Swiprosin-1/EFhD-2 Expression in Cardiac Remodeling and Post-Infarct Repair: Effect of Ischemic Conditioning

 , , ,
, , , 

Abstract
1. Introduction
2. Results
2.1. Swiprosin-1 Expression In Post-Infarcted Mice
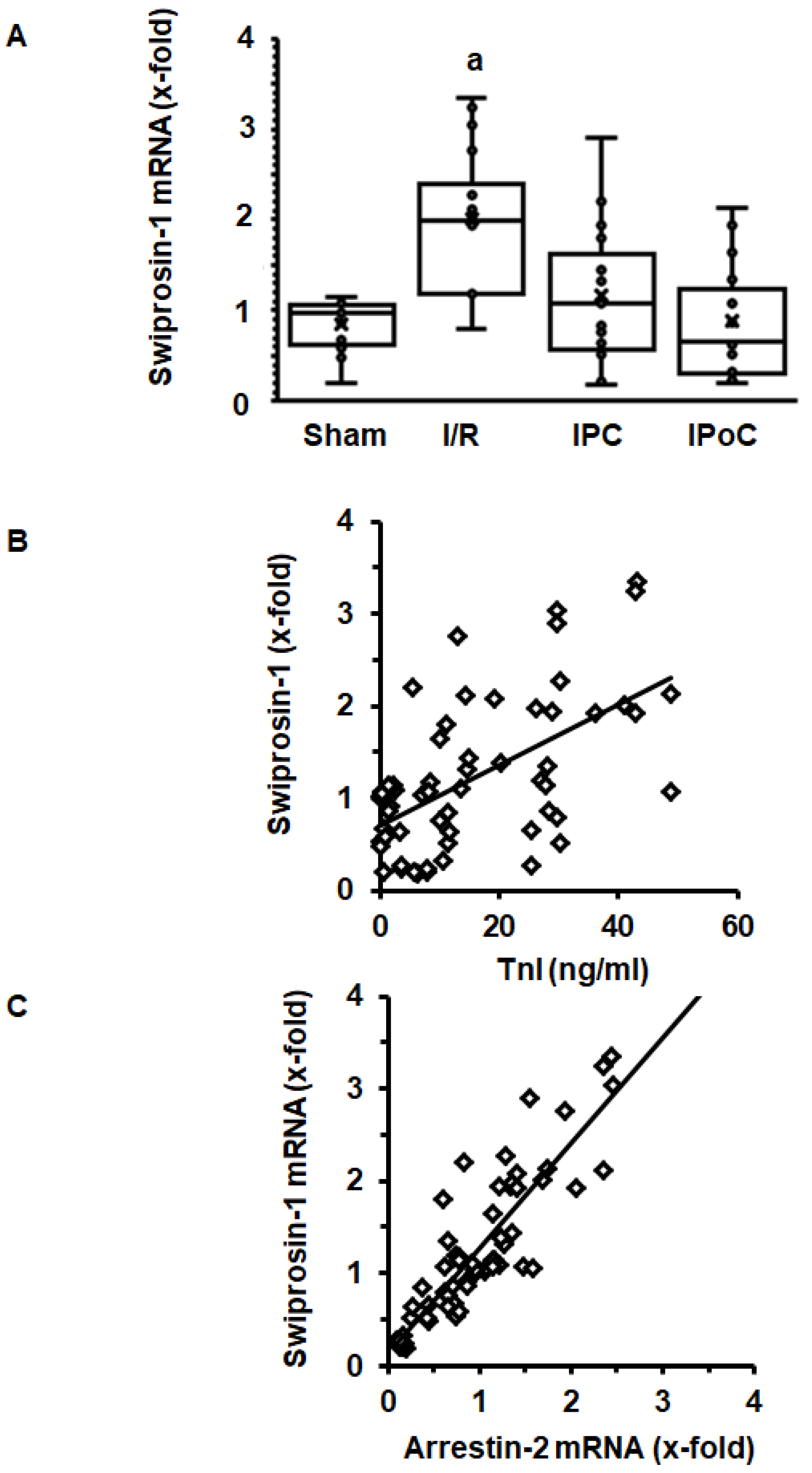
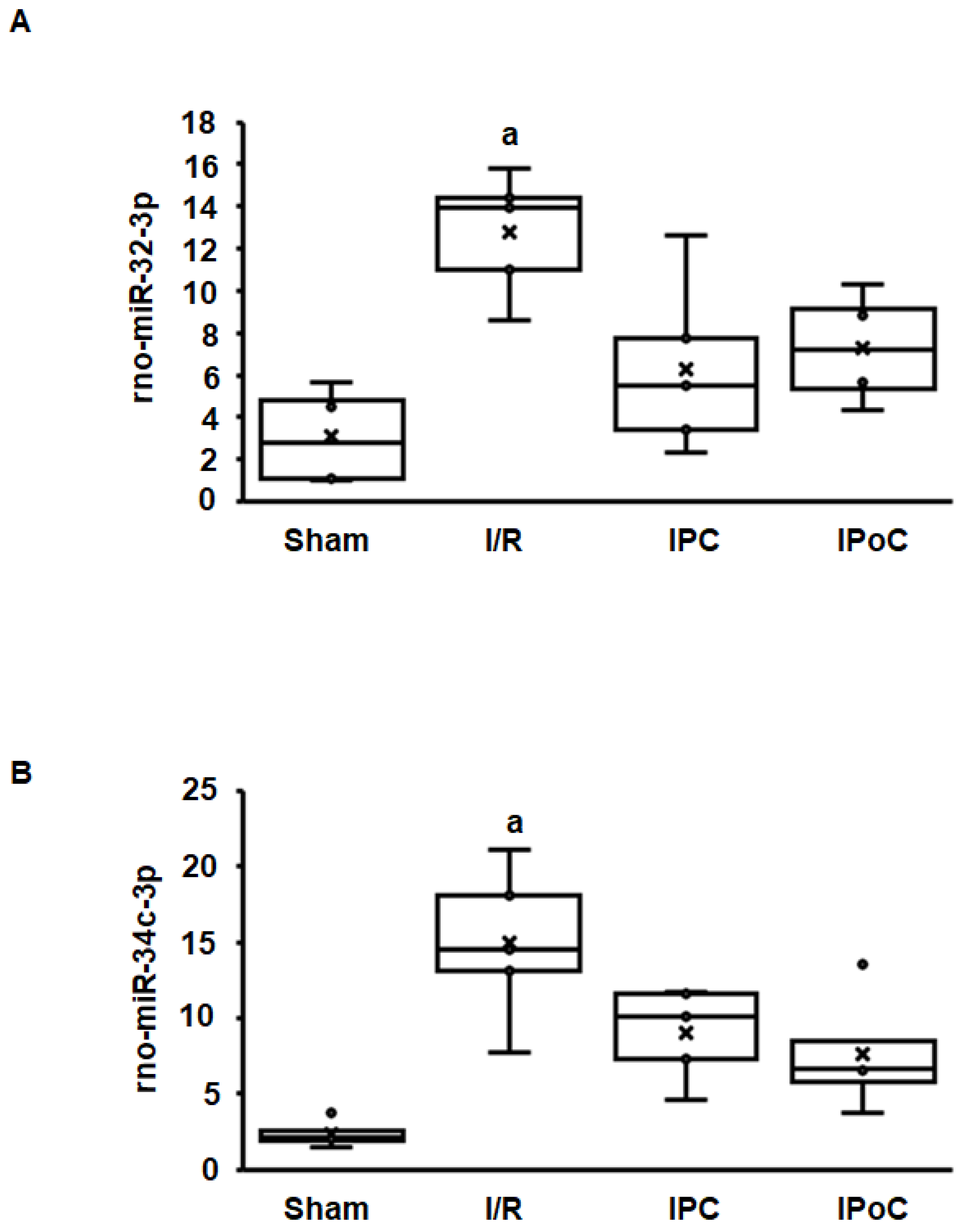
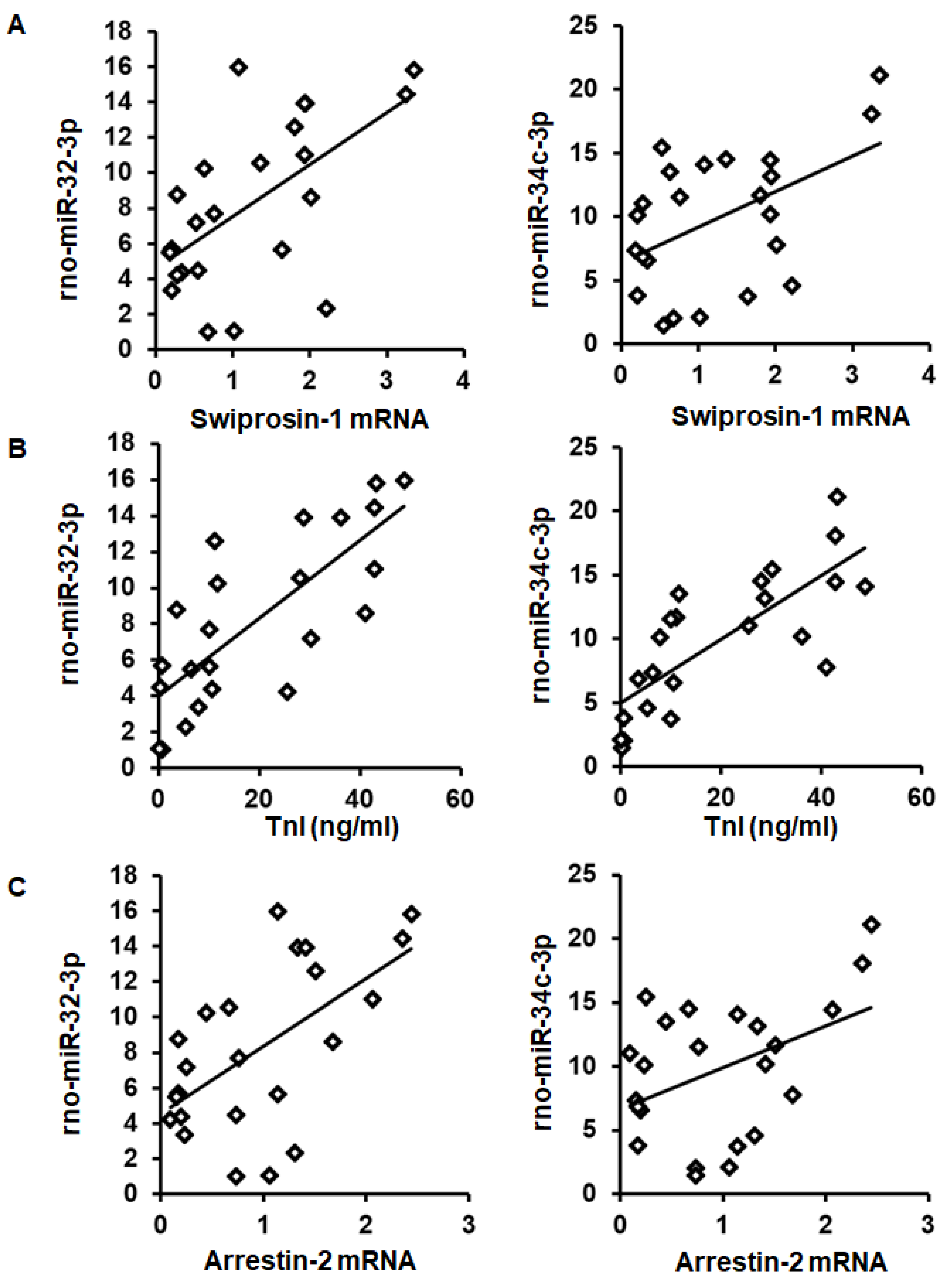
2.2. Induction of Swiprosin-1 in Rat Hearts after Ischemia/Reperfusion
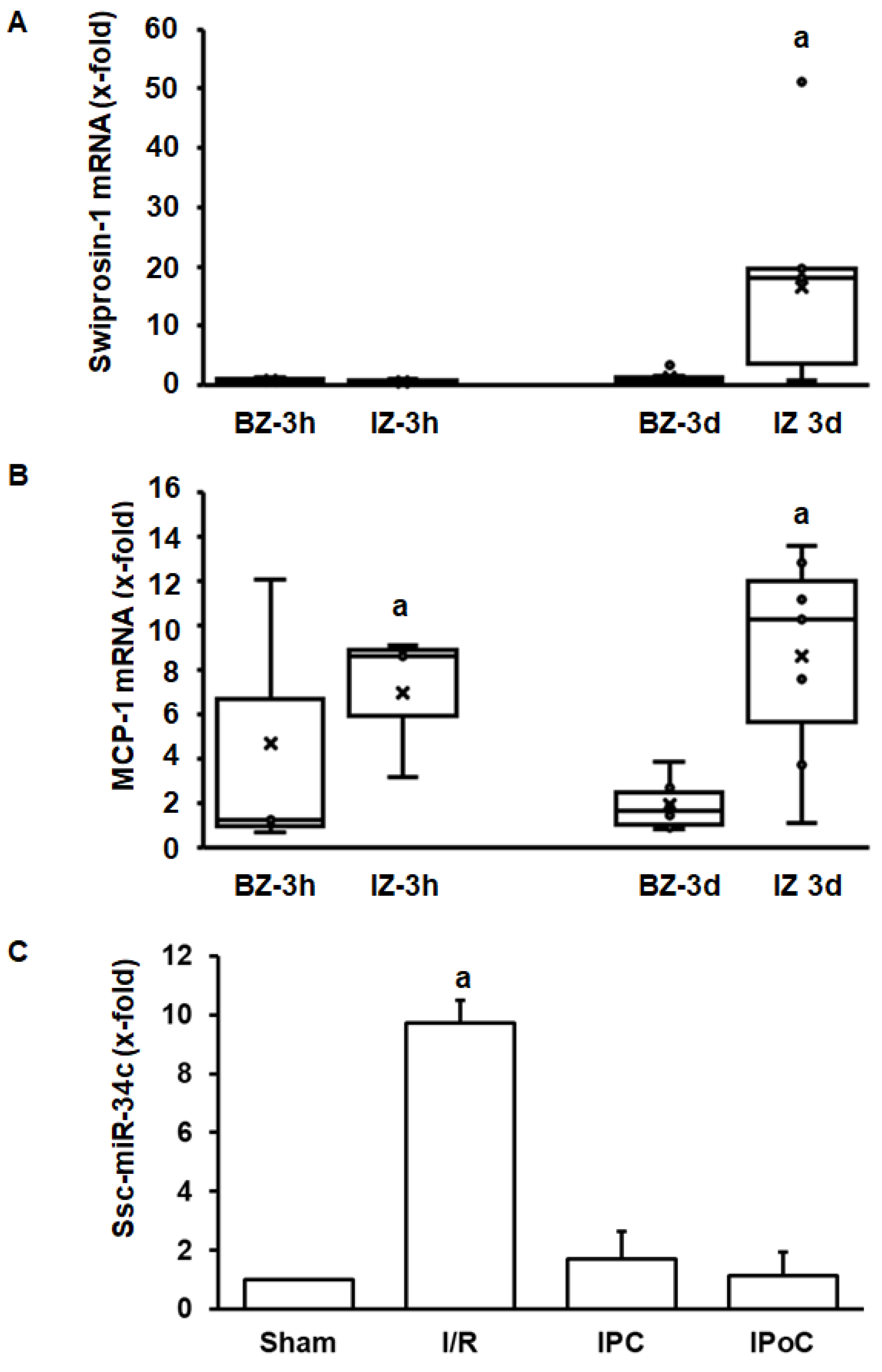
2.3. Regulation of Swiprosin-1 Expression in Pig Hearts
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Western Blot and Histology on Mouse Samples
4.3. RT-PCR on Rat and Pig Tissue
4.4. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IPC | Ischemic Preconditioning |
| IPoC | Ischemic Postconditioning |
| I/R | Ischemia/Reperfusion |
| LAD MI | Left anterior descending coronary artery Myocardial Infarction |
References
- Mielenz, D.; Gunn-Moore, F. Physiological and pathophysiological functions of Swiprosin-1/ EFhd2 in the nervous system. Biochem. J. 2016, 476, 2429–2437. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huh, Y.H.; Kim, S.H.; Chung, K.H.; Oh, S.; Kwon, M.S.; Choi, H.W.; Rhee, S.; Ryu, J.H.; Park, Z.Y.; Jun, C.D.; et al. Swiprosin-1 modulates actin dynamics by regulating the F-actin accessibility to cofilin. Cell. Mol. Life Sci. 2013, 70, 4841–4854. [Google Scholar] [CrossRef] [PubMed]
- Vuadens, F.; Rufer, N.; Kress, A.; Corthesy, P.; Schneider, P.; Tissot, J.D. Identification of swiprosin 1 in human lymphocytes. Proteomics 2014, 4, 2216–2220. [Google Scholar] [CrossRef] [PubMed]
- Mielenz, D.; Vettermann, C.; Hampel, M.; Lang, C.; Avramidou, A.; Karas, M.; Jäck, H.M. Lipid rafts associate with intracellular B cell receptors and exhibit a B cell stage-specific protein composition. J. Immunol. 2005, 175, 3508–3517. [Google Scholar] [CrossRef]
- Thylur, R.P.; Kim, Y.D.; Kwon, M.S.; Oh, H.M.; Kwon, H.K.; Kim, S.H.; Im, S.H.; Chun, J.S.; Park, Z.Y.; Jun, C.D. Swiprosin-a is expressed in mast cells and up-regulated through the protein kinase C beta I/eta pathway. J. Cell. Biochem. 2009, 108, 705–715. [Google Scholar] [CrossRef]
- Kim, Y.D.; Kwon, M.S.; Na, B.R.; Kim, H.R.; Lee, H.S.; Jun, C.D. Swiprosin-1 expression is up-regulated through protein kinase C-ϴ and NF-κB pathway in T cells. Immune Netw. 2013, 13, 55–62. [Google Scholar] [CrossRef][Green Version]
- Purohit, P.; Perez-Branguli, F.; Prots, I.; Borger, E.; Gunn-Moore, F.; Welzel, O.; Loy, K.; Wenzel, E.M.; Grömer, T.W.; Brachs, S.; et al. The Ca2+ Sensor Protein Swiprosin-1/EFhd2 Is Present in Neurites and Involved in Kinesin-Mediated Transport in Neurons. PLoS ONE 2014, 18, e103976. [Google Scholar] [CrossRef]
- Wang, R.M.; Wang, Z.B.; Wang, Y.; Liu, W.Y.; Li, Y.; Tong, L.C.; Zhang, S.; Su, D.F.; Cao, Y.B.; Li, L.; et al. Swiprosin-1 promotes mitochondrial-dependent apoptosis of glomerular podocytes via p38 MAPK pathway in early-stage diabetic nephropathy. Cell. Physiol. Biochem. 2018, 45, 899–916. [Google Scholar] [CrossRef]
- Nippert, F.; Schreckenberg, R.; Hess, A.; Weber, M.; Schlüter, K.-D. The effects of swiprosin-1 on the formation pf pseudopodia-like structures and β-adrenoceptor coupling in cultured adult rat ventricular cardiomyocytes. PLoS ONE 2016, 11, e0167655. [Google Scholar] [CrossRef]
- Hombruch-Freitag, C.; Griemert, B.; Buttgereit, D.; Renkawitz-Pohl, R. Drosophila swiprosin-1/EFHD2 accumulates at the perfusion complex stage during Drosophila myoblast fusion. J. Cell Sci. 2011, 124, 3266–3278. [Google Scholar] [CrossRef]
- Tu, Y.; Zhang, L.; Tong, L.; Wang, Y.; Zhang, S.; Wang, R.; Li, L.; Wang, Z. EFhd2/swiprosin-1 regulates LPS-induced macrophage recruitment via enhancing actin polymerization and cell migration. Int. Immunopharmacol. 2018, 55, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Nippert, F.; Schreckenberg, R.; Schlüter, K.-D. Isolation and cultivation of adult rat cardiomyocytes. J. Vis. Exp. 2017, 128, e56634. [Google Scholar] [CrossRef] [PubMed]
- Perrino, C.; Barabasi, A.L.; Condorelli, G.; Davidson, S.M.; de Windt, L.; Dimmeler, S.; Engel, F.B.; Hausenloy, D.J.; Hill, J.A.; van Laake, L.W.; et al. Epigenomic and transcriptomic approaches in the post-genomic era: Path to novel targest for diagnosis and therapy of the ischaemic heart? Position paper of the European society of cardiology working group on cellular biology of the heart. Cardiovasc. Res. 2017, 113, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Shamagian, G.; Madonna, R.; Taylor, D.; Climent, A.M.; Prosper, F.; Bras-Rosario, L.; Bayes-Genis, A.; Ferdinandy, P.; Fernandez-Aviles, F.; Izpisua Belmonte, J.C.; et al. Perspectives on directions and priorities for future studies in regenerative medicine. Circ. Res. 2019, 124, 938–951. [Google Scholar] [CrossRef]
- Madonna, R.; van Laake, L.W.; Davidson, S.M.; Engel, F.B.; Hausenloy, D.J.; Lecour, S.; Leor, J.; Perrino, C.; Schulz, R.; Ytrehus, K.; et al. Position paper of the European society of cardiology working group cellular biology on the heart: Cell-based therapies for myocardial repair and regeneration in ischemic heart disease and heart failure. Eur. Heart J. 2016, 37, 1789–1798. [Google Scholar] [CrossRef]
- Schreckenberg, R.; Bencsik, P.; Weber, M.; Abdallah, Y.; Csonka, C.; Gömöri, K.; Kiss, K.; Paloczi, J.; Pipis, J.; Sarközy, M.; et al. Adverse effects on β-adrenergic receptor coupling: Ischemic postconditioning failed to preserve long-term cardiac function. J. Am. Heart Assoc. 2017, 6, e006809. [Google Scholar] [CrossRef]
- Schreckenberg, R.; Klein, J.; Kutsche, H.S.; Schulz, R.; Gömöri, K.; Bencsik, P.; Bencsik, B.; Agg, B.; Saghy, E.; Ferdinandy, P.; et al. Ischemic post-conditioning in rats: Responder and nonresponder differ in transcriptome of mitochondrial proteins. J. Cell. Mol. Med. 2020, in press. [Google Scholar] [CrossRef]
- Baranyai, T.; Giricz, Z.; Varga, Z.V.; Koncsos, G.; Lukovic, D.; Makkos, A.; Sarközy, M.; Pavo, N.; Jakab, A.; Czimbalmos, C.; et al. In vivo MRI and ex vivo histology assessment of the cardioprotection induced by ischemic preconditioning, postconditioning and remote conditioning in a closed-chest procine model of reperfused acute myocardial infarction: Importance of microvasculature. J. Transl. Med. 2017, 15, 67. [Google Scholar] [CrossRef]
- Lukovic, D.; Gugerell, A.; Zlabinger, K.; Winkler, J.; Pavo, N.; Baranyai, T.; Giricz, Z.; Varga, Z.V.; Riesenhuber, M.; Spannbauer, A.; et al. Transcriptional Alterations by Ischaemic Postconditioning in a Pig Infarction Model: Impact on Microvascular Protection. Int. J. Mol. Sci. 2019, 20, 344. [Google Scholar] [CrossRef]
- Westermann, D.; Mersmann, J.; Melchior, A.; Freudenberger, T.; Patrik, C.; Schaefer, L.; Lüllmann-Rauch, R.; Letaau, O.; Jacoiby, C.; Schrader, J.; et al. Biglycan is required for adaptive remodeling after myocardial infarction. Circulation 2008, 117, 1269–1276. [Google Scholar] [CrossRef]
- Frangogiannis, N. The extracellular matrix in ischemic and nonischemic heart failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef]
- Schlier, A.; Schreckenberg, R.; Abdallah, Y.; Krasteva, G.; Piper, H.M.; Pfeil, U.; Kummer, W.; Schlüter, K.-D. CGRP-α responsiveness of adult rat ventricular cardiomyocytes from normotensive and spontaneously hypertensive rats. Eur. J. Cell Biol. 2009, 88, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Kutsche, H.S.; Schreckenberg, R.; Weber, M.; Hirschhäuser, C.; Rohrbach, S.; Li, L.; Niemann, B.; Schulz, R.; Schlüter, K.-D. Alteration in glucose metabolism during the transition to heart failure: The contribution of UCP-2. Cells 2020, 9, 552. [Google Scholar] [CrossRef] [PubMed]
- Lörchner, H.; Pöling, J.; Gajawada, P.; Hou, Y.; Polyakova, V.; Kostin, S.; Adrian-Segarra, J.M.; Boettger, T.; Wietelmann, A.; Warnecke, H.; et al. Myocardial healing requires Reg3β-dependent accumulation of macrophages in the ischemic heart. Nat. Med. 2015, 21, 353–362. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rno-miR | R-Coefficient | p-Value | I/R | IPC/IPoC |
|---|---|---|---|---|
| Positive Correlation | ||||
| 32-3p | r = 0.596 | p = 0.003 | yes | yes |
| 34c-3p | r = 0.495 | p = 0.016 | yes | yes |
| 18a-3p | r = 0.465 | p = 0.025 | yes | no |
| 23b-5p | r = 0.475 | p = 0.022 | yes | no |
| 27b-5p | r = 0.436 | p = 0.038 | yes | no |
| 100-3p | r = 0.431 | p = 0.040 | no | - |
| 146a-5p | r = 0.454 | p = 0.029 | no | - |
| 296-5p | r = 0.474 | p = 0.022 | no | - |
| 324-5p | r = 0.520 | p = 0.011 | no | - |
| 342-3p | r = 0.516 | p = 0.012 | no | - |
| 455-3p | r = 0.453 | p = 0.030 | no | - |
| 497-5p | r = 0.458 | p = 0.028 | no | - |
| 505-5p | r = 0.436 | p = 0.038 | no | - |
| 532-3p | r = 0.514 | p = 0.012 | no | - |
| 674-5p | r = 0.616 | p = 0.002 | no | - |
| Negative Correlation | ||||
| 7b | ||||
| 24-2-5p | r = -0.475 | p = 0.022 | no | - |
| 29b-3p | r = -0.425 | p = 0.043 | yes | no |
| 33-5p | r = -0.563 | p = 0.005 | yes | no |
| 99a-5p | r = -0.431 | p = 0.040 | yes | no |
| 145-3p | r = -0.520 | p = 0.011 | yes | no |
| 150-5p | r = -0.414 | p = 0.050 | yes | no |
| 186-5p | r = -0.425 | p = 0.043 | no | - |
| 194-5p | r = -0.495 | p = 0.016 | yes | no |
| 339-3p | r = -0.447 | p = 0.033 | yes | no |
| 345-3p | r = -0.426 | p = 0.042 | no | - |
| 3068-5p | r = -0.422 | p = 0.045 | yes | no |
| rno-miR-32-3p | ||
| Swiprosin-1 | r = 0.596 | p = 0.003 |
| Arrestin-2 | r = 0.585 | p = 0.003 |
| Biglycan | ||
| Collagen-1 | r = 0.581 | p = 0.004 |
| Collagen-3 | r = 0.515 | p = 0.020 |
| Fibronectin | r = 0.452 | p = 0.030 |
| Arginase-2 | r = 0.434 | p = 0.038 |
| iNOS | ||
| RAMP-3 | ||
| MHC-α | ||
| NCX | ||
| Troponin T | r= -0.470 | p = 0.023 |
| HADHA | ||
| UCP-2 | ||
| Slc5a3 | ||
| rno-miR-34c-3p | ||
| Swiprosin-1 | r = 0.495 | p = 0.016 |
| Arrestin-2 | r = 0.440 | p = 0.036 |
| ANP | ||
| MHC-α | ||
| Elfn2 | ||
| Kcnh5 | ||
| Slc5a3 | ||
| Socs7 | ||
| Beta-2-Mircoglobulin | forward: CGT GGC CTT GGT CCT GCT CG | reverse: TCC GTT TTC CGC TGG TGT GC |
| EFhD2 | forward: TCC GGG AGT TCC TCC TGA TT | reverse: AAG CTC TTC GCT CCC TTG AC |
| MCP-1 | forward: CAG CCA CCT TVT GCA CCC AGG | Reverse: CAC AGA TCT CCT TGCCCG CGA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giricz, Z.; Makkos, A.; Schreckenberg, R.; Pöling, J.; Lörchner, H.; Kiss, K.; Bencsik, P.; Braun, T.; Schulz, R.; Ferdinandy, P.; et al. Swiprosin-1/EFhD-2 Expression in Cardiac Remodeling and Post-Infarct Repair: Effect of Ischemic Conditioning. Int. J. Mol. Sci. 2020, 21, 3359. https://doi.org/10.3390/ijms21093359
Giricz Z, Makkos A, Schreckenberg R, Pöling J, Lörchner H, Kiss K, Bencsik P, Braun T, Schulz R, Ferdinandy P, et al. Swiprosin-1/EFhD-2 Expression in Cardiac Remodeling and Post-Infarct Repair: Effect of Ischemic Conditioning. International Journal of Molecular Sciences. 2020; 21(9):3359. https://doi.org/10.3390/ijms21093359
Chicago/Turabian StyleGiricz, Zoltán, András Makkos, Rolf Schreckenberg, Jochen Pöling, Holger Lörchner, Krisztina Kiss, Péter Bencsik, Thomas Braun, Rainer Schulz, Péter Ferdinandy, and et al. 2020. "Swiprosin-1/EFhD-2 Expression in Cardiac Remodeling and Post-Infarct Repair: Effect of Ischemic Conditioning" International Journal of Molecular Sciences 21, no. 9: 3359. https://doi.org/10.3390/ijms21093359
APA StyleGiricz, Z., Makkos, A., Schreckenberg, R., Pöling, J., Lörchner, H., Kiss, K., Bencsik, P., Braun, T., Schulz, R., Ferdinandy, P., & Schlüter, K.-D. (2020). Swiprosin-1/EFhD-2 Expression in Cardiac Remodeling and Post-Infarct Repair: Effect of Ischemic Conditioning. International Journal of Molecular Sciences, 21(9), 3359. https://doi.org/10.3390/ijms21093359