Influence of Platelet-Rich and Platelet-Poor Plasma on Endogenous Mechanisms of Skeletal Muscle Repair/Regeneration


{kind=link}
Abstract
:1. Introduction
2. Adult Skeletal Muscle Repair and Regeneration: Role of Satellite and Non-Satellite Cells
3. Plasma Preparations: Platelet-Rich Plasma (PRP) and Platelet-Poor Plasma (PPP)
3.1. Definition and Biological Properties
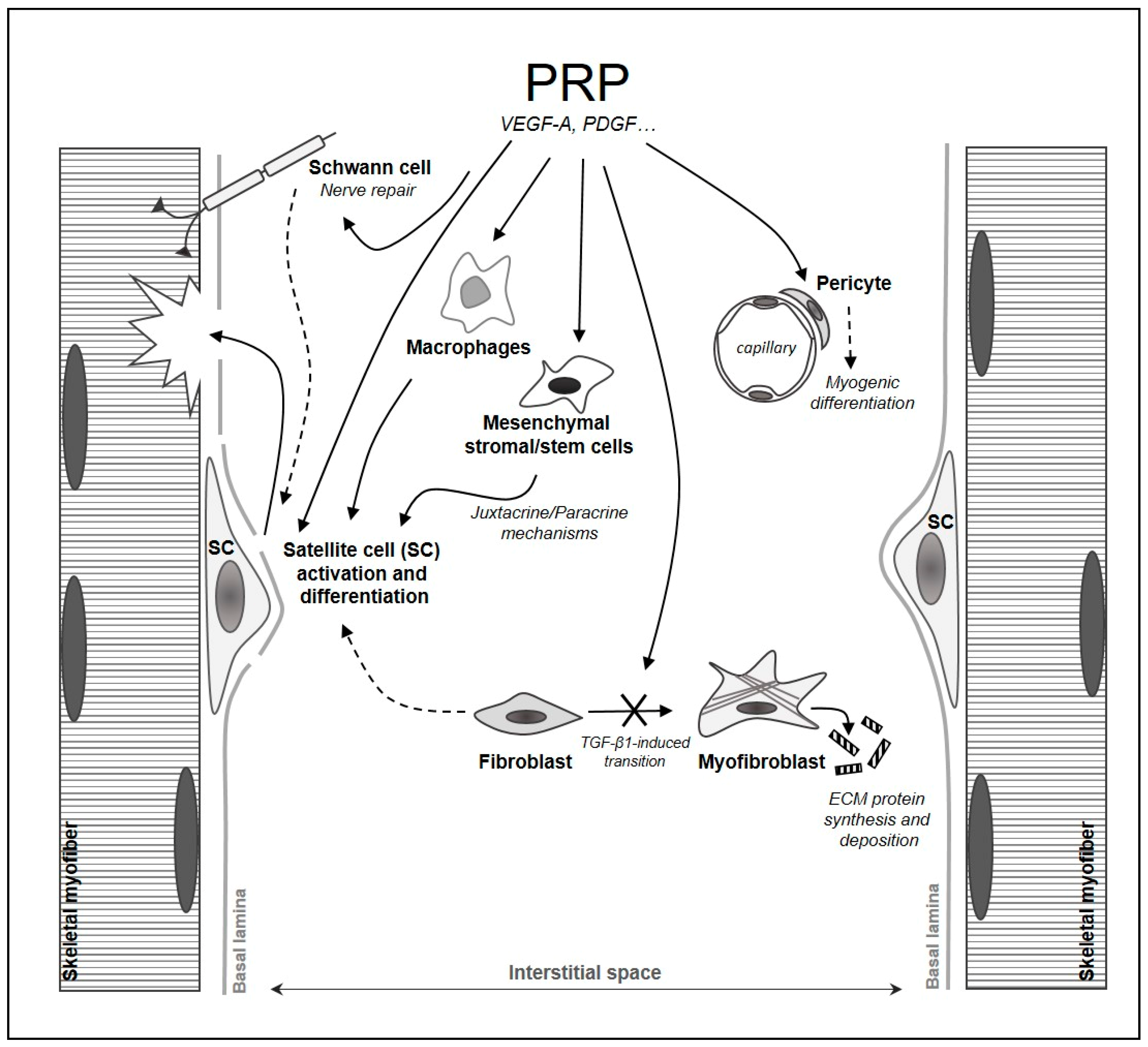
3.2. Contribution of PRP to Skeletal Muscle Repair/Regeneration
3.3. Impact of PRP on Satellite Cells and on Myogenic and Non-Myogenic Interstitial Cells
3.4. Impact of PPP on Skeletal Myogenesis
4. Conclusions and Further Directions
Author Contributions
Funding
Conflicts of Interest
References
- Giudice, J.; Taylor, J.M. Muscle as a paracrine and endocrine organ. Curr. Opin. Pharm. 2017, 34, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Saul, D.; Böker, K.O.; Ernst, J.; Lehman, W.; Schilling, A.F. Current Methods for Skeletal Muscle Tissue Repair and Regeneration. Biomed. Res. Int. 2018, 16, 1984879. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.; Sage, F.; Stok, M.; Brusse, E.; Pijnappel, W.W.M.; Reuser, A.J.; vd Ploeg, A.T. Ex-vivo Expansion of Muscle-Regenerative Cells for the Treatment of Muscle Disorders. J. Stem Cell Res. 2012, S11, 003. [Google Scholar] [CrossRef]
- Sassoli, C.; Pini, A.; Chellini, F.; Mazzanti, B.; Nistri, S.; Nosi, D.; Saccardi, R.; Quercioli, F.; Zecchi-Orlandini, S.; Formigli, L. Bone marrow mesenchymal stromal cells stimulate skeletal myoblast proliferation through the paracrine release of VEGF. PLoS ONE 2012, 7, e37512. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Zecchi-Orlandini, S.; Formigli, L. Trophic actions of bone marrow-derived mesenchymal stromal cells for muscle repair/regeneration. Cells 2012, 1, 832–850. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Nosi, D.; Tani, A.; Chellini, F.; Mazzanti, B.; Quercioli, F.; Zecchi-Orlandini, S.; Formigli, L. Defining the role of mesenchymal stromal cells on the regulation of matrix metalloproteinases in skeletal muscle cells. Exp. Cell Res. 2014, 323, 297–313. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.E. Concise review: Mesoangioblast and mesenchymal stem cell therapy for muscular dystrophy: Progress, challenges, and future directions. Stem Cells Transl. Med. 2015, 4, 91–98. [Google Scholar] [CrossRef] [PubMed]
- De Albornoz, P.M.; Aicale, R.; Forriol, F.; Maffulli, N. Cell Therapies in Tendon, Ligament, and Musculoskeletal System Repair. Sports Med. Arthrosc. Rev. 2018, 26, 48–58. [Google Scholar] [CrossRef]
- Andia, I.; Maffulli, N. New biotechnologies for musculoskeletal injuries. Surgeon 2018. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.; Talovic, M.; Patel, K.; Patel, A.; Marcinczyk, M.; Garg, K. Biomaterial and stem cell-based strategies for skeletal muscle regeneration. J. Orthop. Res. 2019. [Google Scholar] [CrossRef]
- Han, W.M.; Jang, Y.C.; García, A.J. Engineered matrices for skeletal muscle satellite cell engraftment and function. Matrix Biol. 2017, 60–61, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Kwee, B.J.; Mooney, D.J. Biomaterials for skeletal muscle tissue engineering. Curr. Opin. Biotechnol. 2017, 47, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.N.; Fernandes, K.P.; Deana, A.M.; Bussadori, S.K.; Mesquita-Ferrari, R.A. Effects of low-level laser therapy on skeletal muscle repair: A systematic review. Am. J. Phys. Med. Rehabil. 2014, 93, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Chellini, F.; Squecco, R.; Tani, A.; Idrizaj, E.; Nosi, D.; Giannelli, M.; Zecchi-Orlandini, S. Low intensity 635 nm diode laser irradiation inhibits fibroblast-myofibroblast transition reducing TRPC1 channel expression/activity: New perspectives for tissue fibrosis treatment. Lasers Surg. Med. 2016, 48, 318–332. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, H.A.; Antonio, E.L.; Silva, F.A.; de Carvalho, P.T.C.; Feliciano, R.; Yoshizaki, A.; Vieira, S.S.; de Melo, B.L.; Leal-Junior, E.C.P.; Labat, R.; et al. Protective effects of photobiomodulation against resistance exercise-induced muscle damage and inflammation in rats. J. Sports Sci. 2018, 36, 2349–2357. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed]
- Scicchitano, B.M.; Sica, G.; Musarò, A. Stem Cells and Tissue Niche: Two Faces of the Same Coin of Muscle Regeneration. Eur. J. Transl. Myol. 2016, 26, 6125. [Google Scholar] [CrossRef] [PubMed]
- Le Grand, F.; Rudnicki, M.A. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Mounier, R.; Chrétien, F.; Chazaud, B. Blood vessels and the satellite cell niche. Curr. Top. Dev. Biol. 2011, 96, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Dinulovic, I.; Furrer, R.; Handschin, C. Plasticity of the Muscle Stem Cell Microenvironment. Adv. Exp. Med. Biol. 2017, 1041, 141–169. [Google Scholar] [CrossRef]
- Dumont, N.A.; Wang, Y.X.; Rudnicki, M.A. Intrinsic and extrinsic mechanisms regulating satellite cell function. Development 2015, 142, 1572–1581. [Google Scholar] [CrossRef] [PubMed]
- Von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA 2013, 110, 16474–16479. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.T.; Sacco, A. Satellite Cell Heterogeneity in Skeletal Muscle Homeostasis. Trends Cell Biol. 2016, 26, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.; Kuroda, K.; Le Grand, F.; Rudnicki, M.A. Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell 2007, 129, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Bentzinger, C.F.; Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Cellular dynamics in the muscle satellite cell niche. EMBO Rep. 2013, 14, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Urciuolo, A.; Quarta, M.; Morbidoni, V.; Gattazzo, F.; Molon, S.; Grumati, P.; Montemurro, F.; Tedesco, F.S.; Blaauw, B.; Cossu, G.; et al. Collagen VI regulates satellite cell self-renewal and muscle regeneration. Nat. Commun. 2013, 4, 1964. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.; Engler, A.J.; Meyer, G.A. Extracellular matrix regulation in the muscle satellite cell niche. Connect. Tissue Res. 2015, 56, 1–8. [Google Scholar] [CrossRef]
- Malecova, B.; Puri, P.L. “Mix of Mics”-Phenotypic and Biological Heterogeneity of “Multipotent” Muscle Interstitial Cells (MICs). J. Stem Cell Res. 2012, pii:004. [Google Scholar] [CrossRef]
- Ceafalan, L.C.; Popescu, B.O.; Hinescu, M.E. Cellular players in skeletal muscle regeneration. Biomed. Res. Int. 2014, 2014, 957014. [Google Scholar] [CrossRef]
- Čamernik, K.; Barlič, A.; Drobnič, M.; Marc, J.; Jeras, M.; Zupan, J. Mesenchymal Stem Cells in the Musculoskeletal System: From Animal Models to Human Tissue Regeneration? Stem Cell Rev. 2018, 14, 346–369. [Google Scholar] [CrossRef]
- Tonlorenzi, R.; Rossi, G.; Messina, G. Isolation and Characterization of Vessel-Associated Stem/Progenitor Cells from Skeletal Muscle. Methods Mol. Biol. 2017, 1556, 149–177. [Google Scholar] [CrossRef]
- Vezzani, B.; Pierantozzi, E.; Sorrentino, V. Not All Pericytes Are Born Equal: Pericytes from Human Adult Tissues Present Different Differentiation Properties. Stem Cells Dev. 2016, 25, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- Judson, R.N.; Low, M.; Eisner, C.; Rossi, F.M. Isolation, Culture, and Differentiation of Fibro/Adipogenic Progenitors (FAPs) from Skeletal Muscle. Methods Mol. Biol. 2017, 1668, 93–103. [Google Scholar] [CrossRef]
- Marini, M.; Rosa, I.; Ibba-Manneschi, L.; Manetti, M. Telocytes in skeletal, cardiac and smooth muscle interstitium: Morphological and functional aspects. Histol. Histopathol. 2018, 33, 1151–1165. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.J.; Zhou, S.; Tanaka, K.K.; Pisconti, A.; Farina, N.H.; Sorrentino, B.P.; Olwin, B.B. Abcg2 labels multiple cell types in skeletal muscle and participates in muscle regeneration. J. Cell Biol. 2011, 195, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Akatsuka, A.; Okada, Y.; Matsuzaki, Y.; Okano, H.; Kimura, M. Growth and differentiation potential of main- and side-population cells derived from murine skeletal muscle. Exp. Cell Res. 2003, 291, 83–90. [Google Scholar] [CrossRef]
- Mitchell, K.J.; Pannérec, A.; Cadot, B.; Parlakian, A.; Besson, V.; Gomes, E.R.; Marazzi, G.; Sassoon, D.A. Identification and characterization of a non-satellite cell muscle resident progenitor during postnatal development. Nat. Cell. Biol. 2010, 12, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Farup, J.; Madaro, L.; Puri, P.L.; Mikkelsen, U.R. Interactions between muscle stem cells, mesenchymal-derived cells and immune cells in muscle homeostasis, regeneration and disease. Cell Death Dis. 2015, 6, e1830. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G.; Villalta, S.A. Regulatory interactions between muscle and the immune system during muscle regeneration. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1173–R1187. [Google Scholar] [CrossRef] [PubMed]
- Bencze, M.; Negroni, E.; Vallese, D.; Yacoub-Youssef, H.; Chaouch, S.; Wolff, A.; Aamiri, A.; Di Santo, J.P.; Chazaud, B.; Butler-Browne, G.; et al. Proinflammatory macrophages enhance the regenerative capacity of human myoblasts by modifying their kinetics of proliferation and differentiation. Mol. Ther. 2012, 20, 2168–2179. [Google Scholar] [CrossRef] [PubMed]
- Ceafalan, L.C.; Fertig, T.E.; Popescu, A.C.; Popescu, B.O.; Hinescu, M.E.; Gherghiceanu, M. Skeletal muscle regeneration involves macrophage-myoblast bonding. Cell Adhes. Migr. 2018, 12, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, W.; Ransohoff, R.M.; Zhou, L. Infiltrating macrophages are broadly activated at the early stage to support acute skeletal muscleinjury repair. J. Neuroimmunol. 2018, 317, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Sonnet, C.; Lafuste, P.; Arnold, L.; Brigitte, M.; Poron, F.; Authier, F.J.; Chrétien, F.; Gherardi, R.K.; Chazaud, B. Human macrophages rescue myoblasts and myotubes from apoptosis through a set of adhesion molecular systems. J. Cell Sci. 2006, 119, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.L.; Mann, C.J.; Vidal, B.; Ardite, E.; Perdiguero, E.; Muñoz-Cánoves, P. Cellular and molecular mechanisms regulating fibrosis in skeletal muscle repair and disease. Curr. Top. Dev. Biol. 2011, 96, 167–201. [Google Scholar] [CrossRef] [PubMed]
- Joe, A.W.; Yi, L.; Natarajan, A.; Le Grand, F.; So, L.; Wang, J.; Rudnicki, M.A.; Rossi, F.M. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nat. Cell Biol. 2010, 12, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.M.; Lawson, J.A.; Mathew, S.J.; Hutcheson, D.A.; Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 2011, 138, 3625–3637. [Google Scholar] [CrossRef] [PubMed]
- Heredia, J.E.; Mukundan, L.; Chen, F.M.; Mueller, A.A.; Deo, R.C.; Locksley, R.M.; Rando, T.A.; Chawla, A. Type 2 innate signals stimulate fibro/adipogenic progenitors to facilitate muscle regeneration. Cell 2013, 153, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Mackey, A.L.; Magnan, M.; Chazaud, B.; Kjaer, M. Human skeletal muscle fibroblasts stimulate in vitro myogenesis and in vivo muscle regeneration. J. Physiol. 2017, 595, 5115–5127. [Google Scholar] [CrossRef] [PubMed]
- Malecova, B.; Gatto, S.; Etxaniz, U.; Passafaro, M.; Cortez, A.; Nicoletti, C.; Giordani, L.; Torcinaro, A.; De Bardi, M.; Bicciato, S.; et al. Dynamics of cellular states of fibro-adipogenic progenitors during myogenesis and muscular dystrophy. Nat. Commun. 2018, 9, 3670. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Miyaki, S.; Ishitobi, H.; Matsuyama, S.; Nakasa, T.; Kamei, N.; Akimoto, T.; Higashi, Y.; Ochi, M. Mesenchymal-stem-cell-derived exosomes accelerate skeletal muscle regeneration. FEBS Lett. 2015, 589, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Christov, C.; Chrétien, F.; Abou-Khalil, R.; Bassez, G.; Vallet, G.; Authier, F.J.; Bassaglia, Y.; Shinin, V.; Tajbakhsh, S.; Chazaud, B.; et al. Muscle satellite cells and endothelial cells: Close neighbors and privileged partners. Mol. Biol. Cell 2007, 18, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Asakura, Y.; Murakonda, B.S.R.; Pengo, T.; Latroche, C.; Chazaud, B.; McLoon, L.K.; Asakura, A. Muscle Satellite Cell Cross-Talk with a Vascular Niche Maintains Quiescence via VEGF and Notch Signaling. Cell Stem Cell 2018, 23, 530–543.e9. [Google Scholar] [CrossRef] [PubMed]
- Abou-Khalil, R.; Mounier, R.; Chazaud, B. Regulation of myogenic stem cell behavior by vessel cells: The “menage a trois” of satellite cells, periendothelial cells and endothelial cells. Cell Cycle 2010, 9, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Kostallari, E.; Baba-Amer, Y.; Alonso-Martin, S.; Ngoh, P.; Relaix, F.; Lafuste, P.; Gherardi, R.K. Pericytes in the myovascular niche promote post-natal myofiber growth and satellite cell quiescence. Development 2015, 142, 1242–1253. [Google Scholar] [CrossRef] [PubMed]
- Menetrey, J.; Kasemkijwattana, C.; Day, C.S.; Bosch, P.; Vogt, M.; Fu, F.H.; Moreland, M.S.; Huard, J. Growth factors improve muscle healing in vivo. J. Bone Jt. Surg. Br. 2000, 82, 131–137. [Google Scholar] [CrossRef]
- De Perini, A.; Dimauro, I.; Duranti, G.; Fantini, C.; Mercatelli, N.; Ceci, R.; Di Luigi, L.; Sabatini, S.; Caporossi, D. The p75NTR-mediated effect of nerve growth factor in L6C5 myogenic cells. BMC Res. Notes 2017, 10, 686. [Google Scholar] [CrossRef] [PubMed]
- Judson, R.N.; Zhang, R.H.; Rossi, F.M. Tissue-resident mesenchymal stem/progenitor cells in skeletal muscle: Collaborators or saboteurs? FEBS J. 2013, 280, 4100–4108. [Google Scholar] [CrossRef] [PubMed]
- Asakura, A.; Seale, P.; Girgis-Gabardo, A.; Rudnicki, M.A. Myogenic specification of side population cells in skeletal muscle. J. Cell Biol. 2002, 159, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Uezumi, A.; Ojima, K.; Fukada, S.; Ikemoto, M.; Masuda, S.; Miyagoe-Suzuki, Y.; Takeda, S. Functional heterogeneity of side population cells in skeletal muscle. Biochem. Biophys. Res. Commun. 2006, 341, 864–873. [Google Scholar] [CrossRef]
- Tanaka, K.K.; Hall, J.K.; Troy, A.A.; Cornelison, D.D.; Majka, S.M.; Olwin, B.B. Syndecan-4-expressing muscle progenitor cells in the SP engraft as satellite cells during muscle regeneration. Cell Stem Cell 2009, 4, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Dellavalle, A.; Maroli, G.; Covarello, D.; Azzoni, E.; Innocenzi, A.; Perani, L.; Antonini, S.; Sambasivan, R.; Brunelli, S.; Tajbakhsh, S.; et al. Pericytes resident in postnatal skeletal muscle differentiate into muscle fibres and generate satellite cells. Nat. Commun. 2011, 2, 499. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Manera, J.; Gallardo, E.; de Luna, N.; Navas, M.; Soria, L.; Garibaldi, M.; Rojas-García, R.; Tonlorenzi, R.; Cossu, G.; Illa, I. The increase of pericyte population in human neuromuscular disorders supports their role in muscle regeneration in vivo. J. Pathol. 2012, 228, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Birbrair, A.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Enikolopov, G.N.; Mintz, A.; Delbono, O. Skeletal muscle pericyte subtypes differ in their differentiation potential. Stem Cell Res. 2013, 10, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Benchaouir, R.; Meregalli, M.; Farini, A.; D’Antona, G.; Belicchi, M.; Goyenvalle, A.; Battistelli, M.; Bresolin, N.; Bottinelli, R.; Garcia, L.; et al. Restoration of human dystrophin following transplantation of exon-skipping-engineered DMD patient stem cells into dystrophic mice. Cell Stem Cell 2007, 1, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Negroni, E.; Riederer, I.; Chaouch, S.; Belicchi, M.; Razini, P.; Di Santo, J.; Torrente, Y.; Butler-Browne, G.S.; Mouly, V. In vivo myogenic potential of human CD133+ muscle-derived stem cells: A quantitative study. Mol. Ther. 2009, 17, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Chun, S.; Asfahani, R.; Lochmüller, H.; Muntoni, F.; Morgan, J. Human skeletal muscle-derived CD133(+) cells form functional satellite cells after intramuscular transplantation in immunodeficient host mice. Mol. Ther. 2014, 22, 1008–1017. [Google Scholar] [CrossRef]
- Dulauroy, S.; Di Carlo, S.E.; Langa, F.; Eberl, G.; Peduto, L. Lineage tracing and genetic ablation of ADAM12(+) perivascular cells identify a major source of profibrotic cells during acute tissue injury. Nat. Med. 2012, 18, 1262–1270. [Google Scholar] [CrossRef]
- Uezumi, A.; Ikemoto-Uezumi, M.; Tsuchida, K. Roles of non myogenic mesenchymal progenitors in pathogenesis and regeneration of skeletal muscle. Front. Physiol. 2014, 5, 68. [Google Scholar] [CrossRef]
- Lemos, D.R.; Babaeijandaghi, F.; Low, M.; Chang, C.K.; Lee, S.T.; Fiore, D.; Zhang, R.H.; Natarajan, A.; Nedospasov, S.A.; Rossi, F.M. Nilotinib reduces muscle fibrosis in chronic muscle injury by promoting TNF-mediated apoptosis of fibro/adipogenic progenitors. Nat. Med. 2015, 21, 786–794. [Google Scholar] [CrossRef]
- Rudolf, R.; Deschenes, M.R.; Sandri, M. Neuromuscular junction degeneration in muscle wasting. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 177–181. [Google Scholar] [CrossRef] [PubMed]
- DeLong, J.M.; Russell, R.P.; Mazzocca, A.D. Platelet-rich plasma: The PAW classification system. Arthroscopy 2012, 28, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- De Pascale, M.R.; Sommese, L.; Casamassimi, A.; Napoli, C. Platelet derivatives in regenerative medicine: An update. Transfus. Med. Rev. 2015, 29, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Mautner, K.; Malanga, G.A.; Smith, J.; Shiple, B.; Ibrahim, V.; Sampson, S.; Bowen, J.E. A call for a standard classification system for future biologic research: The rationale for new PRP nomenclature. PM R 2015, 7, S53–S59. [Google Scholar] [CrossRef] [PubMed]
- Le, A.D.K.; Enweze, L.; DeBaun, M.R.; Dragoo, J.L. Platelet-Rich Plasma. Clin. Sports Med. 2019, 38, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, V.; Gentile, P.; Scioli, M.G.; Grimaldi, M.; Casciani, C.U.; Spagnoli, L.G.; Orlandi, A. Application of platelet-rich plasma in plastic surgery: Clinical and in vitro evaluation. Tissue Eng. Part C Methods 2009, 15, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.A. Evolution, current status and advances in application of platelet concentrate in periodontics and implantology. World J. Clin. Cases 2017, 5, 159–171. [Google Scholar] [CrossRef]
- Giannaccare, G.; Versura, P.; Buzzi, M.; Primavera, L.; Pellegrini, M.; Campos, E.C. Blood derived eye drops for the treatment of cornea and ocular surface diseases. Transfus. Apher. Sci. 2017, 56, 595–604. [Google Scholar] [CrossRef]
- Tandulwadkar, S.R.; Naralkar, M.V.; Surana, A.D.; Selvakarthick, M.; Kharat, A.H. Autologous intrauterine platelet-rich plasma instillation for suboptimal endometrium in frozen embryo transfer cycles: A pilot study. J. Hum. Reprod. Sci. 2017, 10, 208–212. [Google Scholar] [CrossRef]
- Santos, S.C.N.D.S.; Sigurjonsson, O.E.; Custodio, C.A.; Mano, J.F.C.D.L. Blood plasma derivatives for tissue engineering and regenerative medicine therapies. Tissue Eng. Part B Rev. 2018, 24, 454–462. [Google Scholar] [CrossRef]
- Chicharro-Alcántara, D.; Rubio-Zaragoza, M.; Damiá-Giménez, E.; Carrillo-Poveda, J.M.; Cuervo-Serrato, B.; Peláez-Gorrea, P.; Sopena-Juncosa, J.J. Platelet rich plasma: New insights for cutaneous wound healing management. J. Funct. Biomater. 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, R.T.; Borg-Stein, J.; McInnis, K. Applications of platelet-rich plasma in musculoskeletal and sports medicine: An evidence-based approach. PM R 2011, 3, 226–250. [Google Scholar] [CrossRef] [PubMed]
- Andia, I.; Abate, M. Platelet-rich plasma in the treatment of skeletal muscle injuries. Expert. Opin. Biol. 2015, 15, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Andia, I.; Abate, M. Platelet-rich plasma: Combinational treatment modalities for musculoskeletal conditions. Front. Med. 2018, 12, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Andia, I.; Martin, J.I.; Maffulli, N. Advances with platelet rich plasma therapies for tendon regeneration. Expert Opin. Biol. 2018, 18, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Amable, P.R.; Carias, R.B.; Teixeira, M.V.; da Cruz Pacheco, I.; Corrêa do Amaral, R.J.; Granjeiro, J.M.; Borojevic, R. Platelet-rich plasma preparation for regenerative medicine: Optimization and quantification of cytokines and growth factors. Stem Cell Res. 2013, 4, 67. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.E.; Smith, P.C.; Palma Alvarado, V.A. The influence of platelet-derived products on angiogenesis and tissue repair: A concise update. Front. Physiol. 2015, 6, 290. [Google Scholar] [CrossRef] [PubMed]
- Pochini, A.C.; Antonioli, E.; Bucci, D.Z.; Sardinha, L.R.; Andreoli, C.V.; Ferretti, M.; Ejnisman, B.; Goldberg, A.C.; Cohen, M. Analysis of cytokine profile and growth factors in platelet-rich plasma obtained by open systems and commercial columns. Einstein 2016, 14, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; An, N.; Ouyang, X. Quantification of growth factors in different platelet concentrates. Platelets 2017, 28, 774–778. [Google Scholar] [CrossRef]
- San Sebastian, K.M.; Lobato, I.; Hernández, I.; Burgos-Alonso, N.; Gomez-Fernandez, M.C.; López, J.L.; Rodríguez, B.; March, A.G.; Grandes, G.; Andia, I. Efficacy and safety of autologous platelet rich plasma for the treatment of vascular ulcers in primary care: Phase III study. BMC Fam. Pract. 2014, 15, 211. [Google Scholar] [CrossRef]
- Suthar, M.; Gupta, S.; Bukhari, S.; Ponemone, V. Treatment of chronic non-healing ulcers using autologous platelet rich plasma: A case series. J. Biomed. Sci. 2017, 24, 16. [Google Scholar] [CrossRef] [PubMed]
- Bottegoni, C.; Farinelli, L.; Aquili, A.; Chiurazzi, E.; Gigante, A. Homologous platelet-rich plasma for the treatment of knee involvement in primary Sjögren’s syndrome. J. Biol. Regul. Homeost. Agents 2016, 30, 63–67. [Google Scholar] [PubMed]
- Anitua, E.; Prado, R.; Orive, G. Allogeneic Platelet-Rich Plasma: At the Dawn of an Off-the-Shelf Therapy? Trends Biotechnol. 2017, 35, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Bubnov, R.; Yevseenko, V.; Semeniv, I. Ultrasound guided injections of platelets rich plasma for muscle injury in professional athletes. Comparative study. Med. Ultrason. 2013, 15, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.J.; Patel, R.M.; Terry, M.A. Platelet-rich plasma as an effective treatment for proximal hamstring injuries. Orthopedics 2013, 36, e64–e70. [Google Scholar] [CrossRef] [PubMed]
- A Hamid, M.S.; Mohamed Ali, M.R.; Yusof, A.; George, J.; Lee, L.P. Platelet-rich plasma injections for the treatment of hamstring injuries: A randomized controlled trial. Am. J. Sports Med. 2014, 42, 2410–2418. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.A.; Molina Rómoli, A.R.; Bertona Altieri, B.A.; Burgos Flor, J.A.; Scordo, W.E.; Elizondo, C.M. Does platelet-rich plasma decrease time to return to sports in acute muscle tear? A randomized controlled trial. Knee Surg. Sports Traumatol. Arthrosc. 2017, 25, 3319–3325. [Google Scholar] [CrossRef]
- Borrione, P.; Fossati, C.; Pereira, M.T.; Giannini, S.; Davico, M.; Minganti, C.; Pigozzi, F. The use of platelet-rich plasma (PRP) in the treatment of gastrocnemius strains: A retrospective observational study. Platelets 2018, 29, 596–601. [Google Scholar] [CrossRef]
- Bernuzzi, G.; Petraglia, F.; Pedrini, M.F.; De Filippo, M.; Pogliacomi, F.; Verdano, M.A.; Costantino, C. Use of platelet-rich plasma in the care of sports injuries: Our experience with ultrasound-guided injection. Blood Transfus. 2014, 12, s229–s234. [Google Scholar] [CrossRef]
- Zanon, G.; Combi, F.; Combi, A.; Perticarini, L.; Sammarchi, L.; Benazzo, F. Platelet-rich plasma in the treatment of acute hamstring injuries in professional football players. Joints 2016, 4, 17–23. [Google Scholar] [CrossRef]
- Punduk, Z.; Oral, O.; Ozkayin, N.; Rahman, K.; Varol, R. Single dose of intra-muscular platelet rich plasma reverses the increase in plasma iron levels in exercise-induced muscle damage: A pilot study. J. Sport Health Sci. 2016, 5, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Sheth, U.; Dwyer, T.; Smith, I.; Wasserstein, D.; Theodoropoulos, J.; Takhar, S.; Chahal, J. Does Platelet-Rich Plasma Lead to Earlier Return to Sport When Compared With Conservative Treatment in Acute Muscle Injuries? A Systematic Review and Meta-analysis. Arthroscopy 2018, 34, 281–288.e1. [Google Scholar] [CrossRef] [PubMed]
- Scully, D.; Naseem, K.M.; Matsakas, A. Platelet biology in regenerative medicine of skeletal muscle. Acta Physiol. (Oxf.) 2018, 223, e13071. [Google Scholar] [CrossRef] [PubMed]
- Grassi, A.; Napoli, F.; Romandini, I.; Samuelsson, K.; Zaffagnini, S.; Candrian, C.; Filardo, G. Is Platelet-Rich Plasma (PRP) Effective in the Treatment of Acute Muscle Injuries? A Systematic Review and Meta-Analysis. Sports Med. 2018, 48, 971–989. [Google Scholar] [CrossRef] [PubMed]
- Borrione, P.; Grasso, L.; Chierto, E.; Geuna, S.; Racca, S.; Abbadessa, G.; Ronchi, G.; Faiola, F.; Di Gianfrancesco, A.; Pigozzi, F. Experimental model for the study of the effects of platelet-rich plasma on the early phases of muscle healing. Blood Transfus. 2014, 12, s221–s228. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hicks, J.J.; Wang, L.; Oyster, N.; Philippon, M.J.; Hurwitz, S.; Hogan, M.V.; Huard, J. Customized platelet-rich plasma with transforming growth factor β1 neutralization antibody to reduce fibrosis in skeletal muscle. Biomaterials 2016, 87, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.A.; Camargo, R.C.T.; Koike, T.E.; Ozaki, G.A.T.; Castoldi, R.C.; Camargo Filho, J.C.S. Histological analysis of the association of low level laser therapy and platelet-rich plasma in regeneration of muscle injury in rats. Braz. J. Phys. 2017, 21, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Yu, T.Y.; Chang, G.J.; Lin, L.P.; Lin, M.S.; Pang, J.S. Platelet-Rich Plasma Releasate Promotes Regeneration and Decreases Inflammation and Apoptosis of Injured Skeletal Muscle. Am. J. Sports Med. 2018, 46, 1980–1986. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.W.; Hinton, R.Y.; Curl, L.A.; Muriel, J.M.; Lovering, R.M. Use of autologous platelet-rich plasma to treat muscle strain injuries. Am. J. Sports Med. 2009, 37, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Gigante, A.; Del Torto, M.; Manzotti, S.; Cianforlini, M.; Busilacchi, A.; Davidson, P.A.; Greco, F.; Mattioli-Belmonte, M. Platelet rich fibrin matrix effects on skeletal muscle lesions: An experimental study. J. Biol. Regul. Homeost. Agents 2012, 26, 475–484. [Google Scholar] [PubMed]
- Dimauro, I.; Grasso, L.; Fittipaldi, S.; Fantini, C.; Mercatelli, N.; Racca, S.; Geuna, S.; Di Gianfrancesco, A.; Caporossi, D.; Pigozzi, F.; et al. Platelet-rich plasma and skeletal muscle healing: A molecular analysis of the early phases of the regeneration process in an experimental animal model. PLoS ONE 2014, 9, e102993. [Google Scholar] [CrossRef] [PubMed]
- Cianforlini, M.; Mattioli-Belmonte, M.; Manzotti, S.; Chiurazzi, E.; Piani, M.; Orlando, F.; Provinciali, M.; Gigante, A. Effect of platelet rich plasma concentration on skeletal muscle regeneration: An experimental study. J. Biol. Regul. Homeost. Agents 2015, 29, 47–55. [Google Scholar] [PubMed]
- Contreras-Muñoz, P.; Torrella, J.R.; Serres, X.; Rizo-Roca, D.; De la Varga, M.; Viscor, G.; Martínez-Ibáñez, V.; Peiró, J.L.; Järvinen, T.A.H.; Rodas, G.; et al. Postinjury Exercise and Platelet-Rich Plasma Therapies Improve Skeletal Muscle Healing in Rats But Are Not Synergistic When Combined. Am. J. Sports Med. 2017, 45, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.P.; Hartmann, D.D.; de Moraes, J.P.; Soares, F.A.; Puntel, G.O. Platelet-rich plasma reduces the oxidative damage determined by a skeletal muscle contusion in rats. Platelets 2016, 27, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Cunha, R.C.; Francisco, J.C.; Cardoso, M.A.; Matos, L.F.; Lino, D.; Simeoni, R.B.; Pereira, G.; Irioda, A.C.; Simeoni, P.R.; Guarita-Souza, L.C.; et al. Effect of platelet-rich plasma therapy associated with exercise training in musculoskeletal healing in rats. Transpl. Proc. 2014, 46, 1879–1881. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Pelacho, B.; Prado, R.; Aguirre, J.J.; Sánchez, M.; Padilla, S.; Aranguren, X.L.; Abizanda, G.; Collantes, M.; Hernandez, M.; et al. Infiltration of plasma rich in growth factors enhances in vivo angiogenesis and improves reperfusion and tissue remodeling after severe hind limb ischemia. J. Control. Release 2015, 202, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Ota, S.; Kobayashi, M.; Kobayashi, T.; Mifune, Y.; Takayama, K.; Witt, M.; Vadalà, G.; Oyster, N.; Otsuka, T.; et al. Use of an antifibrotic agent improves the effect of platelet-rich plasma on muscle healing after injury. J. Bone Jt. Surg. Am. 2013, 95, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Denapoli, P.M.; Stilhano, R.S.; Ingham, S.J.; Han, S.W.; Abdalla, R.J. Platelet-Rich Plasma in a Murine Model: Leukocytes, Growth Factors, Flt-1, and Muscle Healing. Am. J. Sports Med. 2016, 44, 1962–1971. [Google Scholar] [CrossRef]
- Pinheiro, C.L.; Peixinho, C.C.; Esposito, C.C.; Manso, J.E.; Machado, J.C. Ultrasound biomicroscopy and claudication test for in vivo follow-up of muscle repair enhancement based on platelet-rich plasma therapy in a rat model of gastrocnemius laceration. Acta Cir. Bras. 2016, 31, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Takase, F.; Inui, A.; Mifune, Y.; Sakata, R.; Muto, T.; Harada, Y.; Ueda, Y.; Kokubu, T.; Kurosaka, M. Effect of platelet-rich plasma on degeneration change of rotator cuff muscles: In vitro and in vivo evaluations. J. Orthop. Res. 2017, 35, 1806–1815. [Google Scholar] [CrossRef] [PubMed]
- Miroshnychenko, O.; Chang, W.T.; Dragoo, J.L. The Use of Platelet-Rich and Platelet-Poor Plasma to Enhance Differentiation of Skeletal Myoblasts: Implications for the Use of Autologous Blood Products for Muscle Regeneration. Am. J. Sports Med. 2017, 45, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Usas, A.; Poddar, M.; Chen, C.W.; Thompson, S.; Ahani, B.; Cummins, J.; Lavasani, M.; Huard, J. Platelet-rich plasma promotes the proliferation of human muscle derived progenitor cells and maintains their stemness. PLoS ONE 2013, 8, e64923. [Google Scholar] [CrossRef] [PubMed]
- Im, W.; Ban, J.J.; Lim, J.; Lee, M.; Chung, J.Y.; Bhattacharya, R.; Kim, S.H. Adipose-derived stem cells extract has a proliferative effect on myogenic progenitors. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 740–766. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Yu, T.Y.; Lin, L.P.; Lin, M.S.; Wu, Y.C.; Liao, C.H.; Pang, J.S. Platelet rich plasma releasate promotes proliferation of skeletal muscle cells in association with upregulation of PCNA, cyclins and cyclin dependent kinases. Platelets 2017, 28, 491–497. [Google Scholar] [CrossRef] [PubMed]
- McClure, M.J.; Garg, K.; Simpson, D.G.; Ryan, J.J.; Sell, S.A.; Bowlin, G.L.; Ericksen, J.J. The influence of platelet-rich plasma on myogenic differentiation. J. Tissue Eng. Regen. Med. 2016, 10, E239–E249. [Google Scholar] [CrossRef] [PubMed]
- McClure, M.J.; Clark, N.M.; Schwartz, Z.; Boyan, B.D. Platelet-rich plasma and alignment enhance myogenin via ERK mitogen activated protein kinase signaling. Biomed. Mater. 2018, 13, 055009. [Google Scholar] [CrossRef] [PubMed]
- Kelc, R.; Trapecar, M.; Gradisnik, L.; Rupnik, M.S.; Vogrin, M. Platelet-rich plasma, especially when combined with a TGF-β inhibitor promotes proliferation, viability and myogenic differentiation of myoblasts in vitro. PLoS ONE 2015, 10, e0117302. [Google Scholar] [CrossRef]
- Deasy, B.M.; Feduska, J.M.; Payne, T.R.; Li, Y.; Ambrosio, F.; Huard, J. Effect of VEGF on the regenerative capacity of muscle stem cells in dystrophic skeletal muscle. Mol. Ther. 2009, 17, 1788–1798. [Google Scholar] [CrossRef]
- Walker, N.; Kahamba, T.; Woudberg, N.; Goetsch, K.; Niesler, C. Dose-dependent modulation of myogenesis by HGF: Implications for c-Met expression and downstream signalling pathways. Growth Factors 2015, 33, 229–241. [Google Scholar] [CrossRef]
- Duan, C.; Ren, H.; Gao, S. Insulin-like growth factors (IGFs), IGF receptors, and IGF-binding proteins: Roles in skeletal muscle growth and differentiation. Gen. Comp. Endocrinol. 2010, 167, 344–351. [Google Scholar] [CrossRef]
- Scully, D.; Sfyri, P.; Verpoorten, S.; Papadopoulos, P.; Muñoz-Turrillas, M.C.; Mitchell, R.; Aburima, A.; Patel, K.; Gutiérrez, L.; Naseem, K.M.; et al. Platelet releasate promotes skeletal myogenesis by increasing muscle stem cell commitment to differentiation and accelerates muscle regeneration following acute injury. Acta Physiol. (Oxf.) 2018, 19, e13207. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Yu, T.Y.; Lin, L.P.; Lin, M.S.; Tsai, T.T.; Pang, J.S. Platelet rich plasma promotes skeletal muscle cell migration in association with up-regulation of FAK, paxillin, and F-Actin formation. J. Orthop. Res. 2017, 35, 2506–2512. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Vallone, L.; Tani, A.; Chellini, F.; Nosi, D.; Zecchi-Orlandini, S. Combined use of bone marrow-derived mesenchymal stromal cells (BM-MSCs) and platelet rich plasma (PRP) stimulates proliferation and differentiation of myoblasts in vitro: New therapeutic perspectives for skeletal muscle repair/regeneration. Cell Tissue Res. 2018, 372, 549–570. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Sankoda, Y.; Tatsumi, R.; Mizunoya, W.; Ikeuchi, Y.; Sunagawa, K.; Allen, R.E. Matrix metalloproteinase-2 mediates stretch-induced activation of skeletal muscle satellite cells in a nitric oxide-dependent manner. Int. J. Biochem. Cell Biol. 2008, 40, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Pallafacchina, G.; François, S.; Regnault, B.; Czarny, B.; Dive, V.; Cumano, A.; Montarras, D.; Buckingham, M. An adult tissue-specific stem cell in its niche: A gene profiling analysis of in vivo quiescent and activated muscle satellite cells. Stem Cell Res. 2010, 4, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Bellayr, I.; Holden, K.; Mu, X.; Pan, H.; Li, Y. Matrix metalloproteinase inhibition negatively affects muscle stem cell behavior. Int. J. Clin. Exp. Pathol. 2013, 6, 124–141. [Google Scholar] [PubMed]
- Chen, X.; Li, Y. Role of matrix metalloproteinases in skeletal muscle: Migration, differentiation, regeneration and fibrosis. Cell Adhes. Migr. 2009, 3, 337–341. [Google Scholar] [CrossRef]
- Miyazaki, D.; Nakamura, A.; Fukushima, K.; Yoshida, K.; Takeda, S.; Ikeda, S. Matrix metalloproteinase-2 ablation in dystrophin-deficient mdx muscles reduces angiogenesis resulting in impaired growth of regenerated muscle fibers. Hum. Mol. Genet. 2011, 20, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Cassano, M.; Dellavalle, A.; Tedesco, F.S.; Quattrocelli, M.; Crippa, S.; Ronzoni, F.; Salvade, A.; Berardi, E.; Torrente, Y.; Cossu, G.; et al. Alpha sarcoglycan is required for FGF-dependent myogenic progenitor cell proliferation in vitro and in vivo. Development 2011, 138, 4523–4533. [Google Scholar] [CrossRef]
- Sartori, R.; Gregorevic, P.; Sandri, M. TGFβ and BMP signaling in skeletal muscle: Potential significance for muscle-related disease. Trends Endocrinol. Metab. 2014, 25, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Escobar, G.; Escobar, A.; Ascui, G.; Tempio, F.I.; Ortiz, M.C.; Pérez, C.A.; López, M.N. Pure platelet-rich plasma and supernatant of calcium-activated P-PRP induce different phenotypes of human macrophages. Regen. Med. 2018, 13, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Papait, A.; Cancedda, R.; Mastrogiacomo, M.; Poggi, A. Allogeneic platelet-rich plasma affects monocyte differentiation to dendritic cells causing an anti-inflammatory microenvironment, putatively fostering wound healing. J. Tissue Eng. Regen. Med. 2018, 12, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Pakshir, P.; Hinz, B. The big five in fibrosis: Macrophages, myofibroblasts, matrix, mechanics, and miscommunication. Matrix Biol. 2018, 68–69, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Chellini, F.; Tani, A.; Vallone, L.; Nosi, D.; Pavan, P.; Bambi, F.; Zecchi Orlandini, S.; Sassoli, C. Platelet-Rich Plasma Prevents In Vitro Transforming Growth Factor-β1-Induced Fibroblast to Myofibroblast Transition: Involvement of Vascular Endothelial Growth Factor (VEGF)-A/VEGF Receptor-1-Mediated Signaling. Cells 2018, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Troya, M.; Orive, G. Plasma rich in growth factors promote gingival tissue regeneration by stimulating fibroblast proliferation and migration and by blocking transforming growth factor-β1-induced myodifferentiation. J. Periodontol. 2012, 83, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; de la Fuente, M.; Muruzabal, F.; Riestra, A.; Merayo-Lloves, J.; Orive, G. Plasma rich in growth factors (PRGF) eye drops stimulates scarless regeneration compared to autologous serum in the ocular surface stromal fibroblasts. Exp. Eye Res. 2015, 135, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Rivlin, M.; Graham, J.G.; Beredjiklian, P.K. Peripheral nerve injury, scarring, and recovery. Connect. Tissue Res. 2018, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Giannessi, E.; Coli, A.; Stornelli, M.R.; Miragliotta, V.; Pirone, A.; Lenzi, C.; Burchielli, S.; Vozzi, G.; De Maria, C.; Giorgetti, M. An autologously generated platelet-rich plasma suturable membrane may enhance peripheral nerve regeneration after neurorraphy in an acute injury model of sciatic nerve neurotmesis. J. Reconstr. Microsurg. 2014, 30, 617–626. [Google Scholar] [CrossRef]
- Zheng, C.; Zhu, Q.; Liu, X.; Huang, X.; He, C.; Jiang, L.; Quan, D.; Zhou, X.; Zhu, Z. Effect of platelet-rich plasma (PRP) concentration on proliferation, neurotrophic function and migration of Schwann cells in vitro. J. Tissue Eng. Regen. Med. 2016, 10, 428–436. [Google Scholar] [CrossRef]
- Sánchez, M.; Garate, A.; Delgado, D.; Padilla, S. Platelet-rich plasma, an adjuvant biological therapy to assist peripheral nerve repair. Neural Regen. Res. 2017, 12, 47–52. [Google Scholar] [CrossRef]
- Teymur, H.; Tiftikcioglu, Y.O.; Cavusoglu, T.; Tiftikcioglu, B.I.; Erbas, O.; Yigitturk, G.; Uyanikgil, Y. Effect of platelet-rich plasma on reconstruction with nerve autografts. Kaohsiung J. Med. Sci. 2017, 33, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, M.; Martínez, C.; Martínez, J.; Smith, P.C. Effects of platelet-rich and -poor plasma on the reparative response of gingival fibroblasts. Clin. Oral Implant. Res. 2012, 23, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Creeper, F.; Ivanovski, S. Effect of autologous and allogenic platelet-rich plasma on human gingival fibroblast function. Oral Dis. 2012, 18, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.E.; González, S.A.; Palma, V.; Smith, P.C. Platelet-Poor and Platelet-Rich Plasma Stimulate Bone Lineage Differentiation in Periodontal Ligament Stem Cells. J. Periodontol. 2016, 87, e18–e26. [Google Scholar] [CrossRef] [PubMed]
- de Mos, M.; van der Windt, A.E.; Jahr, H.; van Schie, H.T.; Weinans, H.; Verhaar, J.A.; van Osch, G.J. Can platelet-rich plasma enhance tendon repair? A cell culture study. Am. J. Sports Med. 2008, 36, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, M.; Vatanmakanian, M.; Arami, M.K.; Sadeghi Shirazi, F.; Esmaeili, N.; Hydarporian, S.; Jafari, S. A comparative study between platelet-rich plasma and platelet-poor plasma effects on angiogenesis. Med. Mol. Morphol. 2018, 51, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Renn, T.Y.; Kao, Y.H.; Wang, C.C.; Burnouf, T. Anti-inflammatory effects of platelet biomaterials in a macrophage cellular model. Vox Sang. 2015, 109, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Rettig, A.C.; Meyer, S.; Bhadra, A.K. Platelet-Rich Plasma in Addition to Rehabilitation for Acute Hamstring Injuries in NFL Players: Clinical Effects and Time to Return to Play. Orthop. J. Sports Med. 2013, 1, 2325967113494354. [Google Scholar] [CrossRef] [PubMed]
- Delos, D.; Leineweber, M.J.; Chaudhury, S.; Alzoobaee, S.; Gao, Y.; Rodeo, S.A. The effect of platelet-rich plasma on muscle contusion healing in a rat model. Am. J. Sports Med. 2014, 42, 2067–2074. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M.S.; Yusof, A.; Mohamed Ali, M.R. Platelet-rich plasma (PRP) for acute muscle injury: A systematic review. PLoS ONE 2014, 9, e90538. [Google Scholar] [CrossRef]
- Hamilton, B.; Tol, J.L.; Almusa, E.; Boukarroum, S.; Eirale, C.; Farooq, A.; Whiteley, R.; Chalabi, H. Platelet-rich plasma does not enhance return to play in hamstring injuries: A randomised controlled trial. Br. J. Sports Med. 2015, 49, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Reurink, G.; Goudswaard, G.J.; Moen, M.H.; Weir, A.; Verhaar, J.A.; Bierma-Zeinstra, S.M.; Maas, M.; Tol, J.L. Dutch HIT-study Investigators. Rationale, secondary outcome scores and 1-year follow-up of a randomised trial of platelet-rich plasma injections in acute hamstring muscle injury: The Dutch Hamstring Injection Therapy study. Br. J. Sports Med. 2015, 49, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Kelc, R.; Vogrin, M. Concerns about fibrosis development after scaffolded PRP therapy of muscle injuries: Commentary on an article by Sanchez et al.: Muscle repair: Platelet-rich plasma derivates as a bridge from spontaneity to intervention. Injury 2015, 46, 428. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Zapata, M.J.; Orozco, L.; Balius, R.; Soler, R.; Bosch, A.; Rodas, G.; Til, L.; Peirau, X.; Urrútia, G.; Gich, I.; et al. PRP-RICE group. Efficacy of autologous platelet-rich plasma for the treatment of muscle rupture with haematoma: A multicentre, randomised, double-blind, placebo-controlled clinical trial. Blood Transfus. 2016, 14, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Guillodo, Y.; Madouas, G.; Simon, T.; Le Dauphin, H.; Saraux, A. Platelet-rich plasma (PRP) treatment of sports-related severe acute hamstring injuries. Muscles Ligaments Tendons J. 2016, 5, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Navani, A.; Li, G.; Chrystal, J. Platelet rich plasma in musculoskeletal pathology: A necessary rescue or a lost cause? Pain Physician 2017, 20, E345–E356. [Google Scholar] [PubMed]
- Tonogai, I.; Hayashi, F.; Iwame, T.; Takasago, T.; Matsuura, T.; Sairyo, K. Platelet-rich plasma does not reduce skeletal muscle fibrosis after distraction osteogenesis. J. Exp. Orthop. 2018, 5, 26. [Google Scholar] [CrossRef]
- Manduca, M.L.; Straub, S.J. Effectiveness of PRP Injection in Reducing Recovery Time of Acute Hamstring Injury: A Critically Appraised Topic. J. Sport Rehabil. 2018, 27, 480–484. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chellini, F.; Tani, A.; Zecchi-Orlandini, S.; Sassoli, C. Influence of Platelet-Rich and Platelet-Poor Plasma on Endogenous Mechanisms of Skeletal Muscle Repair/Regeneration. Int. J. Mol. Sci. 2019, 20, 683. https://doi.org/10.3390/ijms20030683
Chellini F, Tani A, Zecchi-Orlandini S, Sassoli C. Influence of Platelet-Rich and Platelet-Poor Plasma on Endogenous Mechanisms of Skeletal Muscle Repair/Regeneration. International Journal of Molecular Sciences. 2019; 20(3):683. https://doi.org/10.3390/ijms20030683
Chicago/Turabian StyleChellini, Flaminia, Alessia Tani, Sandra Zecchi-Orlandini, and Chiara Sassoli. 2019. "Influence of Platelet-Rich and Platelet-Poor Plasma on Endogenous Mechanisms of Skeletal Muscle Repair/Regeneration" International Journal of Molecular Sciences 20, no. 3: 683. https://doi.org/10.3390/ijms20030683
APA StyleChellini, F., Tani, A., Zecchi-Orlandini, S., & Sassoli, C. (2019). Influence of Platelet-Rich and Platelet-Poor Plasma on Endogenous Mechanisms of Skeletal Muscle Repair/Regeneration. International Journal of Molecular Sciences, 20(3), 683. https://doi.org/10.3390/ijms20030683