Modelling Spatial Scales of Dose Deposition and Radiolysis Products from Gold Nanoparticle Sensitisation of Proton Therapy in A Cell: From Intracellular Structures to Adjacent Cells



Abstract
1. Introduction
2. Results
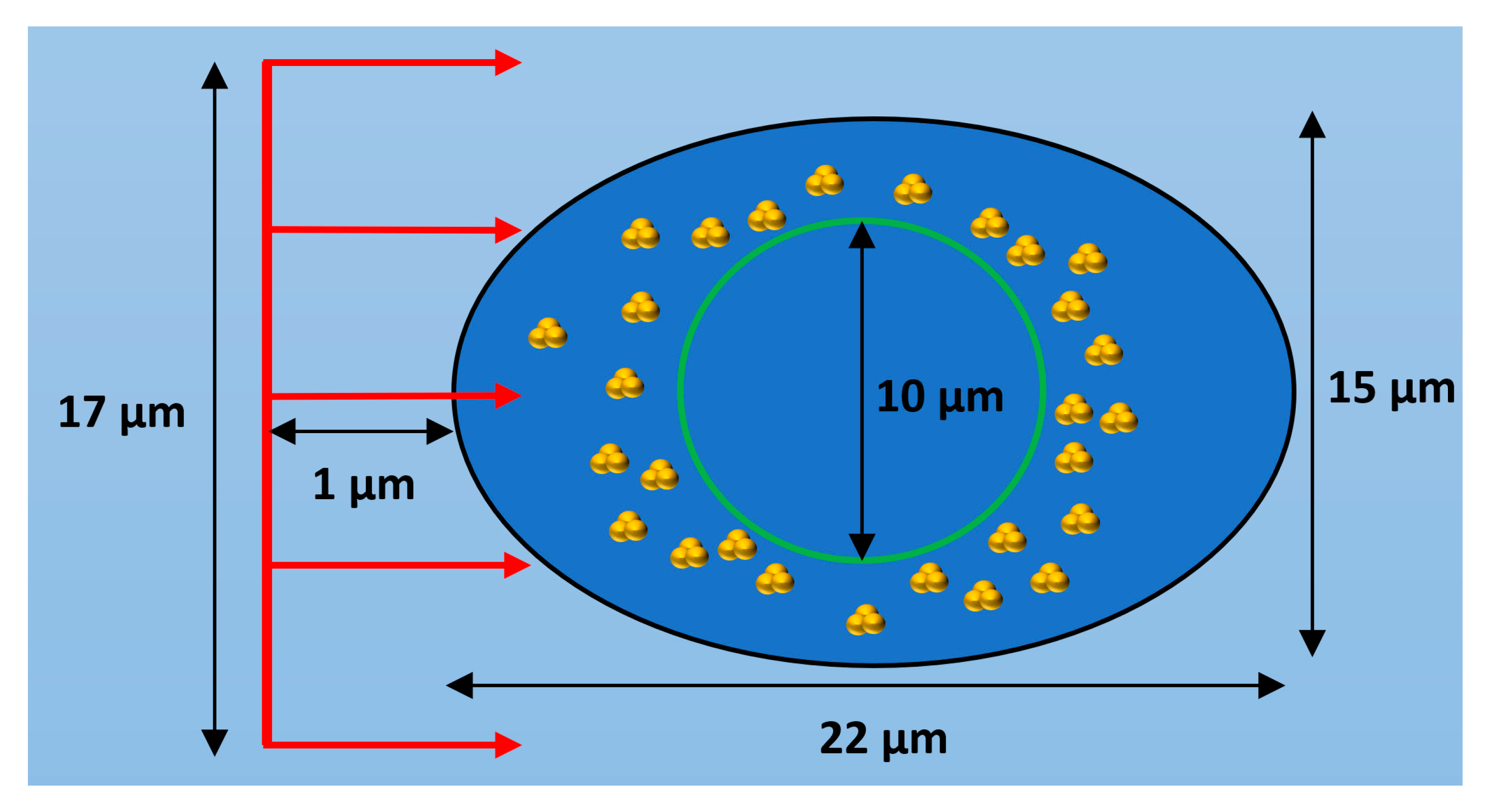
2.1. GNP Cellular Distribution
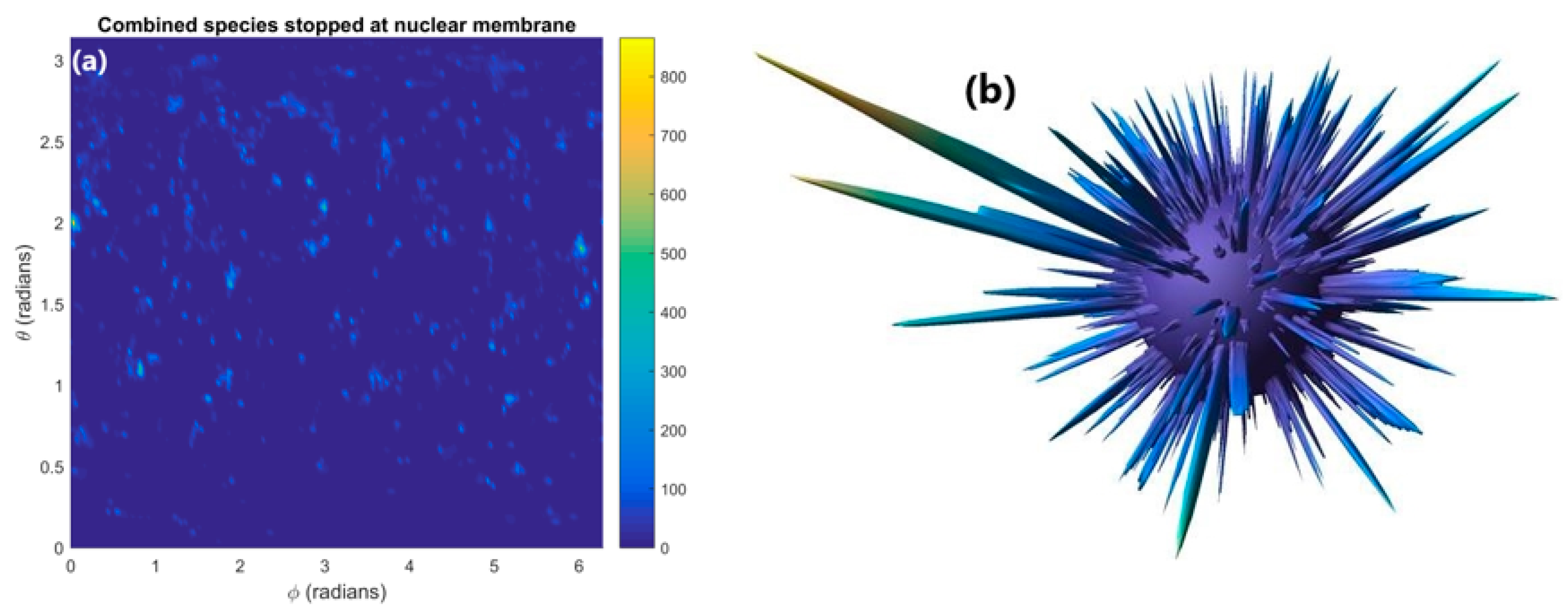
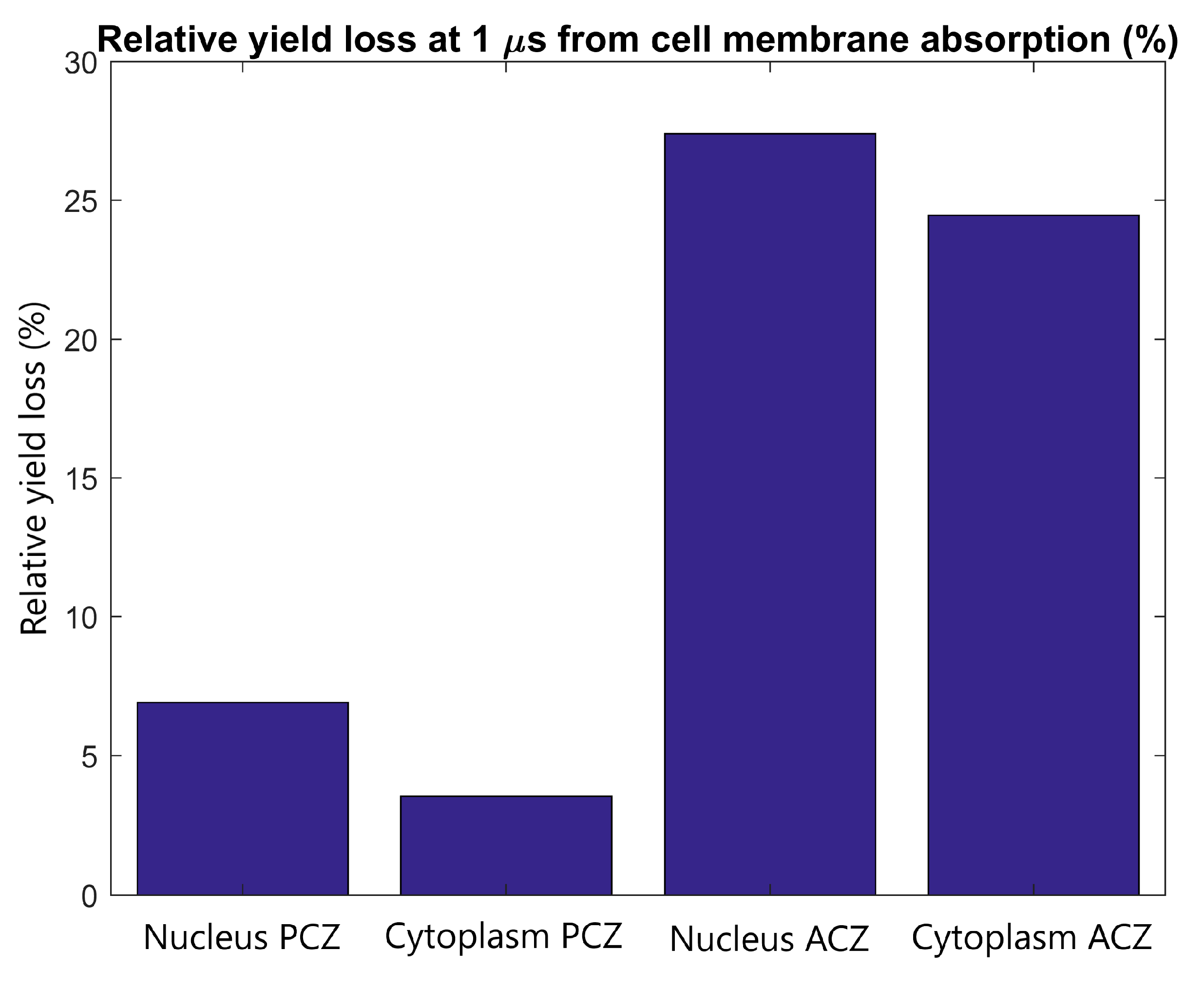
2.2. Absorption in Nuclear Membrane
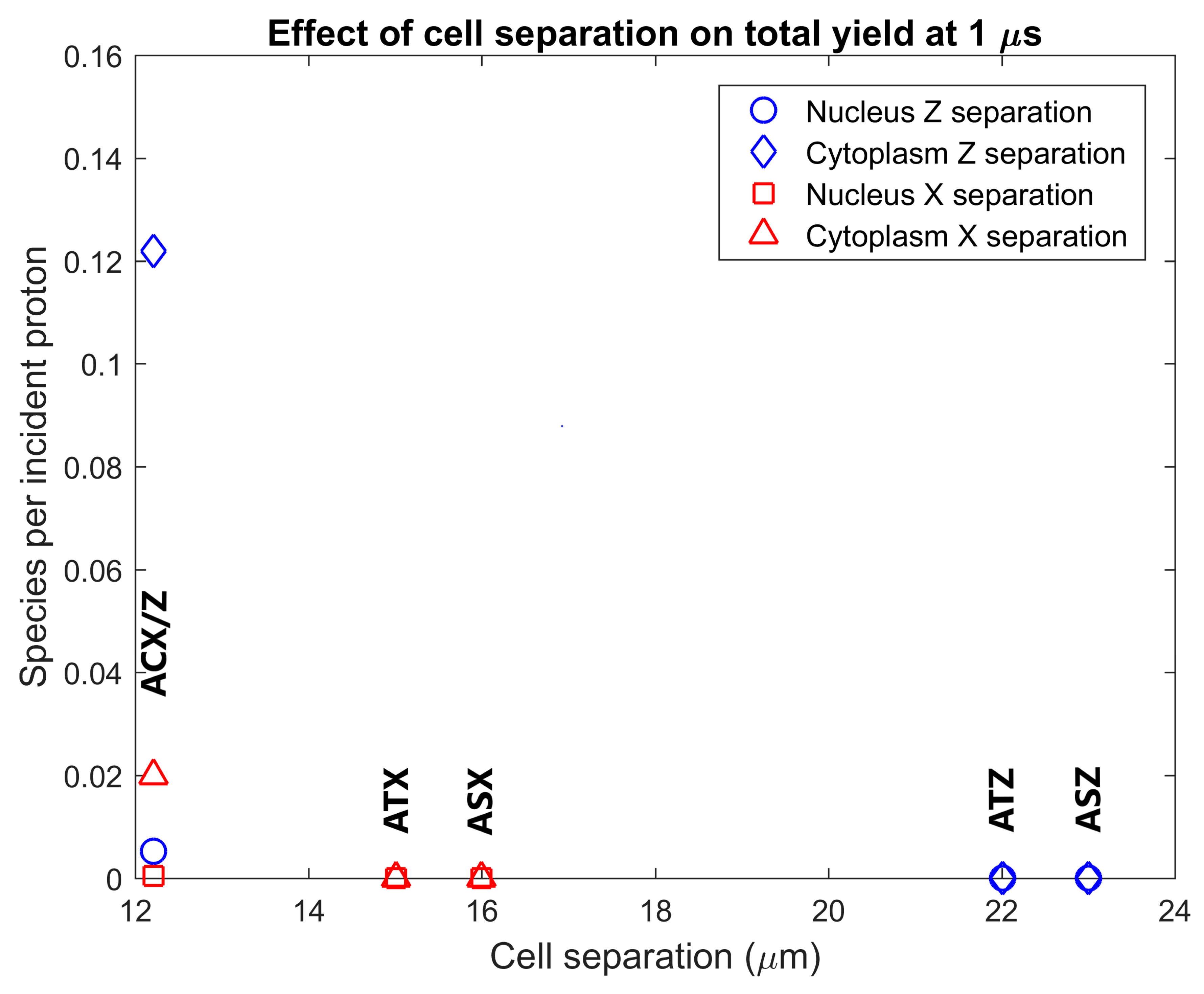
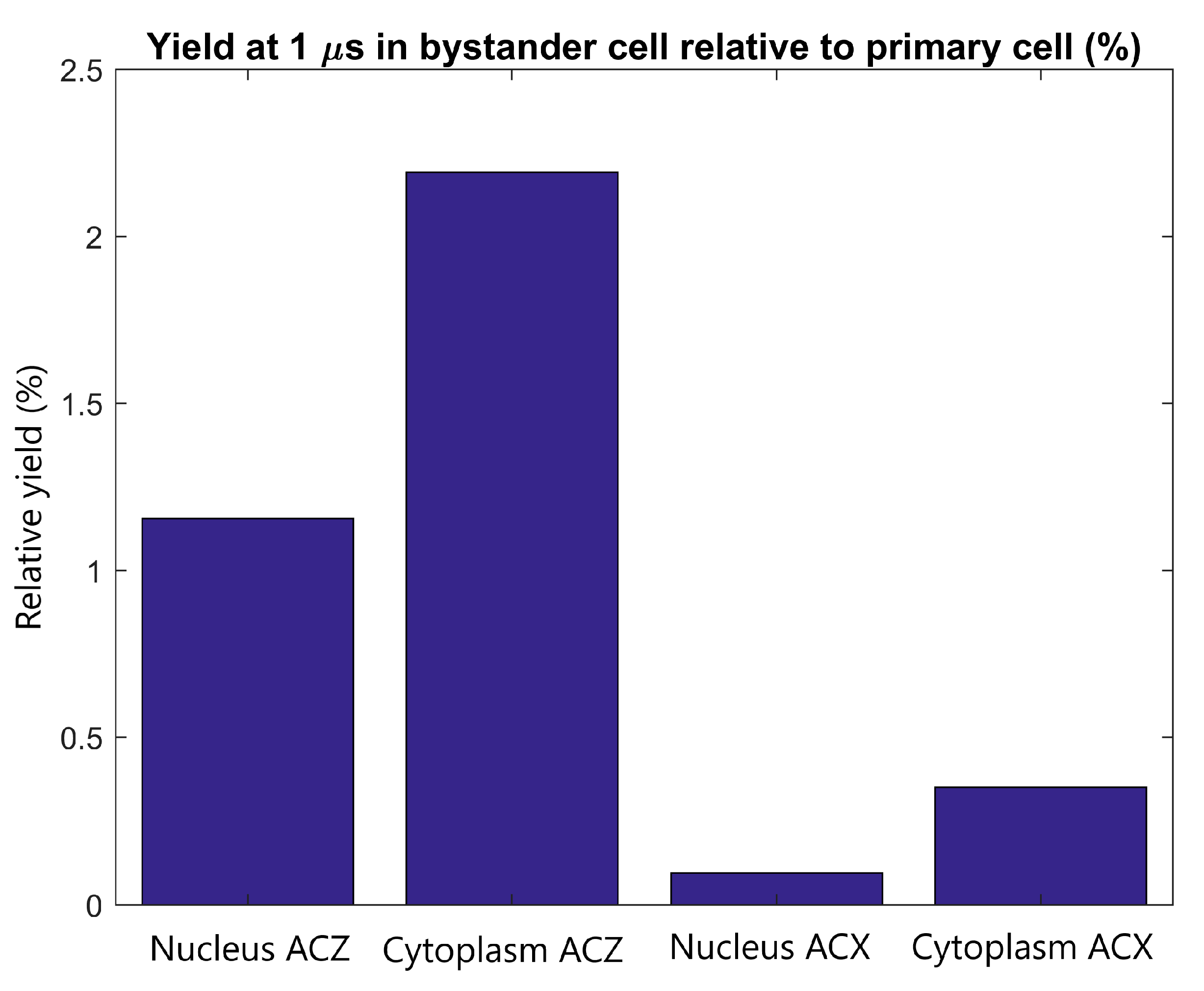
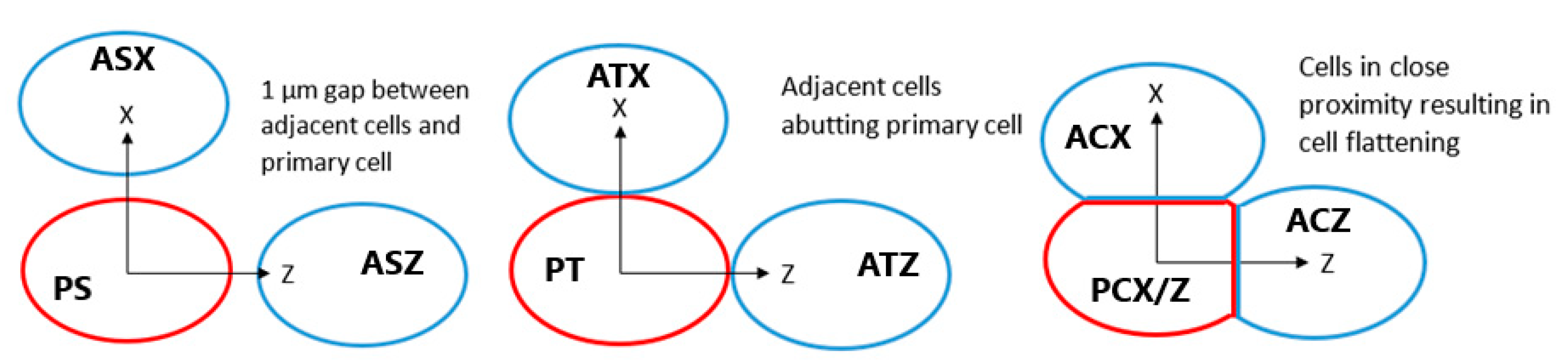
2.3. Inter-Cellular Effects and Cell Membrane Absorption
3. Discussion
3.1. GNP Cellular Distribution
3.2. Absorption in Nuclear Membrane
3.3. Inter-Cellular Effects and Cell Membrane Absorption
3.4. Validation and Limitations
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GNP | Gold Nanoparticle |
| EPR | Enhanced Permeation and Retention |
| LEM | Local Effect Model |
| WNP | Water Nanoparticle |
| LET | Linear Energy Transfer |
References
- Matsumura, Y.; Maeda, H. A New Concept for Macromolecular Therapeutics in Cancer Chemotherapy: Mechanism of Tumoritropic Accumulation of Proteins and the Antitumor Agent Smancs. Cancer Res. 1986, 46, 6387–6392. [Google Scholar] [PubMed]
- Wang, C.-H.; Liu, C.-J.; Chien, C.-C.; Chen, H.-T.; Hua, T.-E.; Leng, W.-H.; Chen, H.-H.; Kempson, I.M.; Hwu, Y.; Hsiao, M.; et al. X-ray synthesized PEGylated (polyethylene glycol coated) gold nanoparticles in mice strongly accumulate in tumors. Mater. Chem. Phys. 2011, 126, 352–356. [Google Scholar] [CrossRef]
- Liu, C.-J.; Wang, C.-H.; Chen, S.-T.; Chen, H.-H.; Leng, W.-H.; Chien, C.-C.; Wang, C.-L.; Kempson, I.M.; Hwu, Y.; Lai, T.-C.; et al. Enhancement of cell radiation sensitivity by pegylated gold nanoparticles. Phys. Med. Biol. 2010, 55, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Polf, J.C.; Bronk, L.F.; Driessen, W.H.P.; Arap, W.; Pasqualini, R.; Gillin, M. Enhanced relative biological effectiveness of proton radiotherapy in tumor cells with internalized gold nanoparticles. Appl. Phys. Lett. 2011, 98, 193702. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Penninckx, S.; Karmani, L.; Heuskin, A.-C.; Watillon, K.; Marega, R.; Zola, J.; Corvaglia, V.; Genard, G.; Gallez, B.; et al. LET-dependent radiosensitization effects of gold nanoparticles for proton irradiation. Nanotechnology 2016, 27, 455101. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Seo, S.-J.; Kim, K.-H.; Kim, T.-J.; Chung, M.-H.; Kim, K.-R.; Yang, T.-K. Therapeutic application of metallic nanoparticles combined with particle-induced X-ray emission effect. Nanotechnology 2010, 21, 425102. [Google Scholar] [CrossRef]
- Kim, J.-K.; Seo, S.-J.; Kim, H.-T.; Kim, K.-H.; Chung, M.-H.; Kim, K.-R.; Ye, S.-J. Enhanced proton treatment in mouse tumors through proton irradiated nanoradiator effects on metallic nanoparticles. Phys. Med. Biol. 2012, 57, 8309–8323. [Google Scholar] [CrossRef] [PubMed]
- Schlathölter, T.; Eustache, P.; Porcel, E.; Salado, D.; Stefancikova, L.; Tillement, O.; Lux, F.; Mowat, P.; Biegun, A.K.; Van Goethem, M.-J.; et al. Improving proton therapy by metal-containing nanoparticles: Nanoscale insights. Int. J. Nanomed. 2016, 11, 1549–1556. [Google Scholar] [CrossRef]
- Howard, D.; Sebastian, S.; Le, Q.V.-C.; Thierry, B.; Kempson, I. Chemical Mechanisms of Nanoparticle Radiosensitization and Radioprotection: A Review of Structure-Function Relationships Influencing Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 579. [Google Scholar] [CrossRef]
- Cho, S.H. Estimation of tumour dose enhancement due to gold nanoparticles during typical radiation treatments: A preliminary Monte Carlo study. Phys. Med. Biol. 2005, 50, N163. [Google Scholar] [CrossRef]
- Cho, J.; Gonzalez-Lepera, C.; Manohar, N.; Kerr, M.; Krishnan, S.; Cho, S.H. Quantitative investigation of physical factors contributing to gold nanoparticle-mediated proton dose enhancement. Phys. Med. Biol. 2016, 61, 2562–2581. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Sutherland, K.; Hashimoto, T.; Shirato, H.; Date, H. Spatial distributions of dose enhancement around a gold nanoparticle at several depths of proton Bragg peak. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2016, 384, 113–120. [Google Scholar] [CrossRef]
- McMahon, S.J.; Hyland, W.B.; Muir, M.F.; Coulter, J.A.; Jain, S.; Butterworth, K.T.; Schettino, G.; Dickson, G.R.; Hounsell, A.R.; O’Sullivan, J.M.; et al. Biological consequences of nanoscale energy deposition near irradiated heavy atom nanoparticles. Sci. Rep. 2011, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; McMahon, S.J.; Paganetti, H.; Schuemann, J. Biological modeling of gold nanoparticle enhanced radiotherapy for proton therapy. Phys. Med. Biol. 2015, 60, 4149–4168. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulos, M.; Henthorn, N.T.; Warmenhoven, J.W.; Mackay, R.I.; Kirkby, K.J.; Merchant, M.J. Modelling direct DNA damage for gold nanoparticle enhanced proton therapy. Nanoscale 2017, 9, 18413–18422. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-H.; Chien, C.-C.; Petibois, C.; Wang, C.-L.; Chu, Y.S.; Lai, S.-F.; Hua, T.-E.; Chen, Y.-Y.; Cai, X.; Kempson, I.M.; et al. Quantitative analysis of nanoparticle internalization in mammalian cells by high resolution X-ray microscopy. J. Nanobiotechnol. 2011, 9, 14. [Google Scholar] [CrossRef]
- Rashkow, J.T.; Patel, S.C.; Tappero, R.; Sitharaman, B. Quantification of single-cell nanoparticle concentrations and the distribution of these concentrations in cell population. J. R. Soc. Interface 2014, 11. [Google Scholar] [CrossRef]
- Rothen-Rutishauser, B.; Kuhn, D.A.; Ali, Z.; Gasser, M.; Amin, F.; Parak, W.J.; Vanhecke, D.; Fink, A.; Gehr, P.; Brandenberger, C. Quantification of gold nanoparticle cell uptake under controlled biological conditions and adequate resolution. Nanomedicine 2014, 9, 607–621. [Google Scholar] [CrossRef]
- Lin, Y.; McMahon, S.J.; Scarpelli, M.; Paganetti, H.; Schuemann, J. Comparing gold nano-particle enhanced radiotherapy with protons, megavoltage photons and kilovoltage photons: A Monte Carlo simulation. Phys. Med. Biol. 2014, 59, 7675–7689. [Google Scholar] [CrossRef]
- Haume, K.; Rosa, S.; Grellet, S.; Śmiałek, M.A.; Butterworth, K.T.; Solov’yov, A.V.; Prise, K.M.; Golding, J.; Mason, N.J. Gold nanoparticles for cancer radiotherapy: A review. Cancer Nanotechnol. 2016, 7, 8. [Google Scholar] [CrossRef]
- Her, S.; Jaffray, D.A.; Allen, C. Gold nanoparticles for applications in cancer radiotherapy: Mechanisms and recent advancements. Adv. Drug Deliv. Rev. 2017, 109, 84–101. [Google Scholar] [CrossRef]
- Peukert, D.; Kempson, I.; Douglass, M.; Bezak, E. Metallic nanoparticle radiosensitisation of ion radiotherapy: A review. Phys. Med. 2018, 47, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.N.; Karamitros, M.; Ivanchenko, V.N.; Guatelli, S.; McKinnon, S.; Murakami, K.; Sasaki, T.; Okada, S.; Bordage, M.C.; Francis, Z.; et al. Geant4 Monte Carlo simulation of absorbed dose and radiolysis yields enhancement from a gold nanoparticle under MeV proton irradiation. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2016, 373, 126–139. [Google Scholar] [CrossRef]
- Agostinelli, S.; Allison, J.; Amako, K.; Apostolakis, J.; Araujo, H.; Arce, P.; Asai, M.; Axen, D.; Banerjee, S.; Barrand, G.; et al. GEANT4—A simulation toolkit. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2003, 506, 250–303. [Google Scholar] [CrossRef]
- Allison, J.; Amako, K.; Apostolakis, J.; Araujo, H.; Dubois, P.A.; Asai, M.; Barrand, G.; Capra, R.; Chauvie, S.; Chytracek, R.; et al. Geant4 developments and applications. IEEE Trans. Nucl. Sci. 2006, 53, 270–278. [Google Scholar] [CrossRef]
- Allison, J.; Amako, K.; Apostolakis, J.; Arce, P.; Asai, M.; Aso, T.; Bagli, E.; Bagulya, A.; Banerjee, S.; Barrand, G.; et al. Recent developments in Geant4. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2016, 835, 186–225. [Google Scholar] [CrossRef]
- Hespeels, F.; Lucas, S.; Tabarrant, T.; Scifoni, E.; Kraemer, M.; Chêne, G.; Strivay, D.; Tran, H.N.; Heuskin, A.C. Experimental measurements validate the use of the binary encounter approximation model to accurately compute proton induced dose and radiolysis enhancement from gold nanoparticles. Phys. Med. Biol. 2019, 64, 65014. [Google Scholar] [CrossRef]
- Peukert, D.; Kempson, I.; Douglass, M.; Bezak, E. Gold nanoparticle enhanced proton therapy: A Monte Carlo simulation of the effects of proton energy, nanoparticle size, coating material and coating thickness on dose and radiolysis yield. Med. Phys. 2020, 47, 651–661. [Google Scholar] [CrossRef]
- Peukert, D.; Kempson, I.; Douglass, M.; Bezak, E. Gold Nanoparticle Enhanced Proton Therapy: Monte Carlo Modeling of Reactive Species’ Distributions Around a Gold Nanoparticle and the Effects of Nanoparticle Proximity and Clustering. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Rudek, B.; McNamara, A.L.; Ramos-Mendez, J.; Byrne, H.; Kuncic, Z.; Schuemann, J. Radio-enhancement by gold nanoparticles and their impact on water radiolysis for x-ray, proton and carbon-ion beams. Phys. Med. Biol. 2019, 64, 175005. [Google Scholar] [CrossRef]
- Peukert, D.; Incerti, S.; Kempson, I.; Douglass, M.; Karamitros, M.; Baldacchino, G.; Bezak, E. Validation and investigation of reactive species yields of Geant4-DNA chemistry models. Med. Phys. 2019, 46, 983–998. [Google Scholar] [CrossRef] [PubMed]
- Incerti, S.; Baldacchino, G.; Bernal, M.A.; CAPRA, R.; CHAMPION, C.; FRANCIS, Z.; GUÈYE, P.; MANTERO, A.; MASCIALINO, B.; MORETTO, P.; et al. THE GEANT4-DNA PROJECT. Int. J. Model. Simul. Sci. Comput. 2010, 1, 157–178. [Google Scholar] [CrossRef]
- Incerti, S.; Ivanchenko, A.; Karamitros, M.; Mantero, A.; Moretto, P.; Tran, H.N.; Mascialino, B.; Champion, C.; Ivanchenko, V.N.; Bernal, M.A.; et al. Comparison of GEANT4 very low energy cross section models with experimental data in water. Med. Phys. 2010, 37, 4692–4708. [Google Scholar] [CrossRef] [PubMed]
- Karamitros, M.; Mantero, A.; Incerti, S.; Friedland, W.; Baldacchino, G.; Barberet, P.; Bernal, M.A.; Capra, R.; Champion, C.; El Bitar, Z.; et al. Modeling radiation chemistry in the Geant4 Toolkit. Prog. Nucl. Sci. Technol. 2011, 2, 503–508. [Google Scholar] [CrossRef]
- Karamitros, M.; Luan, S.; Bernal, M.A.; Allison, J.; Baldacchino, G.; Davidkova, M.; Francis, Z.; Friedland, W.; Ivantchenko, V.; Ivantchenko, A.; et al. Diffusion-controlled reactions modeling in Geant4-DNA. J. Comput. Phys. 2014, 274, 841–882. [Google Scholar] [CrossRef]
- Bernal, M.A.; Bordage, M.C.; Brown, J.M.C.; Davídková, M.; Delage, E.; El Bitar, Z.; Enger, S.A.; Francis, Z.; Guatelli, S.; Ivanchenko, V.N.; et al. Track structure modeling in liquid water: A review of the Geant4-DNA very low energy extension of the Geant4 Monte Carlo simulation toolkit. Phys. Med. 2015, 31, 861–874. [Google Scholar] [CrossRef]
- Incerti, S.; Kyriakou, I.; Bernal, M.A.; Bordage, M.C.; Francis, Z.; Guatelli, S.; Ivanchenko, V.; Karamitros, M.; Lampe, N.; Lee, S.B.; et al. Geant4-DNA example applications for track structure simulations in liquid water: A report from the Geant4-DNA Project. Med. Phys. 2018, 45, e722–e739. [Google Scholar] [CrossRef]
- Turnbull, T.; Thierry, B.; Kempson, I. A quantitative study of intercellular heterogeneity in gold nanoparticle uptake across multiple cell lines. Anal. Bioanal. Chem. 2019, 411, 7529–7538. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulation | Dose Per Proton (eV) (GNP) | Yield Per Proton (GNP) | DEF | REF | Main Results | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | C | N | C | N | C | N | C | |||
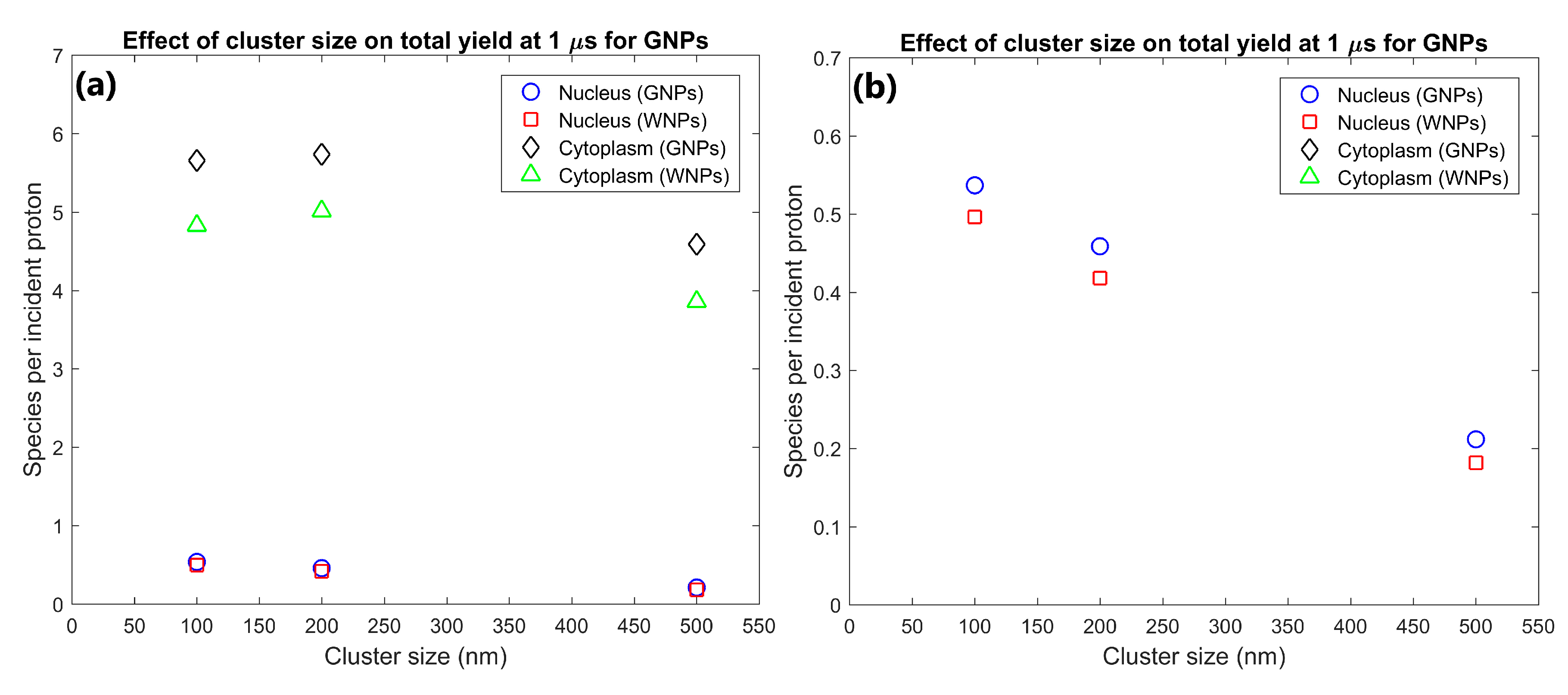
| Cluster size (nm) | 100 | 5.30 | 63.6 | 0.537 | 5.66 | 1.07 | 1.17 | 1.08 | 1.17 | 200 nm clusters had yields 117% and 25% greater than for 500 nm clusters for nucleus and cytoplasm respectively |
| 200 | 4.63 | 64.1 | 0.459 | 5.73 | 1.07 | 1.15 | 1.10 | 1.14 | ||
| 500 | 2.17 | 50.7 | 0.212 | 4.59 | 1.16 | 1.19 | 1.17 | 1.19 | ||
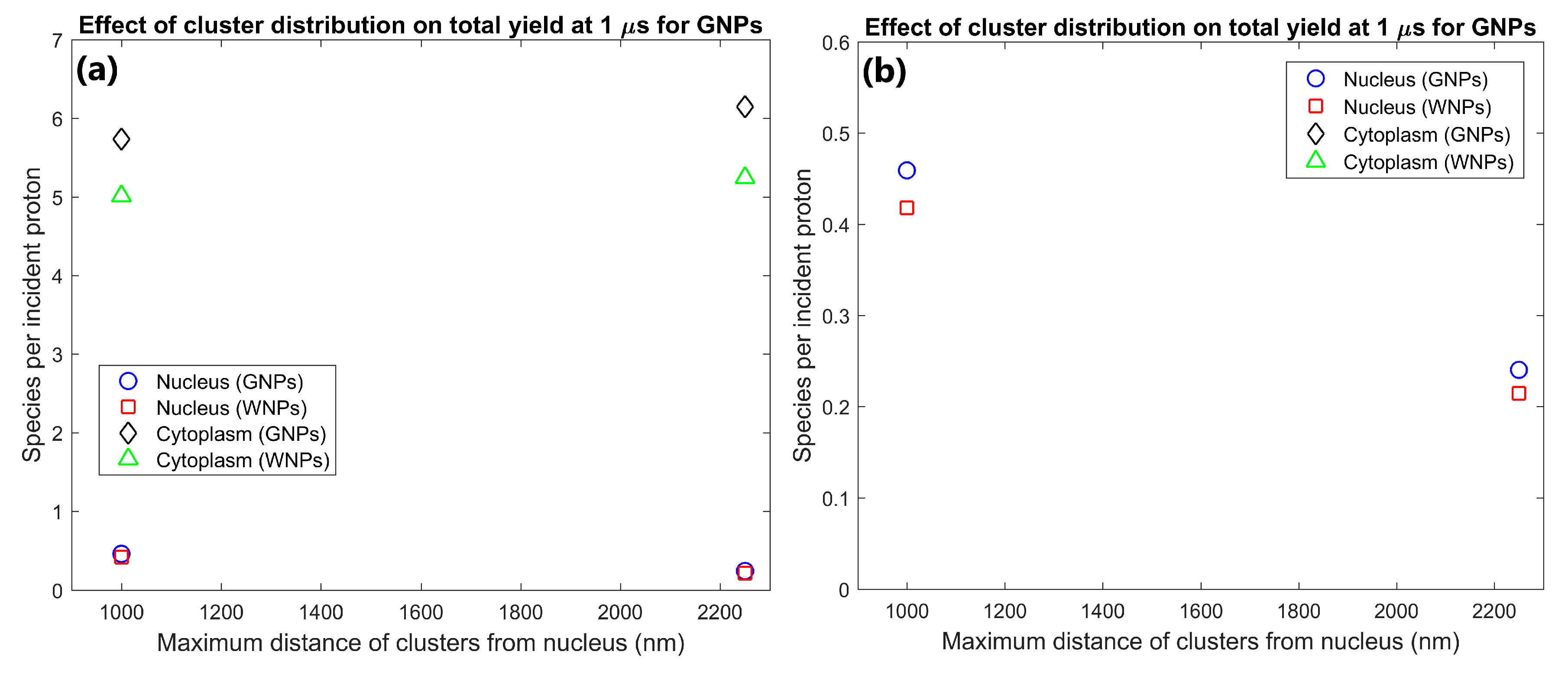
| Cluster distribution | Near nucleus | 4.63 | 64.1 | 0.459 | 5.73 | 1.07 | 1.15 | 1.10 | 1.14 | Clusters near nucleus increased nucleus yield by 91% while reducing cytoplasm yield by 7% compared to a disperse distribution |
| Whole Cytoplasm | 2.47 | 68.4 | 0.240 | 6.15 | 1.09 | 1.18 | 1.12 | 1.17 | ||
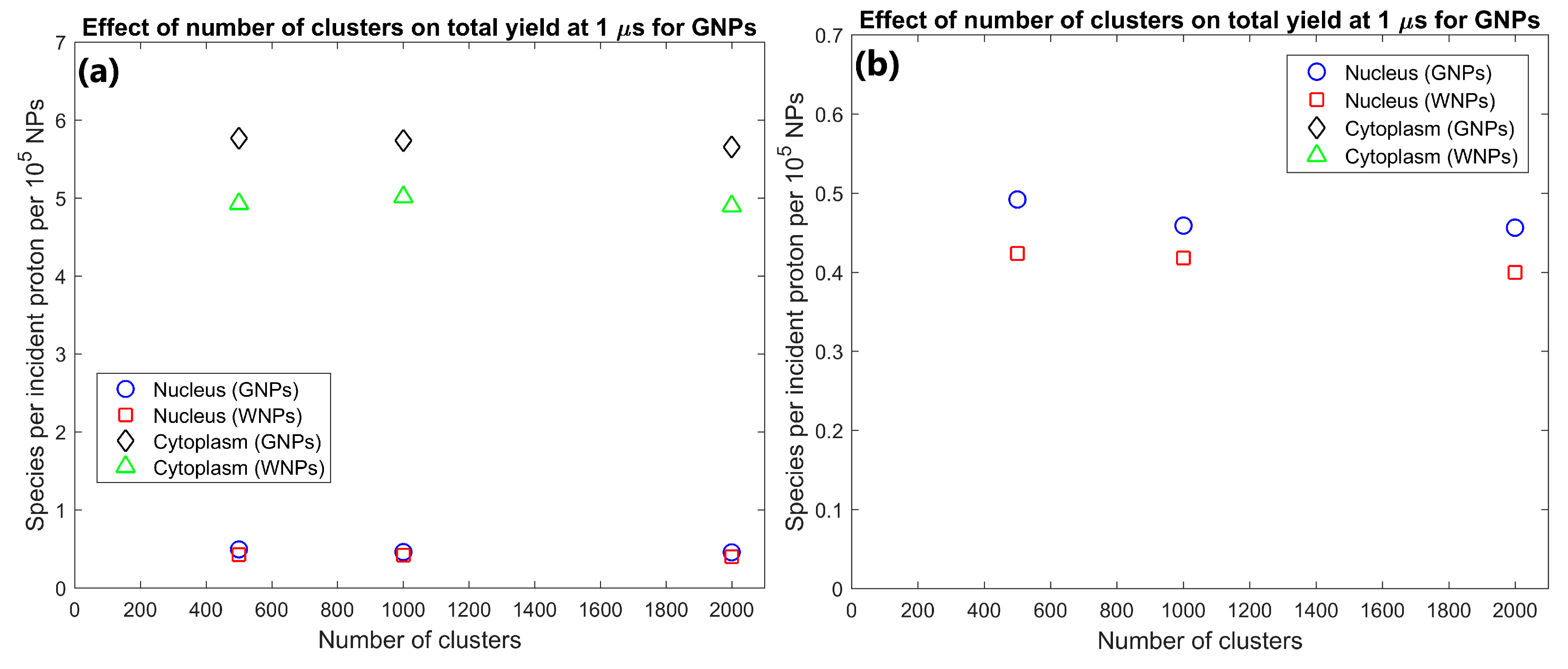
| Number of Clusters | 500 | 2.49 | 32.3 | 0.246 | 2.88 | 1.15 | 1.17 | 1.16 | 1.17 | Dose and yield per GNP changed by less than 7% indicating a good scaling with GNP concentration |
| 1000 | 4.63 | 64.1 | 0.459 | 5.73 | 1.07 | 1.15 | 1.10 | 1.14 | ||
| 2000 | 9.21 | 126.5 | 0.913 | 11.3 | 1.13 | 1.16 | 1.14 | 1.16 | ||
| Simulation | Cluster Diameter (nm) | Total Number of GNPs (104) | Distribution Range from Nucleus (nm) | Nuclear Membrane Permeability (%) | Cell Membrane Permeability (%) | Proton Energy (MeV) |
|---|---|---|---|---|---|---|
| Effect of cluster distribution | 200 | 10 | 1000, 2250 | 100 | 100 | 5 |
| Effect of Cluster size | 100, 200, 500 | 10 | 1000 | 100 | 100 | 5 |
| Effect of GNP concentration | 200 | 5, 10, 20 | 1000 | 100 | 100 | 5 |
| Nuclear membrane absorption | 200 | 10 | 1000 | 0 | 100 | 5 |
| Effect in adjacent cell | 200 | 10 | 1000 | 100 | 100 | 5 |
| Cell membrane absorption | 200 | 10 | 1000 | 100 | 0 | 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peukert, D.; Kempson, I.; Douglass, M.; Bezak, E. Modelling Spatial Scales of Dose Deposition and Radiolysis Products from Gold Nanoparticle Sensitisation of Proton Therapy in A Cell: From Intracellular Structures to Adjacent Cells. Int. J. Mol. Sci. 2020, 21, 4431. https://doi.org/10.3390/ijms21124431
Peukert D, Kempson I, Douglass M, Bezak E. Modelling Spatial Scales of Dose Deposition and Radiolysis Products from Gold Nanoparticle Sensitisation of Proton Therapy in A Cell: From Intracellular Structures to Adjacent Cells. International Journal of Molecular Sciences. 2020; 21(12):4431. https://doi.org/10.3390/ijms21124431
Chicago/Turabian StylePeukert, Dylan, Ivan Kempson, Michael Douglass, and Eva Bezak. 2020. "Modelling Spatial Scales of Dose Deposition and Radiolysis Products from Gold Nanoparticle Sensitisation of Proton Therapy in A Cell: From Intracellular Structures to Adjacent Cells" International Journal of Molecular Sciences 21, no. 12: 4431. https://doi.org/10.3390/ijms21124431
APA StylePeukert, D., Kempson, I., Douglass, M., & Bezak, E. (2020). Modelling Spatial Scales of Dose Deposition and Radiolysis Products from Gold Nanoparticle Sensitisation of Proton Therapy in A Cell: From Intracellular Structures to Adjacent Cells. International Journal of Molecular Sciences, 21(12), 4431. https://doi.org/10.3390/ijms21124431