRef]
- Enoch, M.-A.; Zhou, Z.; Kimura, M.; Mash, D.C.; Yuan, Q.; Goldman, D. GABAergic Gene Expression in Postmortem Hippocampus from Alcoholics and Cocaine Addicts; Corresponding Findings in Alcohol-Naïve P and NP Rats. PLoS ONE 2012, 7, e29369. [Google Scholar] [CrossRef]
- Wearne, T.A.; Parker, L.M.; Franklin, J.L.; Goodchild, A.K.; Cornish, J.L. GABAergic mRNA expression is upregulated in the prefrontal cortex of rats sensitized to methamphetamine. Behav. Brain Res. 2016, 297, 224–230. [Google Scholar] [CrossRef]
- Zhang, X.; Lee, T.H.; Xiong, X.; Chen, Q.; Davidson, C.; Wetsel, W.C.; Ellinwood, E.H. Methamphetamine induces long-term changes in GABAA receptor α2 subunit and GAD67 expression. Biochem. Biophys. Res. Commun. 2006, 351, 300–305. [Google Scholar] [CrossRef]
- Ammon-Treiber, S.; Höllt, V. Morphine-induced Changes of Gene Expression in the Brain. Addict. Biol. 2005, 10, 81–89. [Google Scholar] [CrossRef]
- Spijker, S.; Houtzager, S.W.J.; De Gunst, M.C.M.; De Boer, W.P.H.; Schoffelmeer, A.N.M.; Smit, A.B. Morphine exposure and abstinence define specific stages of gene expression in the rat nucleus accumbens. FASEB J. 2004, 18, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Heikkilä, A.T.; Echenko, O.; Uusi-Oukari, M.; Sinkkonen, S.T.; Korpi, E.R. Morphine withdrawal increases expression of GABA(A) receptor epsilon subunit mRNA in locus coeruleus neurons. Neuroreport 2001, 12, 2981–2985. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.A.; Tipton, A.F.; Zhang, H.; Akbari, A.; Pandey, S.C. Effect of Histone Deacetylase Inhibitor on Ethanol Withdrawal-Induced Hyperalgesia in Rats. Int. J. Neuropsychopharmacol. 2019, 22, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Tapocik, J.D.; Solomon, M.; Flanigan, M.E.; Meinhardt, M.; Barbier, E.; Schank, J.R.; Schwandt, M.; Sommer, W.H.; Heilig, M. Coordinated dysregulation of mRNAs and microRNAs in the rat medial prefrontal cortex following a history of alcohol dependence. Pharm. J. 2013, 13, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Vithlani, M.; Hines, R.M.; Zhong, P.; Terunuma, M.; Hines, D.J.; Revilla-Sanchez, R.; Jurd, R.; Haydon, P.; Rios, M.; Brandon, N.; et al. The ability of BDNF to modify neurogenesis and depressive-like behaviors is dependent upon phosphorylation of tyrosine residues 365/367 in the GABA(A)-receptor γ2 subunit. J. Neurosci. 2013, 33, 15567–15577. [Google Scholar] [CrossRef] [PubMed]
- Staley, K. Molecular mechanisms of epilepsy. Nat. Neurosci. 2015, 18, 367–372. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Benzodiazepine (BZ) Use | Alcohol Use | |
|---|---|---|
| Transcription |
|
|
| Post-Translational Modification | ||
| Total Expression | ||
| Surface Expression |
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barker, J.S.; Hines, R.M. Regulation of GABAA Receptor Subunit Expression in Substance Use Disorders. Int. J. Mol. Sci. 2020, 21, 4445. https://doi.org/10.3390/ijms21124445
Barker JS, Hines RM. Regulation of GABAA Receptor Subunit Expression in Substance Use Disorders. International Journal of Molecular Sciences. 2020; 21(12):4445. https://doi.org/10.3390/ijms21124445
Chicago/Turabian StyleBarker, Jeffrey S., and Rochelle M. Hines. 2020. "Regulation of GABAA Receptor Subunit Expression in Substance Use Disorders" International Journal of Molecular Sciences 21, no. 12: 4445. https://doi.org/10.3390/ijms21124445
APA StyleBarker, J. S., & Hines, R. M. (2020). Regulation of GABAA Receptor Subunit Expression in Substance Use Disorders. International Journal of Molecular Sciences, 21(12), 4445. https://doi.org/10.3390/ijms21124445