Corin Overexpression Reduces Myocardial Infarct Size and Modulates Cardiomyocyte Apoptotic Cell Death

 ,
,  ,
,
Abstract
1. Introduction
2. Results
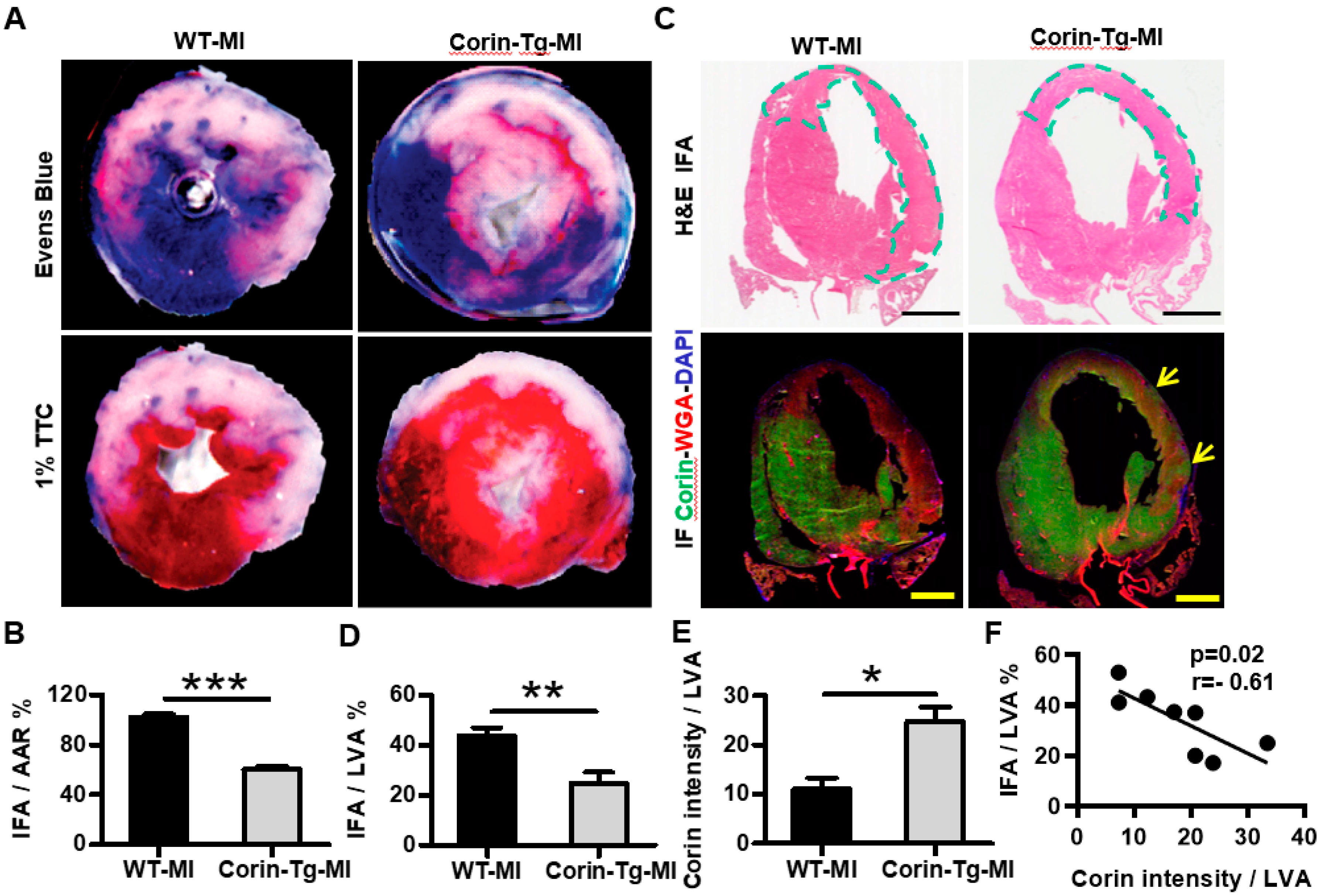
2.1. Cardiac-Specific Overexpression of Corin Reduces Infarct Size
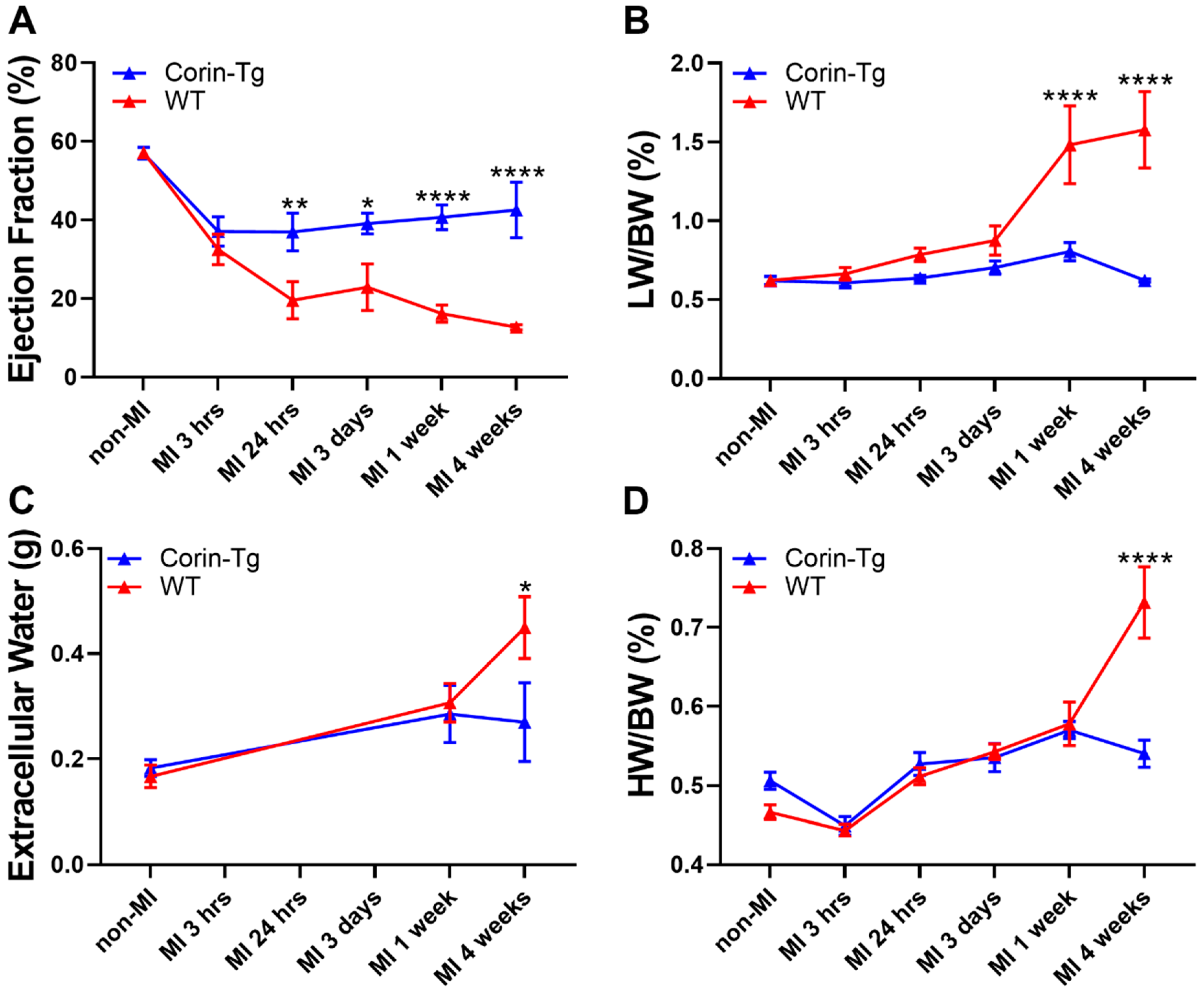
2.2. Cardiac-Specific Overexpression of Corin Significantly Improves Heart Function and Delays Heart Failure Development Post-MI
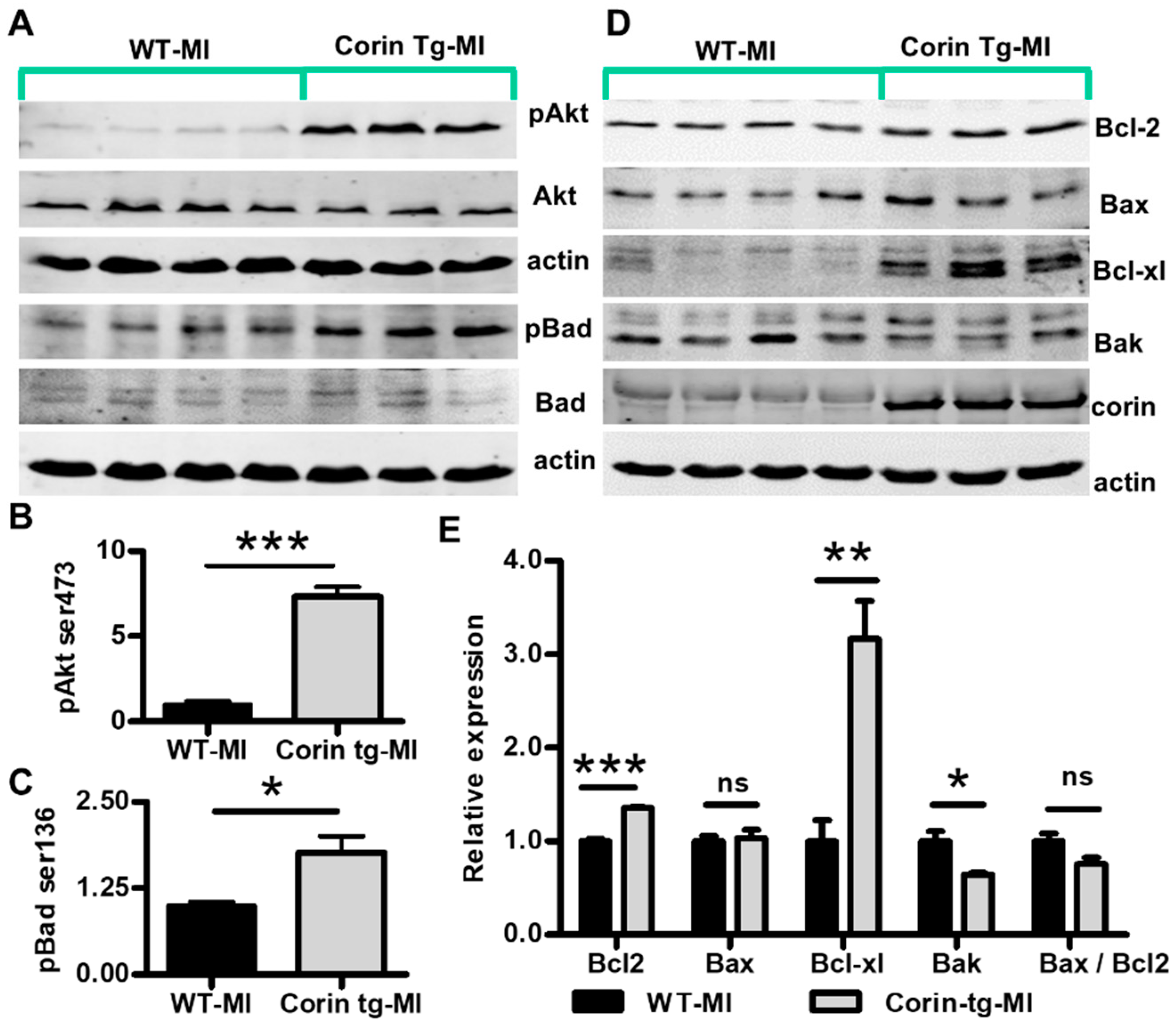
2.3. Cardiac-Specific Overexpression of Corin Attenuates Cardiomyocyte Apoptosis Post-MI
2.4. Corin Overexpression Modulates Bcl-2 Family Proteins in the Infarcted Heart
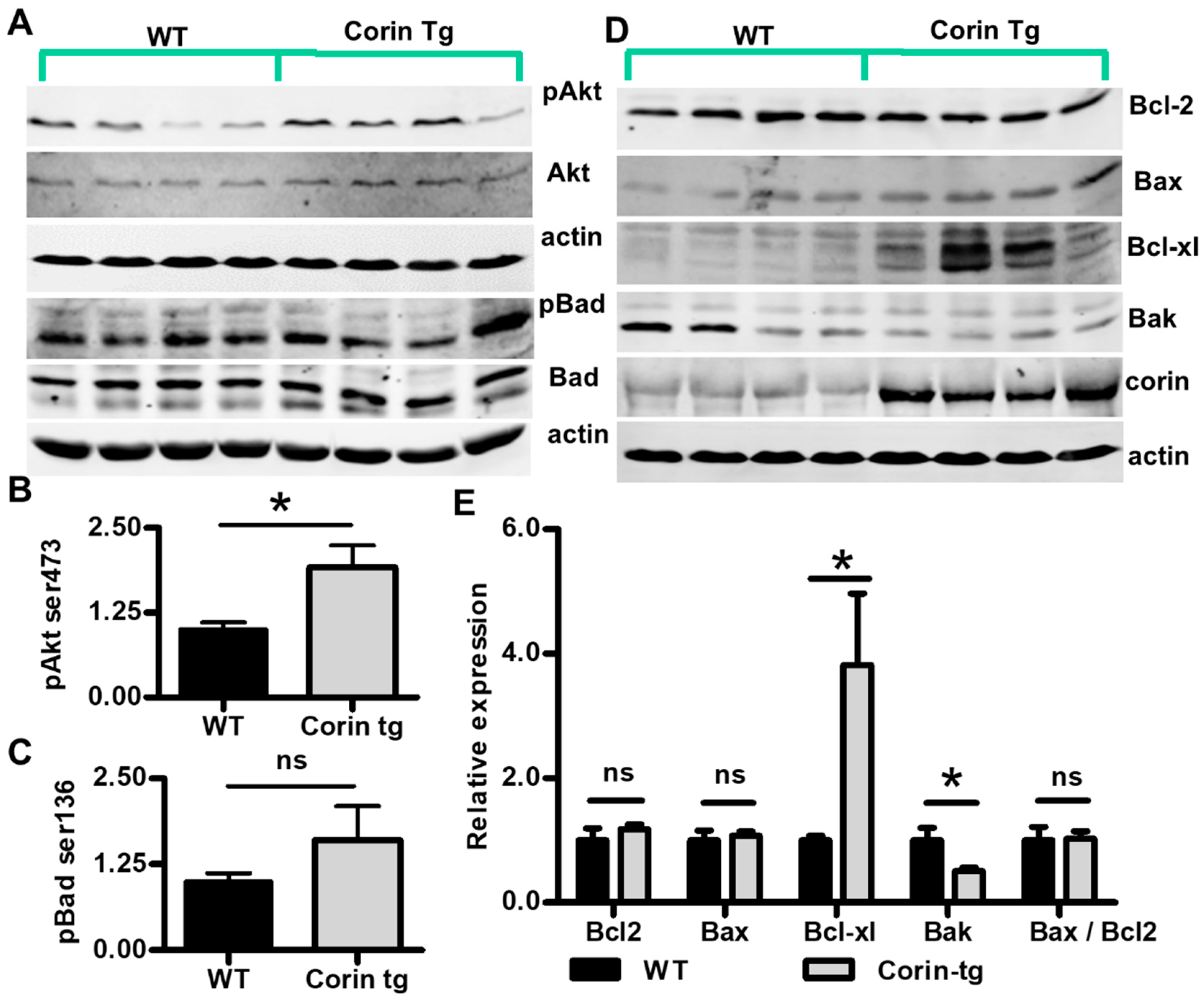
2.5. Corin Overexpression Regulate Bcl-2 Family Proteins under Non-Ischemic Conditions
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Myocardial Infarction Model
4.3. Echocardiography
4.4. Quantitative Magnetic Resonance
4.5. Heart Tissue Lysate Preparation and Western Blot Analysis
4.6. Immunohistological Staining and Analysis
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MI | Myocardial infarction |
| ICM | Ischemic cardiomyopathy |
| DCM | Dilated cardiomyopathy |
| HF | Heart failure |
| WT | Wild-type |
| Corin-Tg | Corin transgenic |
| EF | Ejection fraction |
| FS | Fractional shortening |
| LW/BW | Lung weight to body weight ratio |
| HW/BW | Heart weight to body weight ratio |
| TCC | 2,3,5-triphenyltetrazolium chloride |
| H&E | Hematoxylin and eosin |
| AAR | Area at risk for ischemia |
| IFA | Infarct area |
| IF | Immunofluorescence |
| LV | Left ventricle |
| LVA | Left ventricular area |
| ANP | Atrial natriuretic peptide |
References
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Despres, J.P.; Fullerton, H.J.; et al. Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation 2015. [Google Scholar] [CrossRef]
- Sutton, M.G.; Sharpe, N. Left ventricular remodeling after myocardial infarction: Pathophysiology and therapy. Circulation 2000, 101, 2981–2988. [Google Scholar] [CrossRef]
- Weber, K.T. Aldosterone in congestive heart failure. N. Engl. J. Med. 2001, 345, 1689–1697. [Google Scholar] [CrossRef]
- Pazos-Lopez, P.; Peteiro-Vazquez, J.; Carcia-Campos, A.; Garcia-Bueno, L.; de Torres, J.P.; Castro-Beiras, A. The causes, consequences, and treatment of left or right heart failure. Vasc. Health Risk Manag. 2011, 7, 237–254. [Google Scholar] [CrossRef]
- Armaly, Z.; Assady, S.; Abassi, Z. Corin: A new player in the regulation of salt-water balance and blood pressure. Curr. Opin. Nephrol. Hypertens. 2013, 22, 713–722. [Google Scholar] [CrossRef]
- Dong, N.; Chen, S.; Yang, J.; He, L.; Liu, P.; Zheng, D.; Li, L.; Zhou, Y.; Ruan, C.; Plow, E.; et al. Plasma soluble corin in patients with heart failure. Circ. Heart Fail. 2010, 3, 207–211. [Google Scholar] [CrossRef]
- Ibebuogu, U.N.; Gladysheva, I.P.; Houng, A.K.; Reed, G.L. Decompensated heart failure is associated with reduced corin levels and decreased cleavage of pro-atrial natriuretic peptide. Circ. Heart Fail. 2011, 4, 114–120. [Google Scholar] [CrossRef]
- Lee, R.; Xu, B.; Rame, J.E.; Felkin, L.E.; Barton, P.; Dries, D.L. Regulated inositol-requiring protein 1-dependent decay as a mechanism of corin RNA and protein deficiency in advanced human systolic heart failure. J. Am. Heart Assoc. 2014, 3, e001104. [Google Scholar] [CrossRef]
- Barnet, C.S.; Liu, X.; Body, S.C.; Collard, C.D.; Shernan, S.K.; Muehlschlegel, J.D.; Jarolim, P.; Fox, A.A. Plasma corin decreases after coronary artery bypass graft surgery and is associated with postoperative heart failure: A pilot study. J. Cardiothorac. Vasc. Anesth. 2015, 29, 374–381. [Google Scholar] [CrossRef]
- Rame, J.E.; Tam, S.W.; McNamara, D.; Worcel, M.; Sabolinski, M.L.; Wu, A.H.; Dries, D.L. Dysfunctional corin i555(p568) allele is associated with impaired brain natriuretic peptide processing and adverse outcomes in blacks with systolic heart failure: Results from the Genetic Risk Assessment in Heart Failure substudy. Circ. Heart Fail. 2009, 2, 541–548. [Google Scholar] [CrossRef]
- Feistritzer, H.J.; Klug, G.; Reinstadler, S.J.; Mair, J.; Schocke, M.; Gobel, G.; Franz, W.M.; Metzler, B. Circulating corin concentrations are related to infarct size in patients after ST-segment elevation myocardial infarction. Int. J. Cardiol. 2015, 192, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.M.; Shen, J.X.; Li, H.; Zhao, P.; Xu, G.; Chen, J.C. Association between serum corin levels and risk of acute myocardial infarction. Clin. Chim. Acta 2016, 452, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.; Wang, D.; Sullivan, R.; Fan, T.M.; Gladysheva, I.P.; Reed, G.L. Depressed Corin Levels Indicate Early Systolic Dysfunction Before Increases of Atrial Natriuretic Peptide/B-Type Natriuretic Peptide and Heart Failure Development. Hypertension 2015. [Google Scholar] [CrossRef]
- Gladysheva, I.P.; Wang, D.; McNamee, R.A.; Houng, A.K.; Mohamad, A.A.; Fan, T.M.; Reed, G.L. Corin overexpression improves cardiac function, heart failure, and survival in mice with dilated cardiomyopathy. Hypertension 2013, 61, 327–332. [Google Scholar] [CrossRef]
- Kang, P.M.; Izumo, S. Apoptosis and heart failure: A critical review of the literature. Circ. Res. 2000, 86, 1107–1113. [Google Scholar] [CrossRef]
- Konstantinidis, K.; Whelan, R.S.; Kitsis, R.N. Mechanisms of cell death in heart disease. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1552–1562. [Google Scholar] [CrossRef]
- Abbate, A.; Bussani, R.; Amin, M.S.; Vetrovec, G.W.; Baldi, A. Acute myocardial infarction and heart failure: Role of apoptosis. Int. J. Biochem. Cell Biol. 2006, 38, 1834–1840. [Google Scholar] [CrossRef]
- Krijnen, P.A.; Nijmeijer, R.; Meijer, C.J.; Visser, C.A.; Hack, C.E.; Niessen, H.W. Apoptosis in myocardial ischaemia and infarction. J. Clin. Pathol. 2002, 55, 801–811. [Google Scholar] [CrossRef]
- Wang, D.; Gladysheva, I.P.; Sullivan, R.D.; Fan, T.M.; Mehta, R.M.; Tripathi, R.; Sun, Y.; Reed, G.L. Increases in plasma corin levels following experimental myocardial infarction reflect the severity of ischemic injury. PLoS ONE 2018, 13, e0202571. [Google Scholar] [CrossRef]
- Gill, C.; Mestril, R.; Samali, A. Losing heart: The role of apoptosis in heart disease--a novel therapeutic target? FASEB J. 2002, 16, 135–146. [Google Scholar] [CrossRef]
- Crow, M.T.; Mani, K.; Nam, Y.J.; Kitsis, R.N. The mitochondrial death pathway and cardiac myocyte apoptosis. Circ. Res. 2004, 95, 957–970. [Google Scholar] [CrossRef]
- Anversa, P.; Cheng, W.; Liu, Y.; Leri, A.; Redaelli, G.; Kajstura, J. Apoptosis and myocardial infarction. Basic Res. Cardiol. 1998, 93 (Suppl. 3), 8–12. [Google Scholar] [CrossRef] [PubMed]
- Gomez, L.; Chavanis, N.; Argaud, L.; Chalabreysse, L.; Gateau-Roesch, O.; Ninet, J.; Ovize, M. Fas-independent mitochondrial damage triggers cardiomyocyte death after ischemia-reperfusion. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2153–H2158. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.C. Protein kinase activation and myocardial ischemia/reperfusion injury. Cardiovasc. Res. 2004, 61, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Sussman, M.A.; Volkers, M.; Fischer, K.; Bailey, B.; Cottage, C.T.; Din, S.; Gude, N.; Avitabile, D.; Alvarez, R.; Sundararaman, B.; et al. Myocardial AKT: The omnipresent nexus. Physiol. Rev. 2011, 91, 1023–1070. [Google Scholar] [CrossRef]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef]
- Chiong, M.; Wang, Z.V.; Pedrozo, Z.; Cao, D.J.; Troncoso, R.; Ibacache, M.; Criollo, A.; Nemchenko, A.; Hill, J.A.; Lavandero, S. Cardiomyocyte death: Mechanisms and translational implications. Cell Death Dis. 2011, 2, e244. [Google Scholar] [CrossRef]
- Del Peso, L.; Gonzalez-Garcia, M.; Page, C.; Herrera, R.; Nunez, G. Interleukin-3-induced phosphorylation of BAD through the protein kinase Akt. Science 1997, 278, 687–689. [Google Scholar] [CrossRef]
- Salakou, S.; Kardamakis, D.; Tsamandas, A.C.; Zolota, V.; Apostolakis, E.; Tzelepi, V.; Papathanasopoulos, P.; Bonikos, D.S.; Papapetropoulos, T.; Petsas, T.; et al. Increased Bax/Bcl-2 ratio up-regulates caspase-3 and increases apoptosis in the thymus of patients with myasthenia gravis. In Vivo 2007, 21, 123–132. [Google Scholar]
- Pang, A.; Hu, Y.; Zhou, P.; Long, G.; Tian, X.; Men, L.; Shen, Y.; Liu, Y.; Cui, Y. Corin is down-regulated and exerts cardioprotective action via activating pro-atrial natriuretic peptide pathway in diabetic cardiomyopathy. Cardiovasc. Diabetol. 2015, 14, 134. [Google Scholar] [CrossRef]
- Langenickel, T.H.; Pagel, I.; Buttgereit, J.; Tenner, K.; Lindner, M.; Dietz, R.; Willenbrock, R.; Bader, M. Rat corin gene: Molecular cloning and reduced expression in experimental heart failure. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1516–H1521. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, T.; Boerrigter, G.; Huntley, B.K.; Sangaralingham, S.J.; McKie, P.M.; Harty, G.J.; Harders, G.E.; Burnett, J.C., Jr. Differential expression of the pro-natriuretic peptide convertases corin and furin in experimental heart failure and atrial fibrosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R102–R109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chen, J.; Zhang, Q.; Shao, J.; Du, K.; Xu, X.; Kong, Y. Prognostic Value of Plasma Soluble Corin in Patients with Acute Myocardial Infarction. J. Am. Coll. Cardiol. 2016, 67, 2008–2014. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Doi, D.; Samata, B.; Katsukawa, M.; Kikuchi, T.; Morizane, A.; Ono, Y.; Sekiguchi, K.; Nakagawa, M.; Parmar, M.; Takahashi, J. Isolation of human induced pluripotent stem cell-derived dopaminergic progenitors by cell sorting for successful transplantation. Stem Cell Rep. 2014, 2, 337–350. [Google Scholar] [CrossRef]
- Stone, G.W.; Dixon, S.R.; Grines, C.L.; Cox, D.A.; Webb, J.G.; Brodie, B.R.; Griffin, J.J.; Martin, J.L.; Fahy, M.; Mehran, R.; et al. Predictors of infarct size after primary coronary Angioplasty in acute myocardial infarction from pooled analysis from four contemporary trials. Am. J. Cardiol. 2007, 100, 1370–1375. [Google Scholar] [CrossRef]
- Melenovsky, V.; Andersen, M.J.; Andress, K.; Reddy, Y.N.; Borlaug, B.A. Lung congestion in chronic heart failure: Haemodynamic, clinical, and prognostic implications. Eur. J. Heart Fail. 2015, 17, 1161–1171. [Google Scholar] [CrossRef]
- Page, B.J.; Banas, M.D.; Suzuki, G.; Weil, B.R.; Young, R.F.; Fallavollita, J.A.; Palka, B.A.; Canty, J.M., Jr. Revascularization of chronic hibernating myocardium stimulates myocyte proliferation and partially reverses chronic adaptations to ischemia. J. Am. Coll. Cardiol. 2015, 65, 684–697. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Zhang, X.; Wu, C.; Lange, R.A.; Chilton, R.J.; Lindsey, M.L.; Jin, Y.F. Integrative computational and experimental approaches to establish a post-myocardial infarction knowledge map. PLoS Comput. Biol. 2014, 10, e1003472. [Google Scholar] [CrossRef]
- Lyn, D.; Bao, S.; Bennett, N.A.; Liu, X.; Emmett, N.L. Ischemia elicits a coordinated expression of pro-survival proteins in mouse myocardium. Sci. World J. 2002, 2, 997–1003. [Google Scholar] [CrossRef]
- Matsui, T.; Tao, J.; del Monte, F.; Lee, K.H.; Li, L.; Picard, M.; Force, T.L.; Franke, T.F.; Hajjar, R.J.; Rosenzweig, A. Akt activation preserves cardiac function and prevents injury after transient cardiac ischemia in vivo. Circulation 2001, 104, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.W.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Whelan, R.S.; Kaplinskiy, V.; Kitsis, R.N. Cell death in the pathogenesis of heart disease: Mechanisms and significance. Annu. Rev. Physiol. 2010, 72, 19–44. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ito, Y.; Morikawa, M.; Uchida, H.; Kobune, M.; Sasaki, K.; Abe, T.; Hamada, H. Bcl-xL gene transfer protects the heart against ischemia/reperfusion injury. Biochem. Biophys. Res. Commun. 2003, 311, 64–70. [Google Scholar] [CrossRef]
- Brocheriou, V.; Hagege, A.A.; Oubenaissa, A.; Lambert, M.; Mallet, V.O.; Duriez, M.; Wassef, M.; Kahn, A.; Menasche, P.; Gilgenkrantz, H. Cardiac functional improvement by a human Bcl-2 transgene in a mouse model of ischemia/reperfusion injury. J. Gene Med. 2000, 2, 326–333. [Google Scholar] [CrossRef]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef]
- Kerkela, R.; Ulvila, J.; Magga, J. Natriuretic Peptides in the Regulation of Cardiovascular Physiology and Metabolic Events. J. Am. Heart Assoc. 2015, 4. [Google Scholar] [CrossRef]
- Houng, A.K.; McNamee, R.A.; Kerner, A.; Sharma, P.; Mohamad, A.; Tronolone, J.; Reed, G.L. Atrial natriuretic peptide increases inflammation, infarct size, and mortality after experimental coronary occlusion. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H655–H661. [Google Scholar] [CrossRef]
- Sullivan, R.D.; Mehta, R.M.; Tripathi, R.; Gladysheva, I.P.; Reed, G.L. Normalizing Plasma Renin Activity in Experimental Dilated Cardiomyopathy: Effects on Edema, Cachexia, and Survival. Int. J. Mol. Sci. 2019, 20, 3886. [Google Scholar] [CrossRef]
- Tripathi, R.; Sullivan, R.D.; Fan, T.M.; Houng, A.K.; Mehta, R.M.; Reed, G.L.; Gladysheva, I.P. Cardiac-Specific Overexpression of Catalytically Inactive Corin Reduces Edema, Contractile Dysfunction, and Death in Mice with Dilated Cardiomyopathy. Int. J. Mol. Sci. 2019, 21, 203. [Google Scholar] [CrossRef]
- Tripathi, R.; Sullivan, R.; Fan, T.M.; Wang, D.; Sun, Y.; Reed, G.L.; Gladysheva, I.P. Enhanced heart failure, mortality and renin activation in female mice with experimental dilated cardiomyopathy. PLoS ONE 2017, 12, e0189315. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gladysheva, I.P.; Fan, T.H.; Sullivan, R.; Houng, A.K.; Reed, G.L. Atrial natriuretic peptide affects cardiac remodeling, function, heart failure, and survival in a mouse model of dilated cardiomyopathy. Hypertension 2014, 63, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, I.P.; Robinson, B.R.; Houng, A.K.; Kovats, T.; King, S.M. Corin is co-expressed with pro-ANP and localized on the cardiomyocyte surface in both zymogen and catalytically active forms. J. Mol. Cell Cardiol. 2008, 44, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, I.P.; King, S.M.; Houng, A.K. N-glycosylation modulates the cell-surface expression and catalytic activity of corin. Biochem. Biophys. Res. Commun. 2008, 373, 130–135. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Company | Cat # | Host Species | Clonality | Dilution |
|---|---|---|---|---|---|
| cleaved caspase-3 | Cell Signaling | 9661s | rabbit | polyclonal | 1:1000 |
| caspase-3 | Santa Cruz Biotechnology | sc-7148 | rabbit | polyclonal | 1:400 |
| cleaved caspase-9 | Cell Signaling | 9509s | rabbit | polyclonal | 1:1000 |
| caspase-9 | Cell Signaling | 9508s | mouse | monoclonal | 1:1000 |
| cleaved caspase-8 | Cell Signaling | 8592s | rabbit | monoclonal | 1:1000 |
| caspase-8 | Enzo Life Science | ALX-804-447-C100 | Rat | monoclonal | 1:500 |
| actin | Santa Cruz Biotechnology | sc-1616 | Goat | polyclonal | 1:500 |
| phospho-Akt (Ser473) | Cell Signaling | 4060s | rabbit | monoclonal | 1:1000 |
| Akt1/2/3 | Santa Cruz Biotechnology | sc-8312 | rabbit | polyclonal | 1:500 |
| phospho-Bad (Ser136) | Cell Signaling | 4366s | rabbit | monoclonal | 1:500 |
| bad | Santa Cruz Biotechnology | sc-943 | rabbit | polyclonal | 1:500 |
| Bcl2 | Santa Cruz Biotechnology | sc-492 | rabbit | polyclonal | 1:500 |
| Bax | Santa Cruz Biotechnology | sc-493 | rabbit | polyclonal | 1:500 |
| Bak | Santa Cruz Biotechnology | sc-832 | rabbit | polyclonal | 1:500 |
| Bcl-xL | Santa Cruz Biotechnology | sc-8392 | mouse | monoclonal | 1:500 |
| corin | Lab Generated [53,54] | N/A | rabbit | polyclonal | 1:7000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, R.D.; Houng, A.K.; Gladysheva, I.P.; Fan, T.-H.M.; Tripathi, R.; Reed, G.L.; Wang, D. Corin Overexpression Reduces Myocardial Infarct Size and Modulates Cardiomyocyte Apoptotic Cell Death. Int. J. Mol. Sci. 2020, 21, 3456. https://doi.org/10.3390/ijms21103456
Sullivan RD, Houng AK, Gladysheva IP, Fan T-HM, Tripathi R, Reed GL, Wang D. Corin Overexpression Reduces Myocardial Infarct Size and Modulates Cardiomyocyte Apoptotic Cell Death. International Journal of Molecular Sciences. 2020; 21(10):3456. https://doi.org/10.3390/ijms21103456
Chicago/Turabian StyleSullivan, Ryan D., Aiilyan K. Houng, Inna P. Gladysheva, Tai-Hwang M. Fan, Ranjana Tripathi, Guy L. Reed, and Dong Wang. 2020. "Corin Overexpression Reduces Myocardial Infarct Size and Modulates Cardiomyocyte Apoptotic Cell Death" International Journal of Molecular Sciences 21, no. 10: 3456. https://doi.org/10.3390/ijms21103456
APA StyleSullivan, R. D., Houng, A. K., Gladysheva, I. P., Fan, T.-H. M., Tripathi, R., Reed, G. L., & Wang, D. (2020). Corin Overexpression Reduces Myocardial Infarct Size and Modulates Cardiomyocyte Apoptotic Cell Death. International Journal of Molecular Sciences, 21(10), 3456. https://doi.org/10.3390/ijms21103456